
Ex Vivo Study on the Antioxidant Activity of a Winemaking By-Product Polyphenolic Extract (Taurisolo®) on Human Neutrophils

 , ,
, , 
 ,
,  ,
,  ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Taurisolo® Supplement Preparation
2.3. In Vitro Gastrointestinal Digestion
2.4. Antioxidant Activity of Taurisolo®
2.5. Cell Isolation and Cell Viability Test
2.6. Cell Treatment and Experimental Design
2.7. Hydrogen Peroxide Production
2.8. Oxidative Stress-Related Enzyme Activity
2.9. Malondialdehyde Assay
2.10. Gene Expression
2.11. Statistical Analysis
3. Results
3.1. Chemical Profile of Native and Digested-Taurisolo®
3.2. Characteristics of Study Participants
3.3. Antioxidant Activity of Taurisolo®
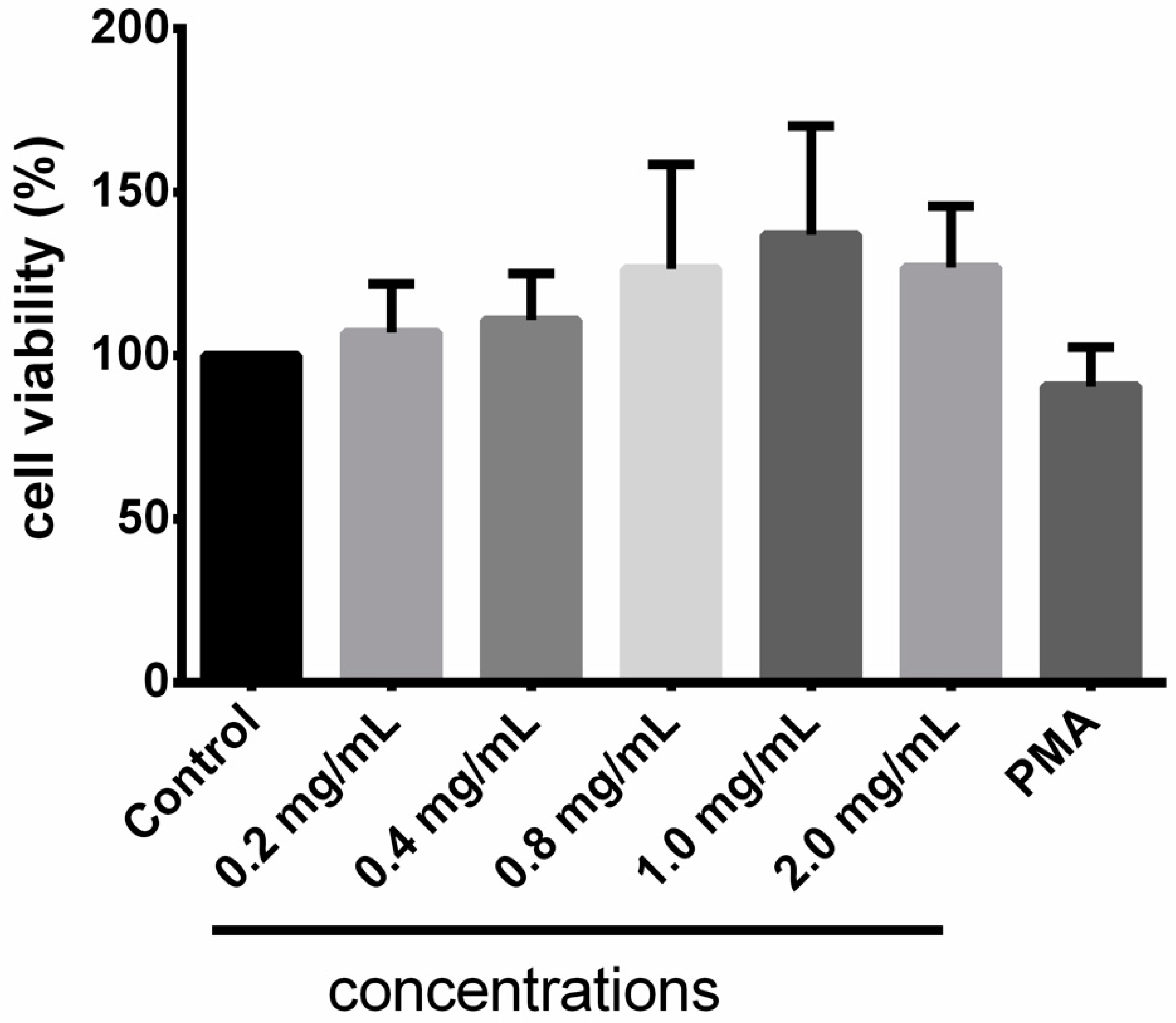
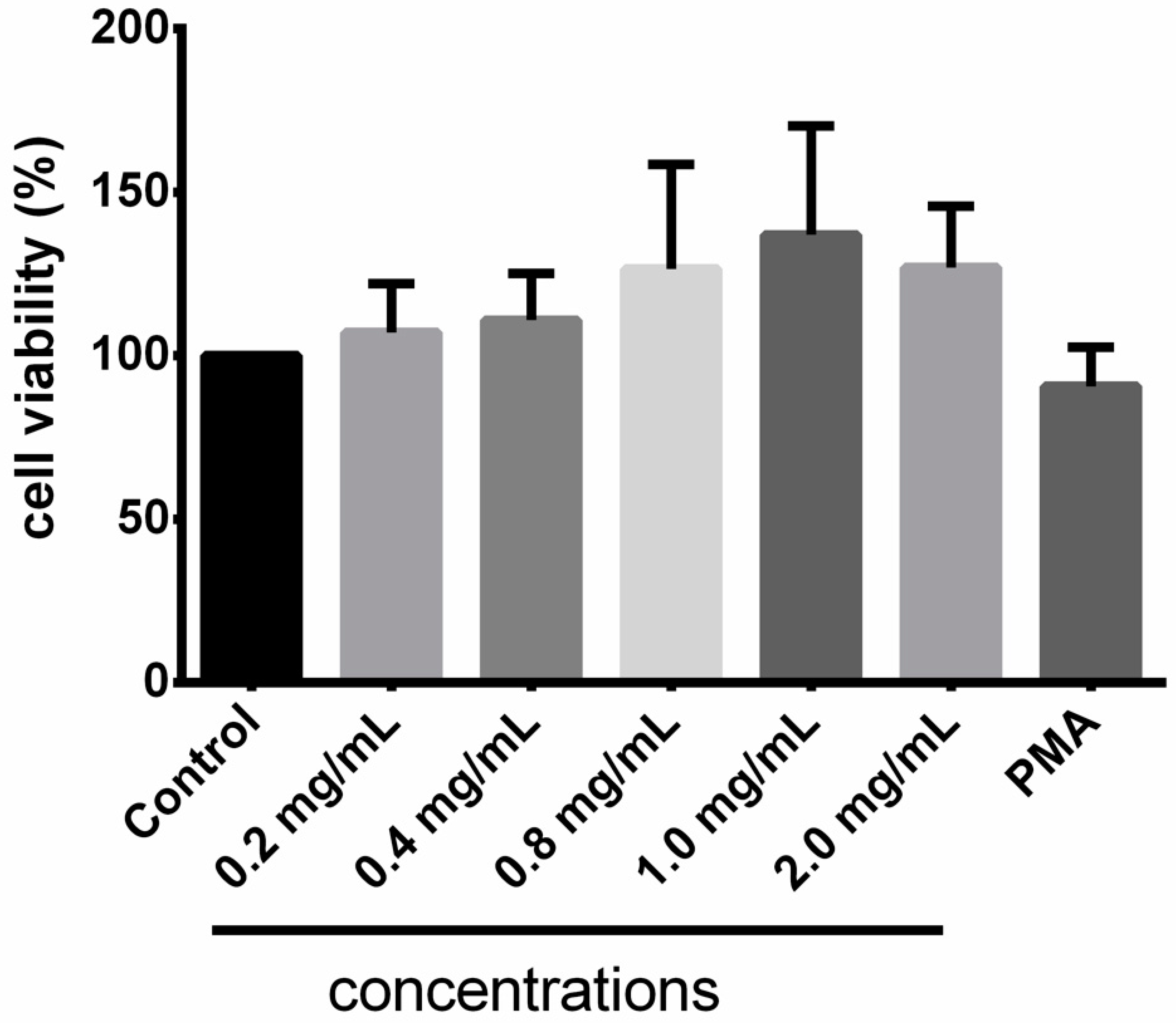
3.4. Cytotoxicity of Taurisolo®
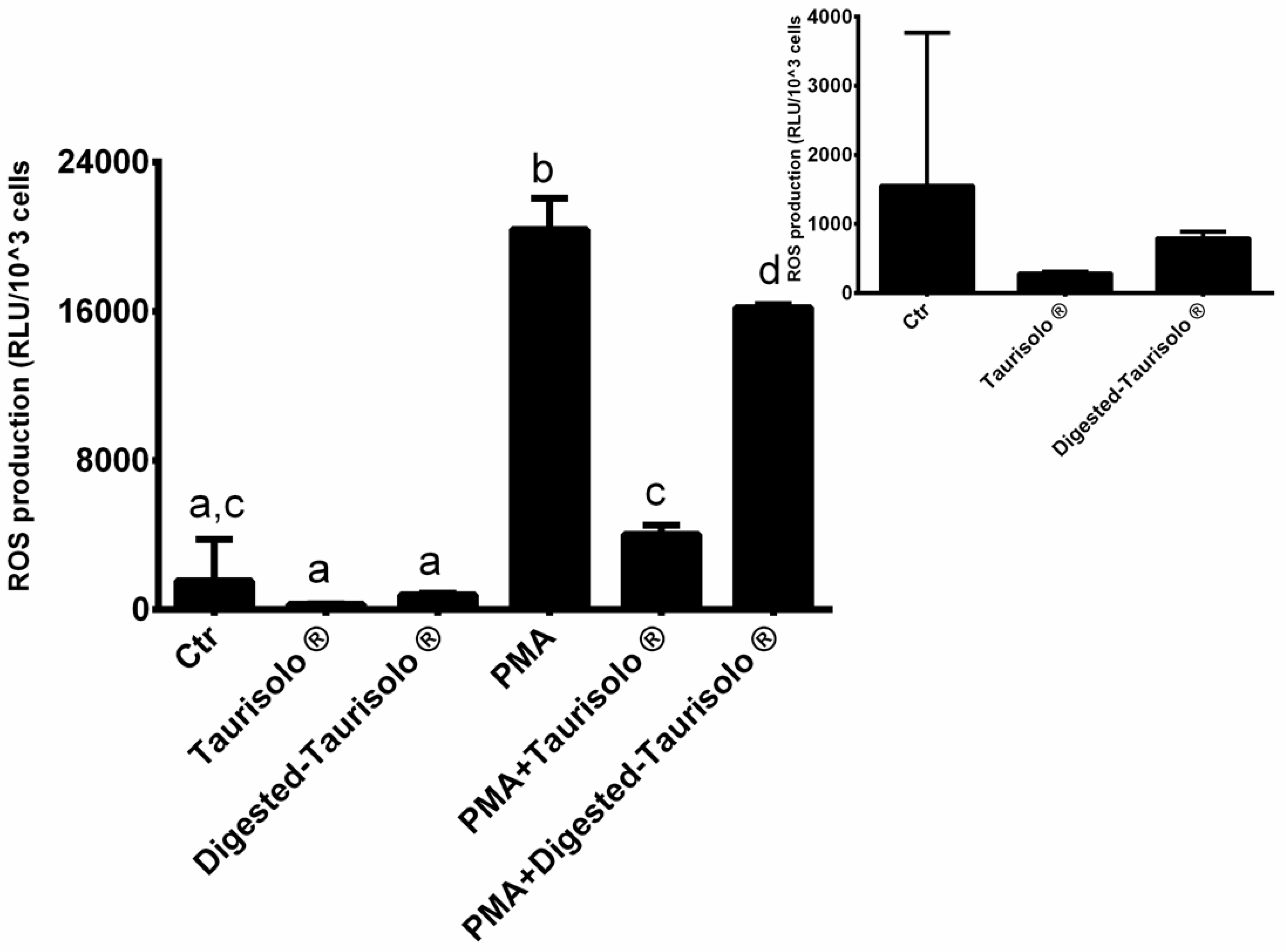
3.5. Taurisolo® Reduces the Levels of ROS
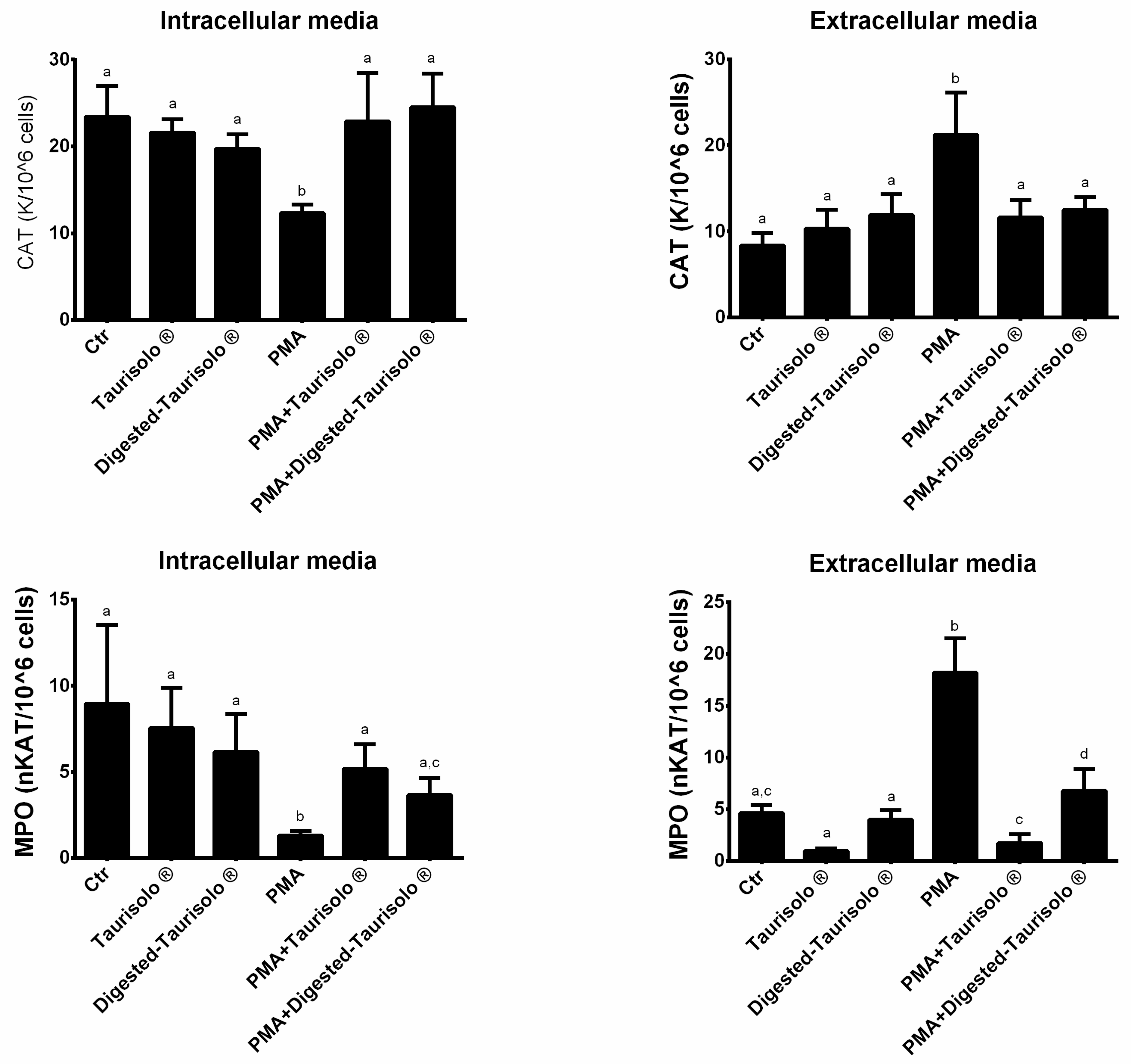
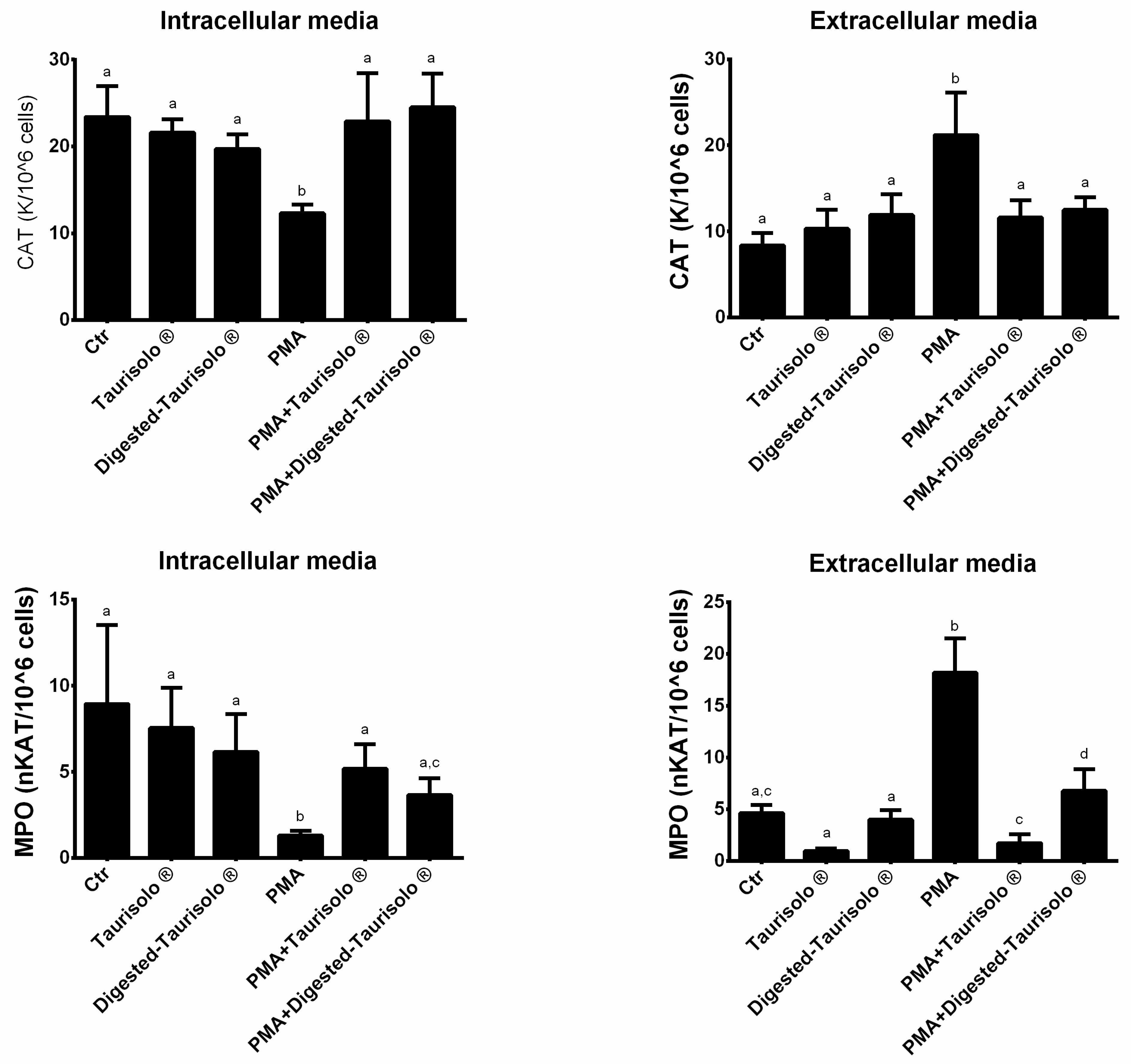
3.6. Taurisolo® Modulates the Enzymatic Activities of CAT and MPO
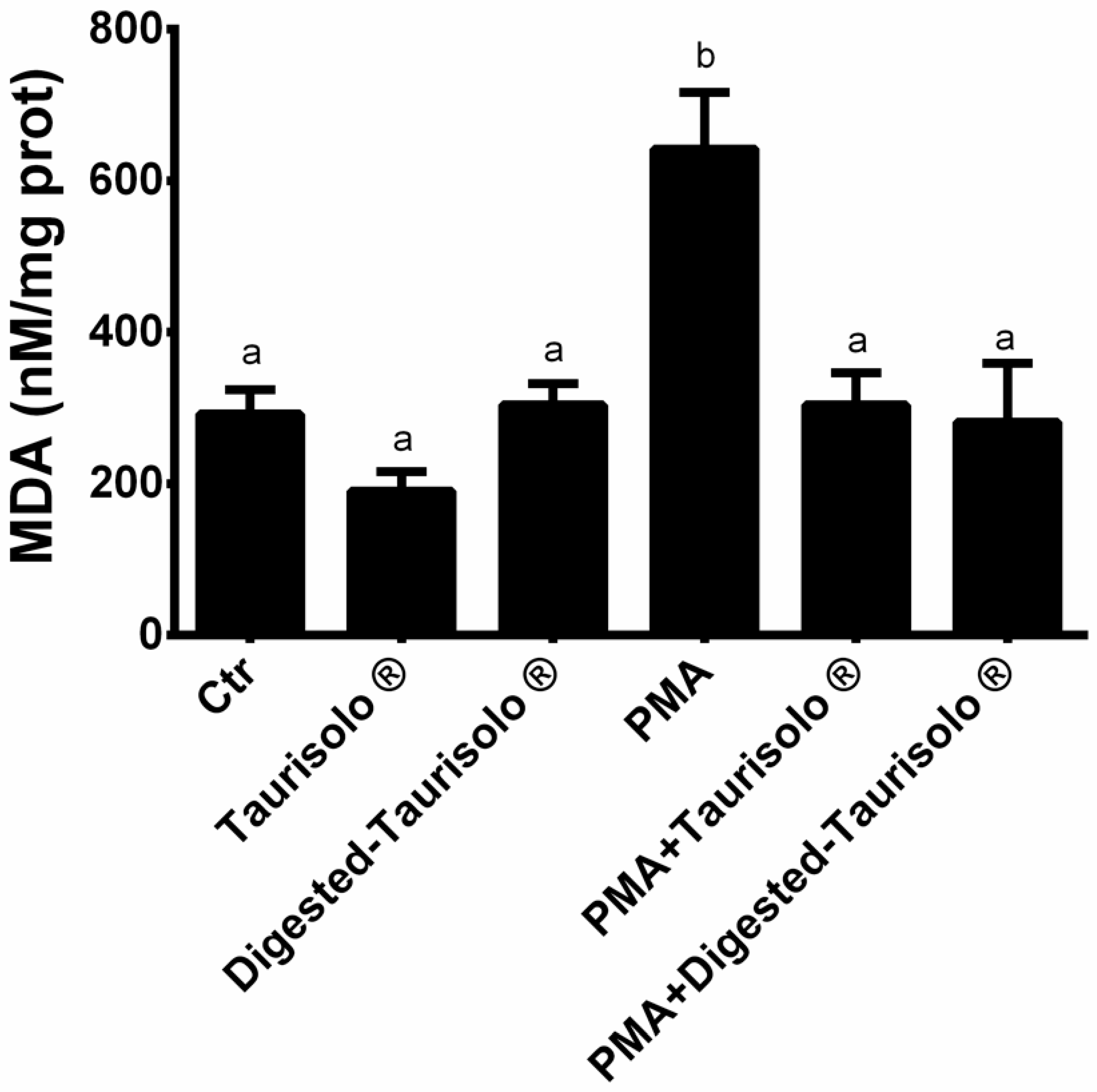
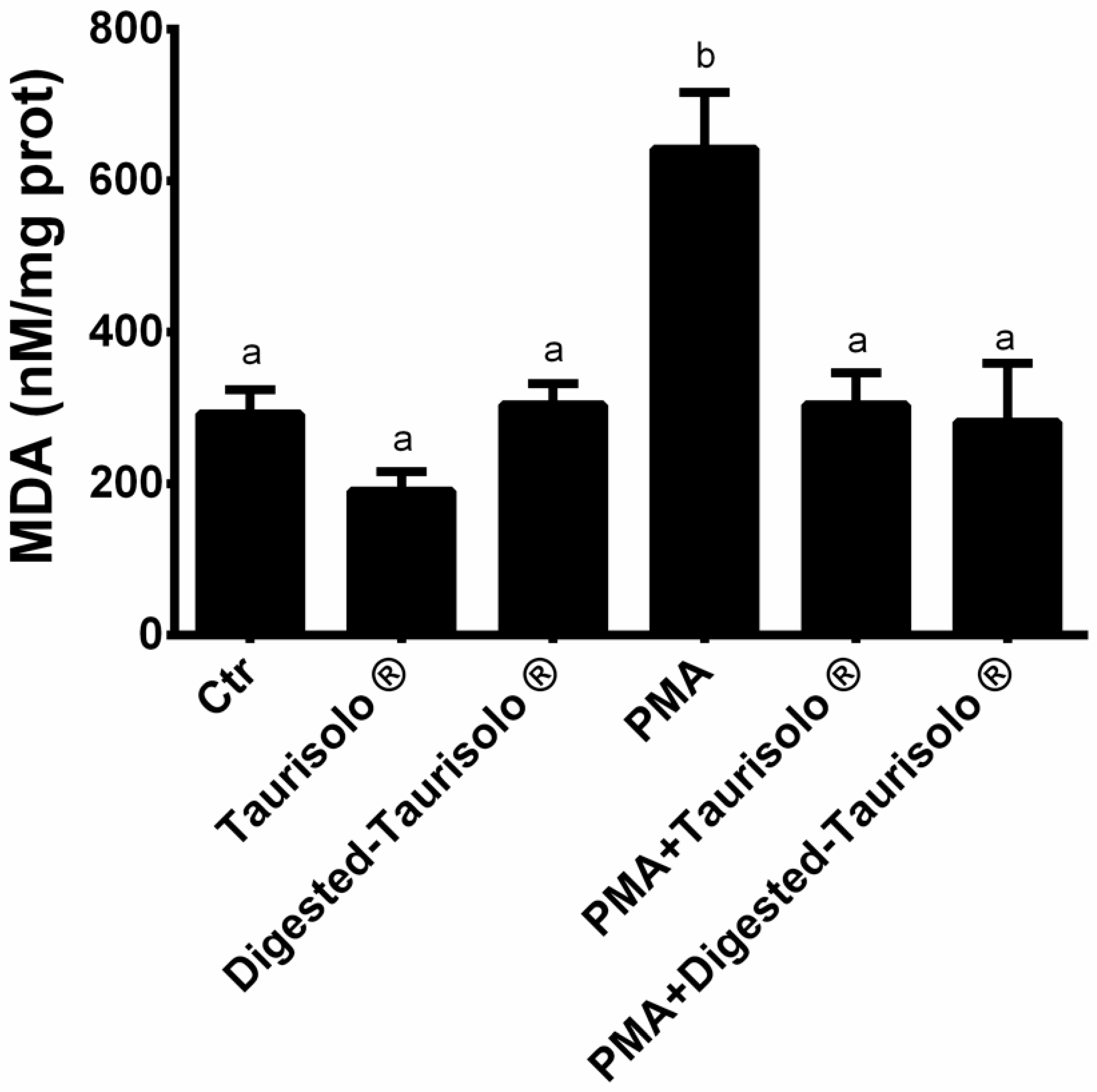
3.7. Taurisolo® Reduces the MDA Levels
3.8. Taurisolo® Modulates the Expression of OxS- and Inflammation-Related Genes
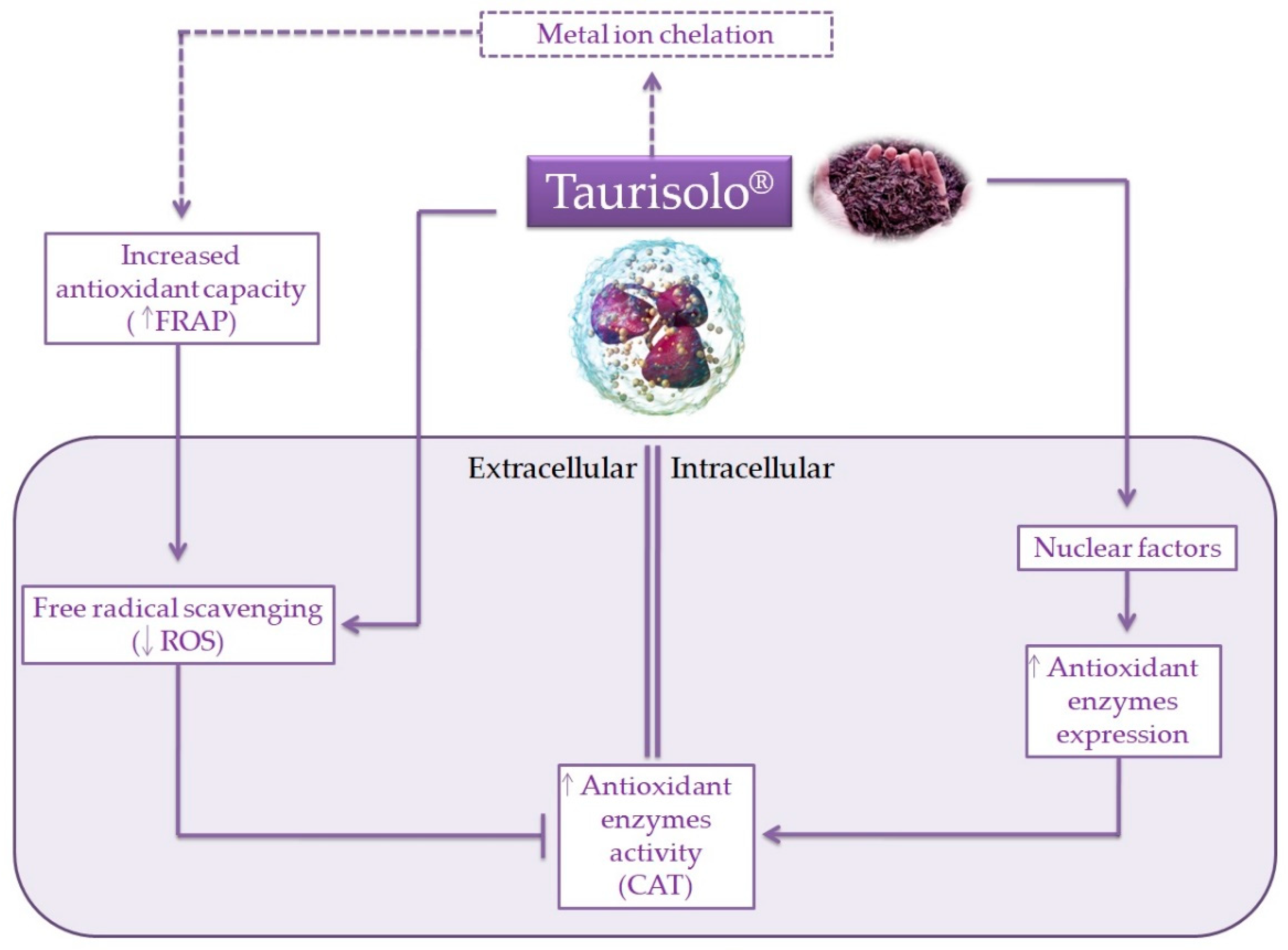
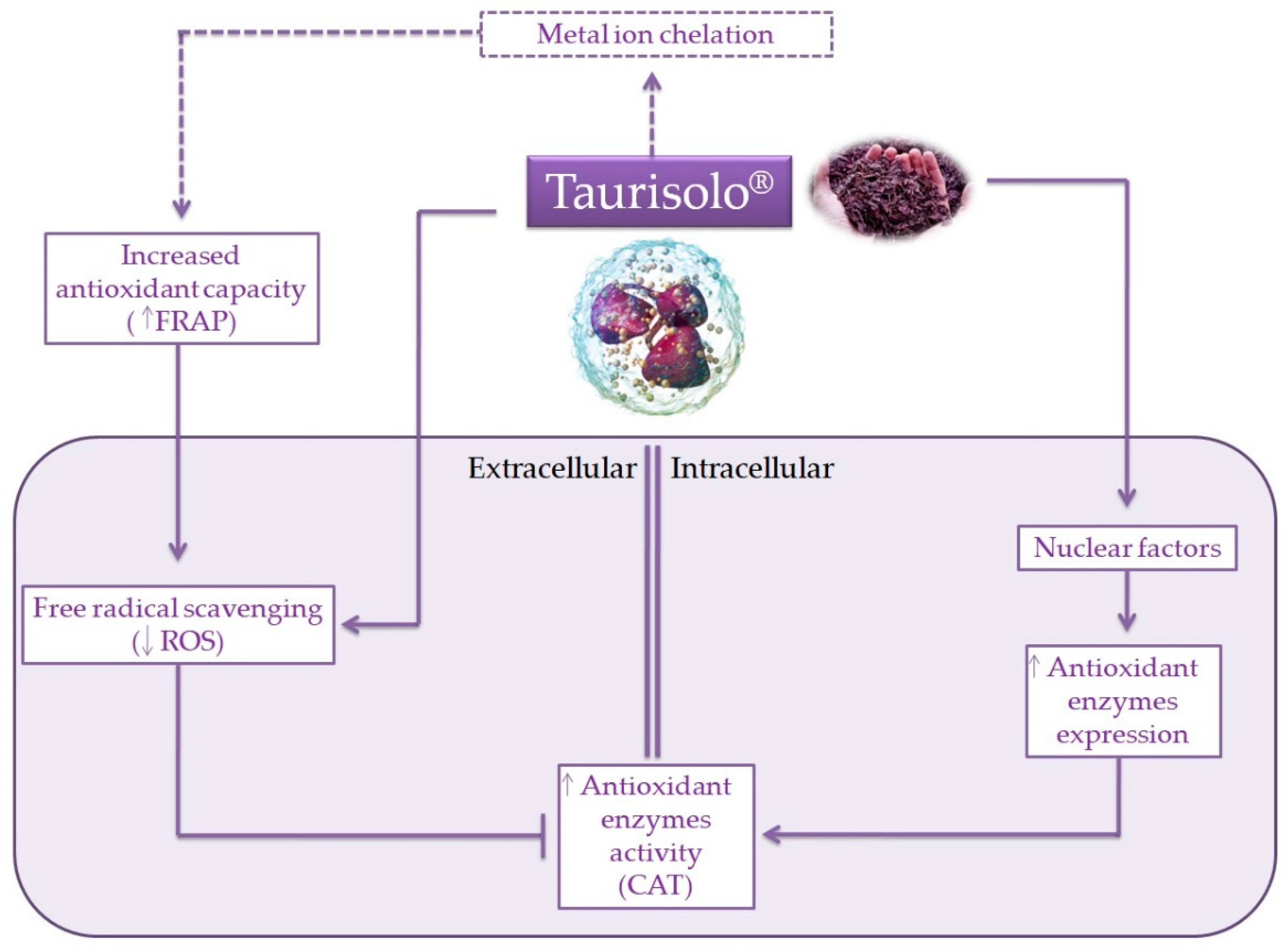
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress: Annual Review of Biochemistry. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Berlett, B.S.; Stadtman, E.R. Protein oxidation in aging, disease, and oxidative stress. J. Biol. Chem. 1997, 272, 20313–20316. [Google Scholar] [CrossRef] [Green Version]
- Dowling, D.K.; Simmons, L.W. Reactive oxygen species as universal constraints in life-history evolution. Proc. R. Soc. B Biol. Sci. 2009, 276, 1737–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherz-Shouval, R.; Elazar, Z. Regulation of autophagy by ROS: Physiology and pathology. Trends Biochem. Sci. 2011, 36, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Mills, K.; le Cessie, S.; Noordam, R.; van Heemst, D. Ageing, age-related diseases and oxidative stress: What to do next? Ageing Res. Rev. 2020, 57, 100982. [Google Scholar] [CrossRef]
- Castro, L.; Freeman, B.A. Reactive oxygen species in human health and disease. Nutrition 2001, 17, 161–165. [Google Scholar] [CrossRef]
- Acharya, A.; Das, I.; Chandhok, D.; Saha, T. Redox regulation in cancer: A double-edged sword with therapeutic potential. Oxid. Med. Cell. Longev. 2010, 3, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Rosales, C. Neutrophils at the crossroads of innate and adaptive immunity. J. Leukoc. Biol. 2020, 108, 377–396. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Tauler, P.; Aguiló, A.; Cases, N.; Fuentespina, E.; Córdova, A.; Tur, J.; Pons, A. Relation between oxidative stress markers and antioxidant endogenous defences during exhaustive exercise. Free Radic. Res. 2005, 39, 1317–1324. [Google Scholar] [CrossRef]
- Truong, V.L.; Jun, M.; Jeong, W.S. Role of resveratrol in regulation of cellular defense systems against oxidative stress. BioFactors 2018, 44, 36–49. [Google Scholar] [CrossRef]
- Sandoval-Acuña, C.; Ferreira, J.; Speisky, H. Polyphenols and mitochondria: An update on their increasingly emerging ROS-scavenging independent actions. Arch. Biochem. Biophys. 2014, 559, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Szabo, K.; Cătoi, A.F.; Vodnar, D.C. Bioactive Compounds Extracted from Tomato Processing by-Products as a Source of Valuable Nutrients. Plant Foods Hum. Nutr. 2018, 73, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Mildner-Szkudlarz, S.; Bajerska, J.; Górnaś, P.; Segliņa, D.; Pilarska, A.; Jesionowski, T. Physical and Bioactive Properties of Muffins Enriched with Raspberry and Cranberry Pomace Powder: A Promising Application of Fruit By-Products Rich in Biocompounds. Plant Foods Hum. Nutr. 2016, 71, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Vodnar, D.C.; Călinoiu, L.F.; Dulf, F.V.; Ştefănescu, B.E.; Crişan, G.; Socaciu, C. Identification of the bioactive compounds and antioxidant, antimutagenic and antimicrobial activities of thermally processed agro-industrial waste. Food Chem. 2017, 231, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, M.; Lucas-Gonzalez, R.; Sayas-Barberá, E.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Bioaccessibility of Phenolic Compounds and Antioxidant Capacity of Chia (Salvia hispanica L.) Seeds. Plant Foods Hum. Nutr. 2018, 73, 47–53. [Google Scholar] [CrossRef] [PubMed]
- de Camargo, A.C.; Schwember, A.R.; Parada, R.; Garcia, S.; Maróstica Júnior, M.R.; Franchin, M.; Regitano-D’arce, M.A.B.; Shahidi, F. Opinion on the hurdles and potential health benefits in value-added use of plant food processing by-products as sources of phenolic compounds. Int. J. Mol. Sci. 2018, 19, 3498. [Google Scholar] [CrossRef] [Green Version]
- García Herrera, P.; Sánchez-Mata, M.C.; Cámara, M. Nutritional characterization of tomato fiber as a useful ingredient for food industry. Innov. Food Sci. Emerg. Technol. 2010, 11, 707–711. [Google Scholar] [CrossRef]
- de Oliveira, W.P.; Biasoto, A.C.T.; Marques, V.F.; dos Santos, I.M.; Magalhães, K.; Correa, L.C.; Negro-Dellacqua, M.; Miranda, M.S.; de Camargo, A.C.; Shahidi, F. Phenolics from Winemaking By-Products Better Decrease VLDL-Cholesterol and Triacylglycerol Levels than Those of Red Wine in Wistar Rats. J. Food Sci. 2017, 82, 2432–2437. [Google Scholar] [CrossRef]
- De Camargo, A.C.; Regitano-D’Arce, M.A.B.; Biasoto, A.C.T.; Shahidi, F. Low molecular weight phenolics of grape juice and winemaking byproducts: Antioxidant activities and inhibition of oxidation of human low-density lipoprotein cholesterol and DNA strand breakage. J. Agric. Food Chem. 2014, 62, 12159–12171. [Google Scholar] [CrossRef]
- Melo, P.S.; Massarioli, A.P.; Denny, C.; Dos Santos, L.F.; Franchin, M.; Pereira, G.E.; Vieira, T.M.F.D.S.; Rosalen, P.L.; De Alencar, S.M. Winery by-products: Extraction optimization, phenolic composition and cytotoxic evaluation to act as a new source of scavenging of reactive oxygen species. Food Chem. 2015, 181, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Oldoni, T.L.C.; Melo, P.S.; Massarioli, A.P.; Moreno, I.A.M.; Bezerra, R.M.N.; Rosalen, P.L.; Da Silva, G.V.J.; Nascimento, A.M.; Alencar, S.M. Bioassay-guided isolation of proanthocyanidins with antioxidant activity from peanut (Arachis hypogaea) skin by combination of chromatography techniques. Food Chem. 2016, 192, 306–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giusti, F.; Caprioli, G.; Ricciutelli, M.; Vittori, S.; Sagratini, G. Determination of fourteen polyphenols in pulses by high performance liquid chromatography-diode array detection (HPLC-DAD) and correlation study with antioxidant activity and colour. Food Chem. 2017. [Google Scholar] [CrossRef]
- Martelli, A.; Flori, L.; Gorica, E.; Piragine, E.; Saviano, A.; Annunziata, G.; Di Minno, M.N.D.; Ciampaglia, R.; Calcaterra, I.; Maione, F.; et al. Vascular Effects of the Polyphenolic Nutraceutical Supplement Taurisolo®: Focus on the Protection of the Endothelial Function. Nutrients 2021, 13, 1540. [Google Scholar] [CrossRef]
- Annunziata, G.; Maisto, M.; Schisano, C.; Ciampaglia, R.; Daliu, P.; Narciso, V.; Tenore, G.C.; Novellino, E. Colon bioaccessibility and antioxidant activity of white, green and black tea polyphenols extract after in vitro simulated gastrointestinal digestion. Nutrients 2018, 10, 1711. [Google Scholar] [CrossRef] [Green Version]
- Annunziata, G.; Maisto, M.; Schisano, C.; Ciampaglia, R.; Narciso, V.; Tenore, G.C.; Novellino, E. Effects of grape pomace polyphenolic extract (Taurisolo ®) in reducing tmao serum levels in humans: Preliminary results from a randomized, placebo-controlled, cross-over study. Nutrients 2019, 11, 139. [Google Scholar] [CrossRef] [Green Version]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Bøyum, A. Separation of white blood cells. Nature 1964, 204, 793–794. [Google Scholar] [CrossRef]
- Capó, X.; Martorell, M.; Sureda, A.; Tur, J.; Pons, A. Effects of docosahexaenoic supplementation and in vitro vitamin C on the oxidative and inflammatory neutrophil response to activation. Oxid. Med. Cell. Longev. 2015, 2015, 187849. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.D.; Sureda, A.; Mestre, A.; Tur, J.A.; Pons, A. The double edge of reactive oxygen species as damaging and signaling molecules in HL60 cell culture. Cell. Physiol. Biochem. 2010, 25, 241–252. [Google Scholar] [CrossRef]
- Capeillère-Blandin, C. Oxidation of guaiacol by myeloperoxidase: A two-electron-oxidized guaiacol transient species as a mediator of NADPH oxidation. Biochem. J. 1998, 336, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, G.; Jimenez-García, M.; Tejada, S.; Moranta, D.; Arnone, A.; Ciampaglia, R.; Tenore, G.C.; Sureda, A.; Novellino, E.; Capó, X. Grape polyphenols ameliorate muscle decline reducing oxidative stress and oxidative damage in aged rats. Nutrients 2020, 12, 1280. [Google Scholar] [CrossRef] [PubMed]
- Rezadoost, M.H.; Kordrostami, M.; Kumleh, H.H. An efficient protocol for isolation of inhibitor-free nucleic acids even from recalcitrant plants. 3 Biotech 2016, 6, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeles, J.G.C.; Laurena, A.C.; Tecson-Mendoza, E.M. Extraction of genomic DNA from the lipid-, polysaccharide-, and polyphenol-rich coconut (Cocos nucifera L.). Plant Mol. Biol. Report. 2005, 23, 297–298. [Google Scholar] [CrossRef]
- Lapi, D.; Stornaiuolo, M.; Sabatino, L.; Sommella, E.; Tenore, G.; Daglia, M.; Scuri, R.; Di Maro, M.; Colantuoni, A.; Novellino, E. The Pomace Extract Taurisolo Protects Rat Brain From Ischemia-Reperfusion Injury. Front. Cell. Neurosci. 2020, 14, 3. [Google Scholar] [CrossRef]
- Annunziata, G.; Maisto, M.; Schisano, C.; Ciampaglia, R.; Narciso, V.; Hassan, S.T.S.; Tenore, G.C.; Novellino, E. Effect of grape pomace polyphenols with or without pectin on TMAO serum levels assessed by LC/MS-based assay: A preliminary clinical study on overweight/obese subjects. Front. Pharmacol. 2019, 10, 575. [Google Scholar] [CrossRef] [Green Version]
- Khaybullina, Z.R. Citation: Khaybullina ZR (2017) Inflammation and Oxidative Stress: Critical Role for Metabolic Syndrome. J. Vasc. Med. Surg. 2017, 5, 302. [Google Scholar] [CrossRef]
- Holvoet, P. Relations between metabolic syndrome, oxidative stress and inflammation and cardiovascular disease. Verh. K. Acad. Geneeskd. Belg. 2008, 70, 193–219. [Google Scholar]
- Devaraj, S.; Goyal, R.; Jialal, I. Inflammation, oxidative stress, and the metabolic syndrome. US Endocrinol. 2008, 4, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Carrier, A. Metabolic Syndrome and Oxidative Stress: A Complex Relationship. Antioxid. Redox Signal. 2017, 26, 429–431. [Google Scholar] [CrossRef]
- Ellulu, M.S.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity & inflammation: The linking mechanism & the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2015, 16, 378–400. [Google Scholar] [CrossRef] [Green Version]
- McMurray, F.; Patten, D.A.; Harper, M.E. Reactive Oxygen Species and Oxidative Stress in Obesity—Recent Findings and Empirical Approaches. Obesity 2016, 24, 2301–2310. [Google Scholar] [CrossRef]
- Lama, S.; Monda, V.; Rizzo, M.R.; Dacrema, M.; Maisto, M.; Annunziata, G.; Tenore, G.C.; Novellino, E.; Stiuso, P. Cardioprotective Effects of Taurisolo® in Cardiomyoblast H9c2 Cells under High-Glucose and Trimethylamine N-Oxide Treatment via De Novo Sphingolipid Synthesis. Oxid. Med. Cell. Longev. 2020, 2020, 2961406. [Google Scholar] [CrossRef] [PubMed]
- Bors, W.; Heller, W.; Michel, C.; Saran, M. Flavonoids as antioxidants: Determination of radical-scavenging efficiencies. Methods Enzymol. 1990, 186, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Amic, D.; Davidovic-Amic, D.; Beslo, D.; Rastija, V.; Lucic, B.; Trinajstic, N. SAR and QSAR of the Antioxidant Activity of Flavonoids. Curr. Med. Chem. 2007, 14, 827–845. [Google Scholar] [CrossRef]
- Niwa, Y.; Ozaki, Y.; Kanoh, T.; Akamatsu, H.; Kurisaka, M. Role of cytokines, tyrosine kinase, and protein kinase C on production of superoxide and induction of scavenging enzymes in human leukocytes. Clin. Immunol. Immunopathol. 1996, 79, 303–313. [Google Scholar] [CrossRef]
- Metzler, K.D.; Goosmann, C.; Lubojemska, A.; Zychlinsky, A.; Papayannopoulos, V. A myeloperoxidase-containing complex regulates neutrophil elastase release and actin dynamics during NETosis. Cell Rep. 2014, 8, 883–896. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, P.A.; Stephenson, K.K.; Wade, K.L.; Liu, H.; Fahey, J.W. Structure-activity analysis of flavonoids: Direct and indirect antioxidant, and antiinflammatory potencies and toxicities. Nutr. Cancer 2013, 65, 1014–1025. [Google Scholar] [CrossRef]
- Ji, L.L.; Gomez-Cabrera, M.C.; Vina, J. Exercise and hormesis: Activation of cellular antioxidant signaling pathway. Ann. N. Y. Acad. Sci. 2006, 1067, 425–435. [Google Scholar] [CrossRef]
- Calabrese, V.; Cornelius, C.; Dinkova-Kostova, A.T.; Calabrese, E.J.; Mattson, M.P. Cellular stress responses, the hormesis paradigm, and vitagenes: Novel targets for therapeutic intervention in neurodegenerative disorders. Antioxid. Redox Signal. 2010, 13, 1763–1811. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Scuto, M.; Fusco, R.; Trovato, A.; Ontario, M.L.; Crea, R.; Di Paola, R.; Cuzzocrea, S.; Calabrese, V. Anti-inflammatory and Anti-oxidant Activity of Hidrox ® in Rotenone-Induced Parkinson’s Disease in Mice. Antioxidants 2020, 9, 824. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, N.; Sfogliano, L.; Calabrese, V.; Rizza, V. Effects of multivitamin treatment on the activity of rat liver tryptophan pyrrolase during ethanol administration. Acta Vitaminol. Enzym. 1981, 3, 199–204. [Google Scholar]
- Calabrese, V.; Renis, M.; Calderone, A.; Russo, A.; Barcellona, M.L.; Rizza, V. Stress proteins and SH-groups in oxidant-induced cell damage after acute ethanol administration in rat. Free Radic. Biol. Med. 1996, 20, 391–397. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Calabrese, V.; Giordano, J. Demonstrated hormetic mechanisms putatively subserve riluzole-induced effects in neuroprotection against amyotrophic lateral sclerosis (ALS): Implications for research and clinical practice. Ageing Res. Rev. 2021, 67, 101273. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Mattson, M.P.; Dhawan, G.; Kapoor, R.; Calabrese, V.; Giordano, J. Hormesis: A potential strategic approach to the treatment of neurodegenerative disease. Int. Rev. Neurobiol. 2020, 155, 271–301. [Google Scholar] [CrossRef]
- Brunetti, G.; Di Rosa, G.; Scuto, M.; Leri, M.; Stefani, M.; Schmitz-Linneweber, C.; Calabrese, V.; Saul, N. Healthspan Maintenance and Prevention of Parkinson’s-like Phenotypes with Hydroxytyrosol and Oleuropein Aglycone in C. elegans. Int. J. Mol. Sci. 2020, 21, 2588. [Google Scholar] [CrossRef] [Green Version]
- Negre-Salvayre, A.; Coatrieux, C.; Ingueneau, C.; Salvayre, R. Advanced lipid peroxidation end products in oxidative damage to proteins. Potential role in diseases and therapeutic prospects for the inhibitors. Br. J. Pharmacol. 2008, 153, 6–20. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Cheeseman, K.H.; Dianzani, M.U.; Poli, G.; Slater, T.F. Separation and characterization of the aldehydic products of lipid peroxidation stimulated by ADP-Fe2+ in rat liver microsomes. Biochem. J. 1982, 208, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Caruso, F.; Tanski, J.; Villegas-Estrada, A.; Rossi, M. Structural basis for antioxidant activity of trans-resveratrol: Ab initio calculations and crystal and molecular structure. J. Agric. Food Chem. 2004, 52, 7279–7285. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Sequence | Temperature (°C) |
|---|---|---|---|
| COX-2 | NM_000963 | Fw: 5′-TTGCCTGGCAGGGTTGCTGGTGGTA-3′ | 95 °C 10 Seg 63 °C 10 Seg 72 °C 15 Seg |
| Rev: 5′-CATCTGCCTGCTCTGGTCAATGGAA-3′ | |||
| IL-10 | NM_000572 | Fw: 5′-AGAACCTGAAGACCCTCAGGC-3′ | 95 °C 10 Seg 58 °C 10 Seg 72 °C 15 Seg |
| Rev: 5′-CCACGGCCTTGCTCTTGTT-3′ | |||
| TNFα | NM_000594 | Fw: 5′-CCCAGGCAGTCAGATCATCTTCTCGGAA-3′ | 95 °C 10 Seg 63 °C 10 Seg 72 °C 15 Seg |
| Rev: 5′-CTGGTTATCTCTCAGCTCCACGCCATT-3′ | |||
| MPO | NM_000250 | Fw: 5′-TGAACATGGGGAGTGTTTCA-3′ | 95 °C 10 Seg 62 °C 10 Seg 72 °C 15 Seg |
| Rev: 5′-CCAGCTCTGCTAACCAGGAC-3′ | |||
| 18S | NR_003286 | Fw: 5′-ATG TGA AGT CAC TGT GCC AG-3′ | 95 °C 10 Seg 60 °C 10 Seg 72 °C 12 Seg |
| Rev: 5′-GTG TAA TCC GTC TCC ACA GA-3′ |
| Compound | Mean Value (µg/g) ± SD | |
|---|---|---|
| Taurisolo® | Digested Taurisolo® | |
| Ferulic acid | 14.59 ± 0.98 | 1.92 ± 0.11 |
| Resveratrol | 12.55 ± 0.02 | 0.245 ± 0.004 |
| Caffeic acid | 35.00 ± 3.00 | n.d. |
| p-cumaric acid | 122.75 ± 2.77 | n.d |
| Rutin | 98.81 ± 7.31 | 20.82 ± 0.20 |
| Quercetin | 135.41 ± 4.69 | n.d. |
| Procyanidin B1 dimer | 946.33 ± 55.20 | 116.72 ± 0.52 |
| Procyanidin B2 dimer | 645.89 ± 59.17 | 169.16 ± 0.77 |
| Syringic acid | 310.95 ± 0.01 | 122.57 ± 0.55 |
| Epicatechin | 1696.55 ± 109.60 | 1.38 ± 0.02 |
| Gallic acid | 199.46 ± 4.59 | n.d. |
| Catechin | 2499.04 ± 307.41 | 77.95 ± 0.29 |
| Parameter | Mean ± SEM (n = 17) |
|---|---|
| Age (years) | 63.4 ± 10.9 |
| Anthropometric characteristics | |
| Weight (kg) | 91.4 ± 17.4 |
| Height (cm) | 169.5 ± 12.1 |
| BMI (kg/m2) | 31.6 ± 3.14 |
| Metabolic parameters | |
| Glucose (mg/dL) | 103.2 ± 27.5 |
| Hb1A (%) | 5.92 ± 1.23 |
| Triglycerides (mg/dL) | 200.6 ± 31.7 |
| HDL-cholesterol (mg/dL) | 40.4 ± 8.57 |
| LDL-cholesterol (mg/dL) | 126.5 ± 28.9 |
| Cholesterol total (mg/dL) | 197.1 ± 40.3 |
| Bilirubin (mg/dL) | 0.683 ± 0.223 |
| AST (U/L) | 21.2 ± 4.02 |
| ALT (U/L) | 24.9 ± 9.96 |
| GGT (U/L) | 39.1 ± 31.4 |
| PKC (mg/dL) | 0.770 ±0.310 |
| Hematological parameters | |
| Hematocrit (%) | 46.0 ± 2.69 |
| Erythrocytes (106/mm3) | 4.99 ± 0.374 |
| Leukocytes (103/mm3) | 7.83 ± 1.63 |
| Neutrophils (103/mm3) | 4.30 ± 1.39 |
| Lymphocytes (103/mm3) | 2.58 ± 0.763 |
| Basophils (103/mm3) | 0.063 ± 0.038 |
| Monocytes (103/mm3) | 0.648 ± 0.189 |
| Eosinophils (103/mm3) | 0.238 ± 0.084 |
| Platelets (103/mm3) | 218.7 ± 62.5 |
| Sample | DPPH (mmol TE/g) | ABTS (mmol TE/g) | FRAP (mM TE) |
|---|---|---|---|
| Taurisolo® | 3.67 ± 0.55 | 3.47 ± 0.11 | 0.705 |
| Digested-Taurisolo® | 2.55 ± 0.76 | 2.33 ± 0.66 | 0.530 |
| Control | Digested-Taurisolo® | PMA | PMA + Digested-Taurisolo® | ANOVA | |||
|---|---|---|---|---|---|---|---|
| T | A | TxA | |||||
| COX-2 | 1.00 ± 0.28 a | 1.87 ± 0.33 a | 10.4 ± 4.59 b | 0.54 ± 0.12 a | x | ||
| IL-10 | 1.00 ± 0.27 | 8.23 ± 2.34 | 5.25 ± 1.82 | 9.87 ± 4.19 | |||
| TNFα | 1.00 ± 0.59 ab | 0.21 ± 0.04 a | 5.32 ± 1.17 c | 2.80 ± 0.86 b | x | x | |
| MPO | 1.00 ± 0.21 a | 0.45 ± 0.07 b | 1.25 ± 0.22 b | 0.41 ± 0.05 b | x | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Annunziata, G.; Capó, X.; Quetglas-Llabrés, M.M.; Monserrat-Mesquida, M.; Tejada, S.; Tur, J.A.; Ciampaglia, R.; Guerra, F.; Maisto, M.; Tenore, G.C.; et al. Ex Vivo Study on the Antioxidant Activity of a Winemaking By-Product Polyphenolic Extract (Taurisolo®) on Human Neutrophils. Antioxidants 2021, 10, 1009. https://doi.org/10.3390/antiox10071009
Annunziata G, Capó X, Quetglas-Llabrés MM, Monserrat-Mesquida M, Tejada S, Tur JA, Ciampaglia R, Guerra F, Maisto M, Tenore GC, et al. Ex Vivo Study on the Antioxidant Activity of a Winemaking By-Product Polyphenolic Extract (Taurisolo®) on Human Neutrophils. Antioxidants. 2021; 10(7):1009. https://doi.org/10.3390/antiox10071009
Chicago/Turabian StyleAnnunziata, Giuseppe, Xavier Capó, Maria Magdalena Quetglas-Llabrés, Margalida Monserrat-Mesquida, Silvia Tejada, Josep A. Tur, Roberto Ciampaglia, Fabrizia Guerra, Maria Maisto, Gian Carlo Tenore, and et al. 2021. "Ex Vivo Study on the Antioxidant Activity of a Winemaking By-Product Polyphenolic Extract (Taurisolo®) on Human Neutrophils" Antioxidants 10, no. 7: 1009. https://doi.org/10.3390/antiox10071009
APA StyleAnnunziata, G., Capó, X., Quetglas-Llabrés, M. M., Monserrat-Mesquida, M., Tejada, S., Tur, J. A., Ciampaglia, R., Guerra, F., Maisto, M., Tenore, G. C., Novellino, E., & Sureda, A. (2021). Ex Vivo Study on the Antioxidant Activity of a Winemaking By-Product Polyphenolic Extract (Taurisolo®) on Human Neutrophils. Antioxidants, 10(7), 1009. https://doi.org/10.3390/antiox10071009