Genetic Relationships Between Ethanol-Induced Conditioned Place Aversion and Other Ethanol Phenotypes in 15 Inbred Mouse Strains

Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Apparatus
2.3. Procedure
2.3.1. Habituation
2.3.2. Conditioning
2.3.3. Place preference tests
2.3.4. Data Analyses
3. Results
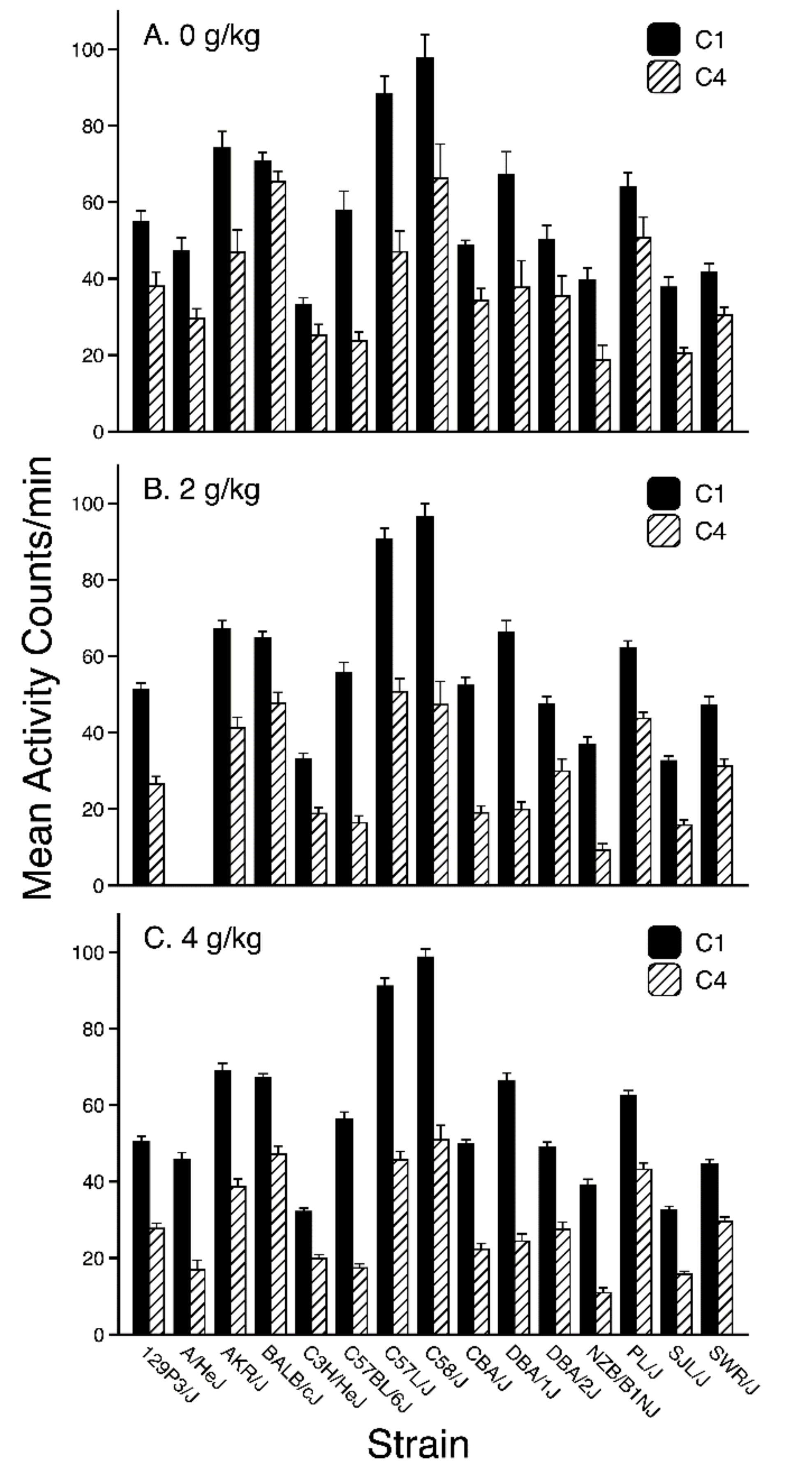
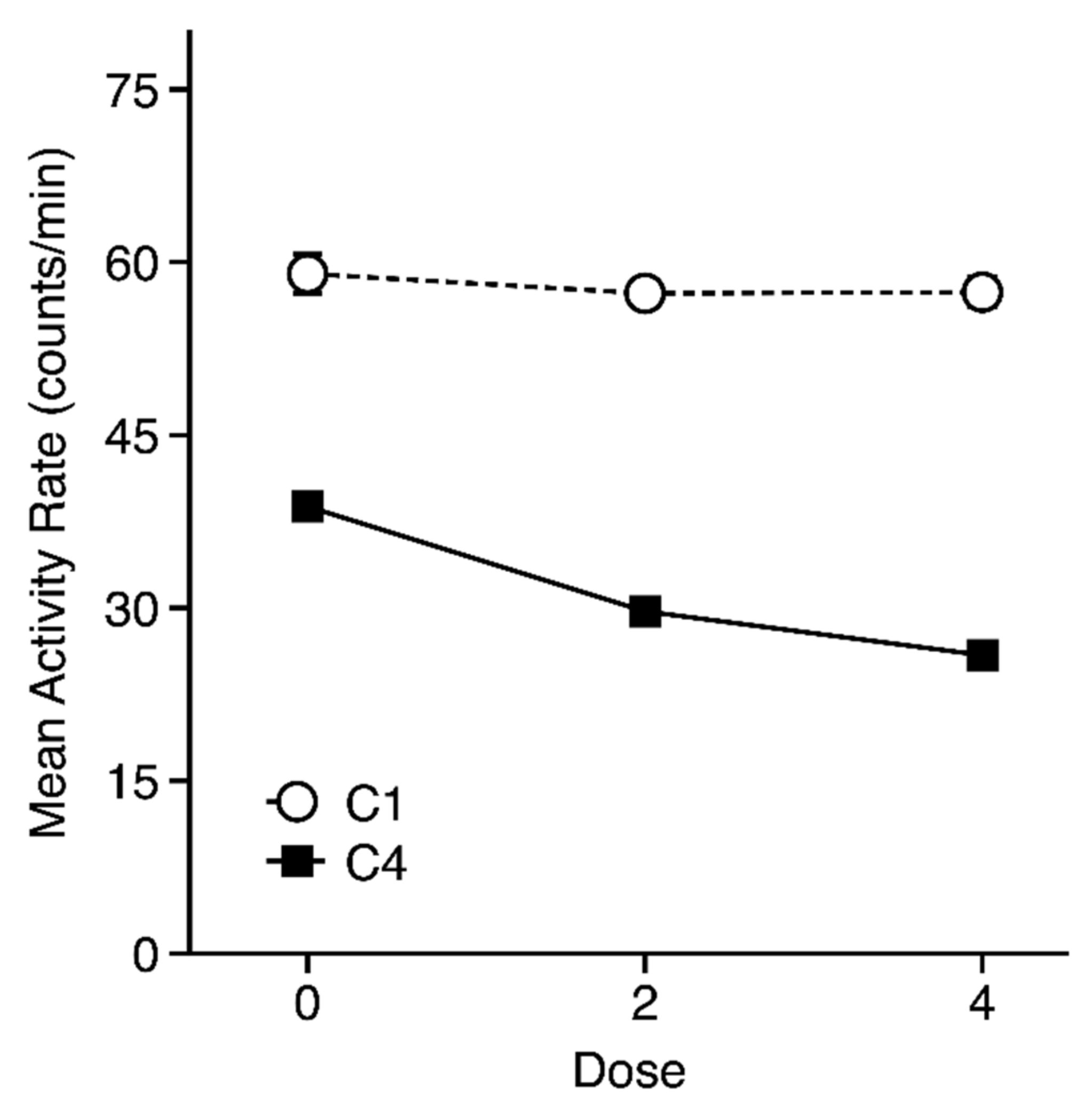
3.1. Habituation Session Activity
3.2. Conditioning Trial Activity
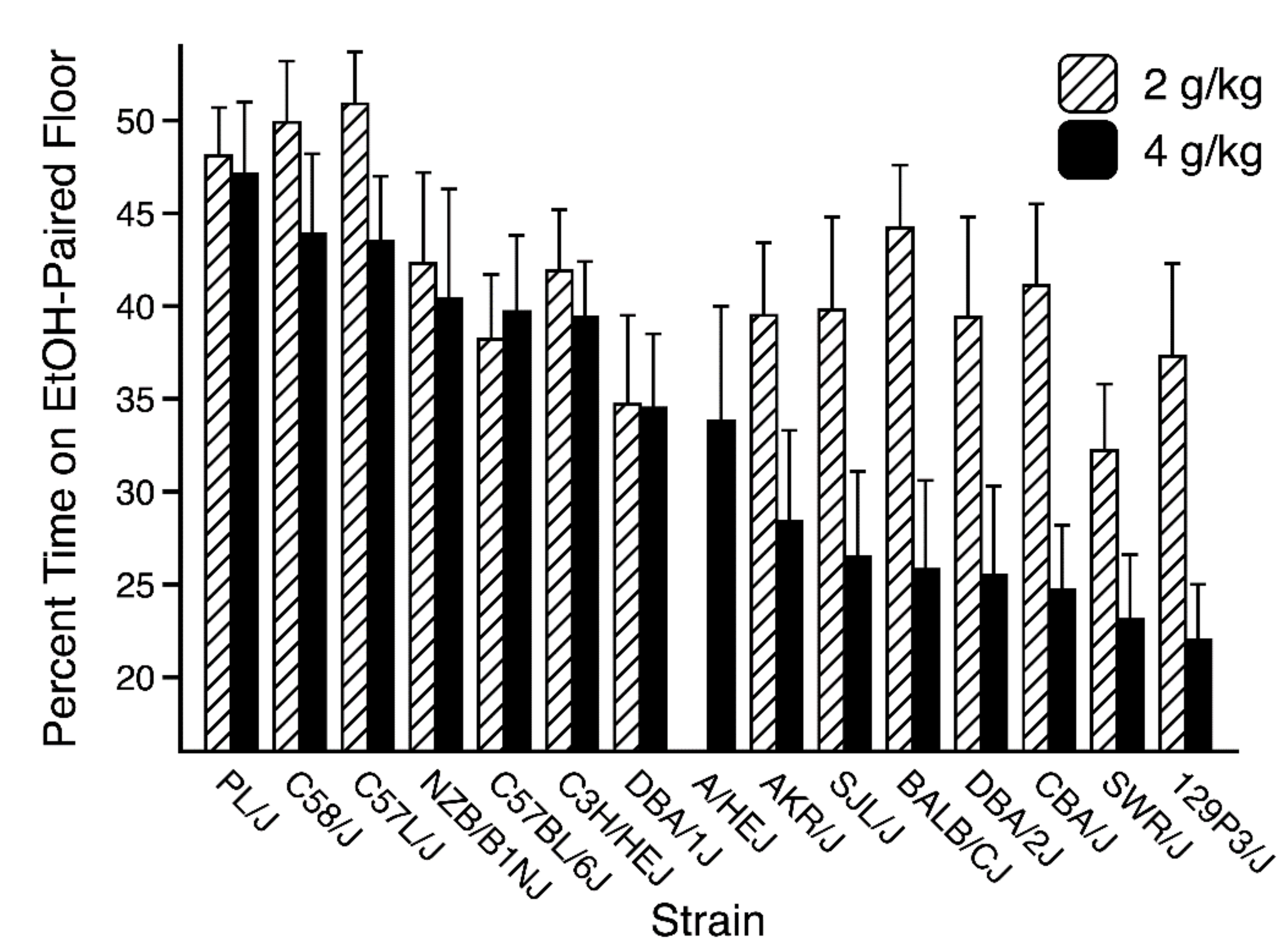
3.3. Place Aversion Test
3.4. Genetic Correlations
3.4.1. Activity rates
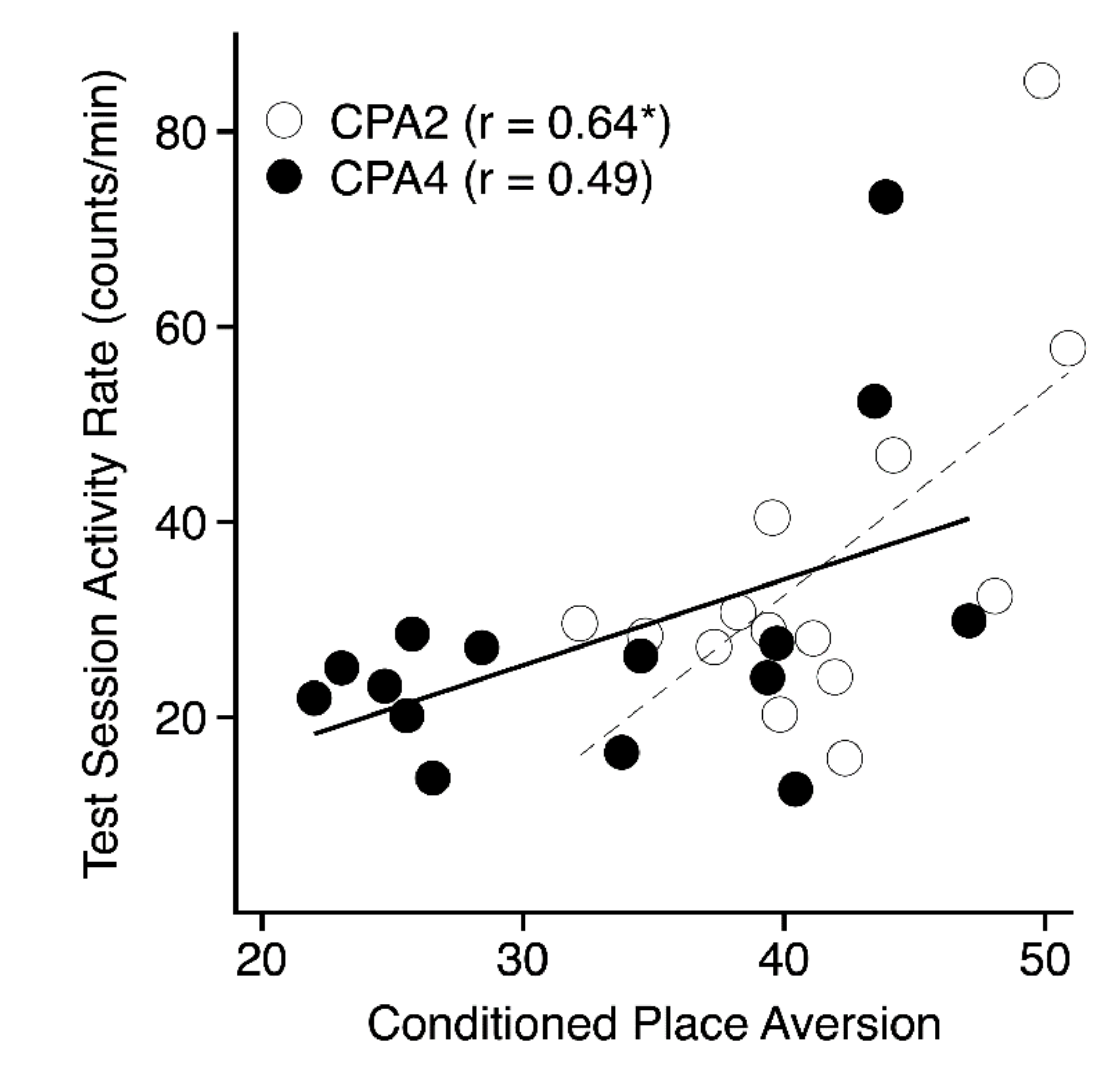
3.4.2. Conditioned Aversion Test
3.4.3. Other Ethanol Phenotypes
4. Discussion
4.1. Conditioned Place Aversion
4.2. Activity Phenotypes
4.3. Genetic Correlations with Other Ethanol Phenotypes
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Phillips, T.J.; Crabbe, J.C. Behavioral studies of genetic differences in alcohol action. In The Genetic Basis of Alcohol and Drug Actions; Crabbe, J.C., Harris, R.A., Eds.; Plenum Press: New York, NY, USA, 1991; pp. 25–104. [Google Scholar]
- Crabbe, J.C.; Gallaher, E.S.; Phillips, T.J.; Belknap, J.K. Genetic determinants of sensitivity to ethanol in inbred mice. Behav. Neurosci. 1994, 108, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Begleiter, H.; Kissin, B. The Genetics of Alcoholism; Oxford: New York, NY, USA, 1995. [Google Scholar]
- Elmer, G.I.; Meisch, R.A.; George, F.R. Differential concentration-response curves for oral ethanol self-administration in C57BL/6J and BALB/cJ mice. Alcohol 1987, 4, 63–68. [Google Scholar] [CrossRef]
- Belknap, J.K.; Crabbe, J.C.; Young, E.R. Voluntary consumption of ethanol in 15 inbred mouse strains. Psychopharm 1993, 112, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Risinger, F.O.; Freeman, P.A.; Greengard, P.; Fienberg, A.A. Motivational effects of ethanol in DARPP-32 knock-out mice. J. Neurosci. 2001, 21, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, N.; Crabbe, J.C.; Ford, M.M.; Murillo, A.; Finn, D.A. Voluntary ethanol consumption in 22 inbred mouse strains. Alcohol 2008, 42, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Crabbe, J.C.; Young, E.R.; Kosobud, A. Genetic correlations with ethanol withdrawal severity. Pharmacol. Biochem. Behav. 1983, 18 (Suppl. 1), 541–547. [Google Scholar] [CrossRef]
- Metten, P.; Crabbe, J.C. Common genetic determinants of severity of acute withdrawal from ethanol, pentobarbital and diazepam in inbred mice. Behav. Pharm. 1994, 5, 533–547. [Google Scholar] [CrossRef][Green Version]
- Metten, P.; Crabbe, J.C. Alcohol withdrawal severity in inbred mouse (Mus musculus) strains. Behav. Neurosci. 2005, 119, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L. Localization of genes influencing ethanol-induced conditioned place preference and locomotor activity in BXD recombinant inbred mice. Psychopharm 1995, 120, 28–41. [Google Scholar] [CrossRef]
- Risinger, F.; Cunningham, C.L. Ethanol-induced conditioned taste aversion in BXD recombinant inbred mice. Alcohol. Clin. Exp. Res. 1998, 22, 1234–1244. [Google Scholar] [CrossRef]
- Broadbent, J.; Muccino, K.J.; Cunningham, C.L. Ethanol-induced conditioned taste aversion in 15 inbred mouse strains. Behav. Neurosci. 2002, 116, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L. Genetic relationship between ethanol-induced conditioned place preference and other ethanol phenotypes in 15 inbred mouse strains. Behav. Neurosci. 2014, 128, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Roberts, A.J.; Schulteis, G.; Parsons, L.H.; Heyser, C.J.; Hyytiä, P.; Merlo-Pich, E.; Weiss, F. Neurocircuitry targets in ethanol reward and dependence. Alcohol. Clin. Exp. Res. 1998, 22, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Tabakoff, B.; Hoffman, P. A neurobiological theory of alcoholism. In Theories on Alcoholism; Chaudron, C.D., Wilkinson, D.A., Eds.; Addiction Research Foundation: Toronto, ON, Canada, 1988; pp. 29–72. [Google Scholar]
- Green, A.S.; Grahame, N.J. Ethanol drinking in rodents: Is free-choice drinking related to the reinforcing effects of ethanol? Alcohol 2008, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Gremel, C.M.; Groblewski, P.A. Genetic influences on conditioned taste aversion. In Conditioned Taste Aversion: Behavioral and Neural Processes; Reilly, S., Schachtman, T.R., Eds.; Oxford: New York, NY, USA, 2009; pp. 387–421. [Google Scholar]
- Cunningham, C.L.; Okorn, D.; Howard, C. Interstimulus interval determines whether ethanol produces conditioned place preference or aversion in mice. Anim. Learn. Behav. 1997, 25, 31–42. [Google Scholar] [CrossRef]
- Cunningham, C.L.; Henderson, C.M.; Bormann, N.M. Extinction of ethanol-induced conditioned place preference and conditioned place aversion: Effects of naloxone. Psychopharm 1998, 139, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Henderson, C.M. Ethanol-induced conditioned place aversion in mice. Behav. Pharmacol. 2000, 11, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Tull, L.E.; Rindal, K.E.; Meyer, P.J. Distal and proximal pre-exposure to ethanol in the place conditioning task: Tolerance to aversive effect, sensitization to activating effect, but no change in rewarding effect. Psychopharm 2002, 160, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Smith, R.; McMullin, C. Competition between ethanol-induced reward and aversion in place conditioning. Anim. Learn. Behav. 2003, 31, 273–280. [Google Scholar] [CrossRef]
- Bechtholt, A.J.; Gremel, C.M.; Cunningham, C.L. Handling blocks expression of conditioned place aversion but not conditioned place preference produced by ethanol in mice. Pharmacol. Biochem. Behav. 2004, 79, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Crabbe, J.C.; Phillips, T.J.; Kosobud, A.; Belknap, J.K. Estimation of genetic correlation: Interpretation of experiments using selectively bred and inbred animals. Alcohol. Clin. Exp. Res. 1990, 14, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Gremel, C.M.; Groblewski, P.A. Drug-induced conditioned place preference and aversion in mice. Nat. Protoc. 2006, 1, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Niehus, D.R.; Malott, D.H.; Prather, L.K. Genetic differences in the rewarding and activating effects of morphine and ethanol. Psychopharm 1992, 107, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Linakis, J.G.; Cunningham, C.L. Effects of concentration of ethanol injected intraperitoneally on taste aversion, body temperature, and activity. Psychopharm 1979, 64, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Ferree, N.K.; Howard, M.A. Apparatus bias and place conditioning with ethanol in mice. Psychopharm 2003, 170, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L. Pavlovian drug conditioning. In Methods in Behavioral Pharmacology; van Haaren, F., Ed.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 349–381. [Google Scholar]
- Cunningham, C.L.; Groblewski, P.A.; Voorhees, C.M. Place conditioning. In Animal Models of Drug Addiction, Neuromethods; Olmstead, M.C., Ed.; Humana Press: New York, NY, USA, 2011; Volume 53, pp. 167–189. [Google Scholar] [CrossRef]
- Metten, P.; Crabbe, J.C. Genetic determinants of severity of acute withdrawal from diazepam in mice: Commonality with ethanol and pentobarbital. Pharmacol. Biochem. Behav. 1999, 63, 473–479. [Google Scholar] [CrossRef]
- Gremel, C.M.; Cunningham, C.L. Role of test activity in ethanol-induced disruption of place preference expression in mice. Psychopharm 2007, 191, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Vezina, P.; Stewart, J. Morphine conditioned place preference and locomotion: The effect of confinement during training. Psychopharm 1987, 93, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Noble, D. Conditioned activation induced by ethanol: Role in sensitization and conditioned place preference. Pharmacol. Biochem. Behav. 1992, 43, 307–313. [Google Scholar] [CrossRef]
- Wong, A.A.; Brown, R.E. Visual detection, pattern discrimination and visual acuity in 14 strains of mice. Genes Brain Behav. 2006, 5, 389–403. [Google Scholar] [CrossRef]
- Brown, R.E.; Wong, A.A. The influence of visual ability on learning and memory performance in 13 strains of mice. Learn. Mem. 2007, 14, 134–144. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, T.P.; Savoie, V.; Brown, R.E. Learning, memory and search strategies of inbred mouse strains with different visual abilities in the Barnes maze. Behav. Brain Res. 2011, 216, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Portugal, G.S.; Wilkinson, D.S.; Kenney, J.W.; Sullivan, C.; Gould, T.J. Strain-dependent effects of acute, chronic, and withdrawal from chronic nicotine on fear conditioning. Behav. Genet. 2011, 42, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Pinhas, A.; Aviel, M.; Koen, M.; Gurgov, S.; Acosta, V.; Israel, M.; Kakuriev, L.; Guskova, E.; Fuzailov, I.; Touzani, K.; et al. Strain differences in sucrose- and fructose-conditioned flavor preferences in mice. Physiol. Behav. 2012, 105, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Ishiwatari, Y.; Bachmanov, A.A. NaCl taste thresholds in 13 inbred mouse strains. Chem. Senses 2012, 37, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Metten, P.; Phillips, T.J.; Crabbe, J.C.; Tarantino, L.M.; McClearn, G.E.; Plomin, R.; Erwin, V.G.; Belknap, J.K. High genetic susceptibility to ethanol withdrawal predicts low ethanol consumption. Mamm. Genome 1998, 9, 983–990. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Phillips, T.J.; Cunningham, C.L.; Belknap, J.K. Genetic determinants of ethanol reinforcement. Ann. N. Y. Acad. Sci. 1992, 654, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Chester, J.; Risinger, F.; Cunningham, C.L. Ethanol reward and aversion in mice bred for sensitivity to ethanol withdrawal. Alcohol. Clin. Exp. Res. 1998, 22, 468–473. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotype * | GT-0 | PDT-0 | PDT-2 | PDT-4 | TACT-0 | TACT-2 | TACT-4 |
|---|---|---|---|---|---|---|---|
| GT-0 | -- | -- | -- | -- | -- | -- | -- |
| PDT-0 | 0.14 | -- | -- | -- | -- | -- | -- |
| PDT-2 | 0.11 | 0.09 | -- | -- | -- | -- | -- |
| PDT-4 | −0.12 | 0.17 | 0.67b | -- | -- | -- | -- |
| TACT-0 | 0.35 | 0.44 | 0.54a | 0.25 | -- | -- | -- |
| TACT-2 | 0.38 | 0.40 | 0.64a | 0.35 | 0.96c | -- | -- |
| TACT-4 | 0.34 | 0.42 | 0.65a | 0.49 | 0.88c | 0.96c | -- |
| HAB-0 | 0.14 | 0.33 | 0.35 | 0.32 | 0.76b | 0.82c | 0.81c |
| HAB-2 | 0.21 | 0.29 | 0.37 | 0.30 | 0.75b | 0.83c | 0.83c |
| HAB-4 | 0.13 | 0.32 | 0.35 | 0.30 | 0.71b | 0.79c | 0.78c |
| C4-0 | 0.44 | 0.44 | 0.56a | 0.14 | 0.86c | 0.82c | 0.71b |
| C4-2 | 0.39 | 0.41 | 0.59a | 0.18 | 0.73b | 0.78c | 0.69b |
| C4-4 | 0.43 | 0.49 | 0.60a | 0.26 | 0.79c | 0.85c | 0.81c |
| (C4-HAB)-0 | 0.11 | −0.12 | −0.07 | −0.30 | −0.37 | −0.47 | −0.54a |
| (C4-HAB)-2 | −0.02 | −0.11 | −0.11 | −0.26 | −0.50 | −0.57a | −0.62a |
| (C4-HAB)-4 | 0.09 | −0.12 | −0.11 | −0.23 | −0.45 | −0.54a | −0.52a |
| c = p < 0.001, b = p < 0.01, a = p < 0.05; significant correlations are shown in boldface | |||||||
| Phenotype | n | CPA PDT-2 | CPA PDT-4 |
|---|---|---|---|
| Belknap et al. (1993) | |||
| 3% ethanol intake | 12/13 | 0.27 | 0.52 * |
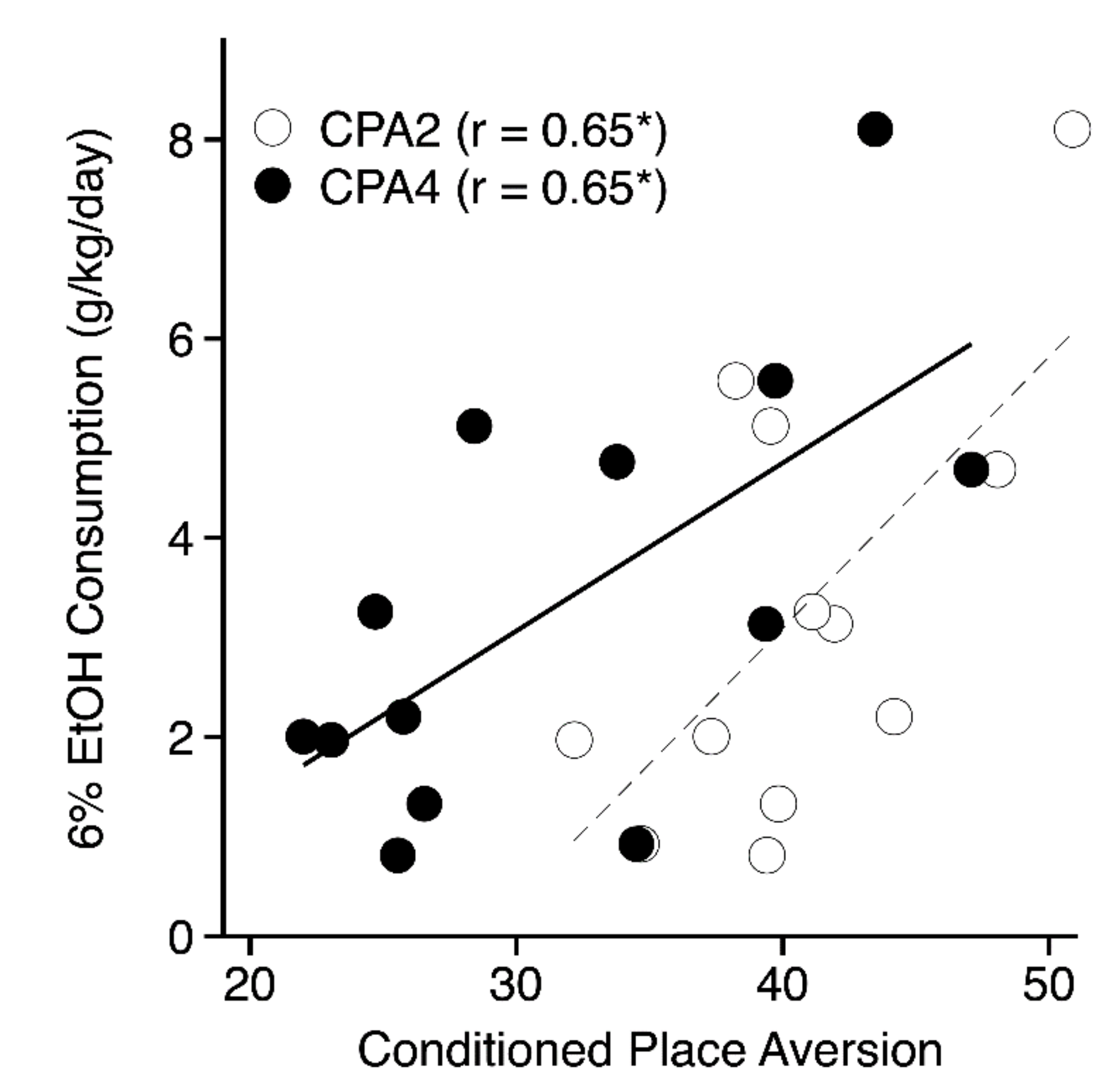
| 6% ethanol intake | 12/13 | 0.65a | 0.65a |
| 10% ethanol intake | 12/13 | 0.41 | 0.30 |
| 3% ethanol preference ratio | 12/13 | 0.08 | 0.44 |
| 6% ethanol preference ratio | 12/13 | 0.52 * | 0.59a |
| 10% ethanol preference ratio | 12/13 | 0.39 | 0.32 |
| Broadbent et al. (2002) | |||
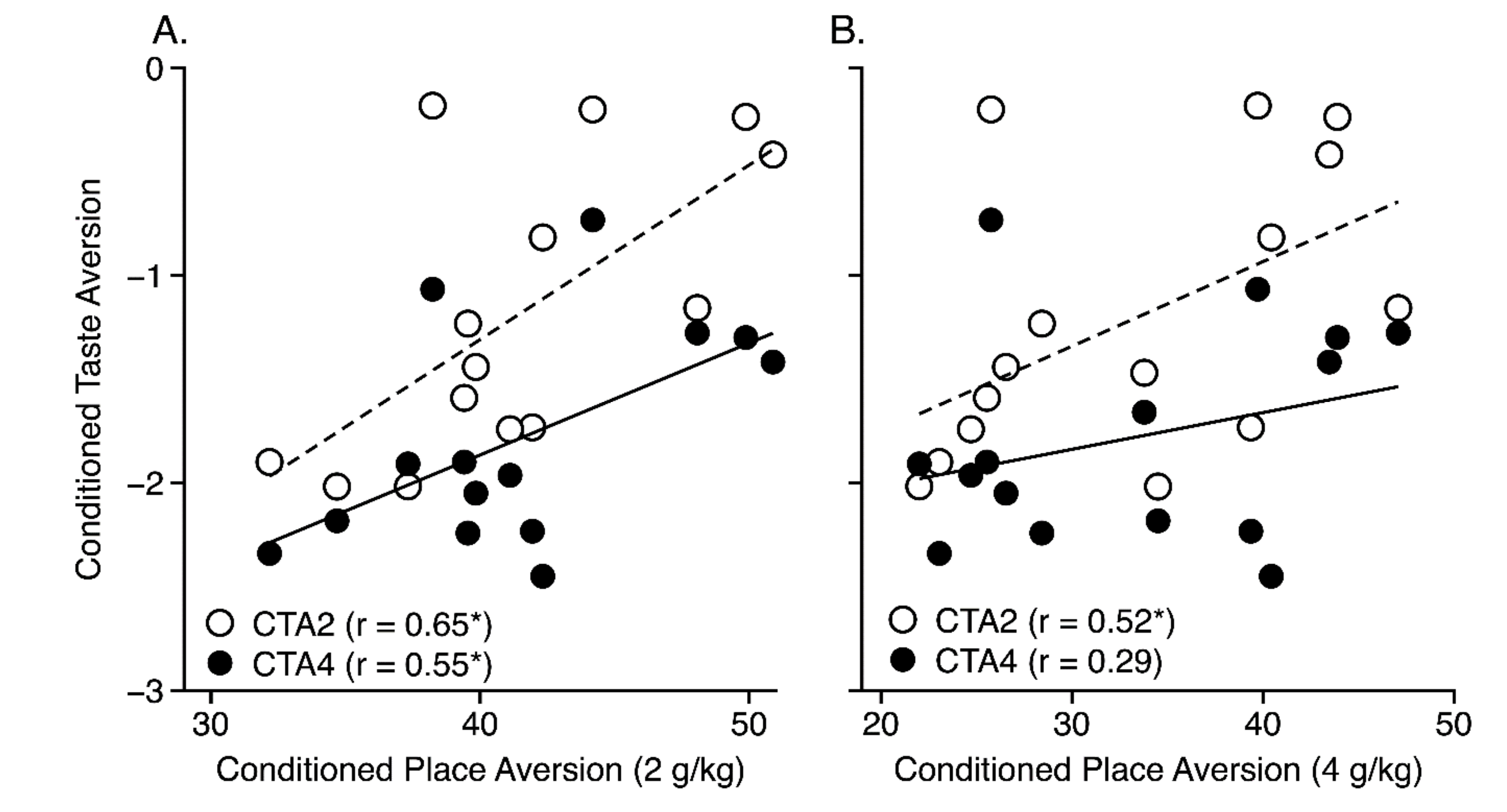
| Conditioned Taste Aversion (2 g/kg) | 14/15 | 0.65a | 0.52a |
| Conditioned Taste Aversion (4 g/kg) | 14/15 | 0.55a | 0.29 |
| Crabbe et al. (1994) | |||
| Blood Ethanol Concentration (1 g/kg) | 12/13 | 0.19 | 0.32 |
| Blood Ethanol Concentration (2 g/kg) | 12/13 | −0.25 | −0.15 |
| Blood Ethanol Concentration (3 g/kg) | 12/13 | 0.18 | 0.45 |
| Blood Ethanol Concentration (4 g/kg) | 12/13 | 0.31 | 0.42 |
| Cunningham (2014) | |||
| Conditioned Place Preference (2 g/kg) | 14/15 | −0.27 | −0.09 |
| Conditioned Place Preference (4 g/kg) | 14/15 | −0.17 | −0.09 |
| Metten & Crabbe (1994) | |||
| Acute Ethanol Withdrawal AREA (uncorrected) | 12/13 | −0.03 | −0.53 * |
| Acute Ethanol Withdrawal Peak (uncorrected) | 12/13 | 0.02 | −0.48 * |
| Acute Diazepam Withdrawal Peak (uncorrected) | 12/13 | −0.29 | −0.66a |
| Metten & Crabbe (1999) | |||
| Acute Diazepam Withdrawal Peak (uncorrected) | 12/13 | −0.30 | −0.63a |
| Metten & Crabbe (2005) | |||
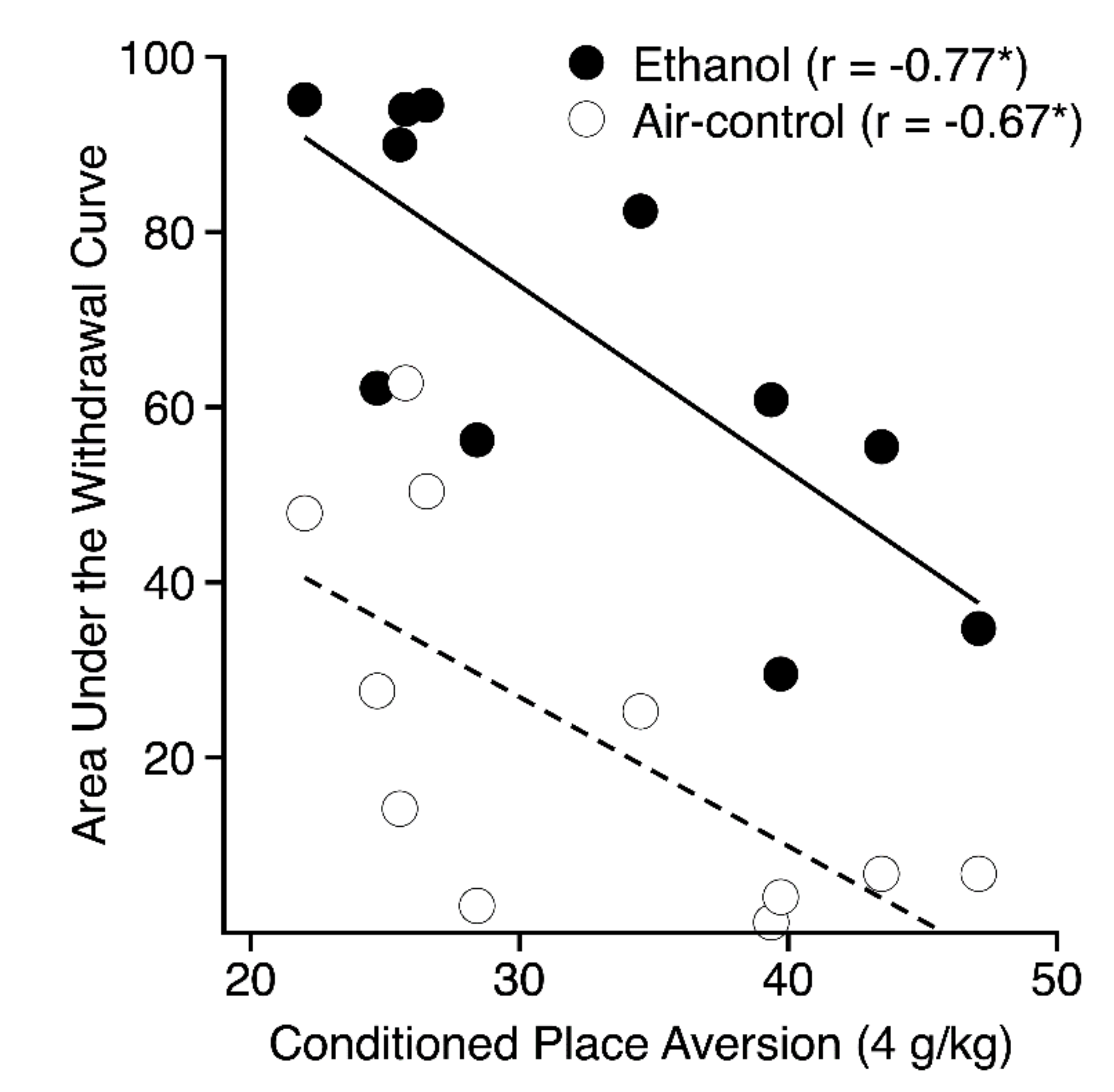
| Air Control AREA 25 | 11 | −0.20 | −0.67a |
| Chronic Ethanol Withdrawal AREA 25 (uncorrected) | 11 | −0.37 | −0.77a |
| Chronic Ethanol Withdrawal (Δ AREA25) | 11 | −0.29 | −0.24 |
| Reference | Phenotype | n | CPA PDT-2 * | CPA PDT-4 * |
|---|---|---|---|---|
| Wong & Brown (2006) | ||||
| Visual detection task (D8 % correct) | 6 | −0.69 | −0.05 | |
| Visual detection task (D8 latency) | 6 | 0.86a | −0.22 | |
| Pattern discrimination task (D8 % correct) | 6 | −0.62 | −0.16 | |
| Pattern discrimination task (D8 latency) | 6 | 0.92a | −0.22 | |
| Swim speed (cm/sec) | 6 | −0.55 | −0.10 | |
| Brown & Wong (2007) | ||||
| Visual detection task (D8 % correct) | 5 | −0.58 | −0.29 | |
| Pattern discrimination task (D8 % correct) | 5 | −0.46 | −0.41 | |
| Reversal latency in MWM (sec) | 5 | 0.81 | 0.09 | |
| Reversal swim distance in MWM (cm) | 5 | 0.92a | 0.10 | |
| Swim speed (cm/sec) | 5 | −0.17 | −0.44 | |
| Probe trial in MWM (% time in correct quadrant) | 5 | −0.77 | −0.02 | |
| Probe trial in MWM (annulus crossings) | 5 | −0.85 | 0.31 | |
| Latency to visible platform in MWM (sec) | 5 | 0.79 | 0.04 | |
| Conditioned odor preference task (% CS+ digging) | 5 | 0.10 | 0.68 | |
| Latency to fall on the Rotarod-males (sec) | 5 | −0.28 | 0.26 | |
| O’Leary et al. (2011) | ||||
| Acquisition latency (s) | 6 | 0.28 | −0.46 | |
| Reversal latency (s) | 6 | −0.01 | −0.10 | |
| Reversal difference latency | 6 | −0.71 | 0.41 | |
| Number of acquisition errors | 6 | −0.02 | −0.37 | |
| Number of reversal errors | 6 | 0.06 | −0.06 | |
| Reversal difference errors | 6 | −0.81 | 0.39 | |
| % time correct quadrant | 6 | −0.67 | 0.52 | |
| Portugal et al. (2011) | ||||
| Contextual Fear Conditioning (% immobility) | 5 | 0.23 | −0.06 | |
| Cued Fear Conditioning (% immobility) | 5 | 0.66 | 0.33 | |
| Pinhas et al. (2012) | ||||
| CS+Sucrose (% CS+ intake) | 7 | −0.16 | −0.63 | |
| CS+Fructose (% CS+ intake) | 7 | 0.23 | −0.09 | |
| CS+Saccharin (% CS+ intake) | 7 | 0.11 | −0.31 | |
| Ishiwatari & Bachmanov (2012) | ||||
| CS intake (mL) Difference (Trial 1–Trial 2) | 6 | 0.66 | 0.41 | |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cunningham, C.L. Genetic Relationships Between Ethanol-Induced Conditioned Place Aversion and Other Ethanol Phenotypes in 15 Inbred Mouse Strains. Brain Sci. 2019, 9, 209. https://doi.org/10.3390/brainsci9080209
Cunningham CL. Genetic Relationships Between Ethanol-Induced Conditioned Place Aversion and Other Ethanol Phenotypes in 15 Inbred Mouse Strains. Brain Sciences. 2019; 9(8):209. https://doi.org/10.3390/brainsci9080209
Chicago/Turabian StyleCunningham, Christopher L. 2019. "Genetic Relationships Between Ethanol-Induced Conditioned Place Aversion and Other Ethanol Phenotypes in 15 Inbred Mouse Strains" Brain Sciences 9, no. 8: 209. https://doi.org/10.3390/brainsci9080209
APA StyleCunningham, C. L. (2019). Genetic Relationships Between Ethanol-Induced Conditioned Place Aversion and Other Ethanol Phenotypes in 15 Inbred Mouse Strains. Brain Sciences, 9(8), 209. https://doi.org/10.3390/brainsci9080209