
Topography of Slow Sigma Power during Sleep is Associated with Processing Speed in Preschool Children


Abstract
:1. Introduction
2. Experimental Section
2.1. Participants
2.2. Procedure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sleep Variables | Mean ± SD | Min | Max |
|---|---|---|---|
| Time in bed (min) | 609.2 ± 67.4 | 483.7 | 647.0 |
| Total sleep time (min) | 545.3 ± 77.2 | 390.0 | 619.7 |
| Stage 1 (%) | 1.1 ± 0.3 | 0.7 | 1.6 |
| Stage 2 (%) | 35.4 ± 4.6 | 27.3 | 44.2 |
| SWS (%) | 31.0 ± 7.3 | 21.1 | 41.6 |
| REM sleep (%) | 32.5 ± 6.1 | 21.8 | 41.8 |
| NREM sleep (%) | 66.4 ± 6.1 | 48.2 | 68.3 |
| Sleep cycle duration (min) | 83.3 ± 19.3 | 50.5 | 120.4 |
2.3. Measures
2.3.1. Processing Speed

2.3.2. Processing and Analysis
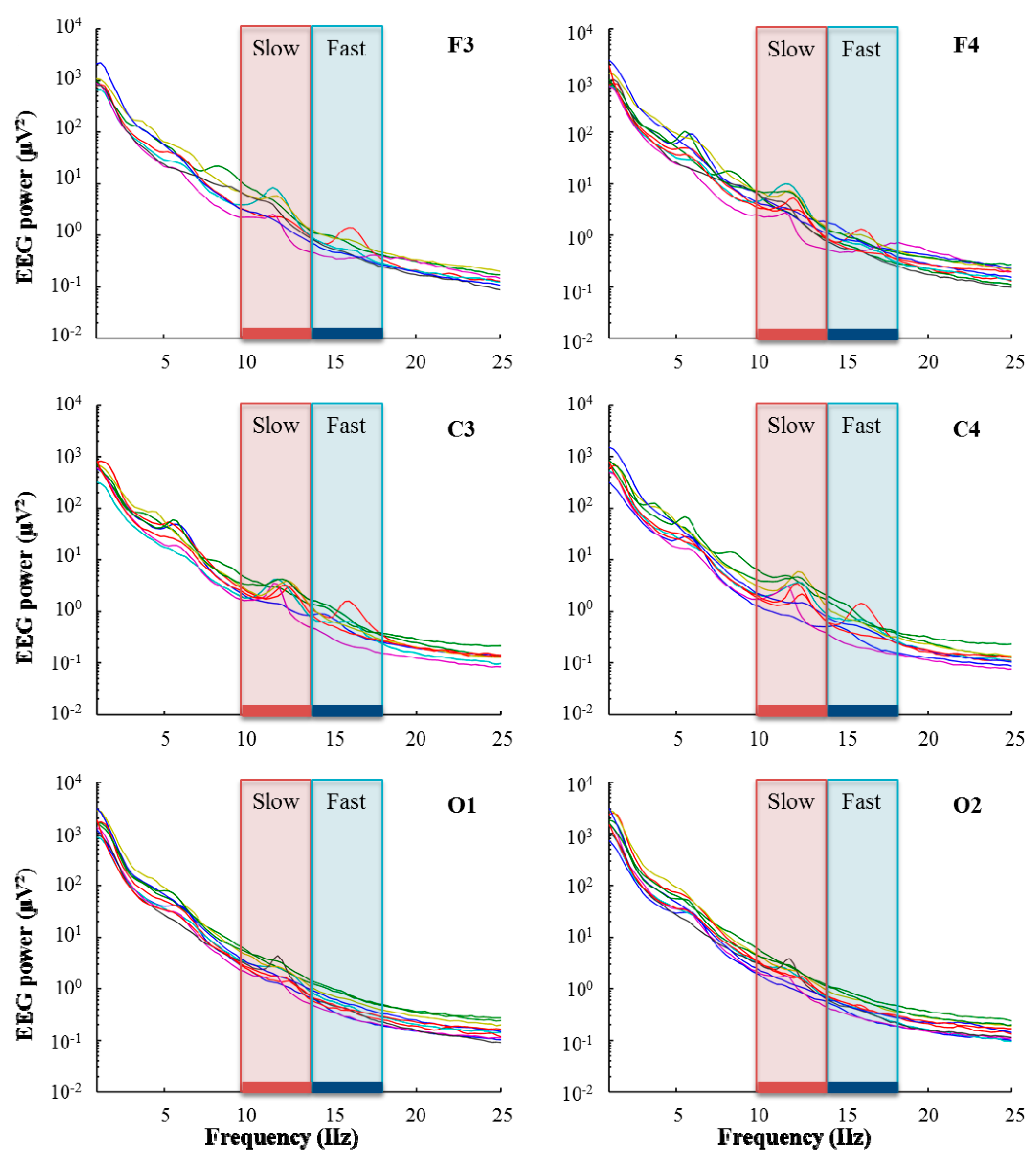
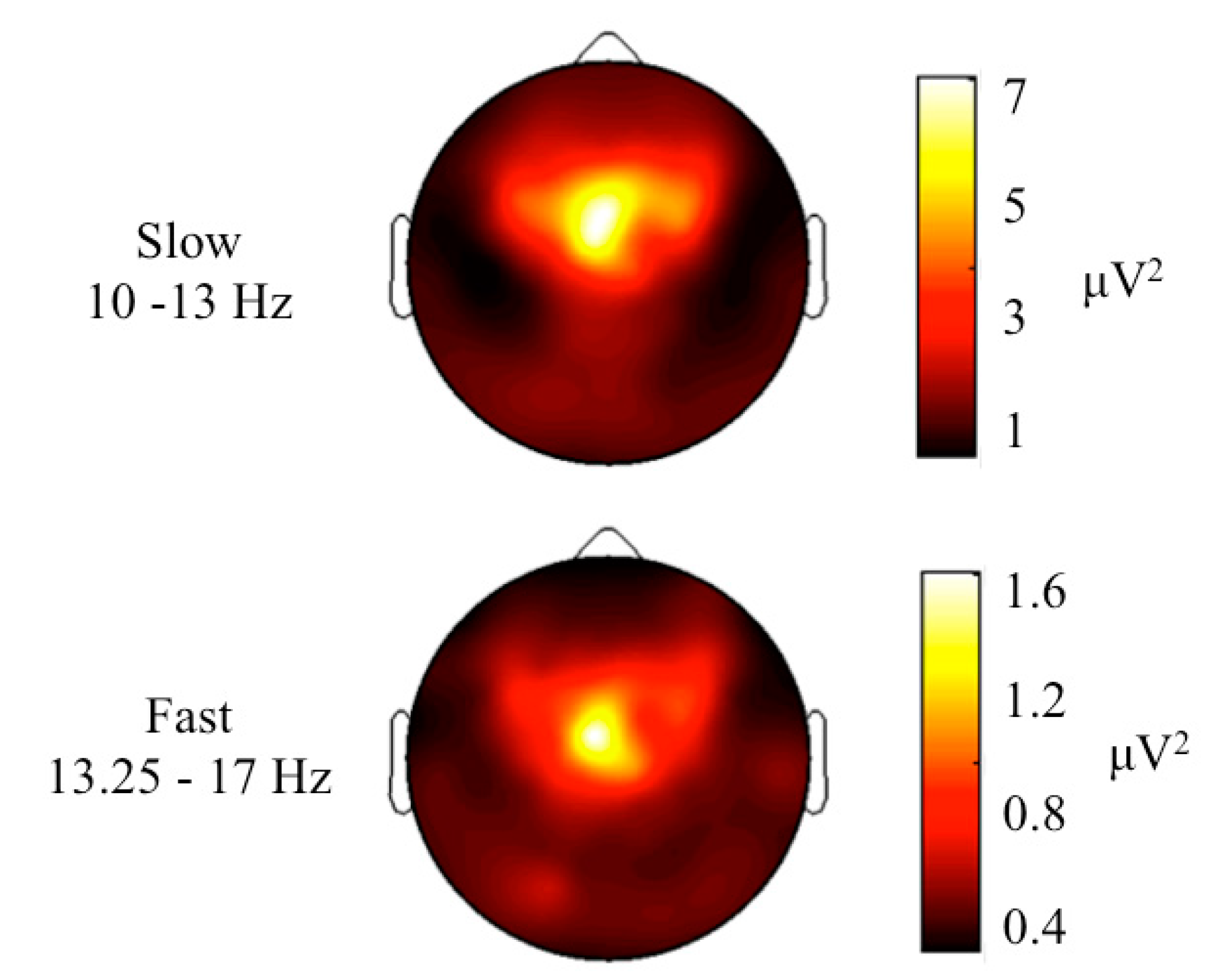
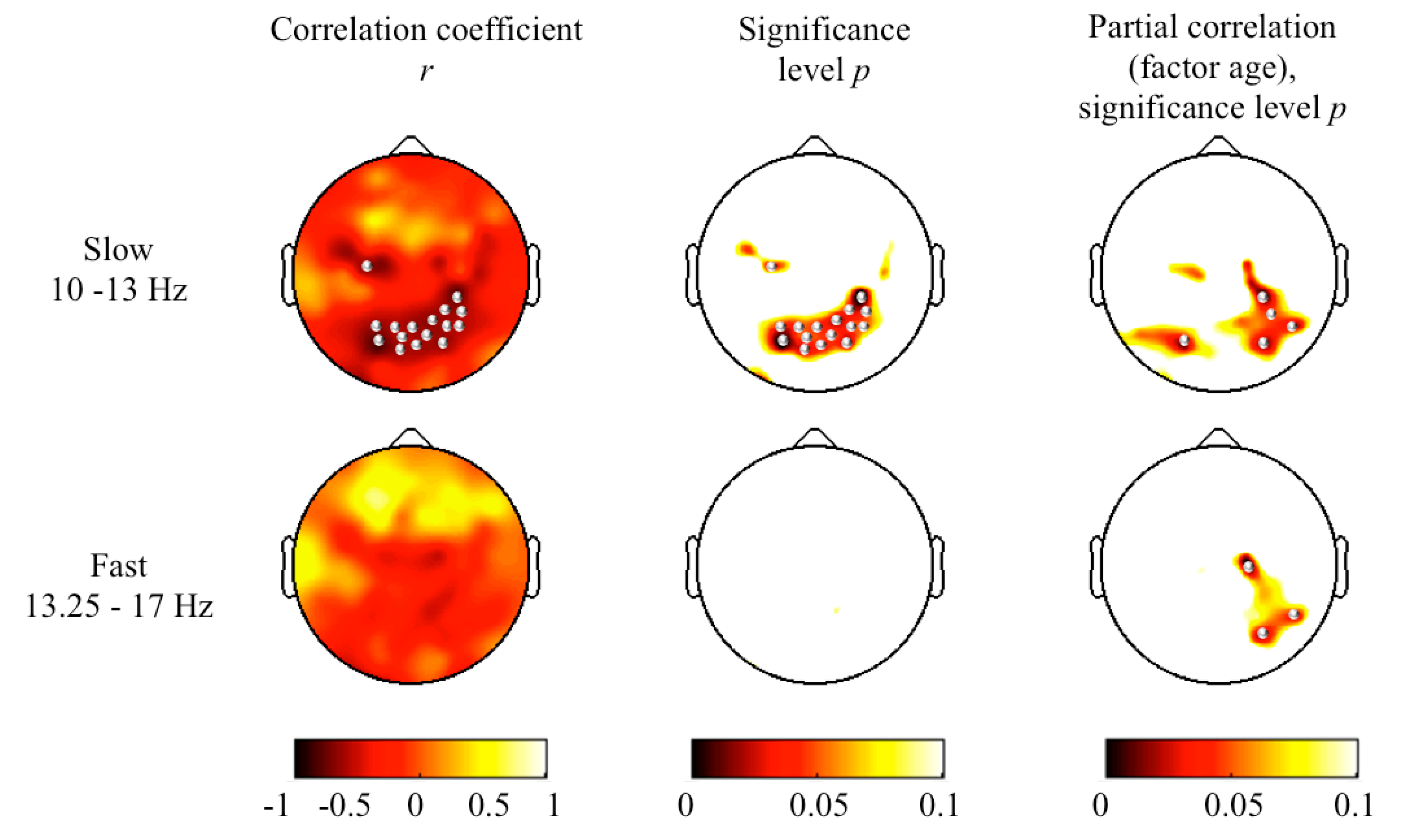
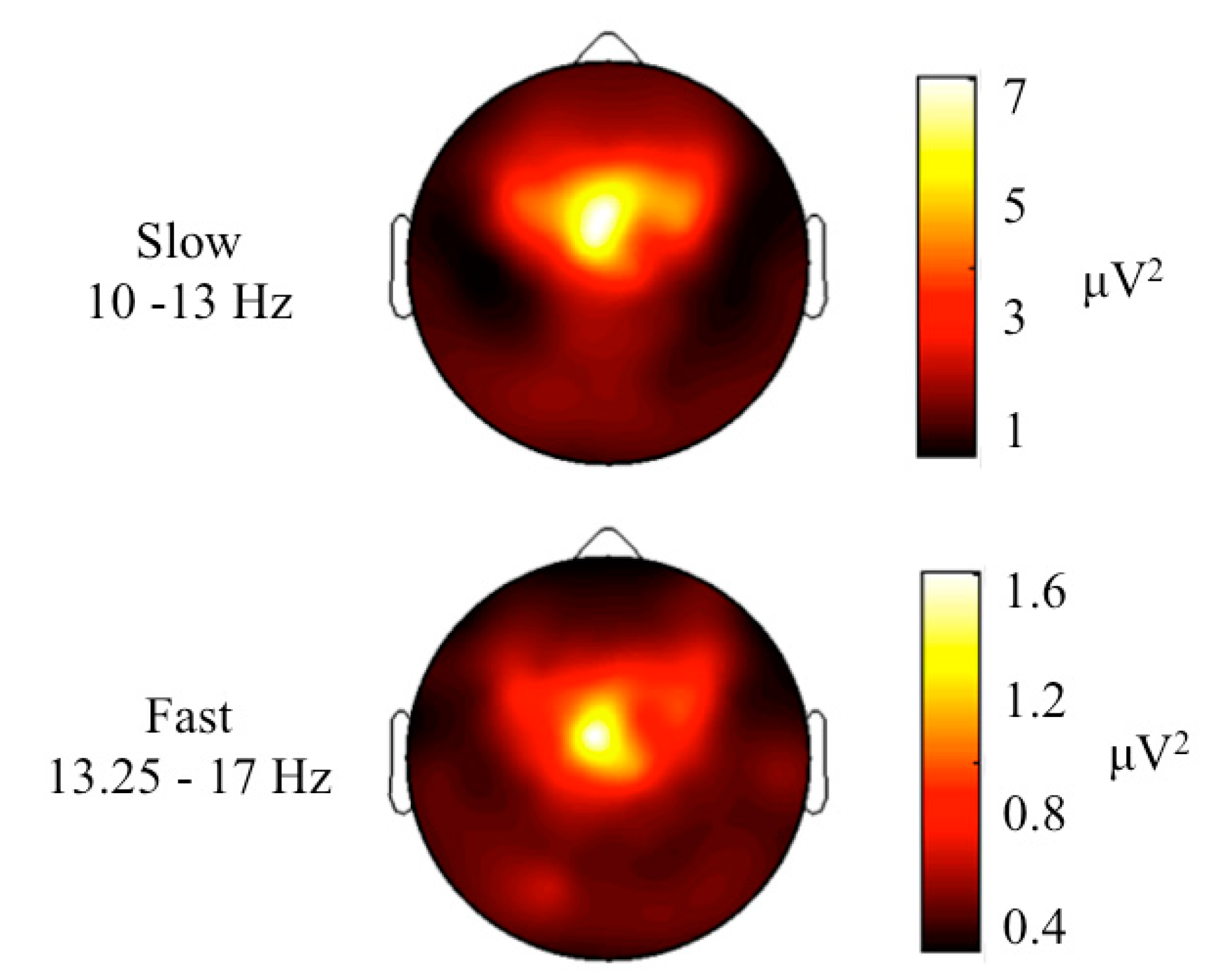
3. Results and Discussion
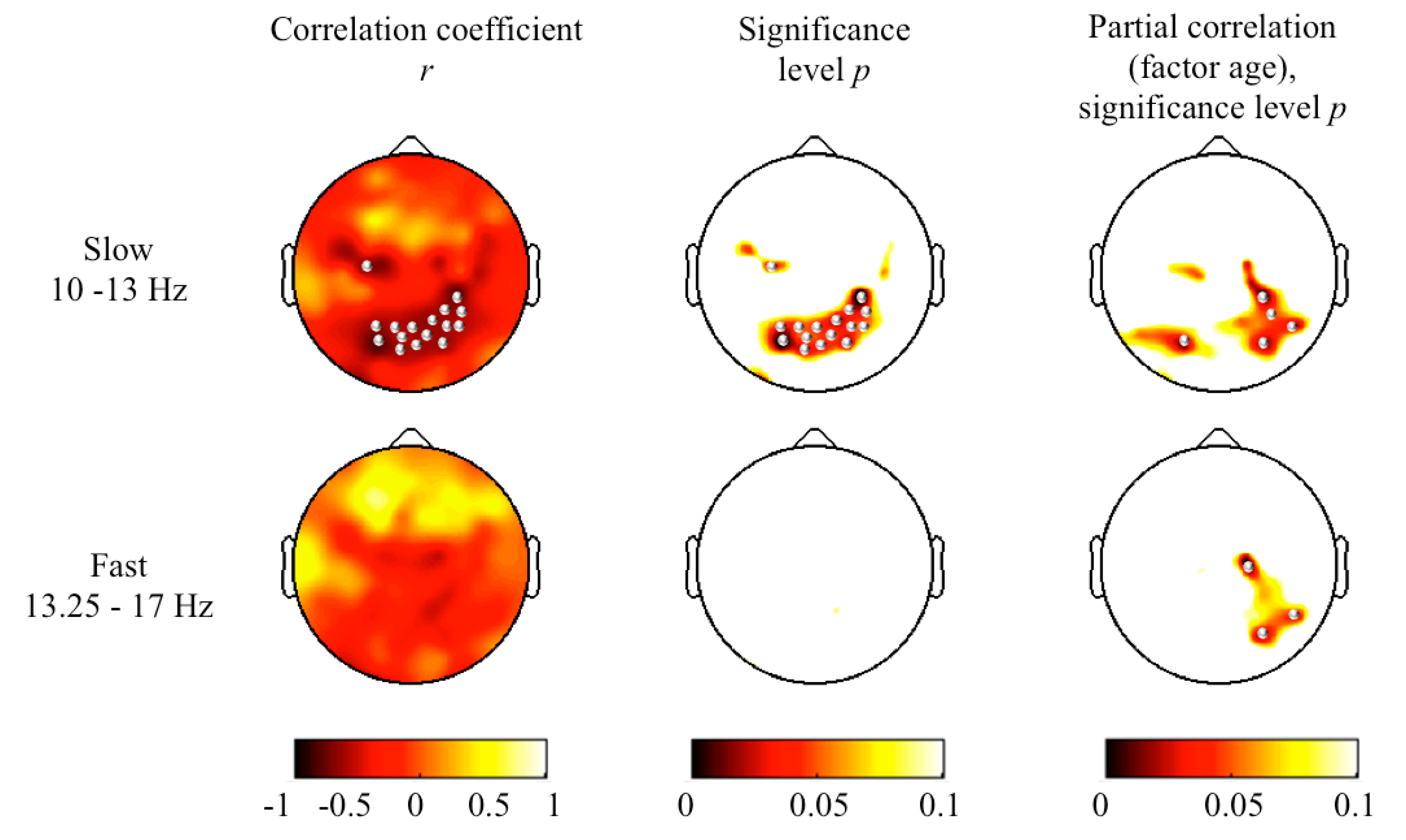
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kail, R.; Wolters, C.A.; Yu, S.L.; Hagen, J.W. Brief report: Speed of information processing in children with insulin-dependent diabetes mellitus. J. Pediatr. Psychol. 2000, 25, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Barrouillet, P.; Bernardin, S.; Camos, V. Time constraints and resource sharing in adults’ working memory spans. J. Exp. Psychol. Gen. 2004, 133, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Towse, J.N.; Hitch, G.J.; Hutton, U. A reevaluation of working memory capacity in children. J. Mem. Lang. 1998, 39, 195–217. [Google Scholar] [CrossRef]
- Fry, A.F.; Hale, S. Processing speed, working memory, and fluid intelligence: Evidence for a developmental cascade. Psychol. Sci. 1996, 7, 237–241. [Google Scholar] [CrossRef]
- Fry, A.F.; Hale, S. Relationships among processing speed, working memory, and fluid intelligence in children. Biol. Psychol. 2000, 54, 1–34. [Google Scholar] [CrossRef]
- Jensen, A.R. Why is reaction time correlated with psychometric g? Curr. Direct. Psychol. Sci. 1993, 2, 53–56. [Google Scholar] [CrossRef]
- Salthouse, T.A. The processing-speed theory of adult age differences in cognition. Psychol. Rev. 1996, 103, 403–428. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.T.; Vernon, P.A. Intelligence, reaction time, and working memory in 4- to 6-year-old children. Intelligence 1996, 22, 155–190. [Google Scholar] [CrossRef]
- Schabus, M.; Dang-Vu, T.T.; Albouy, G.; Balteau, E.; Boly, M.; Carrier, J.; Darsaud, A.; Degueldre, C.; Desseilles, M.; Gais, S.; et al. Hemodynamic cerebral correlates of sleep spindles during human non-rapid eye movement sleep. Proc. Natl. Acad. Sci. USA 2007, 104, 13164–13169. [Google Scholar] [CrossRef] [PubMed]
- McAuley, T.; White, D.A. A latent variables examination of processing speed, response inhibition, and working memory during typical development. J. Exp. Child Psychol. 2011, 108, 453–468. [Google Scholar] [CrossRef] [PubMed]
- Nettelbeck, T.; Burns, N.R. Processing speed, working memory and reasoning ability from childhood to old age. Pers. Indiv. Differ. 2010, 48, 379–384. [Google Scholar] [CrossRef]
- Jung, R.E.; Haier, R.J. The parieto-frontal integration theory (P-FIT) of intelligence: Converging neuroimaging evidence. Behav. Brain Sci. 2007, 30, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Luders, E.; Narr, K.L.; Thompson, P.M.; Toga, A.W. Neuroanatomical correlates of intelligence. Intelligence 2009, 37, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, A.C.; Fink, A. Intelligence and neural efficiency. Neurosci. Biobehav. Rev. 2009, 33, 1004–1023. [Google Scholar] [CrossRef] [PubMed]
- Geiger, A.; Huber, R.; Kurth, S.; Ringli, M.; Jenni, O.G.; Achermann, P. The sleep EEG as a marker of intellectual ability in school age children. Sleep 2011, 34, 181–189. [Google Scholar] [PubMed]
- Astill, R.G.; Piantoni, G.; Raymann, R.J.; Vis, J.C.; Coppens, J.E.; Walker, M.P.; Stickgold, R.; van der Werf, Y.D.; van Someren, E.J. Sleep spindle and slow wave frequency reflect motor skill performance in primary school-age children. Front. Hum. Neurosci. 2014, 8, 910. [Google Scholar] [CrossRef] [PubMed]
- Lustenberger, C.; Maric, A.; Durr, R.; Achermann, P.; Huber, R. Triangular relationship between sleep spindle activity, general cognitive ability and the efficiency of declarative learning. PLoS ONE 2012, 7, e49561. [Google Scholar] [CrossRef] [PubMed]
- Fogel, S.M.; Nader, R.; Cote, K.A.; Smith, C.T. Sleep spindles and learning potential. Behav. Neurosci. 2007, 121, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Schabus, M.; Hoedlmoser, K.; Pecherstorfer, T.; Anderer, P.; Gruber, G.; Parapatics, S.; Sauter, C.; Kloesch, G.; Klimesch, W.; Saletu, B.; et al. Interindividual sleep spindle differences and their relation to learning-related enhancements. Brain Res. 2008, 1191, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Steriade, M. Grouping of brain rhythms in corticothalamic systems. Neuroscience 2006, 137, 1087–1106. [Google Scholar] [CrossRef] [PubMed]
- Rechtschaffen, A.; Kales, A. A Manual of Standardized Terminology, Techniques and Scoring System for Sleep Stages of Human Subjects; US Public Health Service, US Government Printing Office: Washington, DC, USA, 1968.
- Buchmann, A.; Dentico, D.; Peterson, M.J.; Riedner, B.A.; Sarasso, S.; Massimini, M.; Tononi, G.; Ferrarelli, F. Reduced mediodorsal thalamic volume and prefrontal cortical spindle activity in schizophrenia. NeuroImage 2014, 102, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Feld, G.B.; Born, J. Sleep EEG rhythms and system consolidation of memory. In Sleep Brain Activity; Elsevier Inc.: Amsterdam, The Netherlands, 2012; pp. 187–226. [Google Scholar]
- Shinomiya, S.; Nagata, K.; Takahashi, K.; Masumura, T. Development of sleep spindles in young children and adolescents. Clin. EEG 1999, 30, 39–43. [Google Scholar] [CrossRef]
- Tamaki, M.; Matsuoka, T.; Nittono, H.; Hori, T. Fast sleep spindle (13–15 Hz) activity correlates with sleep-dependent improvement in visuomotor performance. Sleep 2008, 31, 204–211. [Google Scholar] [PubMed]
- Werth, E.; Achermann, P.; Dijk, D.J.; Borbely, A.A. Spindle frequency activity in the sleep EEG: Individual differences and topographic distribution. Electroencephalogr. Clin. Neurophysiol. 1997, 103, 535–542. [Google Scholar] [CrossRef]
- Molle, M.; Bergmann, T.O.; Marshall, L.; Born, J. Fast and slow spindles during the sleep slow oscillation: Disparate coalescence and engagement in memory processing. Sleep 2011, 34, 1411–1421. [Google Scholar] [CrossRef] [PubMed]
- Andrillon, T.; Nir, Y.; Staba, R.J.; Ferrarelli, F.; Cirelli, C.; Tononi, G.; Fried, I. Sleep spindles in humans: Insights from intracranial EEG and unit recordings. J. Neurosci. 2011, 31, 17821–17834. [Google Scholar] [CrossRef] [PubMed]
- Dehghani, N.; Cash, S.S.; Halgren, E. Topographical frequency dynamics within EEG and MEG sleep spindles. Clin. Neurophysiol. 2011, 122, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Jenni, O.G.; Carskadon, M.A. Spectral analysis of the sleep electroencephalogram during adolescence. Sleep 2004, 27, 774–783. [Google Scholar] [PubMed]
- Tarokh, L.; Carskadon, M.A. Developmental changes in the human sleep EEG during early adolescence. Sleep 2010, 33, 801–809. [Google Scholar] [PubMed]
- Kurth, S.; Ringli, M.; Geiger, A.; LeBourgeois, M.; Jenni, O.G.; Huber, R. Mapping of cortical activity in the first two decades of life: A high-density sleep electroencephalogram study. J. Neurosci. 2010, 30, 13211–13219. [Google Scholar] [CrossRef] [PubMed]
- Tanguay, P.E.; Ornitz, E.M.; Kaplan, A.; Bozzo, E.S. Evolution of sleep spindles in childhood. Electroencephalogr. Clin. Neurophysiol. 1975, 38, 175–181. [Google Scholar] [CrossRef]
- Nader, R.S.; Smith, C.T. Correlations between adolescent processing speed and specific spindle frequencies. Front. Hum. Neurosci. 2015, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Geiger, A.; Huber, R.; Kurth, S.; Ringli, M.; Achermann, P.; Jenni, O.G. Sleep electroencephalography topography and children’s intellectual ability. Neuroreport 2012, 23, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Walker, M.P. Daytime naps, motor memory consolidation and regionally specific sleep spindles. PLoS ONE 2007, 2, e341. [Google Scholar] [CrossRef] [PubMed]
- Barakat, M.; Doyon, J.; Debas, K.; Vandewalle, G.; Morin, A.; Poirier, G.; Martin, N.; Lafortune, M.; Karni, A.; Ungerleider, L.G.; et al. Fast and slow spindle involvement in the consolidation of a new motor sequence. Behav. Brain Res. 2011, 217, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Tamminen, J.; Lambon Ralph, M.A.; Lewis, P.A. The role of sleep spindles and slow-wave activity in integrating new information in semantic memory. J. Neurosci. 2013, 33, 15376–15381. [Google Scholar] [CrossRef] [PubMed]
- Chatburn, A.; Coussens, S.; Lushington, K.; Kennedy, D.; Baumert, M.; Kohler, M. Sleep spindle activity and cognitive performance in healthy children. Sleep 2013, 36, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Kurdziel, L.; Duclos, K.; Spencer, R.M. Sleep spindles in midday naps enhance learning in preschool children. Proc. Natl. Acad. Sci. USA 2013, 110, 17267–17272. [Google Scholar] [CrossRef] [PubMed]
- Mikoteit, T.; Brand, S.; Beck, J.; Perren, S.; von Wyl, A.; von Klitzing, K.; Holsboer-Trachsler, E.; Hatzinger, M. Visually detected NREM stage 2 sleep spindles in kindergarten children are associated with stress challenge and coping strategies. World J. Biol. Psychiatry 2012, 13, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, M.; Wilhelm, I.; Born, J.; Friederici, A.D. Generalization of word meanings during infant sleep. Nat. Commun. 2015, 6, 6004. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.; Kabani, N.J.; Lerch, J.P.; Eckstrand, K.; Lenroot, R.; Gogtay, N.; Greenstein, D.; Clasen, L.; Evans, A.; Rapoport, J.L.; et al. Neurodevelopmental trajectories of the human cerebral cortex. J. Neurosci. 2008, 28, 3586–3594. [Google Scholar] [CrossRef] [PubMed]
- Achenbach, T.M.; Rescorla, L.A. Manual for Aseba Preschool Forms & Profiles; University of Vermont, Research Center for Children, Youth, & Families: Burlington, VT, USA, 2000. [Google Scholar]
- Mullen, E.M. Mullen Scales of Early Learning; American Guidance Service: Circle Pines, MN, USA, 1995. [Google Scholar]
- Cepeda, N.J.; Blackwell, K.A.; Munakata, Y. Speed isn’t everything: Complex processing speed measures mask individual differences and developmental changes in executive control. Dev. Sci. 2013, 16, 269–286. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.; Graf, T.; Cote, K.A.; Wittmann, L.; Gallmann, E.; Matter, D.; Schuderer, J.; Kuster, N.; Borbely, A.A.; Achermann, P. Exposure to pulsed high-frequency electromagnetic field during waking affects human sleep EEG. Neuroreport 2000, 11, 3321–3325. [Google Scholar] [CrossRef] [PubMed]
- Iber, C.; Ancoli-Israel, S.; Chesson, A.L.; Quan, S.F. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications, 1st ed.; American Academy of Sleep Medicine: Westchester, IL, USA, 2007. [Google Scholar]
- Feinberg, I.; Floyd, T.C. Systematic trends across the night in human sleep cycles. Psychophysiology 1979, 16, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Altena, E.; Vrenken, H.; van der Werf, Y.D.; van den Heuvel, O.A.; van Someren, E.J. Reduced orbitofrontal and parietal gray matter in chronic insomnia: A voxel-based morphometric study. Biol. Psychiatry 2010, 67, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.Y.; Sluming, V.; Mayes, A.; Keller, S.; Barrick, T.; Cezayirli, E.; Roberts, N. Voxel-based morphometry and stereology provide convergent evidence of the importance of medial prefrontal cortex for fluid intelligence in healthy adults. NeuroImage 2005, 25, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Joo, E.Y.; Tae, W.S.; Kim, S.T.; Hong, S.B. Gray matter concentration abnormality in brains of narcolepsy patients. Korean J. Radiol. 2009, 10, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Suh, S.I.; Seol, H.Y.; Oh, K.; Seo, W.K.; Yu, S.W.; Park, K.W.; Koh, S.B. Regional grey matter changes in patients with migraine: A voxel-based morphometry study. Cephalalgia 2008, 28, 598–604. [Google Scholar] [CrossRef] [PubMed]
- O’Donoghue, F.J.; Briellmann, R.S.; Rochford, P.D.; Abbott, D.F.; Pell, G.S.; Chan, C.H.; Tarquinio, N.; Jackson, G.D.; Pierce, R.J. Cerebral structural changes in severe obstructive sleep apnea. Am. J. Respiratory Crit. Care Med. 2005, 171, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Yaouhi, K.; Bertran, F.; Clochon, P.; Mezenge, F.; Denise, P.; Foret, J.; Eustache, F.; Desgranges, B. A combined neuropsychological and brain imaging study of obstructive sleep apnea. J. Sleep Res. 2009, 18, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Bodizs, R.; Kis, T.; Lazar, A.S.; Havran, L.; Rigo, P.; Clemens, Z.; Halasz, P. Prediction of general mental ability based on neural oscillation measures of sleep. J. Sleep Res. 2005, 14, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Huttenlocher, P.R.; Dabholkar, A.S. Regional differences in synaptogenesis in human cerebral cortex. J. Comp. Neurol. 1997, 387, 167–178. [Google Scholar] [CrossRef]
- Kiselev, S.; Espy, K.A.; Sheffield, T. Age-related differences in reaction time task performance in young children. J. Exp. Child Psychol. 2009, 102, 150–166. [Google Scholar] [CrossRef] [PubMed]
- Kail, R. Developmental change in speed of processing during childhood and adolescence. Psychol. Bull. 1991, 109, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.T.; Vernon, P.A. Developmental changes in speed of information processing in young children. Dev. Psychol. 1997, 33, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Schmiedek, F.; Oberauer, K.; Wilhelm, O.; Suss, H.M.; Wittmann, W.W. Individual differences in components their relations to working of reaction time distributions and memory and intelligence. J. Exp. Psychol. Gen. 2007, 136, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Steriade, M. Corticothalamic resonance, states of vigilance and mentation. Neuroscience 2000, 101, 243–276. [Google Scholar] [CrossRef]
- Amzica, F.; Steriade, M. Integration of low-frequency sleep oscillations in corticothalamic networks. Acta Neurobiol. Exp. 2000, 60, 229–245. [Google Scholar]
- De Gennaro, L.; Ferrara, M. Sleep spindles: An overview. Sleep Med. Rev. 2003, 7, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Rypma, B.; Berger, J.S.; Prabhakaran, V.; Bly, B.M.; Kimberg, D.Y.; Biswal, B.B.; D’Esposito, M. Neural correlates of cognitive efficiency. NeuroImage 2006, 33, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.H.; Munakata, Y. Processes of change in brain and cognitive development. Trends Cogn. Sci. 2005, 9, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Tarokh, L.; Carskadon, M.A.; Achermann, P. Trait-like characteristics of the sleep EEG across adolescent development. J. Neurosci. 2011, 31, 6371–6378. [Google Scholar] [CrossRef] [PubMed]
- Kinomura, S.; Larsson, J.; Gulyas, B.; Roland, P.E. Activation by attention of the human reticular formation and thalamic intralaminar nuclei. Science 1996, 271, 512–515. [Google Scholar] [CrossRef] [PubMed]
- LaBerge, D.; Buchsbaum, M.S. Positron emission tomographic measurements of pulvinar activity during an attention task. J. Neurosci. 1990, 10, 613–619. [Google Scholar] [PubMed]
- Paus, T.; Zatorre, R.J.; Hofle, N.; Caramanos, Z.; Gotman, J.; Petrides, M.; Evans, A.C. Time-related changes in neural systems underlying attention and arousal during the performance of an auditory vigilance task. J. Cogn. Neurosci. 1997, 9, 392–408. [Google Scholar] [CrossRef] [PubMed]
- Portas, C.M.; Rees, G.; Howseman, A.M.; Josephs, O.; Turner, R.; Frith, C.D. A specific role for the thalamus in mediating the interaction of attention and arousal in humans. J. Neurosci. 1998, 18, 8979–8989. [Google Scholar] [PubMed]
- Hoedlmoser, K.; Heib, D.P.; Roell, J.; Peigneux, P.; Sadeh, A.; Gruber, G.; Schabus, M. Slow sleep spindle activity, declarative memory, and general cognitive abilities in children. Sleep 2014, 37, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Mason, T.B., 2nd; Teoh, L.; Calabro, K.; Traylor, J.; Karamessinis, L.; Schultz, B.; Samuel, J.; Gallagher, P.R.; Marcus, C.L. Rapid eye movement latency in children and adolescents. Pediatr. Neurol. 2008, 39, 162–169. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doucette, M.R.; Kurth, S.; Chevalier, N.; Munakata, Y.; LeBourgeois, M.K. Topography of Slow Sigma Power during Sleep is Associated with Processing Speed in Preschool Children. Brain Sci. 2015, 5, 494-508. https://doi.org/10.3390/brainsci5040494
Doucette MR, Kurth S, Chevalier N, Munakata Y, LeBourgeois MK. Topography of Slow Sigma Power during Sleep is Associated with Processing Speed in Preschool Children. Brain Sciences. 2015; 5(4):494-508. https://doi.org/10.3390/brainsci5040494
Chicago/Turabian StyleDoucette, Margaret R., Salome Kurth, Nicolas Chevalier, Yuko Munakata, and Monique K. LeBourgeois. 2015. "Topography of Slow Sigma Power during Sleep is Associated with Processing Speed in Preschool Children" Brain Sciences 5, no. 4: 494-508. https://doi.org/10.3390/brainsci5040494
APA StyleDoucette, M. R., Kurth, S., Chevalier, N., Munakata, Y., & LeBourgeois, M. K. (2015). Topography of Slow Sigma Power during Sleep is Associated with Processing Speed in Preschool Children. Brain Sciences, 5(4), 494-508. https://doi.org/10.3390/brainsci5040494