
The Potential of High-Anthocyanin Purple Rice as a Functional Ingredient in Human Health


Abstract
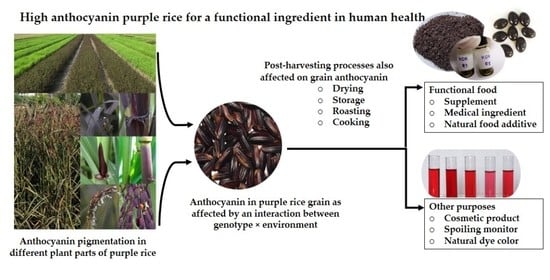
1. Introduction
2. Purple Rice’s Market and Trend
3. Anthocyanins in Different Plant Parts of Purple Rice
3.1. Variation in Grain Anthocyanin
3.2. Occurrence of Anthocyanin in Vegetative Plant Parts
4. Anthocyanin Biosynthesis Genes in Regulation of Anthocyanin in Rice
5. Genotype × Environment Interactions Controlling Anthocyanin in Rice
6. Stability of Grain Anthocyanin in Purple Rice during Post-Harvest Processing
7. Utilization of Anthocyanin in Purple Rice as Functional Compounds
8. Significant Roles of Anthocyanin from Purple Rice in Human Health
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wongwichai, T.; Teeyakasem, P.; Pruksakorn, D.; Kongtawelert, P.; Pothacharoen, P. Anthocyanins and metabolites from purple rice inhibit IL-1β-induced matrix metalloproteinases expression in human articular chondrocytes through the NF-κB and ERK/MAPK pathway. Biomed. Pharmacother. 2019, 112, 108610. [Google Scholar] [CrossRef] [PubMed]
- Fakhar, Z.; Faramarzi, B.; Pacifico, S.; Faramarzi, S. Anthocyanin derivatives as potent inhibitors of SARS-CoV-2 main protease: An in-silico perspective of therapeutic targets against COVID-19 pandemic. J. Biomol. Struct. Dyn. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, I.; Nawaz, A.; Sobhy, R.; Althwab, S.A.; Barakat, H. Polyacylated anthocyanins constructively network with catalytic dyad residues of 3CLpro of 2019-nCoV than monomeric anthocyanins: A structural-relationship activity study with 10 anthocyanins using in-silico approaches. J. Mol. Graph. 2020, 100, 107690. [Google Scholar] [CrossRef] [PubMed]
- Kongthitilerd, P.; Suantawee, T.; Cheng, H.; Thilavech, T.; Marnpae, M.; Adisakwattana, S. Anthocyanin-enriched riceberry rice extract inhibits cell proliferation and adipogenesis in 3T3-L1 preadipocytes by downregulating adipogenic transcription factors and their targeting genes. Nutrients 2020, 12, 2480. [Google Scholar] [CrossRef] [PubMed]
- Palungwachira, P.; Tancharoen, S.; Dararat, P.; Nararatwanchai, T. Anthocyanins isolated from Oryza Sativa L. protect dermal fibroblasts from hydrogen peroxide-induced cell death. J. Nat. Sci. Biol. Med. 2020, 11, 45. [Google Scholar]
- Zhou, J.; Zhu, Y.F.; Chen, X.Y.; Han, B.; Li, F.; Chen, J.Y.; Peng, X.L.; Luo, L.P.; Chen, W.; Yu, X.P. Black rice-derived anthocyanins inhibit HER-2-positive breast cancer epithelial-mesenchymal transition-mediated metastasis in vitro by suppressing FAK signaling. Int. J. Mol. Med. 2017, 40, 1649–1656. [Google Scholar] [CrossRef]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- FAO. 1995. Food and People. Staple Foods: What do People Eat? Available online: http://www.fao.org/3/u8480e/u8480e07.htm (accessed on 13 January 2020).
- Fairhurst, T.; Dobermann, A. Rice in the Global Food Supply. Better Crops Int. 2002, 16, 3–6. [Google Scholar]
- Prom-u-thai, C.; Rashid, A.; Ram, H.; Zou, C.; Guilherme, L.R.G.; Corguinha, A.P.B.; Guo, S.; Kaur, C.; Naeem, A.; Yamuangmorn, S.; et al. Simultaneous biofortification of rice with zinc, iodine, iron and selenium through foliar treatment of a micronutrient cocktail in five countries. Front. Plant Sci. 2020, 11, 589835. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Karbalaii, M.T.; Jaafar, H.Z.E.; Rahmat, A. Phytochemical constituents, antioxidant activity, and antiproliferative properties of black, red, and brown rice bran. Chem. Cent. J. 2018, 12, 17. [Google Scholar] [CrossRef]
- Veni, B.K. Nutrition profiles of different colored rice: A review. J. Pharmacogn. Phytochem. 2019, SP2, 303–305. [Google Scholar]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in solanaceous vegetables: A Review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Magaña Cerino, J.M.; Peniche, H.; Tiessen, A.; Díaz, C. Pigmented maize (Zea mays L.) contains anthocyanins with potential therapeutic action against oxidative stress—A Review. Pol. J. Food Nutr. Sci. 2020, 70, 85–99. [Google Scholar] [CrossRef]
- Pozos, G.; Ruiz, M.; Zamora-Natera, J.; Moya, C.; Ramírez, L.; Reynoso Silva, M.; Macías, R.; Garcia, P.; González-Cruz, R.; Pérez, E.; et al. Antioxidant capacity and antigenotoxic effect of Hibiscus sabdariffa L. extracts obtained with ultrasound-assisted extraction process. Appl. Sci. 2020, 10, 560. [Google Scholar] [CrossRef]
- Song, B.; Xu, H.; Chen, L.; Fan, X.; Jing, Z.; Chen, S.; Xu, Z. Study of the relationship between leaf color formation and anthocyanin metabolism among different purple Pakchoi lines. Molecules 2020, 25, 4809. [Google Scholar] [CrossRef]
- Norkaew, O.; Thitisut, P.; Mahatheeranont, S.; Pawin, B.; Sookwong, P.; Yodpitak, S.; Lungkaphin, A. Effect of wall materials on some physicochemical properties and release characteristics of encapsulated black rice anthocyanin microcapsules. Food Chem. 2019, 294, 493–502. [Google Scholar] [CrossRef]
- Kanha, N.; Regenstein, J.M.; Surawang, S.; Pitchakarn, P.; Laokuldilok, T. Properties and kinetics of the in vitro release of anthocyanin-rich microcapsules produced through spray and freeze-drying complex coacervated double emulsions. Food Chem. 2021, 340, 127950. [Google Scholar] [CrossRef]
- Anuyahong, T.; Chusak, C.; Adisakwattana, S. Incorporation of anthocyanin-rich riceberry rice in yogurts: Effect on physicochemical properties, antioxidant activity and in vitro gastrointestinal digestion. LWT 2020, 129, 109571. [Google Scholar] [CrossRef]
- Papillo, V.A.; Locatelli, M.; Travaglia, F.; Bordiga, M.; Garino, C.; Arlorio, M.; Coïsson, J.D. Spray-dried polyphenolic extract from Italian black rice (Oryza sativa L., var. Artemide) as new ingredient for bakery products. Food Chem. 2018, 269, 603–609. [Google Scholar] [CrossRef]
- Yi, J.; Qiu, M.; Liu, N.; Tian, L.; Zhu, X.; Decker, E.A.; McClements, D.J. Inhibition of lipid and protein oxidation in whey-protein-stabilized emulsions using a natural antioxidant: Black rice anthocyanins. J. Agric. Food Chem. 2020, 68, 10149–10156. [Google Scholar] [CrossRef]
- Okafor, S.; Obonga, W.; Ezeokonkwo, M. Assessment of the health implications of synthetic and natural food Colourants—A Critical Review. UK J. Pharm. Biosci. 2016, 4, 1. [Google Scholar] [CrossRef]
- Chaudhary, R. Speciality rices of the world: Effect of WTO and IPR on its production trend and marketing. J. Food Agric. Environ. 2003, 1, 34–41. [Google Scholar]
- Pal, S.; Bagchi, T.B.; Dhali, K.; Kar, A.; Sanghamitra, P.; Sarkar, S.; Samaddar, M.; Majumder, J. Evaluation of sensory, physicochemical properties and consumer preference of black rice and their products. J. Food Sci. Technol. 2019, 56, 1484–1494. [Google Scholar] [CrossRef]
- Sholikhah, U.; Parjanto, P.; Handoyo, T.; Yunus, A. Genetic diversity of black and aromatic rice cultivar (Oryza sativa L.) from various regions in Indonesia using random amplified polymorphic DNA markers (RAPD). Int. J. Adv. Sci. Eng. Inf. Technol. 2019, 9, 1046–1051. [Google Scholar] [CrossRef]
- Rahmawati, A.; Yuniastuti, E.; Nandariyah. Increased anthocyanin content in seven furrows of Cempo Ireng black rice with mutation induction. IOP Conf. Ser. Earth Environ. Sci. 2020, 466, 012010. [Google Scholar] [CrossRef]
- Dittgen, C.L.; Hoffmann, J.F.; Chaves, F.C.; Rombaldi, C.V.; Filho, J.M.C.; Vanier, N.L. Discrimination of genotype and geographical origin of black rice grown in Brazil by LC-MS analysis of phenolics. Food Chem. 2019, 288, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Appa Rao, S.; Schiller, J.M.; Bounphanousay, C.; Inthapanya, P.; Jackson, M.T. The colored pericarp (black) rice of Laos. In Rice in Laos; Schiller, J.M., Chanphengxay, M.B., Linquist, B., Appa Rao, S., Eds.; International Rice Research Institute: Los Baños, Philippines, 2006; pp. 175–186. [Google Scholar]
- Deng, G.F.; Xu, X.R.; Zhang, Y.; Li, D.; Gan, R.Y.; Li, H.B. Phenolic compounds and bioactivities of pigmented rice. Crit. Rev. Food Sci. Nutr. 2013, 53, 296–306. [Google Scholar] [CrossRef]
- Callcott, E.T.; Santhakumar, A.B.; Strappe, P.; Luo, J.; Blanchard, C.L. Polyphenols from Australian-grown pigmented red and purple rice inhibit adipocyte differentiation. J. Cereal Sci. 2018, 81, 140–146. [Google Scholar] [CrossRef]
- Melini, V.; Panfili, G.; Fratianni, A.; Acquistucci, R. Bioactive compounds in rice on Italian market: Pigmented varieties as a source of carotenoids, total phenolic compounds and anthocyanins, before and after cooking. Food Chem. 2019, 277, 119–127. [Google Scholar] [CrossRef]
- Tumanian, N.; Mukhina, Z.; Esaulova, L.; Papulova, E.; Savenko, E.; Garkusha, S. Colored rice varieties of Russian breeding in terms of grain quality for development of functional rice varieties. E3S Web Conf. 2020, 224, 04021. [Google Scholar] [CrossRef]
- Zelenskaya, O.; Zelensky, G.; Ostapenko, N.; Tumanyan, I. Genetic resources of rice (Oryza sativa L.) with colored pericarp. Vavilov J. Genet. Breed. 2018, 22, 296–303. [Google Scholar] [CrossRef]
- OAE. Rice Export. Office of Agricultural Economics, Thai Ministry of Agriculture and Cooperatives. 2020. Available online: http://impexp.oae.go.th/service/export.php (accessed on 13 January 2020). (In Thai).
- DFT. Notification of Ministry of Commerce. Subject: Standards for Thai Colour Rice B.E. 2562. Department of Foreign Trade. 2019. Available online: https://www.dft.go.th/th-th/Detail-Law/ArticleId/12882/-2562-1-2-3-4-5-6-7-8) (accessed on 8 January 2020).
- Rerkasem, B.; Jumrus, S.; Yimyam, N. Variation of grain nutritional quality among Thai purple rice genotypes grown at two different altitudes. ScienceAsia 2015, 41, 377. [Google Scholar] [CrossRef]
- Jaksomsak, P.; Rerkasem, B.; Prom-u-thai, C. Variation in nutritional quality of pigmented rice varieties under different water regimes. Plant. Prod. Sci. 2020, 24, 244–255. [Google Scholar] [CrossRef]
- Yamuangmorn, S.; Dell, B.; Rerkasem, B.; Prom-u-thai, C. Applying nitrogen fertilizer increased anthocyanin in vegetative shoots but not in grain of purple rice genotypes. J. Sci. Food Agric. 2018, 98, 4527–4532. [Google Scholar] [CrossRef] [PubMed]
- Juliano, B.O.; Tuaño, A.P.P. Gross structure and composition of the rice grain. In Rice Chemistry and Technology, 4th ed.; Bao, J., Ed.; AACC International Press: Washington, DC, USA, 2019; pp. 31–53. [Google Scholar]
- Shao, Y.; Xu, F.; Sun, X.; Bao, J.; Beta, T. Identification and quantification of phenolic acids and anthocyanins as antioxidants in bran, embryo and endosperm of white, red and black rice kernels (Oryza sativa L.). J. Cereal Sci. 2014, 59, 211–218. [Google Scholar] [CrossRef]
- Fang, X.; Mao, Y.; Chen, X. Engineering purple rice for human health. Sci. China-Life Sci. 2018, 61, 365–367. [Google Scholar] [CrossRef]
- Zhu, Q.; Yu, S.; Zeng, D.; Liu, H.; Wang, H.; Yang, Z.; Xie, X.; Shen, R.; Tan, J.; Li, H.; et al. Development of “purple endosperm rice” by Engineering anthocyanin biosynthesis in the endosperm with a high-efficiency transgene stacking system. Mol. Plant. 2017, 10, 918–929. [Google Scholar] [CrossRef]
- Hu, C.; Zawistowski, J.; Ling, W.; Kitts, D.D. Black rice (Oryza sativa L. indica) pigmented fraction suppresses both reactive oxygen species and nitric oxide in chemical and biological model systems. J. Agric. Food Chem. 2003, 51, 5271–5277. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Zaima, N.; Moriyama, T.; Kawamura, Y. Different localization patterns of anthocyanin species in the pericarp of black rice revealed by imaging mass spectrometry. PLoS ONE 2012, 7, e31285. [Google Scholar] [CrossRef]
- Abdel-Aal el, S.M.; Young, J.C.; Rabalski, I. Anthocyanin composition in black, blue, pink, purple, and red cereal grains. J. Agric. Food Chem. 2006, 54, 4696–4704. [Google Scholar] [CrossRef]
- Maisuthisakul, P.; Changchub, L. Effect of extraction on phenolic antioxidant of different Thai rice (Oryza sativa L.) genotypes. Int. J. Food Prop. 2014, 17, 855–865. [Google Scholar] [CrossRef]
- Chen, M.H.; McClung, A.; Bergman, C. Phenolic content, anthocyanins and antiradical capacity of diverse purple bran rice genotypes as compared to other bran colors. J. Cereal Sci. 2017, 77, 110–119. [Google Scholar] [CrossRef]
- Jansom, V.; Jansom, C.; Lerdvuthisopon, N. The relationship between color values in rice to phenolic acids, flavonoids, and antioxidants. J. Med. Assoc. Thai. 2020, 103, 80. [Google Scholar]
- Thuy, P.; Trung, N.; Huong, D. Agronomic characteristics, anthocyanin content, and antioxidant activity of anthocynins extracted from the seeds of black rice accessions. Vietnam. J. Agric. Sci. 2019, 1, 208–219. [Google Scholar]
- Li, T.; Wang, L.; Chen, Z.; Zhang, X.; Zhu, Z. Functional properties and structural changes of rice proteins with anthocyanins complexation. Food Chem. 2020, 331, 127336. [Google Scholar] [CrossRef] [PubMed]
- Jamjod, S.; Yimyam, N.; Lordkaew, S.; Prom-u-thai, C.T.; Rerkasem, B. Characterization of on-farm rice germplasm in an area of the crop’s center of diversity. CMU J. Nat. Sci. 2017, 16, 85–98. [Google Scholar] [CrossRef][Green Version]
- Kristamtini; Taryono, T.; Basunanda, P.; Murti, R.; Supriyanta; Sutarno, S.; Sutarno, N. Morphological of genetic relationships among black rice landraces from yogyakarta and surrounding areas. ARPN J. Agric. Biol. Sci. 2012, 7, 982–989. [Google Scholar]
- Mbanjo, E.G.N.; Jones, H.; Caguiat, X.G.I.; Carandang, S.; Ignacio, J.C.; Ferrer, M.C.; Boyd, L.A.; Kretzschmar, T. Exploring the genetic diversity within traditional Philippine pigmented rice. Rice 2019, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Phonsakhan, W.; Kong-Ngern, K. A comparative proteomic study of white and black glutinous rice leaves. Electron. J. Biotechnol. 2015, 18, 29–34. [Google Scholar] [CrossRef]
- Xiongsiyee, V.; Rerkasem, B.; Veeradittakit, J.; Saenchai, C.; Lordkaew, S.; Prom-u-thai, C.T. Variation in grain quality of upland rice from Luang Prabang province, Lao PDR. Rice Sci. 2018, 25, 94–102. [Google Scholar] [CrossRef]
- Bhuvaneswari, S.; Gopala Krishnan, S.; Bollinedi, H.; Saha, S.; Ellur, R.K.; Vinod, K.K.; Singh, I.M.; Prakash, N.; Bhowmick, P.K.; Nagarajan, M.; et al. Genetic architecture and anthocyanin profiling of aromatic rice from Manipur reveals divergence of Chakhao landraces. Front. Genet. 2020, 11, 1166. [Google Scholar] [CrossRef] [PubMed]
- Pramai, P.; Jiamyangyuen, S. Chemometric classification of pigmented rice varietiesbased on antioxidative properties in relation to color. Songklanakarin J. Sci. Technol. 2016, 38, 463–472. [Google Scholar]
- Sompong, R.; Siebenhandl-Ehn, S.; Linsberger-Martin, G.; Berghofer, E. Physicochemical and antioxidative properties of red and black rice varieties from Thailand, China and Sri Lanka. Food Chem. 2011, 124, 132–140. [Google Scholar] [CrossRef]
- Sangpimpa, W.; Utama-ang, N. Chemical properties of three selected thai rice and texture profiling of cooked Kum Doi Saket rice. Food Appl. Biosci. J. 2018, 6, 117–133. [Google Scholar]
- Juemanee, A.; Meenune, M.; Kijroongrojana, K. Relationships of sensory profile with instrumental measurement and consumer acceptance of Thai unpolished pigmented rice. Int. Food Res. J. 2018, 25, 2112–2120. [Google Scholar]
- Chan-in, P.; Jamjod, S.; Yimyam, N.; Rerkasem, B.; Pusadee, T. Grain quality and allelic variation of the badh2 gene in thai fragrant rice landraces. Agronomy 2020, 10, 779. [Google Scholar] [CrossRef]
- Yamuangmorn, S.; Dell, B.; Prom-u-thai, C. Effects of cooking on anthocyanin concentration and bioactive antioxidant capacity in glutinous and non-glutinous purple rice. Rice Sci. 2018, 25, 270–278. [Google Scholar] [CrossRef]
- Ratseewo, J.; Warren, F.J.; Siriamornpun, S. The influence of starch structure and anthocyanin content on the digestibility of Thai pigmented rice. Food Chem. 2019, 298, 124949. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Landoni, M.; Tomay, F.; Calvenzani, V.; Simonelli, C.; Cormegna, M. Proximate composition, polyphenol content and anti-inflammatory properties of white and pigmented italian rice varieties. Univers. J. Agric. Res. 2017, 5, 312–321. [Google Scholar] [CrossRef]
- Rahaju, R.; Sumardi, D.; Pancoro, A. Total flavonoids and anthocyanins content of pigmented rice. Drug Invent. Today 2019, 12, 363–373. [Google Scholar]
- Murdifin, M.; Pakki, E.; Rahim, A.; Syaiful, S.A.; Ismail; Evary, Y.; Bahar, M.A. Physicochemical properties of Indonesian pigmented rice (Oryza sativa linn.) varieties from south Sulawesi. Asian J. Plant Sci. 2015, 14, 59–65. [Google Scholar] [CrossRef]
- Embate, M.; Calayugan, M.I.; Gentallan, R., Jr.; Hernandez, J.; Sta Cruz, P.; Borromeo, T. Genetic diversity of selected pigmented traditional rice (Oryza sativa L.) varieties from Mindanao, Philippines using agromorphological traits and simple sequence repeats markers. J. Crop. Sci. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Xavier; Caguiat, X.; Rañeses, M.A.; Chico, M.; Arocena, E.; Ticman, H. Molecular analysis of glutinous pigmented rice germplasm through the aid of highly polymorphic markers. Int. J. Res. Agric. Food Sci. 2019, 5, 1–16. [Google Scholar]
- Ahmad, F.; Hanafi, M.; Hakim, M.; Rafii, M.; Wasiu Arolu, I.; Abdullah, S. Genetic divergence and heritability of 42 coloured upland rice genotypes (Oryza sativa) as revealed by microsatellites marker and agro-morphological traits. PLoS ONE 2015, 10, e0138246. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Cros, G.; Yokota, T.; Crozier, A. Phytochemical profiles of black, red, brown, and white rice from the Camargue region of France. J. Agric. Food Chem. 2013, 61, 7976–7986. [Google Scholar] [CrossRef]
- Jiamyangyuen, S.; Nuengchamnong, N.; Ngamdee, P. Changes of anthocyanins in black rice flours prepared by cooking and pregelatinization. CMU J. Nat. Sci. 2019, 18, 535–552. [Google Scholar] [CrossRef]
- Wu, N.N.; Li, H.H.; Tan, B.; Zhang, M.; Xiao, Z.G.; Tian, X.H.; Zhai, X.T.; Liu, M.; Liu, Y.X.; Wang, L.P.; et al. Free and bound phenolic profiles of the bran from different rice varieties and their antioxidant activity and inhibitory effects on ɑ-amylose and ɑ-glucosidase. J. Cereal Sci. 2018, 82, 206–212. [Google Scholar] [CrossRef]
- Yamuangmorn, S.; Dell, B.; Du, X.; Ren, Y.; Prom-u-thai, C. Simultaneous quantification of anthocyanins and phenolic acids in pigmented rice (Oryza sativa) using UPLC-PDA/ESI-Q-TOF. Int. J. Agric. Biol. 2019, 21, 590–596. [Google Scholar]
- Wongsa, P.; Chaiwarith, J.; Voranitikul, J.; Chaiwongkhajorn, J.; Rattanapanone, N.; Lanberg, R. Identification of phenolic compounds in colored rice and their inhibitory potential against α-amylase. Chiang Mai J. Sci. 2019, 46, 672–682. [Google Scholar]
- Chen, P.N.; Chu, S.C.; Chiou, H.L.; Chiang, C.L.; Yang, S.F.; Hsieh, Y.S. Cyanidin 3-glucoside and peonidin 3-glucoside inhibit tumor cell growth and induce apoptosis in vitro and suppress tumor growth in Vivo. Nutr. Cancer 2005, 53, 232–243. [Google Scholar] [CrossRef]
- Yawadio, R.; Tanimori, S.; Morita, N. Identification of phenolic compounds isolated from pigmented rices and their aldose reductase inhibitory activities. Food Chem. 2007, 101, 1616–1625. [Google Scholar] [CrossRef]
- Peanparkdee, M.; Patrawart, J.; Iwamoto, S. Effect of extraction conditions on phenolic content, anthocyanin content and antioxidant activity of bran extracts from Thai rice cultivars. J. Cereal Sci. 2019, 86, 86–91. [Google Scholar] [CrossRef]
- Nakagawa, K.; Yooin, W.; Saenjum, C. EPR and HPLC investigation of pigments in Thai purple rice. J. Oleo Sci. 2018, 67, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Hu, Z.; Yu, Y.; Mou, R.; Zhu, Z.; Beta, T. Phenolic acids, anthocyanins, proanthocyanidins, antioxidant activity, minerals and their correlations in non-pigmented, red, and black rice. Food Chem. 2018, 239, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Lin, Z.; Lin, G.; Chen, S. The anti-photooxidation of anthocyanins-rich leaves of a purple rice cultivar. Sci. China C Life Sci. 2006, 49, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Senakun, C.; Chunta, S.; Somboonwattanakul, I.; Yodsiri, S.; Kurukodt, J.; Senakun, A. Diversity, utilization and cultural significance of purple rice in northeastern Thailand. Int. J. Agric. Res. 2018, 14, 1893–1904. [Google Scholar]
- Chin, H.S.; Wu, Y.P.; Hour, A.L.; Hong, C.Y.; Lin, Y.R. Genetic and evolutionary analysis of purple leaf sheath in rice. Rice 2016, 9, 8. [Google Scholar] [CrossRef]
- Xu, Z.; Mahmood, K.; Rothstein, S.J. ROS induces anthocyanin production via late biosynthetic genes and anthocyanin deficiency confers the hypersensitivity to ROS-generating stresses in Arabidopsis. Plant Cell Physiol. 2017, 58, 1364–1377. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Dai, G.; Zhou, W.; Liang, H.; Huang, J.; Qing, D.; Chen, W.; Wu, H.; Yang, X.; Li, D.; et al. Mapping and identifying a candidate gene Plr4, a recessive gene regulating purple leaf in rice, by using bulked segregant and transcriptome analysis with next-generation sequencing. Int. J. Mol. Sci. 2019, 20, 4335. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Jalil, S.; Cao, H.; Chare, Y.; Hamdaoui, M.; Ziyan, C.; Shi, C.; Jin, X. The purple leaf (pl6) mutation regulates leaf color by altering the anthocyanin and chlorophyll contents in rice. Plants 2020, 9, 1477. [Google Scholar] [CrossRef] [PubMed]
- Oshima, M.; Taniguchi, Y.; Akasaka, M.; Abe, K.; Ichikawa, H.; Tabei, Y.; Tanaka, J. Development of a visible marker trait based on leaf sheath-specific anthocyanin pigmentation applicable to various genotypes in rice. Breed. Sci. 2019, 69, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Cha-um, S. Regulation of anthocyanin accumulation in rice (Oryza sativa L. subsp. indica) using MgSO4 spraying and low temperature. Arch. Agron. Soil Sci. 2018, 64, 1663–1677. [Google Scholar] [CrossRef]
- Jha, P.; Das, A.J.; Deka, S.C. Optimization of ultrasound and microwave assisted extractions of polyphenols from black rice (Oryza sativa cv. Poireton) husk. J. Food Sci. Technol. Mysore 2017, 54, 3847–3858. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, Z.; Chen, C.; Wu, W.; Ren, N.; Jiang, C.; Yu, J.; Zhao, Y.; Zheng, X.; Yang, Q.; et al. The C–S–A gene system regulates hull pigmentation and reveals evolution of anthocyanin biosynthesis pathway in rice. J. Exp. Bot. 2018, 69, 1485–1498. [Google Scholar] [CrossRef]
- Punvittayagul, C.; Chariyakornkul, A.; Sankam, P.; Wongpoomchai, R. Inhibitory effect of Thai purple rice husk extract on chemically induced carcinogenesis in rats. Molecules 2021, 26, 360. [Google Scholar] [CrossRef] [PubMed]
- Chariyakornkul, A.; Punvittayagul, C.; Taya, S.; Wongpoomchai, R. Inhibitory effect of purple rice husk extract on AFB1-induced micronucleus formation in rat liver through modulation of xenobiotic metabolizing enzymes. BMC Complement. Altern. Med. 2019, 19, 237. [Google Scholar] [CrossRef]
- Kim, B.G.; Kim, J.H.; Min, S.Y.; Shin, K.H.; Kim, J.H.; Kim, H.Y.; Ryu, S.N.; Ahn, j.H. Anthocyanin content in rice is related to expression levels of anthocyanin biosynthetic genes. J. Plant Biol. 2007, 50, 156–160. [Google Scholar] [CrossRef]
- Shih, C.H.; Chu, H.; Tang, L.K.; Sakamoto, W.; Maekawa, M.; Chu, I.K.; Wang, M.; Lo, C. Functional characterization of key structural genes in rice flavonoid biosynthesis. Planta 2008, 228, 1043–1054. [Google Scholar] [CrossRef]
- Chen, X.; Itani, T.; Wu, X.; Chikawa, Y.; Irifune, K. Physiological factors affecting transcription of genes involved in the flavonoid biosynthetic pathway in different rice varieties. Plant Signal. Behav. 2013, 8, e27555. [Google Scholar] [CrossRef][Green Version]
- Nayeem, S.; Venkidasamy, B.; Sundararajan, S.; Kuppuraj, S.P.; Ramalingam, S. Differential expression of flavonoid biosynthesis genes and biochemical composition in different tissues of pigmented and non-pigmented rice. J. Food Sci. Technol. 2020, 58, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Yamaguchi, T.; Omoteno, M.; Takarada, T.; Fujita, K.; Murata, K.; Ebitani, T. Genetic dissection of black grain rice by the development of a near isogenic line. Breed Sci. 2014, 64, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Rachasima, L.N.; Sukkasem, R.; Pongjaroenkit, R.; Sangtong, V.; Chowpongpang, S.; Sakulsingharoj, C. Expression analysis and nucleotide variation of OsC1 gene associated with anthocyanin pigmentation in Thai rice cultivars. Genom. Genet. 2017, 10, 1–8. [Google Scholar]
- Kim, D.H.; Park, S.; Lee, J.Y.; Ha, S.H.; Lee, J.G.; Lim, S.H. A rice B-Box protein, OsBBX14, finely regulates anthocyanin biosynthesis in rice. Int. J. Mol. Sci. 2018, 19, 2190. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhou, T.; Han, Z.; Tan, C.; Xing, Y. Dominant complementary interaction between OsC1 and two tightly linked genes, Rb1 and Rb2, controls the purple leaf sheath in rice. Theor. Appl. Genet. 2020, 133, 2555–2566. [Google Scholar] [CrossRef] [PubMed]
- Tsago, Y.; Wang, Z.; Liu, J.; Sunusi, M.; Eshag, J.; Akhter, D.; Shi, C.; Jin, X. Morphological characteristics and gene mapping of purple apiculus formation in rice. Plant Mol. Biol. Rep. 2019, 37, 277–290. [Google Scholar] [CrossRef]
- Akhter, D.; Qin, R.; Nath, U.K.; Eshag, J.; Jin, X.; Shi, C. A rice gene, OsPL, encoding a MYB family transcription factor confers anthocyanin synthesis, heat stress response and hormonal signaling. Gene 2019, 699, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wu, H.; Zhu, H.; Huang, C.; Liu, C.; Chang, Y.; Kong, Z.; Zhou, Z.; Wang, G.; Lin, Y.; et al. Determining factors, regulation system, and domestication of anthocyanin biosynthesis in rice leaves. New Phytol. 2019, 223, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; Lee, H.J.; Kim, J.H.; Kim, D.H.; Kim, H.K.; Cho, Y.G.; Bae, S.; Kang, K.K. CRISPR/Cas9-targeted mutagenesis of F3′H, DFR and LDOX, genes related to anthocyanin biosynthesis in black rice (Oryza sativa L.). Plant Biotechnol. Rep. 2019, 13, 521–531. [Google Scholar] [CrossRef]
- Nuraida, W.O.; Pradipta, P.; Suliartini, N.W.S.; Wijayanto, T.; Muhidin; Sadimantara, G.R. Production and quality of upland red rice under the shade stress. Int. J. Sci. Technol. Res. 2020, 9, 5016–5019. [Google Scholar]
- Chen, C.C.; Huang, M.Y.; Lin, K.H.; Wong, S.L.; Huang, W.D.; Yang, C.M. Effects of light quality on the growth, development and metabolism of rice seedlings (Oryza sativa L.). Res. J. Biotechnol. 2014, 9, 15–24. [Google Scholar]
- Bae, H.K.; Seo, J.H.; Hwang, J.D.; Kim, S.Y. Variation in grain quality and yield of black-colored rice affected by the transplanting time and temperature during ripening stage. Korean J. Crop Sci./Hanguk Jakmul Hakhoe Chi 2019, 64, 63–69. [Google Scholar]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Phisalaphong, M.; Singh, H.P.; Cha-um, S. Promoting water deficit tolerance and anthocyanin fortification in pigmented rice cultivar (Oryza sativa L. subsp. indica) using arbuscular mycorrhizal fungi inoculation. Physiol. Mol. Biol. Plants 2019, 25, 821–835. [Google Scholar] [CrossRef]
- Bennett, C.; Sookwong, P.; Moolkam, S.; Mahatheeranont, S. Effect of plant nutrients on anthocyanin content and yield component of black glutinous rice plants. Int. J. Agric. Biosyst. Eng. 2020, 14, 65–68. [Google Scholar]
- Samart, S.; Chutipaijit, S. Growth of pigmented rice (Oryza sativa L. cv. Riceberry) exposed to ZnO nanoparticles. Mater. Today 2019, 17, 1987–1994. [Google Scholar] [CrossRef]
- Jan, R.; Khan, M.A.; Asaf, S.; Lee, I.J.; Kim, K.M. Modulation of sugar and nitrogen in callus induction media alter PAL pathway, SA and biomass accumulation in rice callus. Plant Cell Tissue Organ Cult. 2020, 143, 517–530. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N.; O’Donnell, C.; Brijesh kumar, T. Effect of thermal processing on anthocyanin stability in foods: Mechanisms and kinetics of degradation. Trends Food Sci. Technol. 2010, 21, 3–11. [Google Scholar] [CrossRef]
- Lang, G.H.; Lindemann, I.D.S.; Ferreira, C.D.; Hoffmann, J.F.; Vanier, N.L.; de Oliveira, M. Effects of drying temperature and long-term storage conditions on black rice phenolic compounds. Food Chem. 2019, 287, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Ratseewo, J.; Meeso, N.; Siriamornpun, S. Changes in amino acids and bioactive compounds of pigmented rice as affected by far-infrared radiation and hot air drying. Food Chem. 2020, 306, 125644. [Google Scholar] [CrossRef] [PubMed]
- Yamuangmorn, S.; Jumrus, S.; Jamjod, S.; Yimyam, N.; Prom-u-thai, C. Stabilizing grain yield and nutrition quality in purple rice varieties by management of planting elevation and storage conditions. Agronomy 2021, 11, 83. [Google Scholar] [CrossRef]
- Ziegler, V.; Ferreira, C.D.; Hoffmann, J.F.; Chaves, F.C.; Vanier, N.L.; de Oliveira, M.; Elias, M.C. Cooking quality properties and free and bound phenolics content of brown, black, and red rice grains stored at different temperatures for six months. Food Chem. 2018, 242, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Norkaew, O.; Boontakham, P.; Dumri, K.; Noenplab, A.N.L.; Sookwong, P.; Mahatheeranont, S. Effect of post-harvest treatment on bioactive phytochemicals of Thai black rice. Food Chem. 2017, 217, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Tananuwong, K.; Tangsrianugul, N. Effects of storage conditions and cooking on colour and antioxidant activities of organic pigmented rice. Int. J. Food Sci. Technol. 2013, 48, 67–73. [Google Scholar] [CrossRef]
- Nayeem, S.; Sundararajan, S.; Ashok, A.K.; Abusaliya, A.; Ramalingam, S. Effects of cooking on phytochemical and antioxidant properties of pigmented and non-pigmented rare Indian rice landraces. Biocatal. Agric. Biotechnol. 2021, 32, 101928. [Google Scholar] [CrossRef]
- Thuengtung, S.; Ogawa, Y. Comparative study of conventional steam cooking and microwave cooking on cooked pigmented rice texture and their phenolic antioxidant. Food Sci. Nutr. 2020, 8, 965–972. [Google Scholar] [CrossRef]
- Fracassetti, D.; Pozzoli, C.; Vitalini, S.; Tirelli, A.; Iriti, M. Impact of cooking on bioactive compounds and antioxidant activity of pigmented rice cultivars. Foods 2020, 9, 967. [Google Scholar] [CrossRef] [PubMed]
- Catena, S.; Turrini, F.; Boggia, R.; Borriello, M.; Gardella, M.; Zunin, P. Effects of different cooking conditions on the anthocyanin content of a black rice (Oryza sativa L. ‘Violet Nori’). Eur. Food Res. Technol. 2019, 245, 2303–2310. [Google Scholar] [CrossRef]
- Arora, S.; Virdi, I.K.; Sharanagat, V.S.; Kheto, A.; Dhua, S.; Suhag, R.; Kumar, R.; Kumar, Y.; Patel, A. Roasting of black rice (Oryza sativa L.): Change in physico-functional, thermo-pasting, antioxidant and anthocyanin content. J. Food Meas. Charact. 2021. [Google Scholar] [CrossRef]
- Yamuangmorn, S.; Sreethong, T.; Saenchai, C.; Rerkasem, B.; Prom-u-thai, C. Effects of roasting conditions on anthocyanin, total phenolic content, and antioxidant capacity in pigmented and non-pigmented rice varieties. Int. Food Res. J. 2021, 28, 73–82. [Google Scholar]
- Qiu, T.; Sun, Y.; Wang, X.; Zheng, L.; Zhang, H.; Jiang, L.; Zhu, X.; Xiong, H. Drum drying-and extrusion-black rice anthocyanins exert anti-inflammatory effects via suppression of the NF-κB /MAPKs signaling pathways in LPS-induced RAW 264.7 cells. Food Biosci. 2021, 41, 100841. [Google Scholar] [CrossRef]
- Jung, Y.K.; Joo, K.S.; Rho, S.J.; Kim, Y.R. pH-dependent antioxidant stability of black rice anthocyanin complexed with cycloamylose. LWT 2020, 129, 109474. [Google Scholar] [CrossRef]
- Peanparkdee, M.; Patrawart, J.; Iwamoto, S. Physicochemical stability and in vitro bioaccessibility of phenolic compounds and anthocyanins from Thai rice bran extracts. Food Chem. 2020, 329, 127157. [Google Scholar] [CrossRef] [PubMed]
- Tobgay, U.; Boonyanuphong, P.; Meunprasertdee, P. Comparison of hot water and methanol extraction combined with ultrasonic pretreatment on antioxidant properties of two pigmented rice cultivars. Food Res. 2019, 4, 547–556. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, W. Microencapsulation of anthocyanins through two-step emulsification and release characteristics during in vitro digestion. Food Chem. 2019, 278, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Zhang, Z.; Jing, P. Black rice anthocyanins embedded in self-assembled chitosan/chondroitin sulfate nanoparticles enhance apoptosis in HCT-116 cells. Food Chem. 2019, 301, 125280. [Google Scholar] [CrossRef] [PubMed]
- Chusak, C.; Pasukamonset, P.; Chantarasinlapin, P.; Adisakwattana, S. Postprandial glycemia, insulinemia, and antioxidant status in healthy subjects after ingestion of bread made from anthocyanin-rich Riceberry rice. Nutrients 2020, 12, 782. [Google Scholar] [CrossRef]
- Thiranusornkij, L.; Thamnarathip, P.; Chandrachai, A.; Kuakpetoon, D.; Adisakwattana, S. Comparative studies on physicochemical properties, starch hydrolysis, predicted glycemic index of Hom Mali rice and Riceberry rice flour and their applications in bread. Food Chem. 2019, 283, 224–231. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, Y.; Zhou, W. Bread fortified with anthocyanin-rich extract from black rice as nutraceutical sources: Its quality attributes and in vitro digestibility. Food Chem. 2016, 196, 910–916. [Google Scholar] [CrossRef]
- Zhao, L.; Pan, F.; Mehmood, A.; Zhang, Y.; Hao, S.; Rehman, A.U.; Li, J.; Wang, C.; Wang, Y. Protective effect and mechanism of action of xanthan gum on the color stability of black rice anthocyanins in model beverage systems. Int. J. Biol. Macromol. 2020, 164, 3800–3807. [Google Scholar] [CrossRef] [PubMed]
- Le, L.; Nguyen, T. Optimization of germination process of “Cam” brown rice by response surface methodology and evaluation of germinated rice quality. Food Res. 2019, 4, 459–467. [Google Scholar]
- Lainumngen, N.; Saengprakai, J.; Tanjor, S.; Phanpho, W.; Phodsoongnoen, A. Development of high anthocyanin crispy rice bar. Suan Sunandha Sci. Technol. J. 2020, 7, 34–42. [Google Scholar]
- Thao, N.; Niwat, C. Effect of germinated colored rice on bioactive compounds and quality of fresh germinated colored rice noodle. KMUTNB Int. J. Appl. Sci. Technol. 2017, 11, 27–37. [Google Scholar] [CrossRef]
- Sethi, S.; Nanda, S.K.; Bala, M. Quality assessment of pasta enriched with anthocyanin-rich black rice bran. J. Food Process Preserv. 2020, 44, e14952. [Google Scholar] [CrossRef]
- Tamprasit, K.; Weerapreeyakul, N.; Sutthanut, K.; Thukhammee, W.; Wattanathorn, J. Harvest age effect on phytochemical content of white and black glutinous rice cultivars. Molecules 2019, 24, 4432. [Google Scholar] [CrossRef]
- Manosroi, J.; Chankhampan, C.; Kitdamrongtham, W.; Zhang, J.; Abe, M.; Akihisa, T.; Manosroi, W.; Manosroi, A. In vivo anti-ageing activity of cream containing niosomes loaded with purple glutinous rice (Oryza sativa Linn.) extract. Int. J. Cosmet. Sci. 2020, 42, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Chaiyasut, C.; Kesika, P.; Sakdakampanat, P.; Peerajan, S.; Sivamaruthi, B. Formulation and evaluation of stability of thai purple rice bran-based cosmetic products. Asian J. Pharm. Clin. Res. 2018, 11, 99–104. [Google Scholar] [CrossRef]
- Silva, H.M.; Mageste, A.B.; Silva, S.J.B.; Dias Ferreira, G.M.; Ferreira, G.M.D. Anthocyanin immobilization in carboxymethylcellulose/starch films: A sustainable sensor for the detection of Al(III) ions in aqueous matrices. Carbohydr. Polym. 2020, 230, 115679. [Google Scholar] [CrossRef]
- Ge, Y.; Li, Y.; Bai, Y.; Yuan, C.; Wu, C.; Hu, Y. Intelligent gelatin/oxidized chitin nanocrystals nanocomposite films containing black rice bran anthocyanins for fish freshness monitorings. Int. J. Biol. Macromol. 2020, 155, 1296–1306. [Google Scholar] [CrossRef]
- Wu, C.; Sun, J.; Zheng, P.; Kang, X.; Chen, M.; Li, Y.; Ge, Y.; Hu, Y.; Pang, J. Preparation of an intelligent film based on chitosan/oxidized chitin nanocrystals incorporating black rice bran anthocyanins for seafood spoilage monitoring. Carbohydr. Polym. 2019, 222, 115006. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.; Liu, J.; Qin, Y.; Bai, R.; Zhang, X. Antioxidant and pH-sensitive films developed by incorporating purple and black rice extracts into chitosan matrix. Int. J. Biol. Macromol. 2019, 137, 307–316. [Google Scholar] [CrossRef]
- Prima, E.; Nugroho, H.; Refantero, G.; Panatarani, C.; Yuliarto, B. Performance of the dye-sensitized quasi-solid state solar cell with combined anthocyanin-ruthenium photosensitizer. RSC Adv. 2020, 2020, 36873–36886. [Google Scholar] [CrossRef]
- Zaidi, S.H.R.; Zakari, S.A.; Zhao, Q.; Khan, A.R.; Shah, J.M.; Cheng, F. Anthocyanin accumulation in black kernel mutant rice and its contribution to ROS detoxification in response to high temperature at the filling stage. Antioxidants 2019, 8, 510. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Srivastava, D.; Chauhan, A.S.; Indoliya, Y.; Singh, P.K.; Tiwari, S.; Fatima, T.; Mishra, S.K.; Dwivedi, S.; Agarwal, L.; et al. Root system architecture, physiological analysis and dynamic transcriptomics unravel the drought-responsive traits in rice genotypes. Ecotox. Environ. Safe. 2021, 207, 111252. [Google Scholar] [CrossRef] [PubMed]
- Ploenlap, P.; Pattanagul, W. Effects of exogenous abscisic acid on foliar anthocyanin accumulation and drought tolerance in purple rice. Biologia 2015, 70, 915–921. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, H.; Jiang, Y.; Wang, J.; Khan, A.; Li, P.; Cao, C. Metabolomic analysis reveals metabolites and pathways involved in grain quality traits of high-quality rice cultivars under a dry cultivation system. Food Chem. 2020, 326, 126845. [Google Scholar] [CrossRef] [PubMed]
- Chunthaburee, S.; Dongsansuk, A.; Sanitchon, J.; Pattanagul, W.; Theerakulpisut, P. Physiological and biochemical parameters for evaluation and clustering of rice cultivars differing in salt tolerance at seedling stage. Saudi J. Biol. Sci. 2016, 23, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Khan, M.A.; Asaf, S.; Lee, I.J.; Kim, K.M. Overexpression of OsF3H modulates WBPH stress by alteration of phenylpropanoid pathway at a transcriptomic and metabolomic level in Oryza sativa. Sci. Rep. 2020, 10, 14685. [Google Scholar] [CrossRef]
- Kim, E.G.; Yun, S.; Park, J.R.; Kim, K.M. Identification of F3H, major secondary metabolite-related gene that confers resistance against whitebacked planthopper through qtl mapping in rice. Plants 2021, 10, 81. [Google Scholar] [CrossRef]
- Sahoo, S.; Sanghamitra, P.; Nanda, N.; Pawar, S.; Pandit, E.; Bastia, R.; Muduli, K.C.; Pradhan, S.K. Association of molecular markers with physio-biochemical traits related to seed vigour in rice. Physiol. Mol. Biol. Plants 2020, 26, 1989–2003. [Google Scholar] [CrossRef]
- Poosri, S.; Thilavech, T.; Pasukamonset, P.; Suparpprom, C.; Adisakwattana, S. Studies on Riceberry rice (Oryza sativa L.) extract on the key steps related to carbohydrate and lipid digestion and absorption: A new source of natural bioactive substances. NFS J. 2019, 17, 17–23. [Google Scholar] [CrossRef]
- Han, M.; Bae, J.S.; Ban, J.J.; Shin, H.S.; Lee, D.H.; Chung, J.H. Black rice (Oryza sativa L.) extract modulates ultraviolet-induced expression of matrix metalloproteinases and procollagen in a skin cell model. Int. J. Mol. Med. 2018, 41, 3073–3080. [Google Scholar] [CrossRef]
- Palungwachira, P.; Tancharoen, S.; Phruksaniyom, C.; Klungsaeng, S.; Srichan, R.; Kikuchi, K.; Nararatwanchai, T. Antioxidant and anti-inflammatory properties of anthocyanins extracted from Oryza sativa L. in primary dermal fibroblasts. Oxidative Med. Cell. Longev. 2019, 2019, 2089817. [Google Scholar] [CrossRef]
- Galanakis, C.M.; Aldawoud, T.M.S.; Rizou, M.; Rowan, N.J.; Ibrahim, S.A. Food ingredients and active compounds against the coronavirus disease (COVID-19) pandemic: A comprehensive review. Foods 2020, 9, 1701. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi Pour, P.; Fakhri, S.; Asgary, S.; Farzaei, M.H.; Echeverría, J. The signaling pathways, and therapeutic targets of antiviral agents: Focusing on the antiviral approaches and clinical perspectives of anthocyanins in the management of viral diseases. Front. Pharmacol. 2019, 10, 1207. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.X.; Qi, S.S.; He, J.; Hu, C.Y.; Han, H.; Jiang, H.; Li, X.S. Cyanidin-3-glucoside from black rice ameliorates diabetic nephropathy via reducing blood glucose, suppressing oxidative stress and inflammation, and regulating transforming growth factor β1/smad expression. J. Agric. Food. Chem. 2020, 68, 4399–4410. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Xu, Y.; Jia, C.; Zhang, B.; Niu, M.; Zhao, S. Structural changes of rice starch and activity inhibition of starch digestive enzymes by anthocyanins retarded starch digestibility. Carbohydr. Polym. 2021, 261, 117841. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Wu, Y.; Chen, B.; Yao, Y.; Wang, Y.; Bai, H.; Li, C.; Yang, Y.; Chen, Y. Cyanidin-3-O-β-glucoside attenuates allergic airway inflammation by modulating the IL-4Rα-STAT6 signaling pathway in a murine asthma model. Int. Immunopharmacol. 2019, 69, 1–10. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Y.; Liu, G.; Hao, S.; Wang, C.; Wang, Y. Black rice anthocyanin-rich extract and rosmarinic acid, alone and in combination, protect against DSS-induced colitis in mice. Food Funct. 2018, 9, 2796–2808. [Google Scholar] [CrossRef]
- Kim, S.H.; Woo, H.J.; Lee, M.H.; Park, M.; Nagendran, T.; Rhee, K.J.; Lee, D.; Jin, Y.B.; Choi, S.W.; Seo, W.D.; et al. Antimicrobial effects of black rice extract on Helicobacter pylori infection in Mongolian gerbil. J. Cereal Sci. 2019, 85, 1–5. [Google Scholar] [CrossRef]
- Wang, H.; Liu, D.; Ji, Y.; Liu, Y.; Xu, L.; Guo, Y. Dietary supplementation of black rice anthocyanin extract regulates cholesterol metabolism and improves gut microbiota dysbiosis in C57BL/6J mice fed a high-fat and cholesterol diet. Mol. Nutr. Food Res. 2020, 64, 1900876. [Google Scholar] [CrossRef]
- Park, Y.M.; Lee, H.Y.; Shin, D.Y.; Lee, Y.H.; Yang, Y.J.; Lee, H.S.; Lee, J.O.; Choi, K.S.; Kang, J.H.; Cho, Y.H.; et al. Immunostimulatory activity of black rice bran in cyclophosphamide-induced immunosuppressed rats. Nat. Prod. Commun. 2020, 15, 1–11. [Google Scholar] [CrossRef]
- Fan, M.J.; Yeh, P.H.; Lin, J.P.; Huang, A.C.; Lien, J.C.; Lin, H.Y.; Chung, J.G. Anthocyanins from black rice (Oryza sativa) promote immune responses in leukemia through enhancing phagocytosis of macrophages in vivo. Exp. Ther. Med. 2017, 14, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Ampawong, S.; Kengkoom, K.; Sukphopetch, P.; Aramwit, P.; Muangkaew, W.; Kanjanapruthipong, T.; Buaban, T. Evaluating the effect of rice (Oryza sativa L.: SRNC05053-6-2) crude extract on psoriasis using in vitro and in vivo models. Sci. Rep. 2020, 10, 17618. [Google Scholar] [CrossRef] [PubMed]
- Tancharoen, S.; Shakya, P.; Narkpinit, S.; Dararat, P.; Kikuchi, K. Anthocyanins Extracted from Oryza sativa L. Prevent fluorouracil-induced nuclear factor-κB activation in oral mucositis: In vitro and in vivo studies. Int. J. Mol. Sci. 2018, 19, 2981. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade | Entirely Dark or Almost Entirely Dark Kernels (%) | Allowed Off Color Kernels (%) | Other Color Kernels (%) | Other Type Kernels (%) | Size of Head Rice (Part) (mm) |
|---|---|---|---|---|---|
| Non-glutinous rice | |||||
| Prime quality | ≥80 | ≤20 | ≤4.0 | ≤1.0 | ≥7 |
| Superb quality | ≥65 | ≤35 | ≤4.0 | ≤1.0 | ≥7 |
| Glutinous rice | |||||
| Best quality | ≥30 | ≤70 | ≤6 | ≤1.0 | ≥6.5 |
| Standard quality | ≥15–<30 | >70–≤ 85 | ≤6 | ≤1.0 | ≥6.5 |
| Variety | Ecotype | TAC (mg /100 g) | Amylose (%) | Aroma Property | Rice Sample | Measurement/Extract Solution | Reference |
|---|---|---|---|---|---|---|---|
| Glutinous Rice | |||||||
| Niaw Dam Pleuk Dam | unknown | 381 | 9.7 | nd | whole grain | pH different/acidified methanol | [57,58] |
| Niaw Dam Pleuk Khow | lowland | 442 | 8.9 | nd | whole grain | pH different/acidified methanol | [57,58] |
| Kum Doi Saket | lowland | 119 | 6.2 | A | whole grain | pH different/ acidified methanol | [59,60] |
| Leum pua | upland | 212 | 3.4 | A | flour | pH different/nd | [60,61] |
| Pieisu | upland | 271 | nd | nd | whole grain | pH different/acidified methanol | [62] |
| KHCMU | upland | 58 | nd | A | whole grain | pH different/acidified methanol | [62] |
| Kam Med-dam | upland | 222 | nd | nd | whole grain | pH different/acidified methanol | [55] |
| Kam Leuang-dam | upland | 150 | nd | nd | whole grain | pH different/ acidified methanol | [55] |
| Kalobhat | unknown | 57 | 5.4 | nd | whole grain | Colorimetric/acidified methanol | [24] |
| Chakhao | lowland | 276 | 6.0 | A | flour | HPLC/acidified methanol | [56] |
| Non-Glutinous Rice | |||||||
| Hom Nil | lowland | 65 | 14.2 | nd | whole grain | Colorimetric/distilled water | [63] |
| Riceberry | lowland | 129 | 17.5 | nd | flour | pH different/nd | [62] |
| Malinil Surin | lowland | 75 | 13.8 | nd | flour | pH different/nd | [60] |
| KJ CMU 107 | lowland | 12 | nd | A | whole grain | pH different/acidified methanol | [62] |
| Mamihunger | unknown | 34 | 17.6 | nd | whole grain | colorimetric/ acidified methanol | [24] |
| Italian rice | lowland | 54 | 9.9 | nd | whole grain | pH different /acidified methanol | [64] |
| Jatinangor | unknown | 40 | nd | nd | Whole grain | colorimetric/ethanol and citric acid | [65] |
| Variety | Rice Sample | Concentration (mg/100 g) | Ref. | |
|---|---|---|---|---|
| C3G | P3G | |||
| Kum Doi Saket CMU 125 | whole grain whole grain | 50 200 | 48 65 | [73] |
| Hom Nil Riceberry | whole grain whole grain | 133 48 | 20 7 | [74] |
| Hom Nil Leum Pua | Bran bran | 926 2277 | 1422 792 | [77] |
| WC 320 | bran | 1103 | 33 | [47] |
| Khao Gam Pah E-Kaw | whole grain | 88 | 32 | [78] |
| Artemide | whole grain | 199 | 11 | [31] |
| Chakhao | whole grain | 208 | 45 | [56] |
| IAC 600 | whole grain | 130 | 10 | [27] |
| Heimi 2420 | whole grain | 111 | 31 | [79] |
| PER-Z#3 PER-Z#14 | polished grain polished grain | 53 27 | 44 26 | [42] |
| Purple Rice Anthocyanin Application | Ref. |
|---|---|
| Functional Food | |
| Bread made from anthocyanin-rich purple rice improved postprandial plasma glucose and antioxidant status in healthy subjects | [130] |
| Anthocyanin-rich purple rice flour can be used as a gluten-free ingredient in bread providing FRAP antioxidant activity | [131] |
| Bread fortified with 1–4% of anthocyanin-rich rice powder has a low digestion rate that provides health benefits | [132] |
| Anthocyanin-rich rice beverage added with xanthan gum has high thermal stability | [133] |
| Germinated purple rice that retains anthocyanins has good sensory characteristics | [134] |
| Crispy rice bar made purple rice provides high anthocyanin | [135] |
| Fresh germinated purple rice noodles provide total anthocyanin and DPPH and FRAP antioxidant capacity | [136] |
| 15% of anthocyanin-rich purple rice extract supplemented pasta contains high anthocyanins and antioxidant capacity (DPPH and FRAP) | [137] |
| Purple rice sprouts present high total anthocyanin and could be developed for natural health products | [138] |
| 0.25% of anthocyanin-rice purple rice extract supplemented yogurt suppresses postprandial glucose level and improved plasma antioxidant capacity in healthy volunteers | [19] |
| 0.06% of purple rice anthocyanin extract inhibited lipid and protein oxidation in whey-protein-stabilized food emulsions | [21] |
| Other Purposes | |
| Anthocyanin extract used in cream exhibited in vitro antioxidant activity and in vivo anti-ageing activity on human skin | [139] |
| Rice bran extracts containing anthocyanins have been investigated as ingredients in the cosmetic formulations that exhibit antioxidant capacity | [140] |
| Purple rice anthocyanin is used in the colorimetric sensing of Al+3 | [141] |
| Packaging film based on oxidized chitin nanocrystals/gelatin incorporating purple rice bran anthocyanins has potential in freshness monitoring | [142] |
| 3% of purple rice bran anthocyanins added in oxidized-chitin nanocrystals/chitosan matrix are able to monitor the spoiling of seafoods | [143] |
| 1% of purple rice anthocyanins incorporated into chitosan packing films can be used to monitor pork spoilage | [144] |
| Anthocyanin dye increased the performance of dye-sensitized solar cells | [145] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamuangmorn, S.; Prom-u-Thai, C. The Potential of High-Anthocyanin Purple Rice as a Functional Ingredient in Human Health. Antioxidants 2021, 10, 833. https://doi.org/10.3390/antiox10060833
Yamuangmorn S, Prom-u-Thai C. The Potential of High-Anthocyanin Purple Rice as a Functional Ingredient in Human Health. Antioxidants. 2021; 10(6):833. https://doi.org/10.3390/antiox10060833
Chicago/Turabian StyleYamuangmorn, Supapohn, and Chanakan Prom-u-Thai. 2021. "The Potential of High-Anthocyanin Purple Rice as a Functional Ingredient in Human Health" Antioxidants 10, no. 6: 833. https://doi.org/10.3390/antiox10060833
APA StyleYamuangmorn, S., & Prom-u-Thai, C. (2021). The Potential of High-Anthocyanin Purple Rice as a Functional Ingredient in Human Health. Antioxidants, 10(6), 833. https://doi.org/10.3390/antiox10060833