
A Balance between the Activities of Chloroplasts and Mitochondria Is Crucial for Optimal Plant Growth

 , , , ,
, , , , 
Abstract
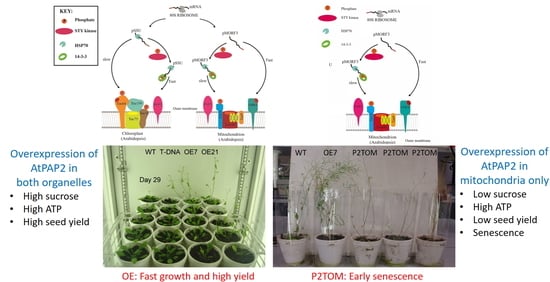
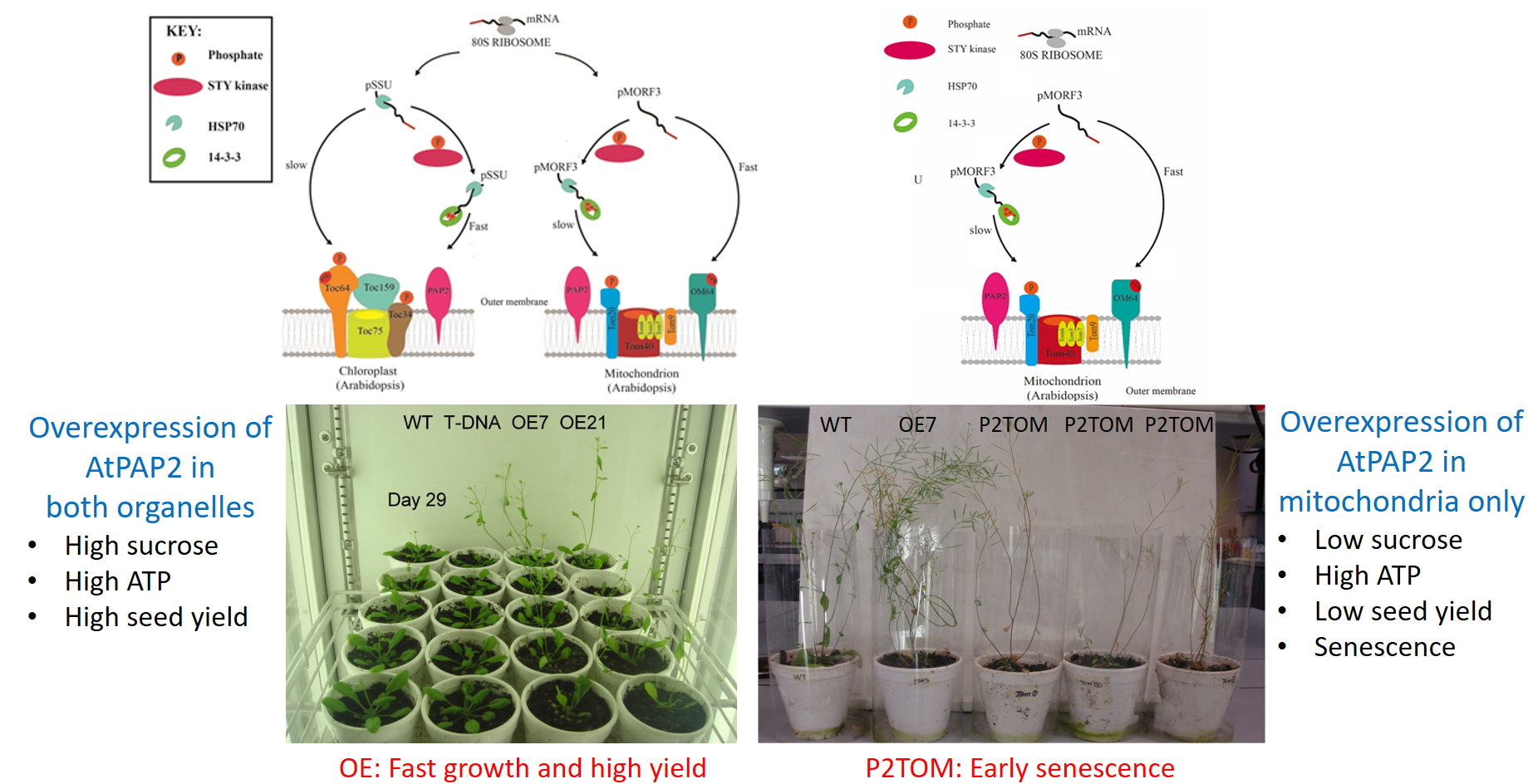
1. Introduction
2. Materials and Methods
2.1. Plant Lines and Growth Conditions
2.2. Cloning of C-tails into Enhanced Green Fluorescent (EGFP) Constructs
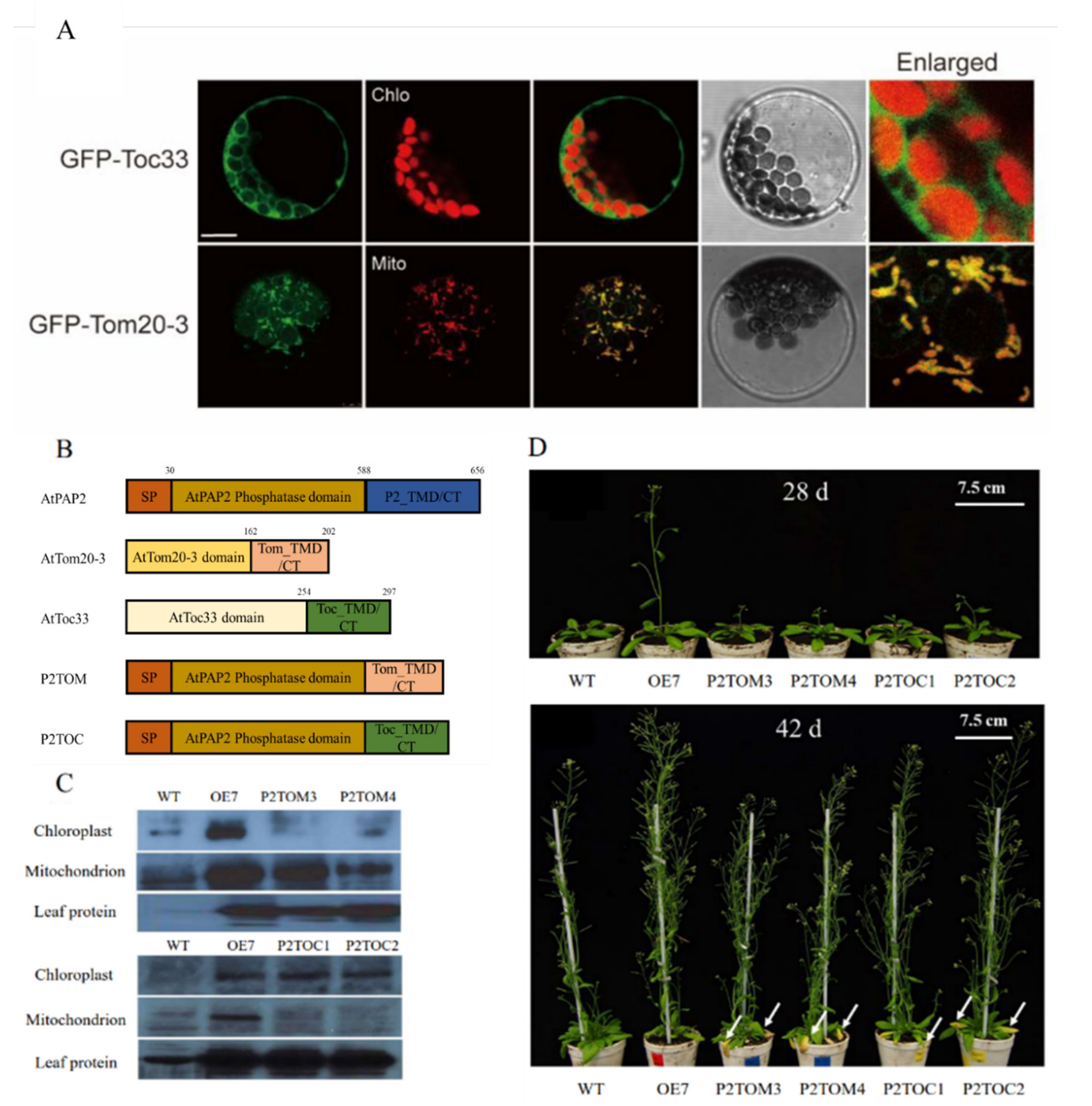
2.3. Generation of Transgenic Line of P2TOM and P2TOC in Arabidopsis
2.4. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis (SDS-PAGE)/Western Blotting
2.5. Isolation of Mitochondria from Arabidopsis Leaves Using Continuous Gradient Centrifugation
2.6. Isolation of Chloroplast from Arabidopsis Leaves Using Discontinuous Gradient Centrifugation
2.7. Protoplast Transient Expression Assay
2.8. Enzymatic Assays
2.9. Chlorophyll Fluorescence Measurement
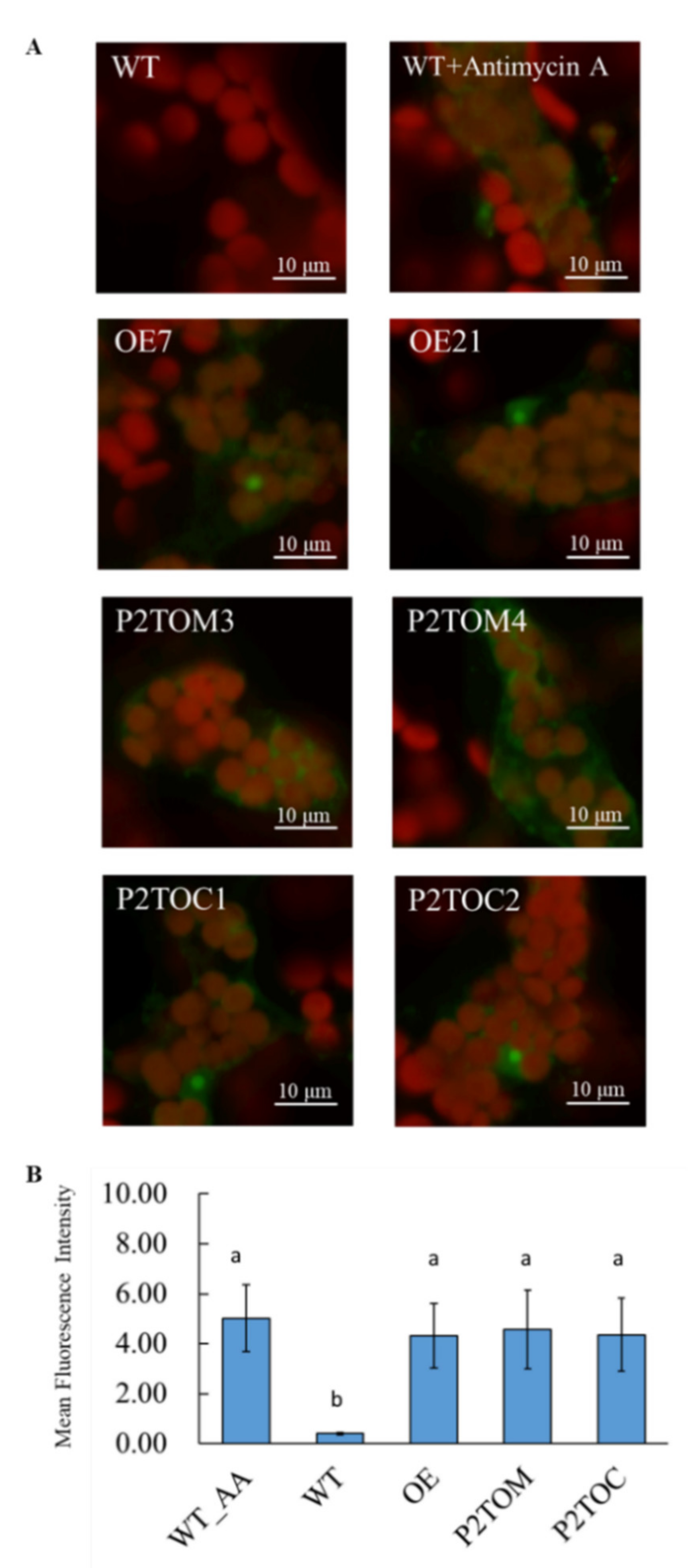
2.10. Imaging of Leaf Mitochondrial Superoxide (O2●−)
2.11. Luciferase ATP/ADP Measurement
2.12. Gas Chromatography-Mass Spectrometry (GC-MS/MS) Measurement
3. Results
3.1. Growth Phenotypes of P2TOM and P2TOC Lines
3.2. Mitochondrial Activity of OE7, P2TOM, and P2TOC Lines was Higher Than WT
3.3. Chloroplastic Activity was Higher in OE7 and P2TOC but Lower in P2TOM Than in WT
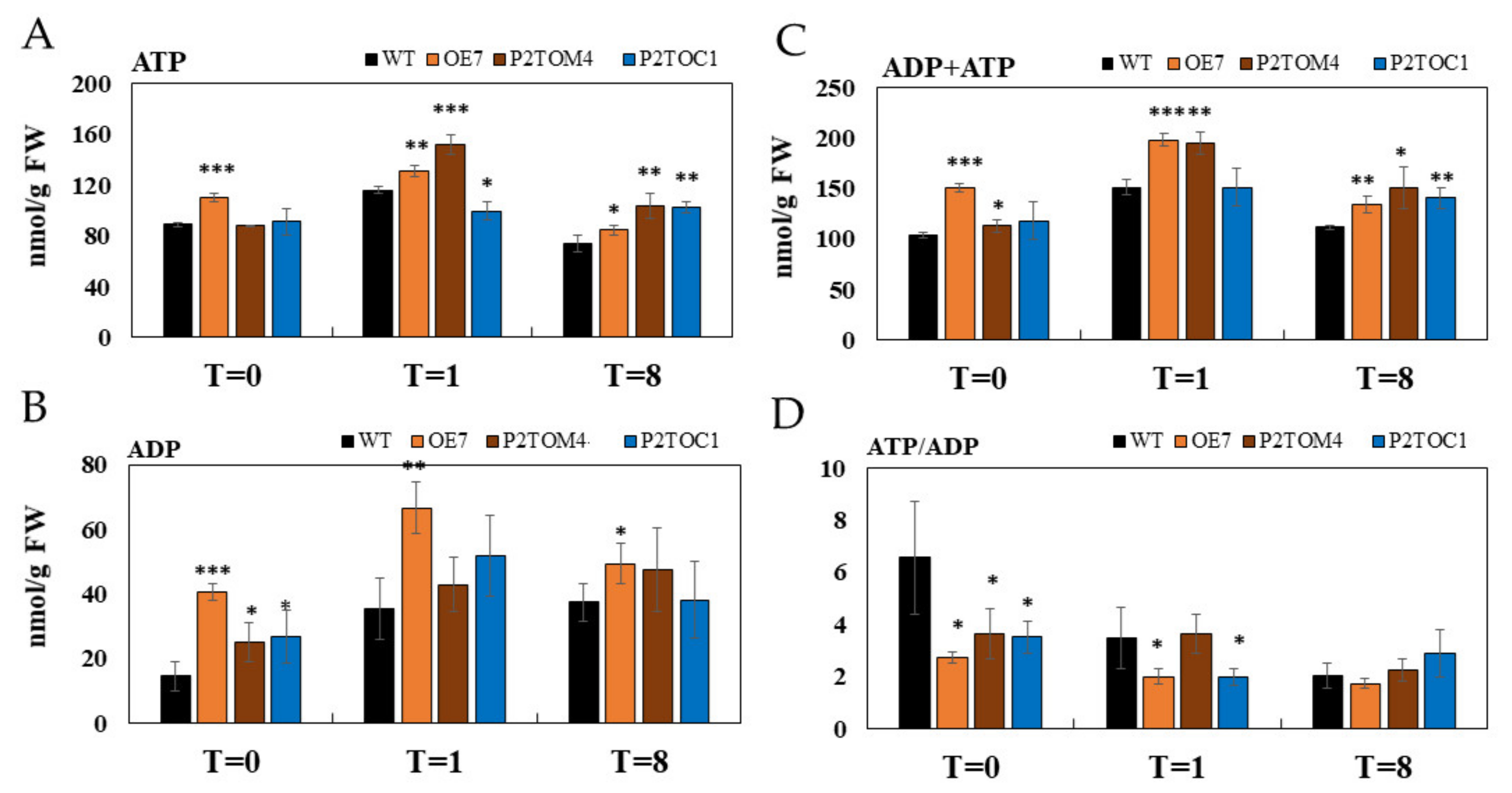
3.4. Impacts of AtPAP2 Overexpression on the Plant Energy Status
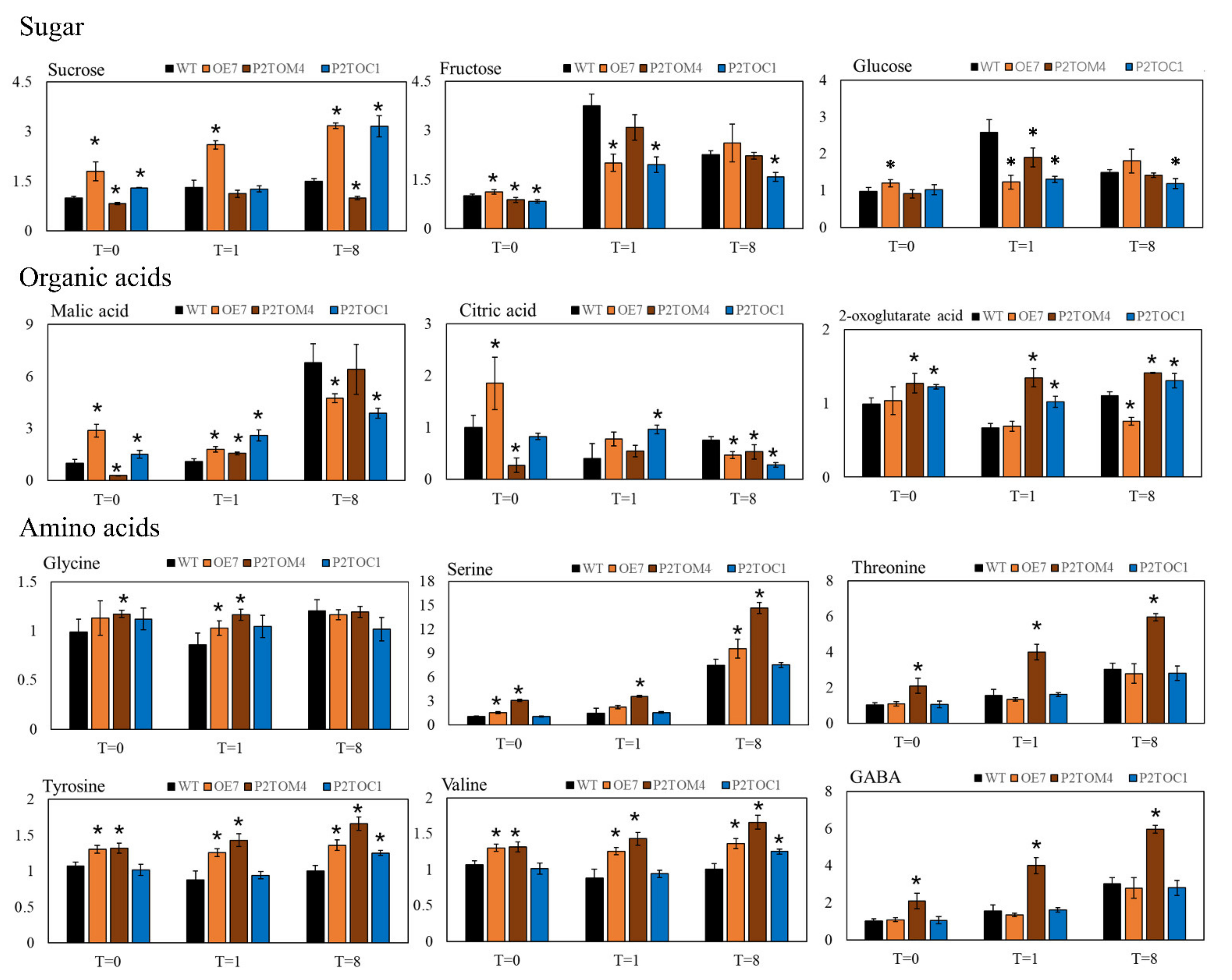
3.5. Impact of AtPAP2 Overexpression on Leaf Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nunes-Nesi, A.; Araújo, W.L.; Fernie, A.R. Targeting mitochondrial metabolism and machinery as a means to enhance photosynthesis. Plant Physiol. 2011, 155, 101–107. [Google Scholar] [CrossRef]
- Kong, F.; Burlacot, A.; Liang, Y.; Légeret, B.; Alseekh, S.; Brotman, Y.; Fernie, A.R.; Krieger-Liszkay, A.; Beisson, F.; Peltier, G. Interorganelle Communication: Peroxisomal MALATE DEHYDROGENASE2 connects lipid catabolism to photosynthesis through redox coupling in Chlamydomonas. Plant Cell 2018, 30, 1824–1847. [Google Scholar] [CrossRef] [PubMed]
- Voon, C.P.; Lim, B.L. ATP translocation and chloroplast biology. Natl. Sci. Rev. 2019, 6, 1073–1076. [Google Scholar] [CrossRef]
- Voon, C.P.; Law, Y.S.; Guan, X.; Lim, S.L.; Xu, Z.; Chu, W.T.; Zhang, R.; Sun, F.; Labs, M.; Pribil, M.; et al. Modulating the activities of chloroplasts and mitochondria promotes ATP production and plant growth. Quant. Plant Biol. 2021, 2, e7. [Google Scholar] [CrossRef]
- Horton, P.; Ruban, A.; Walters, R. Regulation of light harvesting in green plants. Annu. Rev. Plant Biol. 1996, 47, 655–684. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef]
- Niyogi, K.K. Safety valves for photosynthesis. Curr. Opin. Plant Biol. 2000, 3, 455–460. [Google Scholar] [CrossRef]
- Scheibe, R. Maintaining homeostasis by controlled alternatives for energy distribution in plant cells under changing conditions of supply and demand. Photosynth. Res. 2019, 139, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Scheibe, R. Malate valves to balance cellular energy supply. Physiol. Plant. 2004, 120, 21–26. [Google Scholar] [CrossRef]
- Bailleul, B.; Berne, N.; Murik, O.; Petroutsos, D.; Prihoda, J.; Tanaka, A.; Villanova, V.; Bligny, R.; Flori, S.; Falconet, D. Energetic coupling between plastids and mitochondria drives CO 2 assimilation in diatoms. Nature 2015, 524, 366. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.-V.; Plet, J.; Tolleter, D.; Jokel, M.; Cuiné, S.; Carrier, P.; Auroy, P.; Richaud, P.; Johnson, X.; Alric, J. Combined increases in mitochondrial cooperation and oxygen photoreduction compensate for deficiency in cyclic electron flow in Chlamydomonas reinhardtii. Plant Cell 2014, 26, 3036–3050. [Google Scholar] [CrossRef]
- Voon, C.P.; Guan, X.; Sun, Y.; Sahu, A.; Chan, M.N.; Gardeström, P.; Wagner, S.; Fuchs, P.; Nietzel, T.; Versaw, W.K.; et al. ATP compartmentation in plastids and cytosol of Arabidopsis thaliana revealed by fluorescent protein sensing. Proc. Natl. Acad. Sci. USA 2018, 115, E10778–E10787. [Google Scholar] [CrossRef]
- Lim, S.-L.; Voon, C.P.; Guan, X.; Yang, Y.; Gardeström, P.; Lim, B.L. In planta study of photosynthesis and photorespiration using NADPH and NADH/NAD+ fluorescent protein sensors. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, R.; Stitt, M.; Heldt, H.W. Subcellular metabolite levels in spinach leaves: Regulation of sucrose synthesis during diurnal alterations in photosynthetic partitioning. Plant Physiol. 1987, 83, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Carrie, C.; Law, S.; Murcha, M.W.; Zhang, R.; Law, Y.S.; Suen, P.K.; Whelan, J.; Lim, B.L. AtPAP2 is a tail-anchored protein in the outer membrane of chloroplasts and mitochondria. Plant Signal. Behav. 2012, 7, 927–932. [Google Scholar] [CrossRef]
- Sun, F.; Suen, P.K.; Zhang, Y.; Liang, C.; Carrie, C.; Whelan, J.; Ward, J.L.; Hawkins, N.D.; Jiang, L.; Lim, B.L. A dual-targeted purple acid phosphatase in Arabidopsis thaliana moderates carbon metabolism and its overexpression leads to faster plant growth and higher seed yield. New Phytol. 2012, 194, 206–219. [Google Scholar] [CrossRef]
- Zhang, R.; Guan, X.; Law, Y.-S.; Sun, F.; Chen, S.; Wong, K.B.; Lim, B.L. AtPAP2 modulates the import of the small subunit of Rubisco into chloroplasts. Plant Signal. Behav. 2016, 11, e1239687. [Google Scholar] [CrossRef] [PubMed]
- Law, Y.S.; Ngan, L.; Yan, J.; Kwok, L.Y.; Sun, Y.; Cheng, S.; Schwenkert, S.; Lim, B.L. Multiple kinases can phosphorylate the N-terminal sequences of mitochondrial proteins in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 982. [Google Scholar] [CrossRef] [PubMed]
- Law, Y.S.; Zhang, R.; Guan, X.; Cheng, S.; Sun, F.; Duncan, O.; Murcha, M.W.; Whelan, J.; Lim, B.L. Phosphorylation and Dephosphorylation of the Presequence of Precursor MULTIPLE ORGANELLAR RNA EDITING FACTOR3 during Import into Mitochondria from Arabidopsis. Plant Physiology 2015, 169, 1344–1355. [Google Scholar] [CrossRef]
- Liang, C.; Zhang, Y.; Cheng, S.; Osorio, S.; Sun, Y.; Fernie, A.R.; Cheung, C.; Lim, B.L. Impacts of high ATP supply from chloroplasts and mitochondria on the leaf metabolism of Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 922. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, L.; Yung, K.-F.; Leung, D.Y.; Sun, F.; Lim, B.L. Over-expression of AtPAP2 in Camelina sativa leads to faster plant growth and higher seed yield. Biotechnol. Biofuels 2012, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Kurien, B.T.; Scofield, R.H. Western blotting. Methods 2006, 38, 283–293. [Google Scholar] [CrossRef]
- Schägger, H. Tricine–sds-page. Nat. Protoc. 2006, 1, 16. [Google Scholar] [CrossRef]
- Day, D.; Neuburger, M.; Douce, R. Biochemical characterization of chlorophyll-free mitochondria from pea leaves. Funct. Plant Biol. 1985, 12, 219–228. [Google Scholar] [CrossRef]
- Lister, R.; Carrie, C.; Duncan, O.; Ho, L.H.; Howell, K.A.; Murcha, M.W.; Whelan, J. Functional definition of outer membrane proteins involved in preprotein import into mitochondria. Plant Cell 2007, 19, 3739–3759. [Google Scholar] [CrossRef] [PubMed]
- Aronsson, H.; Jarvis, P. A simple method for isolating import-competent Arabidopsis chloroplasts. FEBS Lett. 2002, 529, 215–220. [Google Scholar] [CrossRef]
- Lamkemeyer, P.; Laxa, M.; Collin, V.; Li, W.; Finkemeier, I.; Schöttler, M.A.; Holtkamp, V.; Tognetti, V.B.; Issakidis-Bourguet, E.; Kandlbinder, A. Peroxiredoxin Q of Arabidopsis thaliana is attached to the thylakoids and functions in context of photosynthesis. Plant J. 2006, 45, 968–981. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Huang, S.; Lee, C.P.; Millar, A.H. Activity assay for plant mitochondrial enzymes. In Plant Mitochondria; Springer: Berlin/Heidelberg, Germany, 2015; pp. 139–149. [Google Scholar]
- Schaeffer, G.W.; Sharpe, F.T.; Sicher, R.C. Fructose 1, 6-bisphosphate aldolase activity in leaves of a rice mutant selected for enhanced lysine. Phytochemistry 1997, 46, 1335–1338. [Google Scholar] [CrossRef]
- Winter, K.; Foster, J.G.; Edwards, G.E.; Holtum, J.A. Intracellular localization of enzymes of carbon metabolism in Mesembryanthemum crystallinum exhibiting C3 photosynthetic characteristics or performing Crassulacean acid metabolism. Plant Physiol. 1982, 69, 300–307. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Robinson, K.M.; Janes, M.S.; Beckman, J.S. The selective detection of mitochondrial superoxide by live cell imaging. Nat. Protoc. 2008, 3, 941. [Google Scholar] [CrossRef]
- Cvetkovska, M.; Vanlerberghe, G.C. Alternative oxidase modulates leaf mitochondrial concentrations of superoxide and nitric oxide. New Phytol. 2012, 195, 32–39. [Google Scholar] [CrossRef]
- Meyer, E.H.; Tomaz, T.; Carroll, A.J.; Estavillo, G.; Delannoy, E.; Tanz, S.K.; Small, I.D.; Pogson, B.J.; Millar, A.H. Remodeled respiration in ndufs4 with low phosphorylation efficiency suppresses Arabidopsis germination and growth and alters control of metabolism at night. Plant Physiol. 2009, 151, 603–619. [Google Scholar] [CrossRef]
- Ford, S.R.; Leach, F.R. Bioluminescent assay of the adenylate energy charge. Methods Mol. Biol. 1998, 102, 69–81. [Google Scholar]
- Lu, Y.; Lam, H.; Pi, E.; Zhan, Q.; Tsai, S.; Wang, C.; Kwan, Y.; Ngai, S. Comparative metabolomics in Glycine max and Glycine soja under salt stress to reveal the phenotypes of their offspring. J. Agric. Food Chem. 2013, 61, 8711–8721. [Google Scholar] [CrossRef] [PubMed]
- Dhanoa, P.K.; Richardson, L.G.; Smith, M.D.; Gidda, S.K.; Henderson, M.P.; Andrews, D.W.; Mullen, R.T. Distinct pathways mediate the sorting of tail-anchored proteins to the plastid outer envelope. PLoS ONE 2010, 5, e10098. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M. Plant mitochondria and oxidative stress: Electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu. Rev. Plant Biol. 2001, 52, 561–591. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef]
- Nie, S.; Yue, H.; Zhou, J.; Xing, D. Mitochondrial-derived reactive oxygen species play a vital role in the salicylic acid signaling pathway in Arabidopsis thaliana. PLoS ONE 2015, 10, e0119853. [Google Scholar] [CrossRef]
- Khlyntseva, S.; Bazel, Y.R.; Vishnikin, A.; Andruch, V. Methods for the determination of adenosine triphosphate and other adenine nucleotides. J. Anal. Chem. 2009, 64, 657–673. [Google Scholar] [CrossRef]
- Liang, F.; Lindberg, P.; Lindblad, P. Engineering photoautotrophic carbon fixation for enhanced growth and productivity. Sustain. Energy Fuels 2018, 2, 2583–2600. [Google Scholar] [CrossRef]
- Raines, C.A. The Calvin cycle revisited. Photosynth. Res. 2003, 75, 1–10. [Google Scholar] [CrossRef]
- Martinoia, E.; Rentsch, D. Malate compartmentation-responses to a complex metabolism. Annu. Rev. Plant Biol. 1994, 45, 447–467. [Google Scholar] [CrossRef]
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol. 2011, 62, 79–104. [Google Scholar] [CrossRef]
- Gauthier, P.P.; Bligny, R.; Gout, E.; Mahe, A.; Nogues, S.; Hodges, M.; Tcherkez, G.G. In folio isotopic tracing demonstrates that nitrogen assimilation into glutamate is mostly independent from current CO2 assimilation in illuminated leaves of Brassica napus. New Phytol. 2010, 185, 988–999. [Google Scholar] [CrossRef] [PubMed]
- Popova, T.N.; De Carvalho, M.Â.A.P. Citrate and isocitrate in plant metabolism. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 1998, 1364, 307–325. [Google Scholar] [CrossRef]
- Sweetlove, L.J.; Beard, K.F.; Nunes-Nesi, A.; Fernie, A.R.; Ratcliffe, R.G. Not just a circle: Flux modes in the plant TCA cycle. Trends Plant Sci. 2010, 15, 462–470. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | INF | NRF | Seed Weight per Plant (mg) | Yield Change (%) |
|---|---|---|---|---|
| WT | 27.5 ± 0.9 a | 12.6 ± 0.9 a | 235.7 ± 31.9 | - |
| OE7 | 20.4 ± 1.0 d | 6.8 ± 1.1 c | 331.6 ± 33.1 ** | +40.7 |
| P2TOC1 | 25.8 ± 0.5 b | 11.4 ± 1.0 b | 292.7 ± 39.1 * | +24.2 |
| P2TOC2 | 25.6 ± 0.9 b | 11.6 ± 1.2 ab | 281.9 ± 28.6 * | +19.6 |
| P2TOM3 | 23.8 ± 1.3 c | 10.6 ± 1.3 b | 61.1 ± 15.4 ** | −74.1 |
| P2TOM4 | 24.5 ± 0.6 c | 10.6 ± 0.8 b | 62.4 ± 31.4 ** | −73.5 |
| Enzyme | Source | Enzyme Activity (nmol min−1 mg−1 Protein) | |||
|---|---|---|---|---|---|
| WT | OE7 | P2TOM4 | P2TOC1 | ||
| FBA | Chloroplast | 154 ± 0.6 | 180 ± 6.6 * | 134 ± 4.8 * | 158 ± 3.8 |
| NADP+-GAPDH | Chloroplast | 164 ± 5.6 | 176 ± 3.3 * | 148 ± 4.5 * | 183 ± 1.8 * |
| NADP+-MDH | Chloroplast | 474 ± 4.5 | 552 ± 24 * | 482 ± 37 | 584 ± 27 * |
| NAD+-MDH | Leaf | 3273 ± 51 | 3289 ± 26 | 3790 ± 58 * | 3511 ± 36 * |
| NAD+-MDH | Mitochondria | 17,244 ± 130 | 18,144 ± 350 * | 17,950 ± 80 * | 17,455 ± 490 |
| NAD+-ME | Mitochondria | 137 ± 1.0 | 151 ± 1.7 * | 162 ± 1.1* | 149 ± 1.6 * |
| Citrate synthase | Mitochondria | 5.4 ± 0.6 | 5.3 ± 0.9 | 5.5 ± 0.4 | 5.3 ± 0.0 |
| Aconitase | Mitochondria | 80.0 ± 4.6 | 79.1 ± 3.4 | 113.4 ± 1.7* | 99.4 ± 0.3 * |
| NAD+-ICDH | Mitochondria | 65.8 ± 1.7 | 84.8 ± 3.7 * | 86.4 ± 1.4* | 69.3 ± 0.9 * |
| NADP+-ICDH | Leaf | 59.8 ± 1.7 | 41.3 ± 0.5 * | 62.8 ± 1.7 | 61.9 ± 0.8 |
| 2OGDH | Mitochondria | 18.1 ± 0.3 | 20.9 ± 0.5 * | 19.5 ± 0.2* | 18.3 ± 0.1 * |
| SDH | Mitochondria | 63.1 ± 5.3 | 96.3 ± 8.0 * | 76.4 ± 2.3* | 83.0 ± 0.81 * |
| Fumarase | Mitochondria | 440 ± 11 | 404 ± 15 * | 441 ± 19 | 390 ± 12 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.; Zhang, R.; Yang, M.; Law, Y.-S.; Sun, F.; Hon, N.L.; Ngai, S.M.; Lim, B.L. A Balance between the Activities of Chloroplasts and Mitochondria Is Crucial for Optimal Plant Growth. Antioxidants 2021, 10, 935. https://doi.org/10.3390/antiox10060935
Xu Z, Zhang R, Yang M, Law Y-S, Sun F, Hon NL, Ngai SM, Lim BL. A Balance between the Activities of Chloroplasts and Mitochondria Is Crucial for Optimal Plant Growth. Antioxidants. 2021; 10(6):935. https://doi.org/10.3390/antiox10060935
Chicago/Turabian StyleXu, Zhou, Renshan Zhang, Meijing Yang, Yee-Song Law, Feng Sun, Ngai Lung Hon, Sai Ming Ngai, and Boon Leong Lim. 2021. "A Balance between the Activities of Chloroplasts and Mitochondria Is Crucial for Optimal Plant Growth" Antioxidants 10, no. 6: 935. https://doi.org/10.3390/antiox10060935
APA StyleXu, Z., Zhang, R., Yang, M., Law, Y.-S., Sun, F., Hon, N. L., Ngai, S. M., & Lim, B. L. (2021). A Balance between the Activities of Chloroplasts and Mitochondria Is Crucial for Optimal Plant Growth. Antioxidants, 10(6), 935. https://doi.org/10.3390/antiox10060935