
Impact of Simulated Gastrointestinal Conditions on Antiglycoxidant and α-Glucosidase Inhibition Capacities of Cyanidin-3-O-Glucoside

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
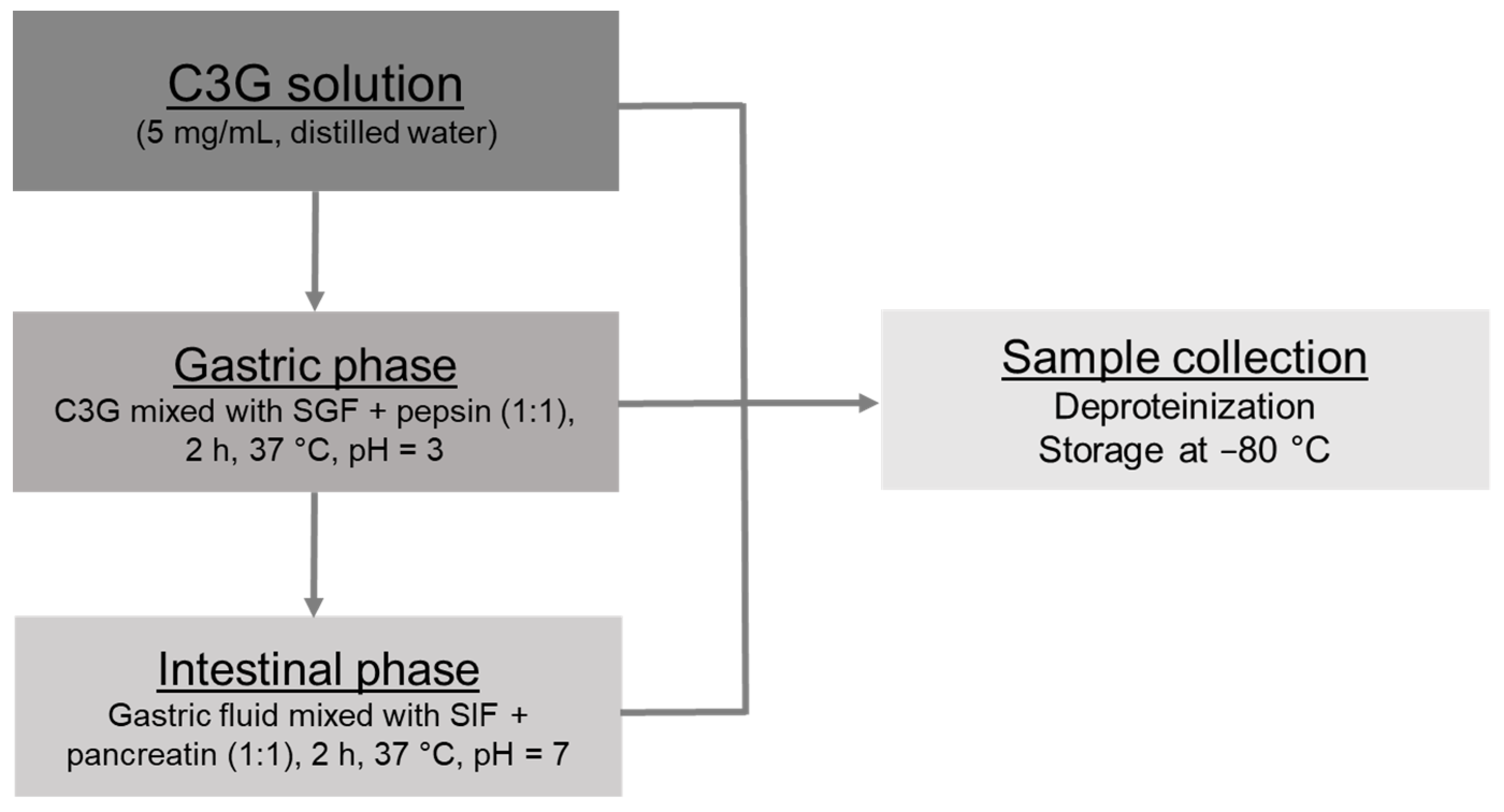
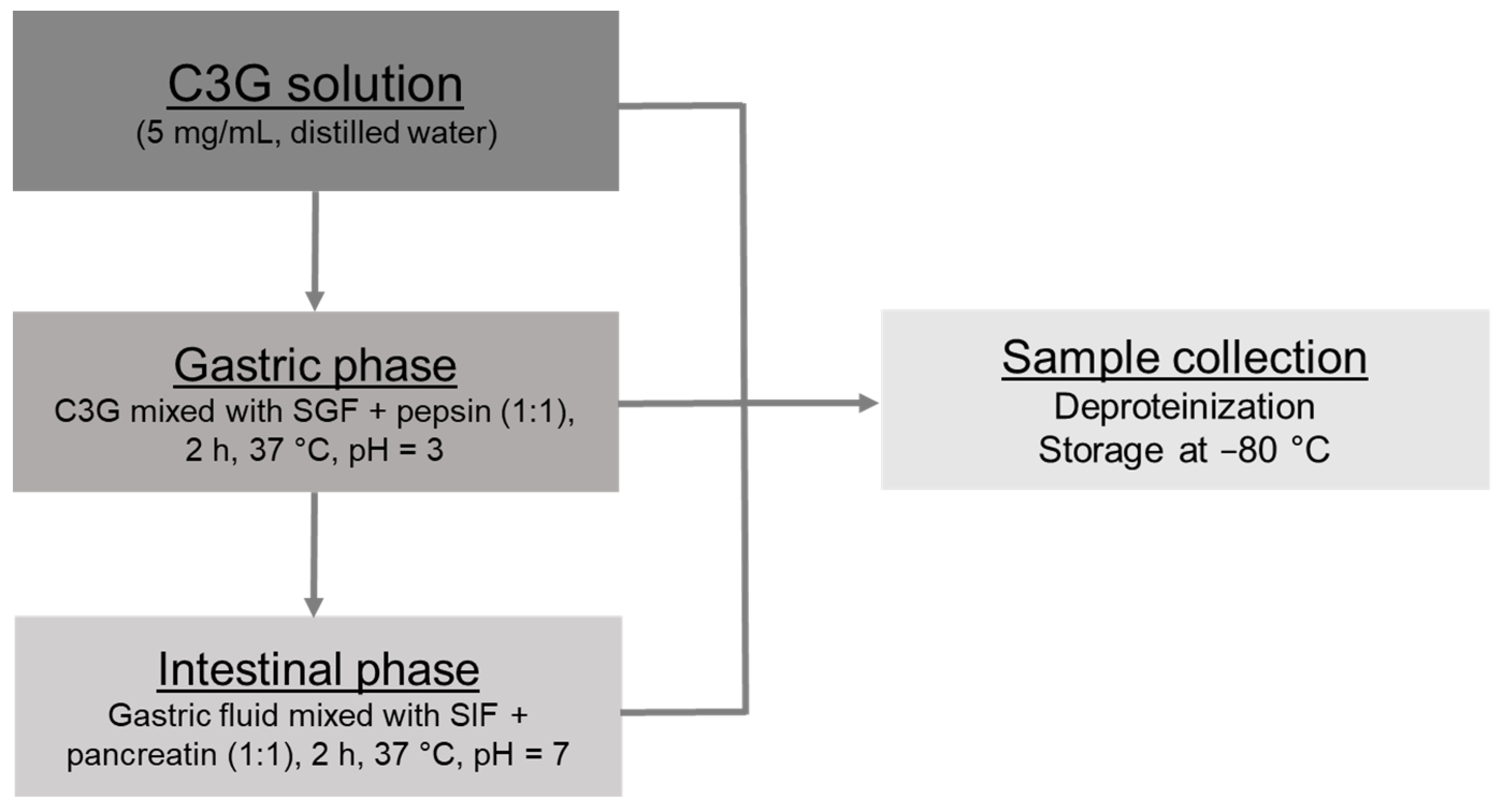
2.2. In Vitro Gastrointestinal Digestion
2.2.1. Gastric and Intestinal Phases
2.2.2. Sample Management
2.3. Spectrometric and Fluorometric Evaluations
2.4. HPLC Analysis
2.5. Statistical Analyses
3. Results and Discussion
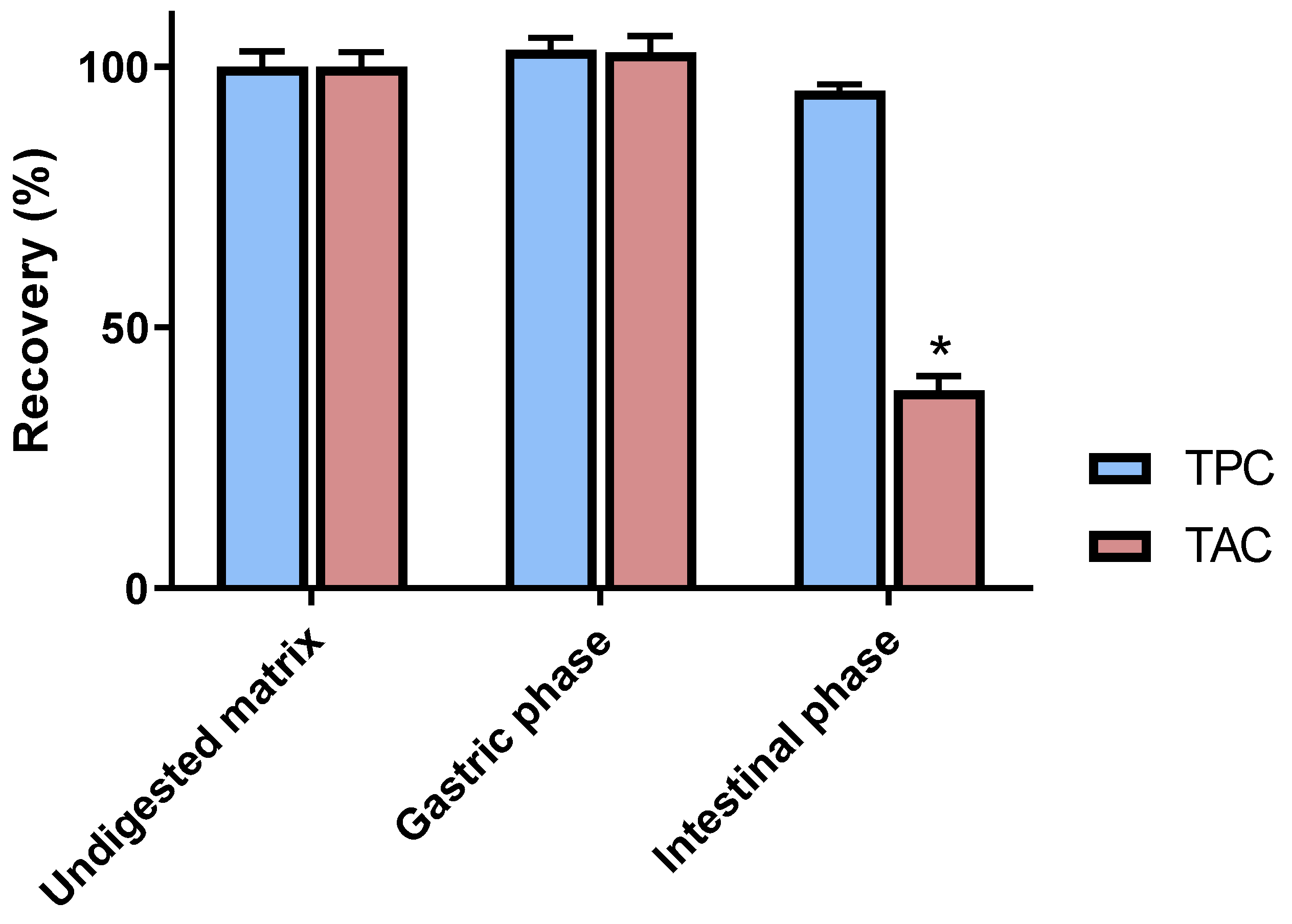
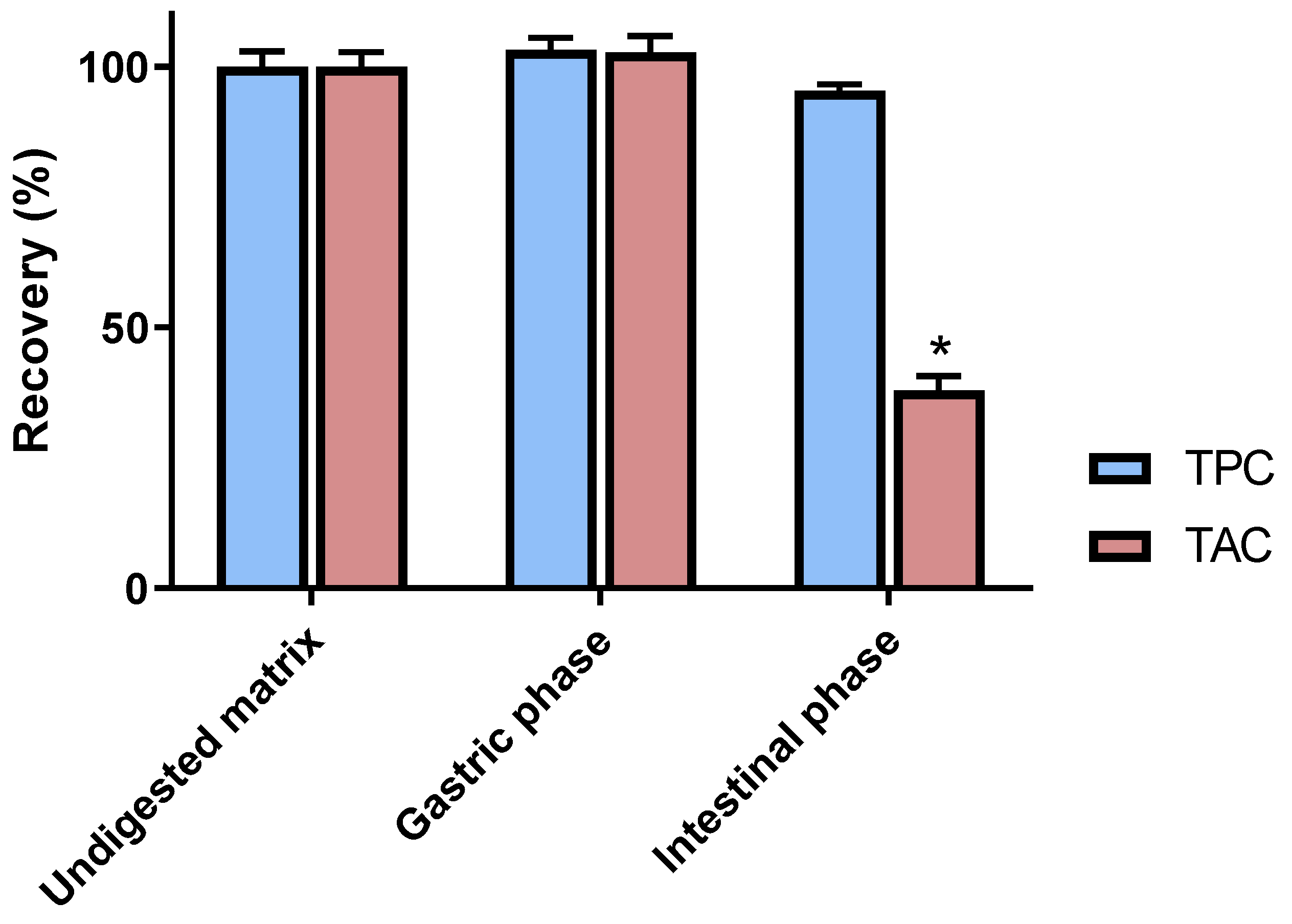
3.1. Chemical Analyses of Digested C3G Samples
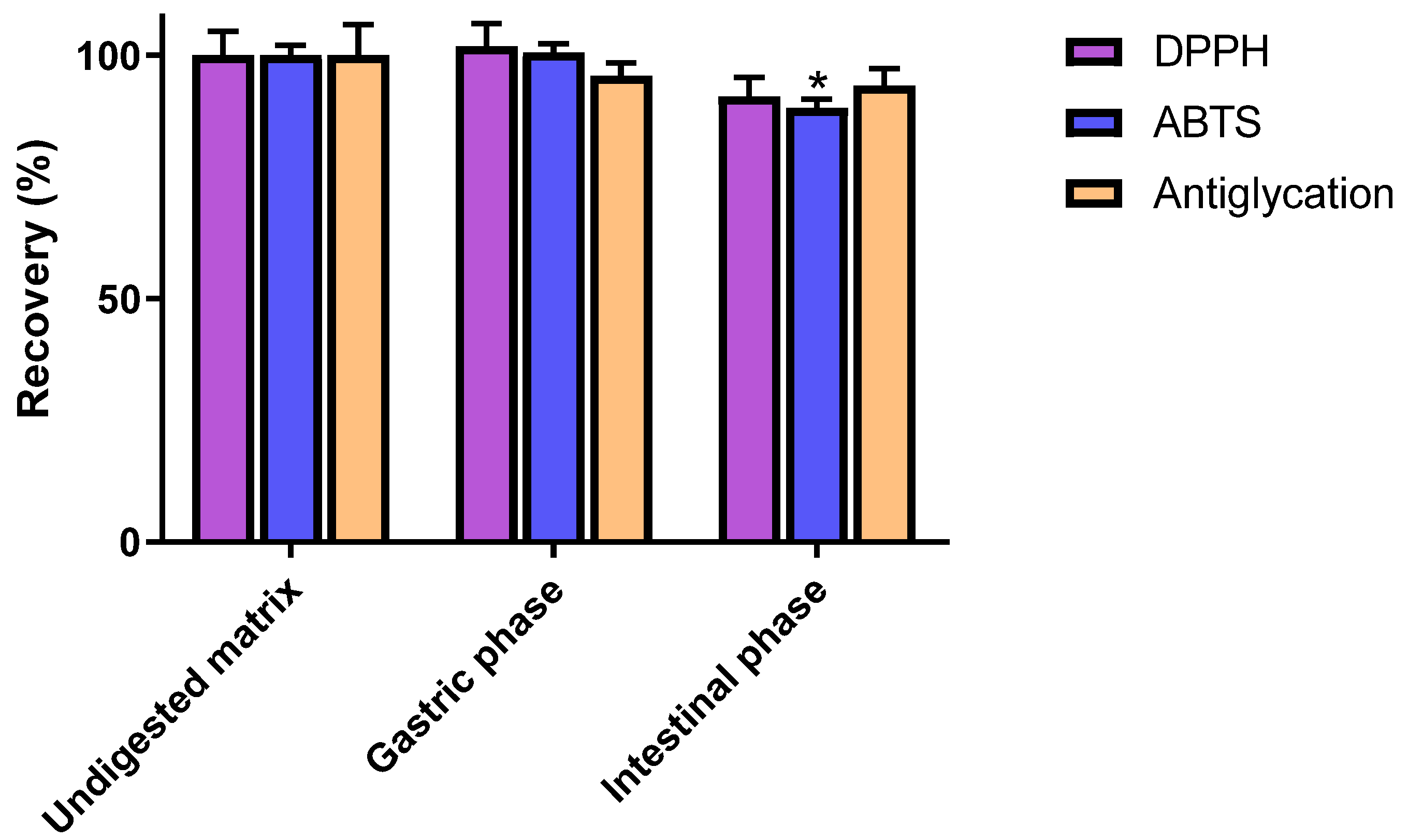
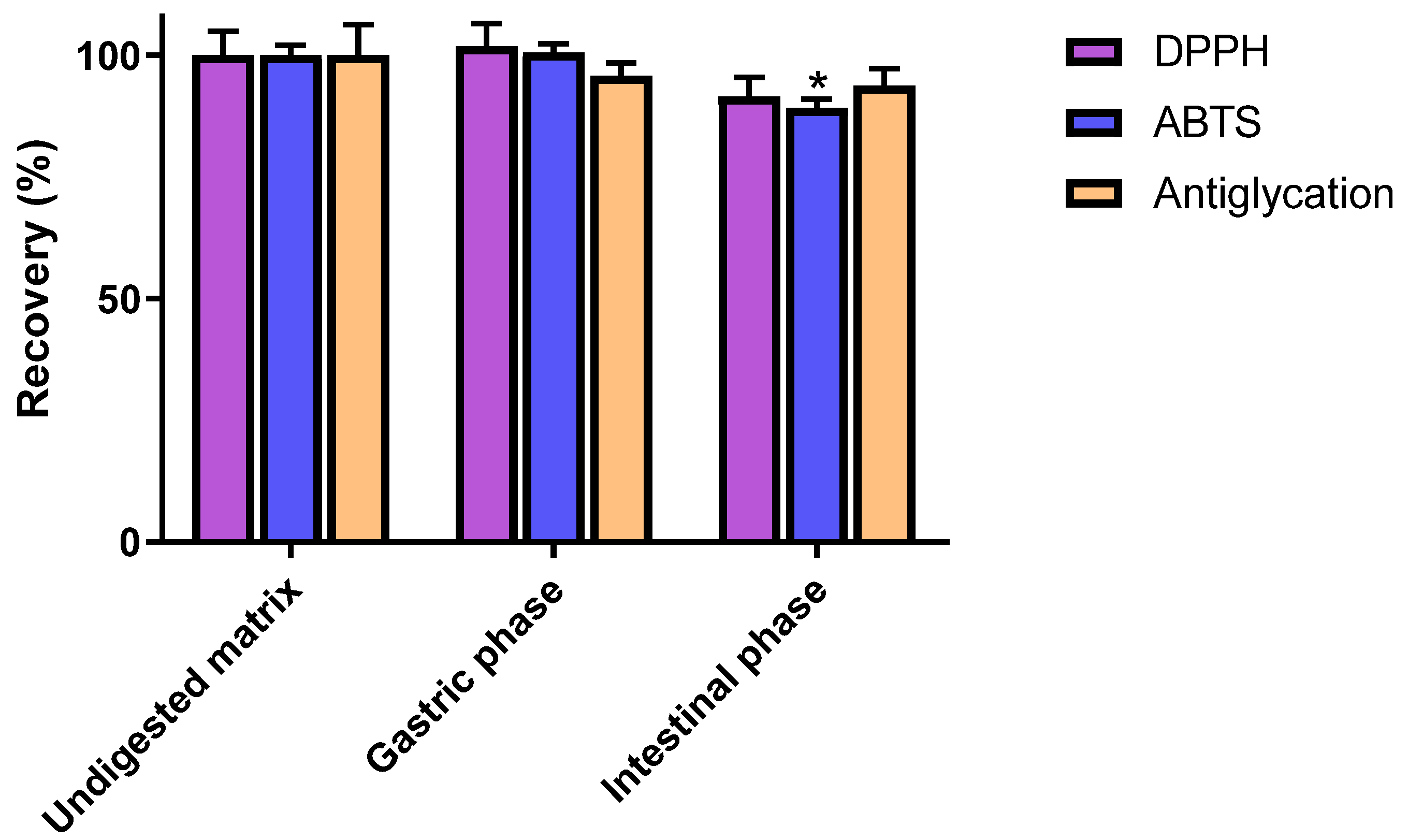
3.2. Impact of In Vitro Digestion on Radical Scavenging and Antiglycation Activities of C3G
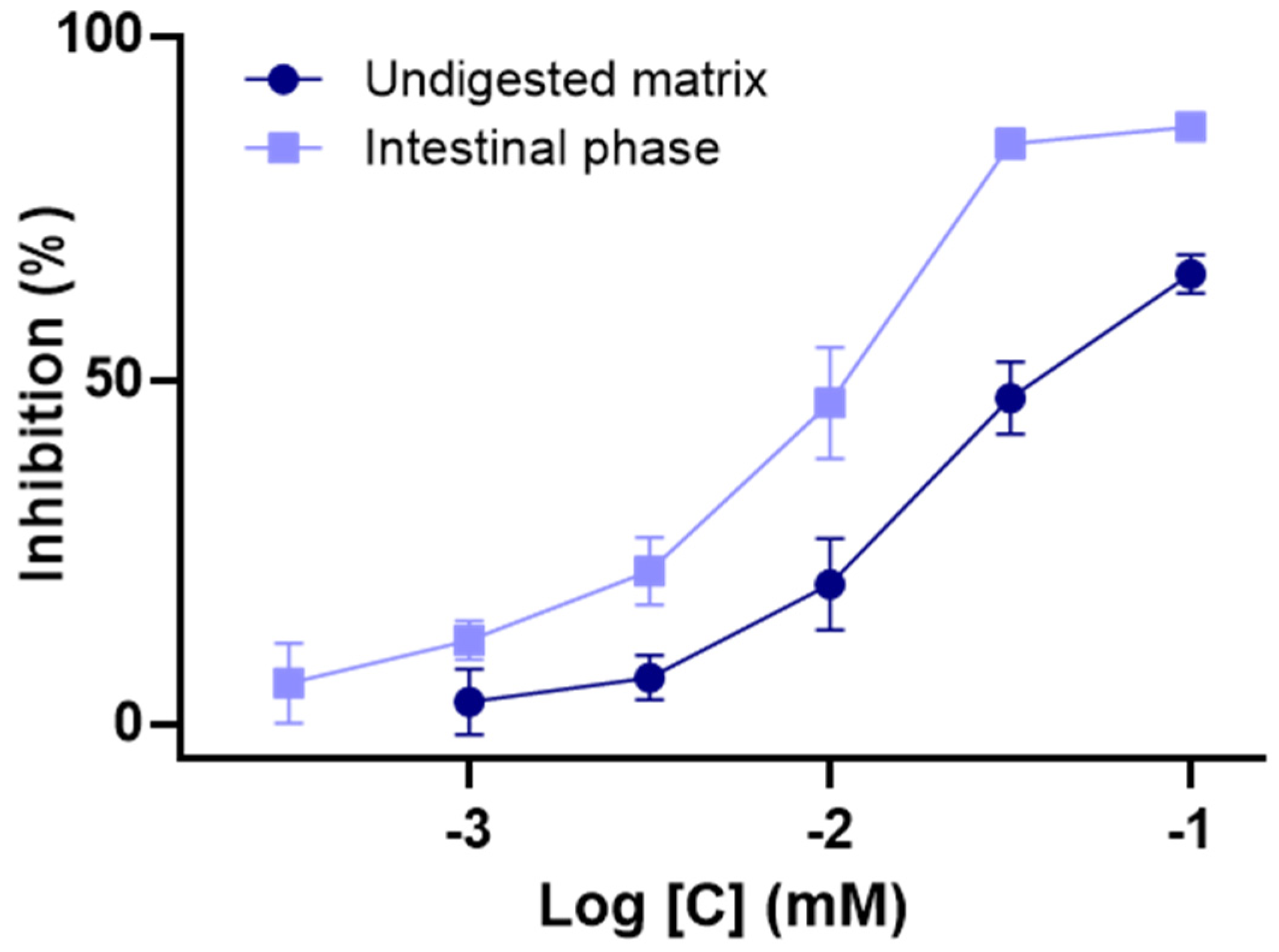
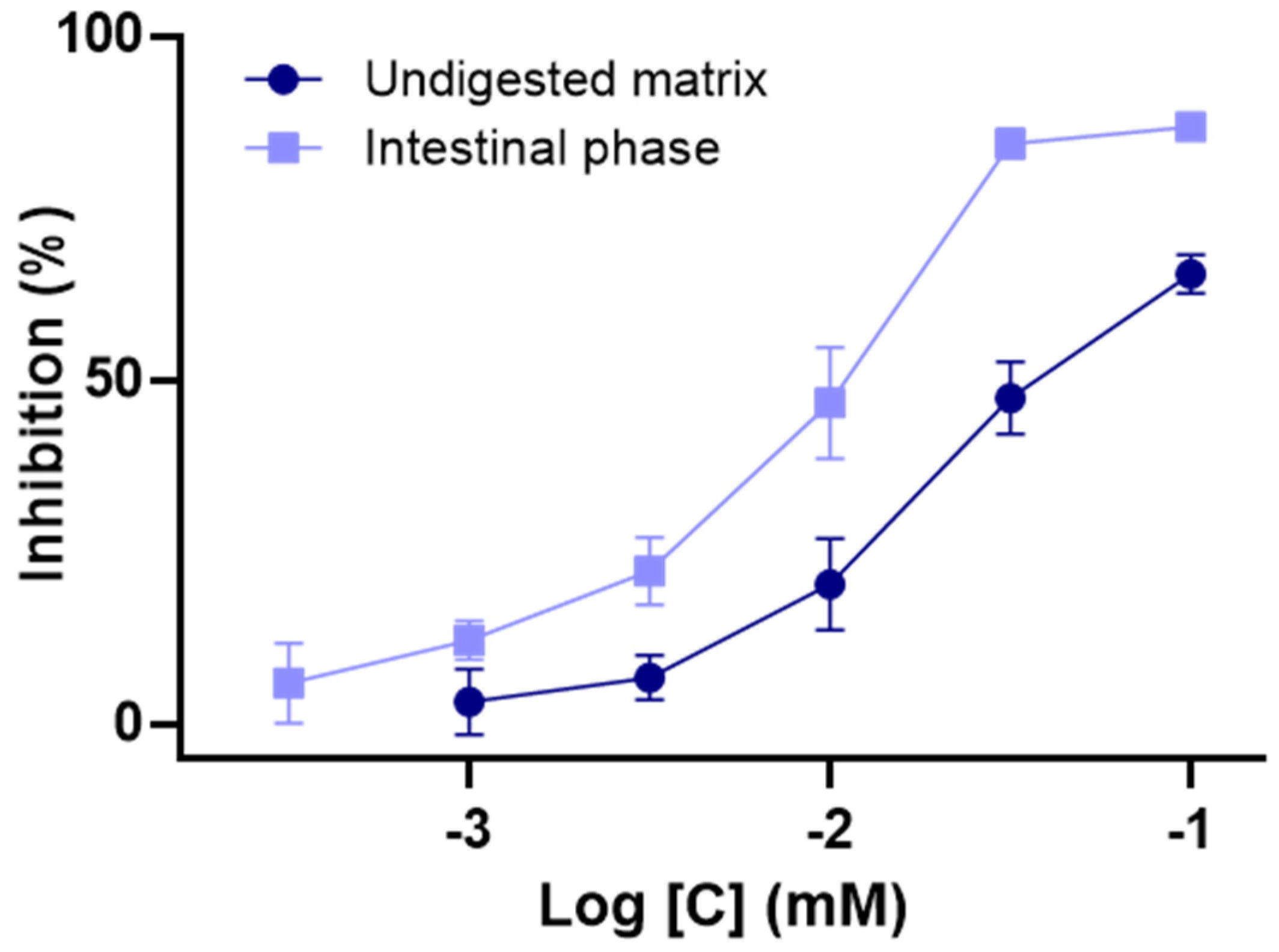
3.3. Impact of In Vitro Digestion on α-Glucosidase Inhibition Properties of C3G
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castañeda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical Studies of Anthocyanins: A Review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Wallace, T.C.; Giusti, M.M. Anthocyanins—Nature’s Bold, Beautiful, and Health-Promoting Colors. Foods 2019, 8, 550. [Google Scholar] [CrossRef] [Green Version]
- Eker, M.E.; Aaby, K.; Budic-Leto, I.; Brnčić, S.R.; El, S.N.; Karakaya, S.; Simsek, S.; Manach, C.; Wiczkowski, W.; de Pascual-Teresa, S. A Review of Factors Affecting Anthocyanin Bioavailability: Possible Implications for the Inter-Individual Variability. Foods 2019, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.-M.; Chia, L.-S.; Goh, N.-K.; Chia, T.-F.; Brouillard, R. Analysis and Biological Activities of Anthocyanins. Phytochemistry 2003, 64, 923–933. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Nunes, A.R.; Falcão, A.; Alves, G.; Silva, L.R. Dietary Effects of Anthocyanins in Human Health: A Comprehensive Review. Pharmaceuticals 2021, 14, 690. [Google Scholar] [CrossRef]
- Olivas-Aguirre, F.J.; Rodrigo-García, J.; Martínez-Ruiz, N.D.R.; Cárdenas-Robles, A.I.; Mendoza-Díaz, S.O.; Álvarez-Parrilla, E.; González-Aguilar, G.A.; de la Rosa, L.A.; Ramos-Jiménez, A.; Wall-Medrano, A. Cyanidin-3-O-Glucoside: Physical-Chemistry, Foodomics and Health Effects. Molecules 2016, 21, 1264. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Gu, L.; Prior, R.L.; McKay, S. Characterization of Anthocyanins and Proanthocyanidins in Some Cultivars of Ribes, Aronia, and Sambucus and Their Antioxidant Capacity. J. Agric. Food Chem. 2004, 52, 7846–7856. [Google Scholar] [CrossRef]
- Cho, M.J.; Howard, L.R.; Prior, R.L.; Clark, J.R. Flavonoid Glycosides and Antioxidant Capacity of Various Blackberry, Blueberry and Red Grape Genotypes Determined by High-Performance Liquid Chromatography/Mass Spectrometry. J. Sci. Food Agric. 2004, 84, 1771–1782. [Google Scholar] [CrossRef]
- Määttä-Riihinen, K.R.; Kamal-Eldin, A.; Törrönen, A.R. Identification and Quantification of Phenolic Compounds in Berries of Fragaria and Rubus Species (Family Rosaceae). J. Agric. Food Chem. 2004, 52, 6178–6187. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Heinonen, M. Antioxidant Activity of Anthocyanins and Their Aglycons. J. Agric. Food Chem. 2003, 51, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Speer, H.; D’Cunha, N.M.; Alexopoulos, N.I.; McKune, A.J.; Naumovski, N. Anthocyanins and Human Health—A Focus on Oxidative Stress, Inflammation and Disease. Antioxidants 2020, 9, 366. [Google Scholar] [CrossRef]
- Ma, H.; Johnson, S.; Liu, W.; DaSilva, N.; Meschwitz, S.; Dain, J.; Seeram, N. Evaluation of Polyphenol Anthocyanin-Enriched Extracts of Blackberry, Black Raspberry, Blueberry, Cranberry, Red Raspberry, and Strawberry for Free Radical Scavenging, Reactive Carbonyl Species Trapping, Anti-Glycation, Anti-β-Amyloid Aggregation, and Microglial Neuroprotective Effects. IJMS 2018, 19, 461. [Google Scholar] [CrossRef] [Green Version]
- Fraisse, D.; Bred, A.; Felgines, C.; Senejoux, F. Screening and Characterization of Antiglycoxidant Anthocyanins from Vaccinium Myrtillus Fruit Using DPPH and Methylglyoxal Pre-Column HPLC Assays. Antioxidants 2020, 9, 512. [Google Scholar] [CrossRef]
- Konczak, I.; Zhang, W. Anthocyanins—More Than Nature’s Colours. J. Biomed. Biotechnol. 2004, 2004, 239–240. [Google Scholar] [CrossRef] [Green Version]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A Comprehensive Review of Their Chemical Properties and Health Effects on Cardiovascular and Neurodegenerative Diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef]
- Jaiswal, V.; Park, M.; Lee, H.-J. Comparative Transcriptome Analysis of the Expression of Antioxidant and Immunity Genes in the Spleen of a Cyanidin 3-O-Glucoside-Treated Alzheimer’s Mouse Model. Antioxidants 2021, 10, 1435. [Google Scholar] [CrossRef]
- Putta, S.; Yarla, N.S.; Kumar, K.E.; Lakkappa, D.B.; Kamal, M.A.; Scotti, L.; Scotti, M.T.; Ashraf, G.M.; Rao, B.S.B.; Reddy, G.V.; et al. Preventive and Therapeutic Potentials of Anthocyanins in Diabetes and Associated Complications. CMC 2019, 25, 5347–5371. [Google Scholar] [CrossRef] [PubMed]
- Mcdougall, G.J.; Stewart, D. The Inhibitory Effects of Berry Polyphenols on Digestive Enzymes. BioFactors 2005, 23, 189–195. [Google Scholar] [CrossRef]
- Fang, J. Bioavailability of Anthocyanins. Drug Metab. Rev. 2014, 46, 508–520. [Google Scholar] [CrossRef]
- Kahle, K.; Kraus, M.; Scheppach, W.; Ackermann, M.; Ridder, F.; Richling, E. Studies on Apple and Blueberry Fruit Constituents: Do the Polyphenols Reach the Colon after Ingestion? Mol. Nutr. Food Res. 2006, 50, 418–423. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A Standardised Static in vitro Digestion Method Suitable for Food—An International Consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- McDougall, G.J.; Fyffe, S.; Dobson, P.; Stewart, D. Anthocyanins from Red Cabbage—Stability to Simulated Gastrointestinal Digestion. Phytochemistry 2007, 68, 1285–1294. [Google Scholar] [CrossRef]
- Fraisse, D.; Bred, A.; Felgines, C.; Senejoux, F. Stability and Antiglycoxidant Potential of Bilberry Anthocyanins in Simulated Gastrointestinal Tract Model. Foods 2020, 9, 1695. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Séro, L.; Sanguinet, L.; Blanchard, P.; Dang, B.T.; Morel, S.; Richomme, P.; Seraphin, D.; Derbré, S. Tuning a 96-Well Microtiter Plate Fluorescence-Based Assay to Identify AGE Inhibitors in Crude Plant Extracts. Molecules 2013, 18, 14320–14339. [Google Scholar] [CrossRef] [Green Version]
- Mohamed Sham Shihabudeen, H.; Hansi Priscilla, D.; Thirumurugan, K. Cinnamon Extract Inhibits α-Glucosidase Activity and Dampens Postprandial Glucose Excursion in Diabetic Rats. Nutr. Metab. 2011, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loypimai, P.; Moongngarm, A.; Chottanom, P. Thermal and PH Degradation Kinetics of Anthocyanins in Natural Food Colorant Prepared from Black Rice Bran. J. Food Sci. Technol. 2016, 53, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangles, O.; Fenger, J.-A. The Chemical Reactivity of Anthocyanins and Its Consequences in Food Science and Nutrition. Molecules 2018, 23, 1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Yuan, C.; Wang, H.; Han, F.; Liu, Y.; Wang, L.; Liu, Y. Stability of Anthocyanins and Their Degradation Products from Cabernet Sauvignon Red Wine under Gastrointestinal PH and Temperature Conditions. Molecules 2018, 23, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Chen, H.; Zhang, W.; Ding, Y.; Zhao, T.; Zhang, M.; Mao, G.; Feng, W.; Wu, X.; Yang, L. Bioaccessibility and Biotransformation of Anthocyanin Monomers Following in Vitro Simulated Gastric-Intestinal Digestion and in Vivo Metabolism in Rats. Food Funct. 2019, 10, 6052–6061. [Google Scholar] [CrossRef]
- Liang, T.; Guan, R.; Wang, Z.; Shen, H.; Xia, Q.; Liu, M. Comparison of Anticancer Activity and Antioxidant Activity between Cyanidin-3-O-Glucoside Liposomes and Cyanidin-3-O-Glucoside in Caco-2 Cells in Vitro. RSC Adv. 2017, 7, 37359–37368. [Google Scholar] [CrossRef] [Green Version]
- Bedekar, A.; Shah, K.; Koffas, M. Natural Products for Type II Diabetes Treatment. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2010; Volume 71, pp. 21–73. ISBN 978-0-12-380993-3. [Google Scholar]
- Yang, H.K.; Lee, S.-H.; Shin, J.; Choi, Y.-H.; Ahn, Y.-B.; Lee, B.-W.; Rhee, E.J.; Min, K.W.; Yoon, K.-H. Acarbose Add-on Therapy in Patients with Type 2 Diabetes Mellitus with Metformin and Sitagliptin Failure: A Multicenter, Randomized, Double-Blind, Placebo-Controlled Study. Diabetes Metab. J. 2019, 43, 287. [Google Scholar] [CrossRef] [PubMed]
- Cásedas, G.; Les, F.; González-Burgos, E.; Gómez-Serranillos, M.P.; Smith, C.; López, V. Cyanidin-3-O-Glucoside Inhibits Different Enzymes Involved in Central Nervous System Pathologies and Type-2 Diabetes. S. Afr. J. Bot. 2019, 120, 241–246. [Google Scholar] [CrossRef]
- Choi, K.; Choi, S.I.; Park, M.H.; Han, J.S. Cyanidin-3-O-glucoside Ameliorates Postprandial Hyperglycemia in Diabetic Mice. J. Life Sci. 2017, 27, 32–37. [Google Scholar] [CrossRef]
- Salvador, Â.; Król, E.; Lemos, V.; Santos, S.; Bento, F.; Costa, C.; Almeida, A.; Szczepankiewicz, D.; Kulczyński, B.; Krejpcio, Z.; et al. Effect of Elderberry (Sambucus nigra L.) Extract Supplementation in STZ-Induced Diabetic Rats Fed with a High-Fat Diet. IJMS 2016, 18, 13. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.-D.; Zhang, B.; Zhang, J.-K.; Xu, C.-J.; Wu, Y.-L.; Li, X.; Chen, K.-S. Cyanidin-3-Glucoside-Rich Extract from Chinese Bayberry Fruit Protects Pancreatic β Cells and Ameliorates Hyperglycemia in Streptozotocin-Induced Diabetic Mice. J. Med. Food 2012, 15, 288–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, D.; Sarkar, S.; De Silva, A.B.K.H.; D’Souza, K.; Kienesberger, P.; Rupasinghe, H.P.V.; Pulinilkunnil, T. Cyanidin-3-O-Glucoside Rich Extract From Haskap Berry Improves Glucose Homeostasis and Insulin Sensitivity in Diet-Induced Obese Mice. Can. J. Diabetes 2018, 42, S55. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Undigested Matrix | Gastric Phase | Intestinal Phase |
|---|---|---|---|
| DPPH scavenging activity (μmol of Trolox eq/g) | 3761 ± 189 a | 3834 ± 176 a | 3445 ± 145 a |
| ABTS scavenging activity (μmol of Trolox eq/g) | 9218 ± 195 a | 9268 ± 172 a | 8225 ± 147 b |
| Antiglycation activity (IC50, µg/mL) | 40.52 ± 2.61 a | 42.00 ± 1.10 a | 42.97 ± 1.55 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraisse, D.; Bred, A.; Felgines, C.; Senejoux, F. Impact of Simulated Gastrointestinal Conditions on Antiglycoxidant and α-Glucosidase Inhibition Capacities of Cyanidin-3-O-Glucoside. Antioxidants 2021, 10, 1670. https://doi.org/10.3390/antiox10111670
Fraisse D, Bred A, Felgines C, Senejoux F. Impact of Simulated Gastrointestinal Conditions on Antiglycoxidant and α-Glucosidase Inhibition Capacities of Cyanidin-3-O-Glucoside. Antioxidants. 2021; 10(11):1670. https://doi.org/10.3390/antiox10111670
Chicago/Turabian StyleFraisse, Didier, Alexis Bred, Catherine Felgines, and François Senejoux. 2021. "Impact of Simulated Gastrointestinal Conditions on Antiglycoxidant and α-Glucosidase Inhibition Capacities of Cyanidin-3-O-Glucoside" Antioxidants 10, no. 11: 1670. https://doi.org/10.3390/antiox10111670
APA StyleFraisse, D., Bred, A., Felgines, C., & Senejoux, F. (2021). Impact of Simulated Gastrointestinal Conditions on Antiglycoxidant and α-Glucosidase Inhibition Capacities of Cyanidin-3-O-Glucoside. Antioxidants, 10(11), 1670. https://doi.org/10.3390/antiox10111670