
GABAergic Influences on Medulloblastoma

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Origins of MB
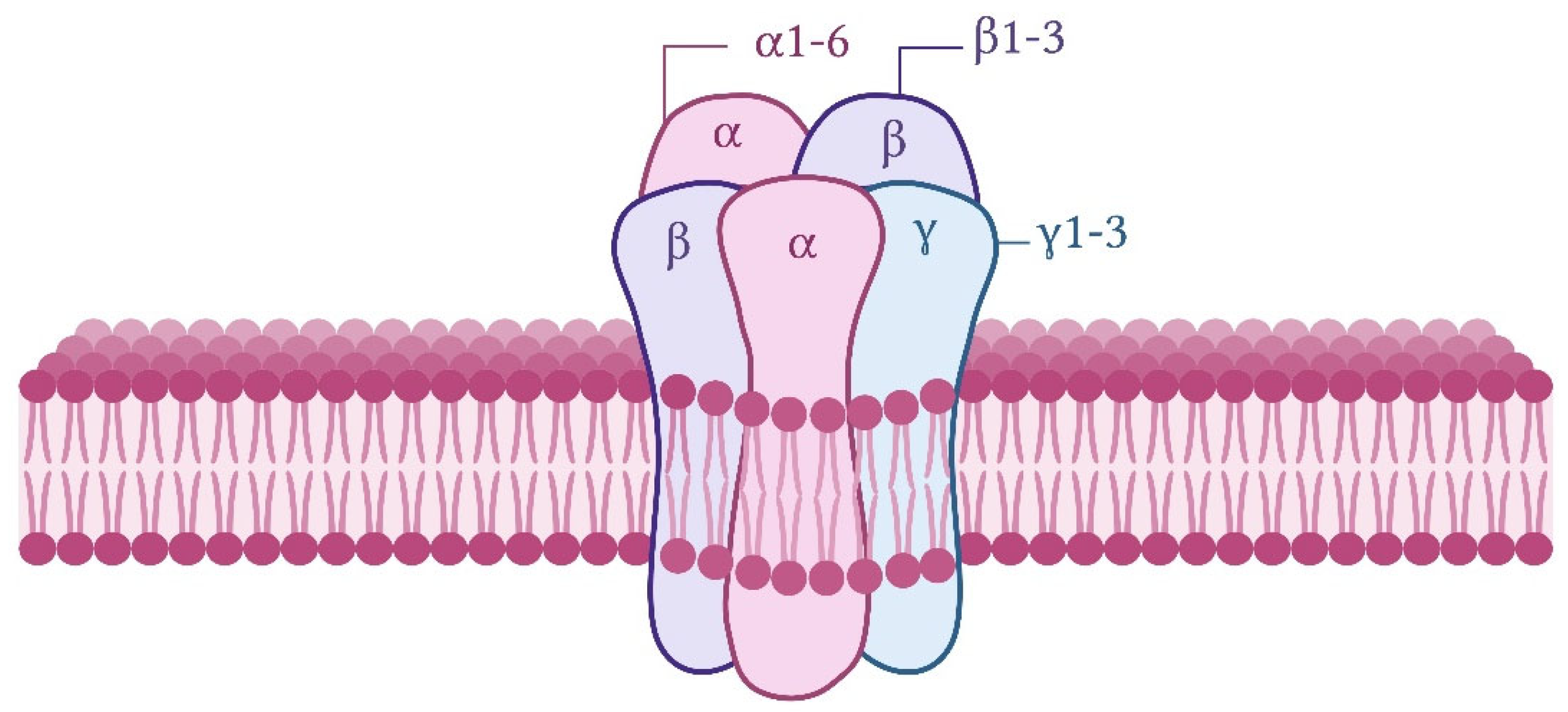
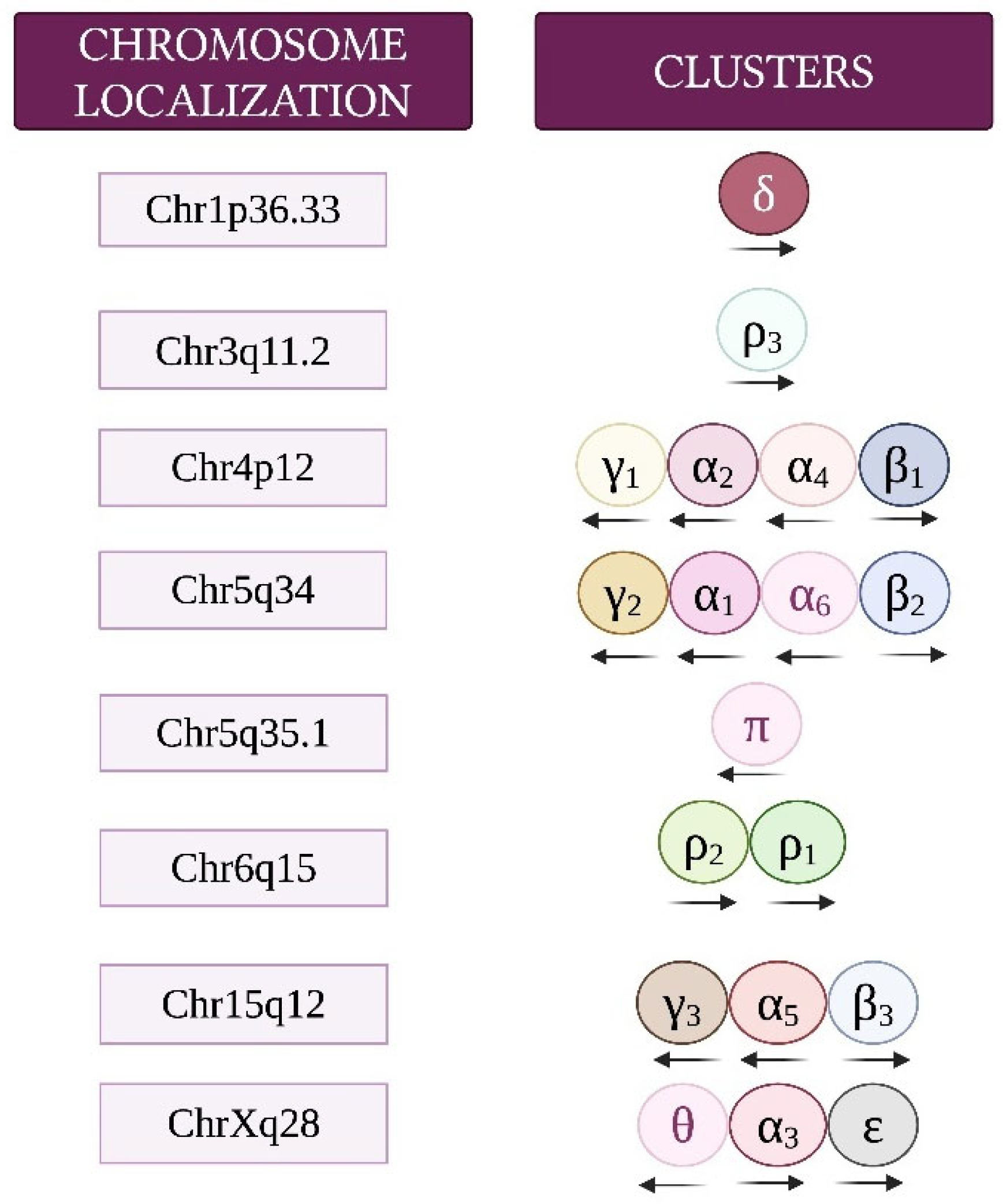
3. The GABAergic System and Its Role in Inhibitory Neurotransmission, Brain Development, and Synaptic Plasticity
4. GABAergic Regulation of Cerebellar Development and Function: Focus on GPCs and CGCs
5. The GABAergic System and Cancer
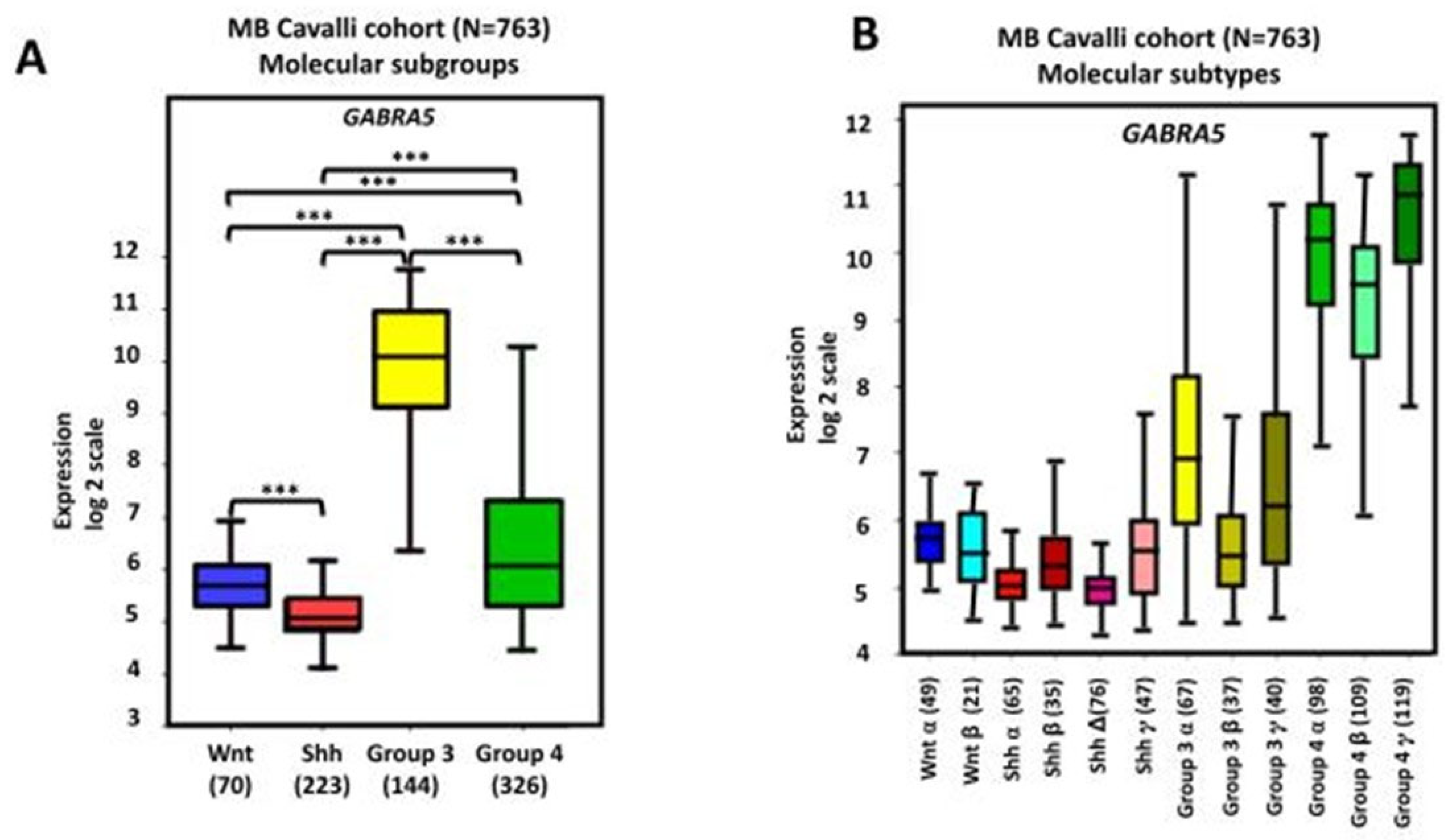
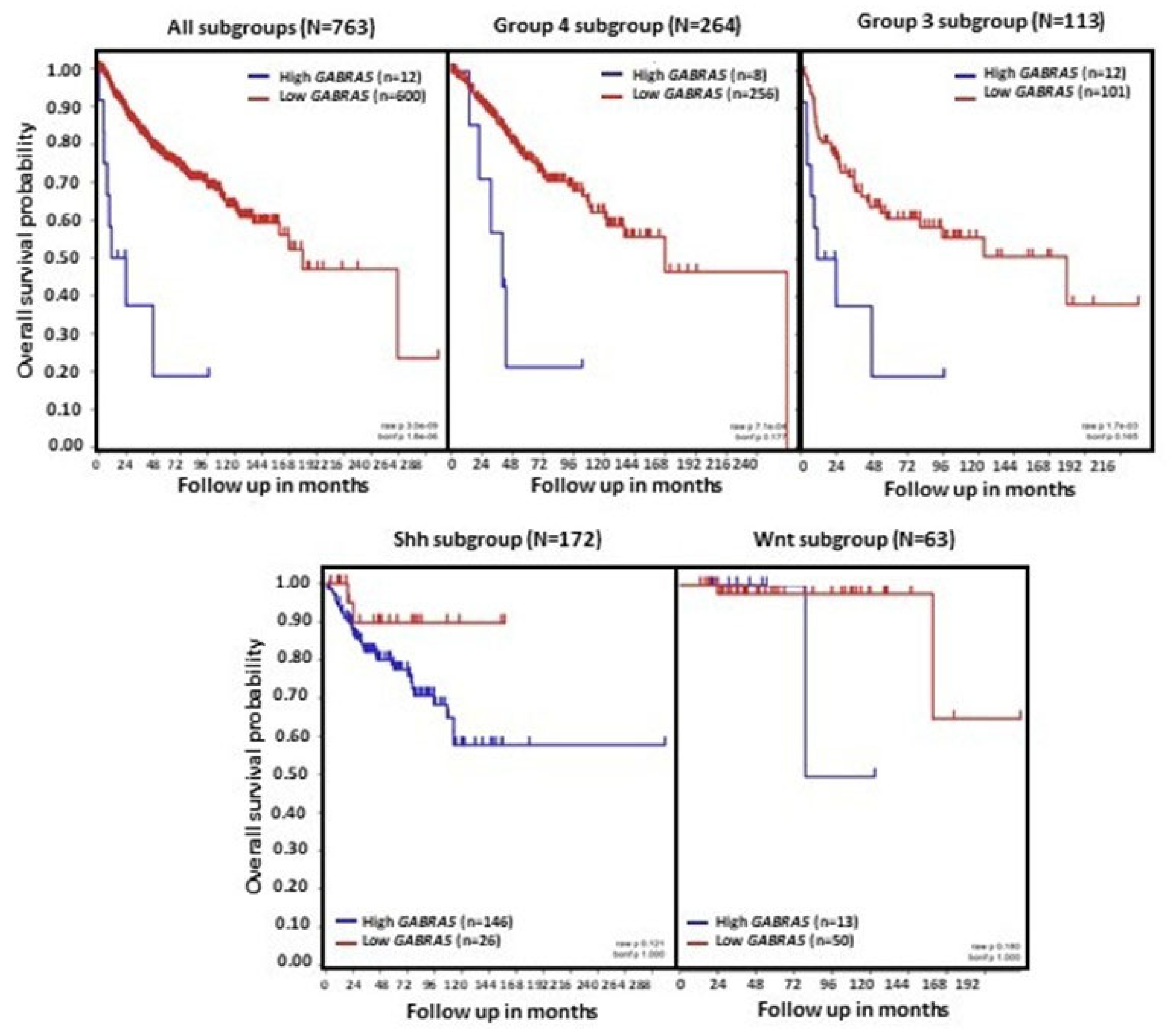
6. GABA Receptor Influences on MB
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Northcott, P.A.; Robinson, G.W.; Kratz, C.P.; Mabbott, D.J.; Pomeroy, S.L.; Clifford, S.C.; Rutkowski, S.; Ellison, D.W.; Malkin, D.; Taylor, M.D.; et al. Medulloblastoma. Nat. Rev. Dis. Primers 2019, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Ghit, A.; Assal, D.; Al-Shami, A.S.; Hussein, D.E.E. GABAA receptors: Structure, function, pharmacology, and related disorders. J. Genet. Eng. Biotechnol. 2021, 19, 123. [Google Scholar] [CrossRef]
- Simeone, T.A.; Donevan, S.D.; Rho, J.M. Molecular biology and ontogeny of gamma-aminobutyric acid (GABA) receptors in the mammalian central nervous system. J. Child Neurol. 2003, 18, 39–48, discussion 49. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, D.; Di Palma, M.; Cuppini, R.; Ambrogini, P. GABAergic input affects intracellular calcium levels in developing granule cells of adult rat hippocampus. Int. J. Mol. Sci. 2020, 21, 1715. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Guo, K.; Wu, B.; Wang, H. Overexpression of Shrm4 promotes proliferation and differentiation of neural stem cells through activation of GABA signaling pathway. Mol. Cell. Biochem. 2020, 463, 115–126. [Google Scholar] [CrossRef]
- Nakayama, H.; Miyazaki, T.; Kitamura, K.; Hashimoto, K.; Yanagawa, Y.; Obata, K.; Sakimura, K.; Watanabe, M.; Kano, M. GABAergic inhibition regulates developmental synapse elimination in the cerebellum. Neuron 2012, 74, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Zonouzi, M.; Scafidi, J.; Li, P.; McEllin, B.; Edwards, J.; Dupree, J.L.; Harvey, L.; Sun, D.; Hübner, C.A.; Cull-Candy, S.G.; et al. GABAergic regulation of cerebellar NG2 cell development is altered in perinatal white matter injury. Nat. Neurosci. 2015, 18, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthi, K.; Lin, Y. The contribution of GABAergic dysfunction to neurodevelopmental disorders. Trends Mol. Med. 2011, 17, 452–462. [Google Scholar] [CrossRef]
- Kallay, L.; Keskin, H.; Ross, A.; Rupji, M.; Moody, O.A.; Wang, X.; Li, G.; Ahmed, T.; Rashid, F.; Stephen, M.R.; et al. Modulating native GABAA receptors in medulloblastoma with positive allosteric benzodiazepine-derivatives induces cell death. J. Neurooncol. 2019, 142, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Weeraratne, S.D.; Sun, H.; Phallen, J.; Rallapalli, S.K.; Teider, N.; Kosaras, B.; Amani, V.; Pierre-Francois, J.; Tang, Y.; et al. α5-GABAA receptors negatively regulate MYC-amplified medulloblastoma growth. Acta Neuropathol. 2014, 127, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.M.; Dalmolin, M.; Fernandes, M.A.C.; Ramos, J.I.R.; Ribas, C.A.P.M.; Tabushi, F.I.; Roesler, R.; Isolan, G.R. High expression of GABAA receptor β subunit genes is associated with longer overall survival in medulloblastoma. Brain Sci. 2024, 14, 1146. [Google Scholar] [CrossRef]
- Gold, M.P.; Ong, W.; Masteller, A.M.; Ghasemi, D.R.; Galindo, J.A.; Park, N.R.; Huynh, N.C.; Donde, A.; Pister, V.; Saurez, R.A.; et al. Developmental basis of SHH medulloblastoma heterogeneity. Nat. Commun. 2024, 15, 270. [Google Scholar] [CrossRef] [PubMed]
- Jessa, S.; Blanchet-Cohen, A.; Krug, B.; Vladoiu, M.; Coutelier, M.; Faury, D.; Poreau, B.; De Jay, N.; Hébert, S.; Monlong, J.; et al. Stalled developmental programs at the root of pediatric brain tumors. Nat. Genet. 2019, 51, 1702–1713. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Xia, M.; Shi, W.; Zhao, C.; Wang, J.; Xin, D.; Dong, X.; Xiong, Y.; Zhang, F.; Berry, K.; et al. Human fetal cerebellar cell atlas informs medulloblastoma origin and oncogenesis. Nature 2022, 612, 787–794. [Google Scholar] [CrossRef]
- Northcott, P.A.; Buchhalter, I.; Morrissy, A.S.; Hovestadt, V.; Weischenfeldt, J.; Ehrenberger, T.; Gröbner, S.; Segura-Wang, M.; Zichner, T.; Rudneva, V.A.; et al. The whole-genome landscape of medulloblastoma subtypes. Nature 2017, 547, 311–317. [Google Scholar] [CrossRef]
- Thomaz, A.; Jaeger, M.; Brunetto, A.L.; Brunetto, A.T.; Gregianin, L.; de Farias, C.B.; Ramaswamy, V.; Nör, C.; Taylor, M.D.; Roesler, R. Neurotrophin signaling in medulloblastoma. Cancers 2020, 12, 2542. [Google Scholar] [CrossRef] [PubMed]
- Vladoiu, M.C.; El-Hamamy, I.; Donovan, L.K.; Farooq, H.; Holgado, B.L.; Sundaravadanam, Y.; Ramaswamy, V.; Hendrikse, L.D.; Kumar, S.; Mack, S.C.; et al. Childhood cerebellar tumours mirror conserved fetal transcriptional programs. Nature 2019, 572, 67–73. [Google Scholar] [CrossRef]
- Northcott, P.A.; Korshunov, A.; Witt, H.; Hielscher, T.; Eberhart, C.G.; Mack, S.; Bouffet, E.; Clifford, S.C.; Hawkins, C.E.; French, P.; et al. Medulloblastoma comprises four distinct molecular variants. J. Clin. Oncol. 2011, 29, 1408–1414. [Google Scholar] [CrossRef]
- Taylor, M.D.; Northcott, P.A.; Korshunov, A.; Remke, M.; Cho, Y.J.; Clifford, S.C.; Eberhart, C.G.; Parsons, D.W.; Rutkowski, S.; Gajjar, A.; et al. Molecular subgroups of medulloblastoma: The current consensus. Acta Neuropathol. 2012, 123, 465–472. [Google Scholar] [CrossRef]
- Juraschka, K.; Taylor, M.D. Medulloblastoma in the age of molecular subgroups: A review. J. Neurosurg. Pediatr. 2019, 24, 353–363. [Google Scholar] [CrossRef]
- Northcott, P.A.; Korshunov, A.; Pfister, S.M.; Taylor, M.D. The clinical implications of medulloblastoma subgroups. Nat. Rev. Neurol. 2012, 8, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, F.M.G.; Remke, M.; Rampasek, L.; Peacock, J.; Shih, D.J.H.; Luu, B.; Garzia, L.; Torchia, J.; Nor, C.; Morrissy, A.S.; et al. Intertumoral heterogeneity within medulloblastoma subgroups. Cancer Cell 2017, 31, 737–754.e6. [Google Scholar] [CrossRef]
- Northcott, P.A.; Shih, D.J.; Peacock, J.; Garzia, L.; Morrissy, A.S.; Zichner, T.; Stütz, A.M.; Korshunov, A.; Reimand, J.; Schumacher, S.E.; et al. Subgroup-specific structural variation across 1,000 medulloblastoma genomes. Nature 2012, 488, 49–56. [Google Scholar] [CrossRef]
- Schwalbe, E.C.; Lindsey, J.C.; Nakjang, S.; Crosier, S.; Smith, A.J.; Hicks, D.; Rafiee, G.; Hill, R.M.; Iliasova, A.; Stone, T.; et al. Novel molecular subgroups for clinical classification and outcome prediction in childhood medulloblastoma: A cohort study. Lancet Oncol. 2017, 18, 958–971. [Google Scholar] [CrossRef]
- Sharma, T.; Schwalbe, E.C.; Williamson, D.; Sill, M.; Hovestadt, V.; Mynarek, M.; Rutkowski, S.; Robinson, G.W.; Gajjar, A.; Cavalli, F.; et al. Second-generation molecular subgrouping of medulloblastoma: An international meta-analysis of Group 3 and Group 4 subtypes. Acta Neuropathol. 2019, 138, 309–326. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.; Tong, Y.; Robinson, G.; Thompson, M.C.; Currle, D.S.; Eden, C.; Kranenburg, T.A.; Hogg, T.; Poppleton, H.; Martin, J.; et al. Subtypes of medulloblastoma have distinct developmental origins. Nature 2010, 468, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Aldinger, K.A.; Thomson, Z.; Phelps, I.G.; Haldipur, P.; Deng, M.; Timms, A.E.; Hirano, M.; Santpere, G.; Roco, C.; Rosenberg, A.B.; et al. Spatial and cell type transcriptional landscape of human cerebellar development. Nat. Neurosci. 2021, 24, 1163–1175. [Google Scholar] [CrossRef]
- Haldipur, P.; Aldinger, K.A.; Bernardo, S.; Deng, M.; Timms, A.E.; Overman, L.M.; Winter, C.; Lisgo, S.N.; Razavi, F.; Silvestri, E.; et al. Spatiotemporal expansion of primary progenitor zones in the developing human cerebellum. Science 2019, 366, 454–460. [Google Scholar] [CrossRef]
- Oliver, T.G.; Read, T.A.; Kessler, J.D.; Mehmeti, A.; Wells, J.F.; Huynh, T.T.; Lin, S.M.; Wechsler-Reya, R.J. Loss of patched and disruption of granule cell development in a pre-neoplastic stage of medulloblastoma. Development 2005, 132, 2425–2439. [Google Scholar] [CrossRef]
- Wallace, V.A. Purkinje-cell-derived Sonic hedgehog regulates granule neuron precursor cell proliferation in the developing mouse cerebellum. Curr. Biol. 1999, 9, 445–448. [Google Scholar] [CrossRef]
- Wechsler-Reya, R.J.; Scott, M.P. Control of neuronal precursor proliferation in the cerebellum by Sonic Hedgehog. Neuron 1999, 22, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.J.; Ellis, T.; Markant, S.L.; Read, T.A.; Kessler, J.D.; Bourboulas, M.; Schüller, U.; Machold, R.; Fishell, G.; Rowitch, D.H.; et al. Medulloblastoma can be initiated by deletion of Patched in lineage-restricted progenitors or stem cells. Cancer Cell 2008, 14, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Kool, M.; Jones, D.T.; Jäger, N.; Northcott, P.A.; Pugh, T.J.; Hovestadt, V.; Piro, R.M.; Esparza, L.A.; Markant, S.L.; Remke, M.; et al. ICGC PedBrain Tumor Project. Genome sequencing of SHH medulloblastoma predicts genotype-related response to smoothened inhibition. Cancer Cell 2014, 25, 393–405. [Google Scholar] [CrossRef]
- Ramaswamy, V.; Remke, M.; Bouffet, E.; Bailey, S.; Clifford, S.C.; Doz, F.; Kool, M.; Dufour, C.; Vassal, G.; Milde, T.; et al. Risk stratification of childhood medulloblastoma in the molecular era: The current consensus. Acta Neuropathol. 2016, 131, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Zhukova, N.; Ramaswamy, V.; Remke, M.; Pfaff, E.; Shih, D.J.; Martin, D.C.; Castelo-Branco, P.; Baskin, B.; Ray, P.N.; Bouffet, E.; et al. Subgroup-specific prognostic implications of TP53 mutation in medulloblastoma. J. Clin. Oncol. 2013, 31, 2927–2935. [Google Scholar] [CrossRef]
- Smith, K.S.; Bihannic, L.; Gudenas, B.L.; Haldipur, P.; Tao, R.; Gao, Q.; Li, Y.; Aldinger, K.A.; Iskusnykh, I.Y.; Chizhikov, V.V.; et al. Unified rhombic lip origins of group 3 and group 4 medulloblastoma. Nature 2022, 609, 1012–1020. [Google Scholar] [CrossRef]
- Visvanathan, A.; Saulnier, O.; Chen, C.; Haldipur, P.; Orisme, W.; Delaidelli, A.; Shin, S.; Millman, J.; Bryant, A.; Abeysundara, N.; et al. Early rhombic lip Protogenin+ve stem cells in a human-specific neurovascular niche initiate and maintain group 3 medulloblastoma. Cell 2024, 187, 4733–4750.e26. [Google Scholar] [CrossRef]
- Lee, J.J.Y.; Tao, R.; You, Z.; Haldipur, P.; Erickson, A.W.; Farooq, H.; Hendriske, L.D.; Abeysundara, N.; Richman, C.M.; Wang, E.Y.; et al. ZIC1 is a context-dependent medulloblastoma driver in the rhombic lip. Nat. Genet. 2025, 57, 88–102. [Google Scholar] [CrossRef]
- Fan, X.; Eberhart, C.G. Medulloblastoma stem cells. J. Clin. Oncol. 2008, 26, 2821–2827. [Google Scholar] [CrossRef]
- Freire, N.H.; Herlinger, A.L.; Vanini, J.; Dalmolin, M.; Fernandes, M.A.C.; Nör, C.; Ramaswamy, V.; de Farias, C.B.; Brunetto, A.T.; Brunetto, A.L.; et al. Modulation of stemness and differentiation regulators by valproic acid in medulloblastoma neurospheres. Cells 2025, 14, 72. [Google Scholar] [CrossRef]
- Hemmati, H.D.; Nakano, I.; Lazareff, J.A.; Masterman-Smith, M.; Geschwind, D.H.; Bronner-Fraser, M.; Kornblum, H.I. Cancerous stem cells can arise from pediatric brain tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 15178–15183. [Google Scholar] [CrossRef] [PubMed]
- Manoranjan, B.; Venugopal, C.; McFarlane, N.; Doble, B.W.; Dunn, S.E.; Scheinemann, K.; Singh, S.K. Medulloblastoma stem cells: Modeling tumor heterogeneity. Cancer Lett. 2013, 338, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Manoranjan, B.; Wang, X.; Hallett, R.M.; Venugopal, C.; Mack, S.C.; McFarlane, N.; Nolte, S.M.; Scheinemann, K.; Gunnarsson, T.; Hassell, J.A.; et al. FoxG1 interacts with Bmi1 to regulate self-renewal and tumorigenicity of medulloblastoma stem cells. Stem Cells 2013, 31, 1266–1277. [Google Scholar] [CrossRef]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of human brain tumour initiating cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Johnston, G.A. Advantages of an antagonist: Bicuculline and other GABA antagonists. Br. J. Pharmacol. 2013, 169, 328–336. [Google Scholar] [CrossRef]
- Johnston, G.A. Flavonoid nutraceuticals and ionotropic receptors for the inhibitory neurotransmitter GABA. Neurochem. Int. 2015, 89, 120–125. [Google Scholar] [CrossRef]
- Möhler, H. The rise of a new GABA pharmacology. Neuropharmacology 2011, 60, 1042–1049. [Google Scholar] [CrossRef]
- Cossette, P.; Lachance-Touchette, P.; Rouleau, G.A. Mutated GABAA receptor subunits in idiopathic generalized epilepsy. In Jasper’s Basic Mechanisms of the Epilepsies, 4th ed.; Noebels, J.L., Avoli, M., Rogawski, M.A., Olsen, R.W., Delgado-Escueta, A.V., Eds.; National Center for Biotechnology Information: Bethesda, MD, USA, 2012. [Google Scholar]
- Russek, S.J. Evolution of GABA(A) receptor diversity in the human genome. Gene 1999, 227, 213–222. [Google Scholar] [CrossRef]
- Simon, J.; Wakimoto, H.; Fujita, N.; Lalande, M.; Barnard, E.A. Analysis of the set of GABA(A) receptor genes in the human genome. J. Biol. Chem. 2004, 279, 41422–41435. [Google Scholar] [CrossRef]
- Chapman, C.A.; Perez, Y.; Lacaille, J.C. Effects of GABA(A) inhibition on the expression of long-term potentiation in CA1 pyramidal cells are dependent on tetanization parameters. Hippocampus 1998, 8, 289–298. [Google Scholar] [CrossRef]
- Hirai, H.; Tomita, H.; Okada, Y. Inhibitory effect of GABA (gamma-aminobutyric acid) on the induction of long-term potentiation in guinea pig superior colliculus slices. Neurosci. Lett. 1993, 149, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Field, M.; Dorovykh, V.; Thomas, P.; Smart, T.G. Physiological role for GABAA receptor desensitization in the induction of long-term potentiation at inhibitory synapses. Nat. Commun. 2021, 12, 2112. [Google Scholar] [CrossRef] [PubMed]
- Kullmann, D.M.; Lamsa, K.P. LTP and LTD in cortical GABAergic interneurons: Emerging rules and roles. Neuropharmacology 2011, 60, 712–719. [Google Scholar] [CrossRef]
- Cherubini, E.; Gaiarsa, J.L.; Ben-Ari, Y. GABA: An excitatory transmitter in early postnatal life. Trends Neurosci. 1991, 14, 515–519. [Google Scholar] [CrossRef]
- Ji, X.; Liu, S.; Li, S.; Li, X.; Luo, A.; Zhang, X.; Zhao, Y. GABA in early brain development: A dual role review. Int. J. Dev. Neurosci. 2024, 84, 843–856. [Google Scholar] [CrossRef] [PubMed]
- LoTurco, J.J.; Owens, D.F.; Heath, M.J.; Davis, M.B.; Kriegstein, A.R. GABA and glutamate depolarize cortical progenitor cells and inhibit DNA synthesis. Neuron 1995, 15, 1287–1298. [Google Scholar] [CrossRef]
- Owens, D.F.; Kriegstein, A.R. Is there more to GABA than synaptic inhibition? Nat. Rev. Neurosci. 2002, 3, 715–727. [Google Scholar] [CrossRef]
- Topchiy, I.; Mohbat, J.; Folorunso, O.O.; Wang, Z.Z.; Lazcano-Etchebarne, C.; Engin, E. GABA system as the cause and effect in early development. Neurosci. Biobehav. Rev. 2024, 161, 105651. [Google Scholar] [CrossRef]
- Chen, G.; Trombley, P.Q.; van den Pol, A.N. GABA receptors precede glutamate receptors in hypothalamic development; differential regulation by astrocytes. J. Neurophysiol. 1995, 74, 1473–1484. [Google Scholar] [CrossRef]
- Ben-Ari, Y.; Tseeb, V.; Raggozzino, D.; Khazipov, R.; Gaiarsa, J.L. γ-Aminobutyric acid (GABA): A fast excitatory transmitter which may regulate the development of hippocampal neurones in early postnatal life. Prog. Brain Res. 1994, 102, 261–273. [Google Scholar] [CrossRef]
- Wu, C.; Sun, D. GABA receptors in brain development, function, and injury. Metab. Brain Dis. 2015, 30, 367–379. [Google Scholar] [CrossRef]
- Hashimoto, M.; Hibi, M. Development and evolution of cerebellar neural circuits. Dev. Growth Differ. 2012, 54, 373–389. [Google Scholar] [CrossRef]
- Hatten, M.E.; Heintz, N. Mechanisms of neural patterning and specification in the developing cerebellum. Annu. Rev. Neurosci. 1995, 18, 385–408. [Google Scholar] [CrossRef]
- Kuhar, S.G.; Feng, L.; Vidan, S.; Ross, M.E.; Hatten, M.E.; Heintz, N. Changing patterns of gene expression define four stages of cerebellar granule neuron differentiation. Development 1993, 117, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Sidman, R.L.; Rakic, P. Neuronal migration, with special reference to developing human brain: A review. Brain Res. 1973, 62, 1–35. [Google Scholar] [CrossRef]
- Rossi, D.J.; Slater, N.T. The developmental onset of NMDA receptor-channel activity during neuronal migration. Neuropharmacology 1993, 32, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.C.; Wang, L.Y.; Howe, J.R. Heterogeneous conductance levels of native AMPA receptors. J. Neurosci. 2000, 20, 2073–2085. [Google Scholar] [CrossRef] [PubMed]
- Dave, K.A.; Bordey, A. GABA increases Ca2+ in cerebellar granule cell precursors via depolarization: Implications for proliferation. IUBMB Life 2009, 61, 496–503. [Google Scholar] [CrossRef]
- Takayama, C.; Inoue, Y. Transient expression of GABAA receptor α2 and α3 subunits in differentiating cerebellar neurons. Brain Res. Dev. Brain Res. 2004, 148, 169–177. [Google Scholar] [CrossRef]
- Takayama, C. Formation of GABAergic synapses in the cerebellum. Cerebellum 2005, 4, 171–177. [Google Scholar] [CrossRef]
- Takayama, C.; Inoue, Y. Extrasynaptic localization of GABA in the developing mouse cerebellum. Neurosci. Res. 2004, 50, 447–458. [Google Scholar] [CrossRef]
- Studler, B.; Fritschy, J.M.; Brünig, I. GABAergic and glutamatergic terminals differentially influence the organization of GABAergic synapses in rat cerebellar granule cells in vitro. Neuroscience 2002, 114, 123–133. [Google Scholar] [CrossRef]
- Luján, R. Subcellular regulation of metabotropic GABA receptors in the developing cerebellum. Cerebellum 2007, 6, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Luján, R.; Shigemoto, R. Localization of metabotropic GABA receptor subunits GABAB1 and GABAB2 relative to synaptic sites in the rat developing cerebellum. Eur. J. Neurosci. 2006, 23, 1479–1490. [Google Scholar] [CrossRef]
- Turgeon, S.M.; Albin, R.L. Pharmacology, distribution, cellular localization, and development of GABAB binding in rodent cerebellum. Neuroscience 1993, 55, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Mejía, C.; García-Alcocer, G.; Berumen, L.C.; Rosas-Arellano, A.; Miledi, R.; Martínez-Torres, A. Expression of GABArho subunits during rat cerebellum development. Neurosci. Lett. 2008, 432, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.J.; Hamann, M. Spillover-mediated transmission at inhibitory synapses promoted by high affinity alpha6 subunit GABA(A) receptors and glomerular geometry. Neuron 1998, 20, 783–795. [Google Scholar] [CrossRef]
- Rossi, D.J.; Hamann, M.; Attwell, D. Multiple modes of GABAergic inhibition of rat cerebellar granule cells. J. Physiol. 2003, 548, 97–110. [Google Scholar] [CrossRef]
- Tia, S.; Wang, J.F.; Kotchabhakdi, N.; Vicini, S. Developmental changes of inhibitory synaptic currents in cerebellar granule neurons: Role of GABA(A) receptor alpha 6 subunit. J. Neurosci. 1996, 16, 3630–3640. [Google Scholar] [CrossRef]
- Fiszman, M.L.; Borodinsky, L.N.; Neale, J.H. GABA induces proliferation of immature cerebellar granule cells grown in vitro. Brain Res. Dev. Brain Res. 1999, 115, 1–8. [Google Scholar] [CrossRef]
- Carlson, B.X.; Elster, L.; Schousboe, A. Pharmacological and functional implications of developmentally-regulated changes in GABA(A) receptor subunit expression in the cerebellum. Eur. J. Pharmacol. 1998, 352, 1–14. [Google Scholar] [CrossRef]
- Hansen, G.H.; Meier, E.; Schousboe, A. GABA influences the ultrastructure composition of cerebellar granule cells during development in culture. Int. J. Dev. Neurosci. 1984, 2, 247–257. [Google Scholar] [CrossRef]
- Meier, E.; Hansen, G.H.; Schousboe, A. The trophic effect of gaba on cerebellar granule cells is mediated by gaba-receptors. Int. J. Dev. Neurosci. 1985, 3, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Nikas, P.; Gatta, E.; Cupello, A.; Di Braccio, M.; Grossi, G.; Pellistri, F.; Robello, M. Study of the interaction of 1,4- and 1,5-benzodiazepines with GABAA receptors of rat cerebellum granule cells in culture. J. Mol. Neurosci. 2015, 56, 768–772. [Google Scholar] [CrossRef]
- Yamashita, M.; Marszalec, W.; Yeh, J.Z.; Narahashi, T. Effects of ethanol on tonic GABA currents in cerebellar granule cells and mammalian cells recombinantly expressing GABA(A) receptors. J. Pharmacol. Exp. Ther. 2006, 319, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Barberis, A.; Lu, C.; Vicini, S.; Mozrzymas, J.W. Developmental changes of GABA synaptic transient in cerebellar granule cells. Mol. Pharmacol. 2005, 67, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Ives, J.H.; Drewery, D.L.; Thompson, C.L. Neuronal activity and its influence on developmentally regulated GABA(A) receptor expression in cultured mouse cerebellar granule cells. Neuropharmacology 2002, 43, 715–725. [Google Scholar] [CrossRef]
- Mellor, J.R.; Merlo, D.; Jones, A.; Wisden, W.; Randall, A.D. Mouse cerebellar granule cell differentiation: Electrical activity regulates the GABAA receptor alpha 6 subunit gene. J. Neurosci. 1998, 18, 2822–2833. [Google Scholar] [CrossRef]
- Balduzzi, R.; Cupello, A.; Robello, M. Modulation of the expression of GABA(A) receptors in rat cerebellar granule cells by protein tyrosine kinases and protein kinase C. Biochim. Biophys. Acta 2002, 1564, 263–270. [Google Scholar] [CrossRef]
- Khatri, S.N.; Wu, W.C.; Yang, Y.; Pugh, J.R. Mechanisms of GABAB receptor enhancement of extrasynaptic GABAA receptor currents in cerebellar granule cells. Sci. Rep. 2019, 9, 16683. [Google Scholar] [CrossRef]
- Rossi, P.; Mapelli, L.; Roggeri, L.; Gall, D.; de Kerchove d’Exaerde, A.; Schiffmann, S.N.; Taglietti, V.; D’Angelo, E. Inhibition of constitutive inward rectifier currents in cerebellar granule cells by pharmacological and synaptic activation of GABA receptors. Eur. J. Neurosci. 2006, 24, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Cao, L. Targeting GABA signalling for cancer treatment. Nat. Cell Biol. 2022, 24, 131–132. [Google Scholar] [CrossRef]
- Jiang, S.H.; Hu, L.P.; Wang, X.; Li, J.; Zhang, Z.G. Neurotransmitters: Emerging targets in cancer. Oncogene 2020, 39, 503–515. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Gawali, V.S.; Kallay, L.; Toukam, D.K.; Koehler, A.; Stambrook, P.; Krummel, D.P.; Sengupta, S. Therapeutically leveraging GABAA receptors in cancer. Exp. Biol. Med. 2021, 246, 2128–2135. [Google Scholar] [CrossRef]
- Watanabe, M.; Maemura, K.; Oki, K.; Shiraishi, N.; Shibayama, Y.; Katsu, K. Gamma-aminobutyric acid (GABA) and cell proliferation: Focus on cancer cells. Histol. Histopathol. 2006, 21, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wang, Y.; Thompson, J.W.; Yin, T.; Alexander, P.B.; Qin, D.; Mudgal, P.; Wu, H.; Liang, Y.; Tan, L.; et al. Cancer-cell-derived GABA promotes β-catenin-mediated tumour growth and immunosuppression. Nat. Cell Biol. 2022, 24, 230–241. [Google Scholar] [CrossRef]
- Brzozowska, A.; Burdan, F.; Duma, D.; Solski, J.; Mazurkiewicz, M. γ-amino butyric acid (GABA) level as an overall survival risk factor in breast cancer. Ann. Agric. Environ. Med. 2017, 24, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Bundy, J.; Shaw, J.; Hammel, M.; Nguyen, J.; Robbins, C.; Mercier, I.; Suryanarayanan, A. Role of β3 subunit of the GABA type A receptor in triple negative breast cancer proliferation, migration, and cell cycle progression. Cell Cycle 2024, 23, 448–465. [Google Scholar] [CrossRef]
- Bundy, J.; Ahmed, Y.; Weller, S.; Nguyen, J.; Shaw, J.; Mercier, I.; Suryanarayanan, A. GABA Type A receptors expressed in triple negative breast cancer cells mediate chloride ion flux. Front. Pharmacol. 2024, 15, 1449256. [Google Scholar] [CrossRef]
- Sizemore, G.M.; Sizemore, S.T.; Seachrist, D.D.; Keri, R.A. GABA(A) receptor pi (GABRP) stimulates basal-like breast cancer cell migration through activation of extracellular-regulated kinase 1/2 (ERK1/2). J. Biol. Chem. 2014, 289, 24102–24113. [Google Scholar] [CrossRef]
- Takehara, A.; Hosokawa, M.; Eguchi, H.; Ohigashi, H.; Ishikawa, O.; Nakamura, Y.; Nakagawa, H. Gamma-aminobutyric acid (GABA) stimulates pancreatic cancer growth through overexpressing GABAA receptor pi subunit. Cancer Res. 2007, 67, 9704–9712. [Google Scholar] [CrossRef]
- Dou, Z.; Li, M.; Shen, Z.; Jiang, H.; Pang, X.; Li, T.; Liang, X.; Tang, Y. GAD1-mediated GABA elicits aggressive characteristics of human oral cancer cells. Biochem. Biophys. Res. Commun. 2023, 681, 80–89. [Google Scholar] [CrossRef]
- Taylor, R.A.; Watt, M.J. Unsuspected protumorigenic signaling role for the oncometabolite GABA in advanced prostate cancer. Cancer Res. 2019, 79, 4580–4581. [Google Scholar] [CrossRef] [PubMed]
- Solorzano, S.R.; Imaz-Rosshandler, I.; Camacho-Arroyo, I.; García-Tobilla, P.; Morales-Montor, G.; Salazar, P.; Arena-Ortiz, M.L.; Rodríguez-Dorantes, M. GABA promotes gastrin-releasing peptide secretion in NE/NE-like cells: Contribution to prostate cancer progression. Sci. Rep. 2018, 8, 10272. [Google Scholar] [CrossRef]
- Gao, Y.; Chen, L.; Du, Z.; Gao, W.; Wu, Z.; Liu, X.; Huang, H.; Xu, D.; Li, Q. Glutamate decarboxylase 65 signals through the androgen receptor to promote castration resistance in prostate cancer. Cancer Res. 2019, 79, 4638–4649. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xu, X.; Liu, D.; Gao, J.; Gao, Y.; Wu, X.; Sheng, H.; Li, Q.; Mi, J. The delta subunit of the GABAA receptor is necessary for the GPT2-promoted breast cancer metastasis. Theranostics 2023, 13, 1355–1369. [Google Scholar] [CrossRef]
- Thaker, P.H.; Yokoi, K.; Jennings, N.B.; Li, Y.; Rebhun, R.B.; Rousseau, D.L., Jr.; Fan, D.; Sood, A.K. Inhibition of experimental colon cancer metastasis by the GABA-receptor agonist nembutal. Cancer Biol. Ther. 2005, 4, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Qin, H.; Liu, L.; Wu, J.; Zhao, Z.; Zhao, Y.; Fang, Y.; Yu, X.; Su, C. GABA regulates metabolic reprogramming to mediate the development of brain metastasis in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 2025, 44, 61. [Google Scholar] [CrossRef]
- Labrakakis, C.; Patt, S.; Hartmann, J.; Kettenmann, H. Functional GABA(A) receptors on human glioma cells. Eur. J. Neurosci. 1998, 10, 231–238. [Google Scholar] [CrossRef]
- El-Habr, E.A.; Dubois, L.G.; Burel-Vandenbos, F.; Bogeas, A.; Lipecka, J.; Turchi, L.; Lejeune, F.X.; Coehlo, P.L.; Yamaki, T.; Wittmann, B.M.; et al. A driver role for GABA metabolism in controlling stem and proliferative cell state through GHB production in glioma. Acta Neuropathol. 2017, 133, 645–660. [Google Scholar] [CrossRef]
- Synowitz, M.; Ahmann, P.; Matyash, M.; Kuhn, S.A.; Hofmann, B.; Zimmer, C.; Kirchhoff, F.; Kiwit, J.C.; Kettenmann, H. GABA(A)-receptor expression in glioma cells is triggered by contact with neuronal cells. Eur. J. Neurosci. 2001, 14, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Babateen, O.; Jin, Z.; Bhandage, A.; Korol, S.V.; Westermark, B.; Forsberg Nilsson, K.; Uhrbom, L.; Smits, A.; Birnir, B. Etomidate, propofol and diazepam potentiate GABA-evoked GABAA currents in a cell line derived from human glioblastoma. Eur. J. Pharmacol. 2015, 748, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Smits, A.; Jin, Z.; Elsir, T.; Pedder, H.; Nistér, M.; Alafuzoff, I.; Dimberg, A.; Edqvist, P.H.; Pontén, F.; Aronica, E.; et al. GABA-A channel subunit expression in human glioma correlates with tumor histology and clinical outcome. PLoS ONE 2012, 7, e37041. [Google Scholar] [CrossRef] [PubMed]
- Badalotti, R.; Dalmolin, M.; Malafaia, O.; Ribas Filho, J.M.; Roesler, R.; Fernandes, M.A.C.; Isolan, G.R. Gene expression of GABAA receptor subunits and association with patient survival in glioma. Brain Sci. 2024, 14, 275. [Google Scholar] [CrossRef]
- Shard, C.; Jones, A.C.; Fouladzadeh, A.; Palethorpe, H.M.; Francis, A.; Boyle, Y.; Ormsby, R.J.; Dewdney, B.; Yeow, Y.; Mahajan, I.; et al. Novel GABAAR antagonists target networked gene hubs at the leading-edge in high-grade gliomas. Neuro Oncol. 2025, noaf143. [Google Scholar] [CrossRef]
- Codina, C.; Kraft, R.; Pietsch, T.; Prinz, M.; Steinhäuser, C.; Cervós-Navarro, J.; Patt, S. Voltage- and gamma-aminobutyric acid-activated membrane currents in the human medulloblastoma cell line MHH-MED-3. Neurosci. Lett. 2000, 287, 53–56. [Google Scholar] [CrossRef]
- Cho, Y.J.; Tsherniak, A.; Tamayo, P.; Santagata, S.; Ligon, A.; Greulich, H.; Berhoukim, R.; Amani, V.; Goumnerova, L.; Eberhart, C.G.; et al. Integrative genomic analysis of medulloblastoma identifies a molecular subgroup that drives poor clinical outcome. J. Clin. Oncol. 2011, 29, 1424–1430. [Google Scholar] [CrossRef]
- Robinson, G.; Parker, M.; Kranenburg, T.A.; Lu, C.; Chen, X.; Ding, L.; Phoenix, T.N.; Hedlund, E.; Wei, L.; Zhu, X.; et al. Novel mutations target distinct subgroups of medulloblastoma. Nature 2012, 488, 43–48. [Google Scholar] [CrossRef]
- Mancusi, R.; Monje, M. The neuroscience of cancer. Nature 2023, 618, 467–479. [Google Scholar] [CrossRef]
- Prillaman, M. How cancer hijacks the nervous system to grow and spread. Nature 2024, 626, 22–24. [Google Scholar] [CrossRef]
- Mravec, B. Neurobiology of cancer: Definition, historical overview, and clinical implications. Cancer Med. 2022, 11, 903–921. [Google Scholar] [CrossRef] [PubMed]
- Winkler, F.; Venkatesh, H.S.; Amit, M.; Batchelor, T.; Demir, I.E.; Deneen, B.; Gutmann, D.H.; Hervey-Jumper, S.; Kuner, T.; Mabbott, D.; et al. Cancer neuroscience: State of the field, emerging directions. Cell 2023, 186, 1689–1707. [Google Scholar] [CrossRef] [PubMed]
- Barron, T.; Yalçın, B.; Su, M.; Byun, Y.G.; Gavish, A.; Shamardani, K.; Xu, H.; Ni, L.; Soni, N.; Mehta, V.; et al. GABAergic neuron-to-glioma synapses in diffuse midline gliomas. Nature 2025, 639, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buffon, V.A.; Filho, J.M.R.; Malafaia, O.; Tassinari, I.D.; Roesler, R.; Isolan, G.R. GABAergic Influences on Medulloblastoma. Brain Sci. 2025, 15, 746. https://doi.org/10.3390/brainsci15070746
Buffon VA, Filho JMR, Malafaia O, Tassinari ID, Roesler R, Isolan GR. GABAergic Influences on Medulloblastoma. Brain Sciences. 2025; 15(7):746. https://doi.org/10.3390/brainsci15070746
Chicago/Turabian StyleBuffon, Viviane Aline, Jurandir M. Ribas Filho, Osvaldo Malafaia, Isadora D. Tassinari, Rafael Roesler, and Gustavo R. Isolan. 2025. "GABAergic Influences on Medulloblastoma" Brain Sciences 15, no. 7: 746. https://doi.org/10.3390/brainsci15070746
APA StyleBuffon, V. A., Filho, J. M. R., Malafaia, O., Tassinari, I. D., Roesler, R., & Isolan, G. R. (2025). GABAergic Influences on Medulloblastoma. Brain Sciences, 15(7), 746. https://doi.org/10.3390/brainsci15070746