

Exploring Neural Signaling Patterns and Their Physiological Origins in Fibromyalgia by Means of Functional MRI Guided by a Review of the Literature

Abstract
1. Introduction
2. Materials and Methods
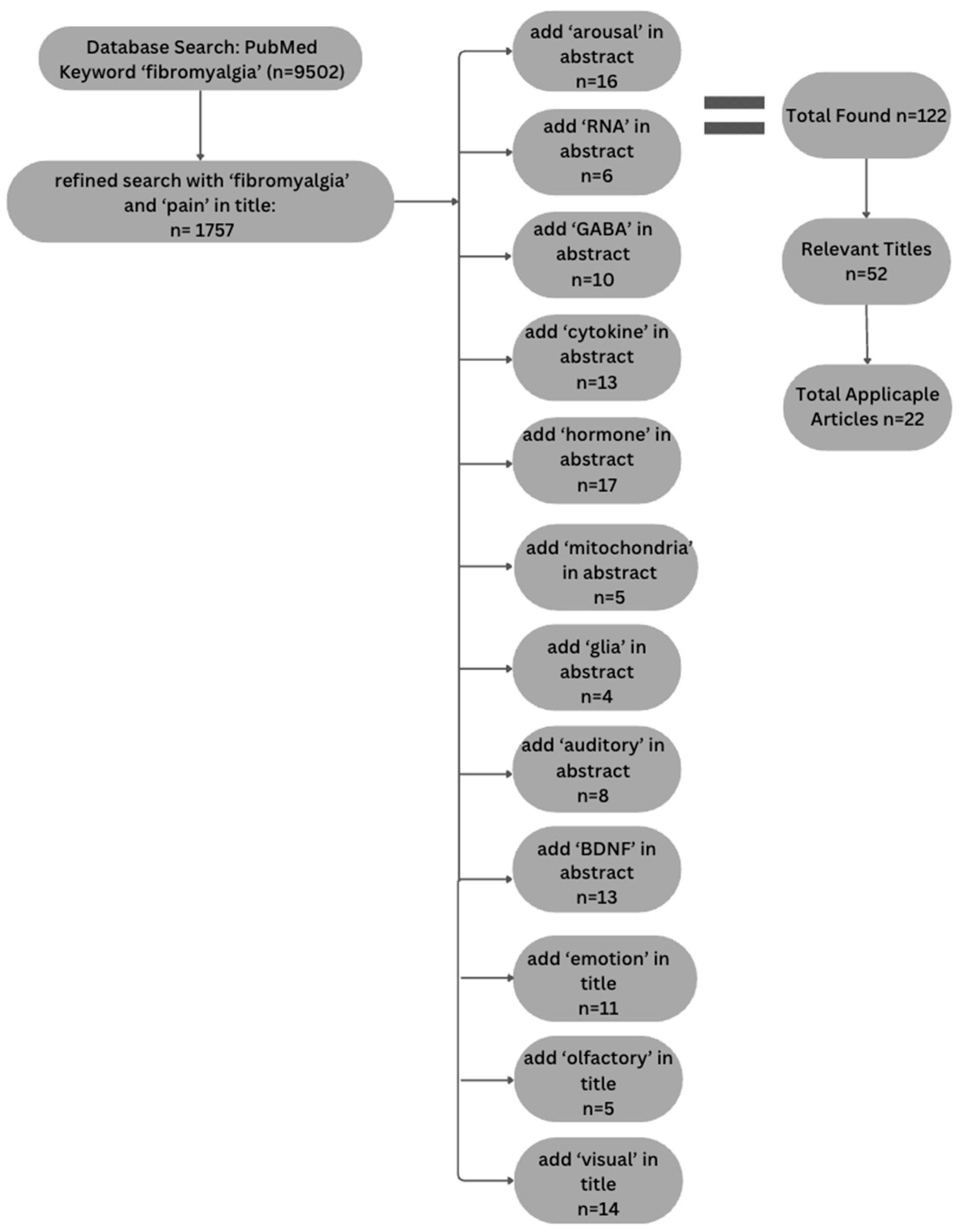
2.1. Component 1: Literature Review
2.2. Component 2: Analysis of Existing fMRI Data
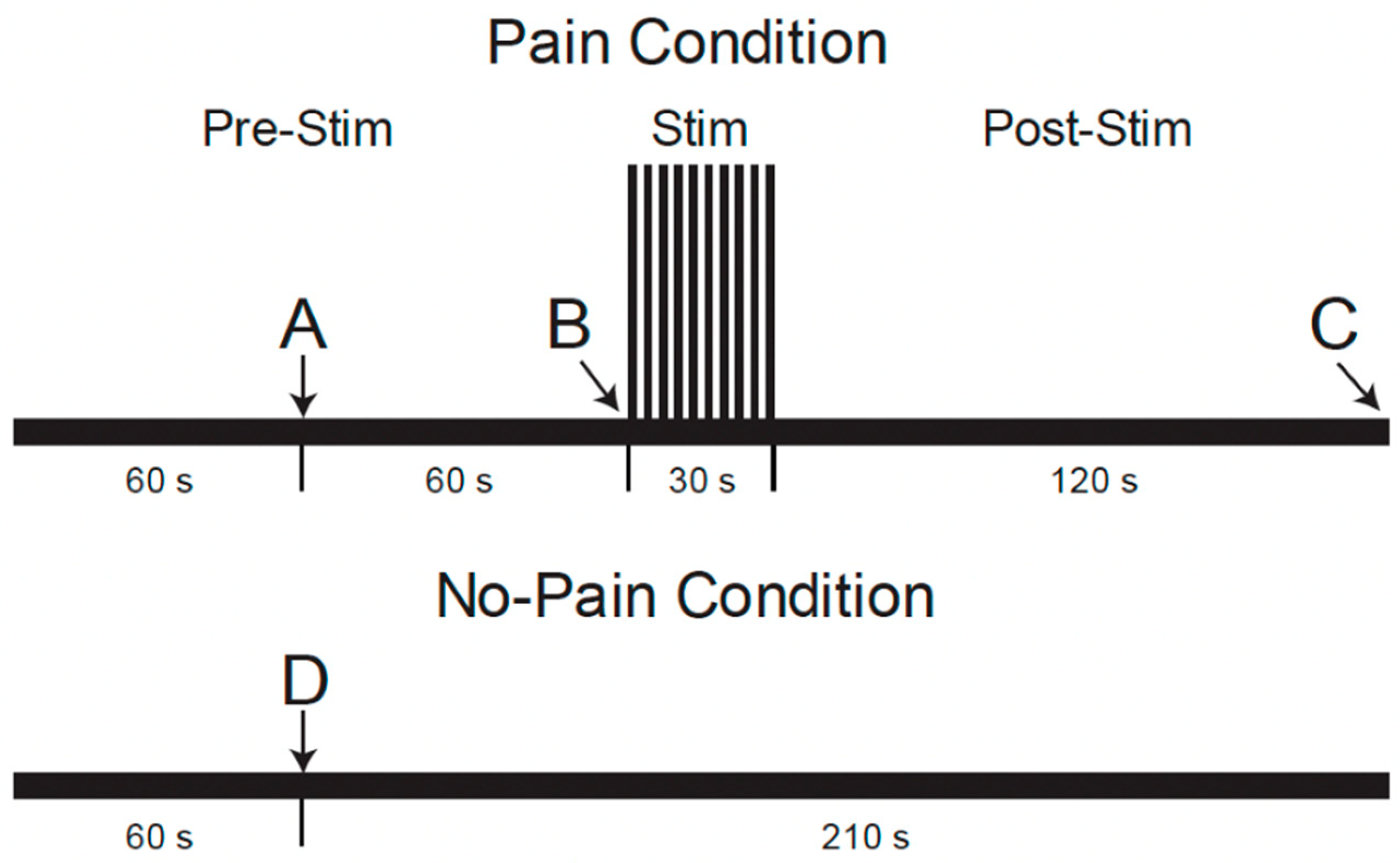
2.2.1. Source of the Existing Data and Supporting Participant Information
2.2.2. Analysis Methods for the Present Study
3. Results
3.1. Study Component 1: Literature Review
3.1.1. Cognitive Influences
Emotional Arousal
Sensitivity to Sensory Stimuli (Auditory, Olfactory, and Visual)
3.1.2. Physiological Influences
BDNF and GABA Relationship
Mitochondria Dysfunction
Changes in Glial Cells
DNA and RNA
Proteins: Cytokines and Translocator Protein Prevalence
Presence and Regulation of Dopamine
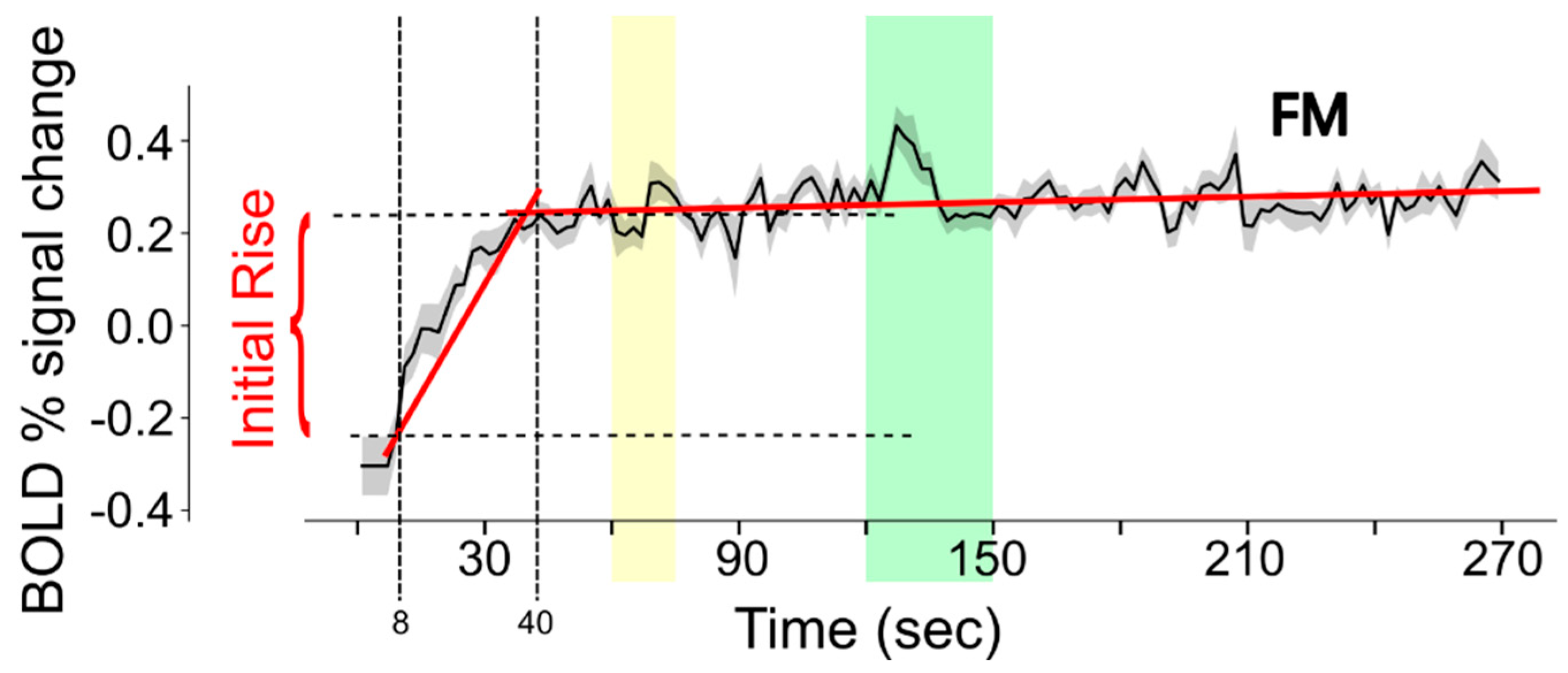
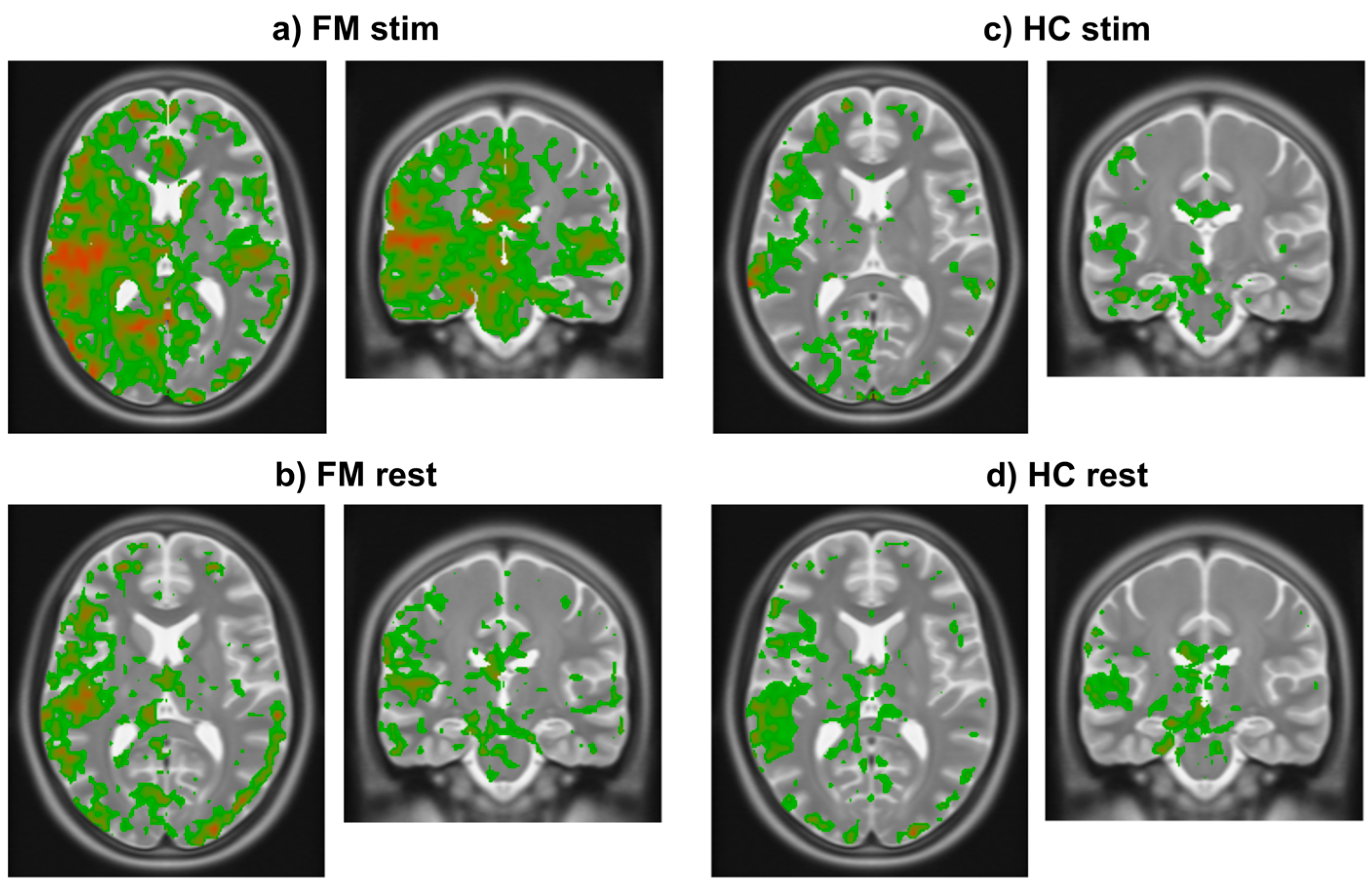
3.2. Study Component 2: Analysis of FMRI Data
4. Discussion
4.1. Interpretation of Literature Review
4.2. Interpretation of fMRI Data Analysis
4.3. Limitations and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ichesco, E.; Puiu, T.; Hampson, J.P.; Kairys, A.E.; Clauw, D.J.; Harte, S.E.; Peltier, S.J.; Harris, R.E.; Schmidt-Wilcke, T. Altered fMRI resting-state connectivity in individuals with fibromyalgia on acute pain stimulation. Eur. J. Pain 2016, 20, 1079–1089. [Google Scholar] [CrossRef]
- Staud, R.; Godfrey, M.M.; Stroman, P.W. Fibromyalgia is associated with hypersensitivity but not with abnormal pain modulation: Evidence from QST trials and spinal fMRI. Front. Pain Res. 2023, 4, 1284103. [Google Scholar] [CrossRef]
- Ioachim, G.; Warren, H.J.M.; Powers, J.M.; Staud, R.; Pukall, C.F.; Stroman, P.W. Distinct neural signaling characteristics between fibromyalgia and provoked vestibulodynia revealed by means of functional magnetic resonance imaging in the brainstem and spinal cord. Front. Pain Res. 2023, 4, 1171160. [Google Scholar] [CrossRef]
- Ioachim, G.; Warren, H.J.M.; Powers, J.M.; Staud, R.; Pukall, C.F.; Stroman, P.W. Altered Pain in the Brainstem and Spinal Cord of Fibromyalgia Patients During the Anticipation and Experience of Experimental Pain. Front. Neurol. 2022, 13, 862976. [Google Scholar] [CrossRef]
- Staud, R.; Boissoneault, J.; Lai, S.; Mejia, M.S.; Ramanlal, R.; Godfrey, M.M.; Stroman, P.W. Descending Modulation of Spinal Cord Activity in Fibromyalgia Patients During Temporal Summation of Pain: An fMRI Study of the Spinal Cord. Eur. J. Pain, 2021; Submitted. [Google Scholar]
- Warren, H.J.M.; Ioachim, G.; Powers, J.M.; Stroman, P.W. How fMRI Analysis Using Structural Equation Modeling Techniques Can Improve Our Understanding of Pain Processing in Fibromyalgia. J. Pain Res. 2021, 14, 381–398. [Google Scholar] [CrossRef]
- Pando-Naude, V.; Barrios, F.A.; Alcauter, S.; Pasaye, E.H.; Vase, L.; Brattico, E.; Vuust, P.; Garza-Villarreal, E.A. Functional connectivity of music-induced analgesia in fibromyalgia. Sci. Rep. 2019, 9, 15486. [Google Scholar] [CrossRef]
- Martucci, K.T.; Weber, K.A., 2nd; Mackey, S.C. Altered Cervical Spinal Cord Resting-State Activity in Fibromyalgia. Arthritis Rheumatol. 2019, 71, 441–450. [Google Scholar] [CrossRef]
- Lopez-Sola, M.; Woo, C.W.; Pujol, J.; Deus, J.; Harrison, B.J.; Monfort, J.; Wager, T.D. Towards a neurophysiological signature for fibromyalgia. Pain 2017, 158, 34–47. [Google Scholar] [CrossRef]
- Jarrahi, B.; Martucci, K.T.; Nilakantan, A.S.; Mackey, S. Investigating the BOLD spectral power of the intrinsic connectivity networks in fibromyalgia patients: A resting-state fMRI study. In Proceedings of the 2017 39th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Jeju, Republic of Korea, 11–15 July 2017; pp. 497–500. [Google Scholar] [CrossRef]
- Bosma, R.L.; Mojarad, E.A.; Leung, L.; Pukall, C.; Staud, R.; Stroman, P.W. FMRI of spinal and supra-spinal correlates of temporal pain summation in fibromyalgia patients. Hum. Brain Mapp. 2016, 37, 1349–1360. [Google Scholar] [CrossRef]
- Cagnie, B.; Coppieters, I.; Denecker, S.; Six, J.; Danneels, L.; Meeus, M. Central sensitization in fibromyalgia? A systematic review on structural and functional brain MRI. Semin. Arthritis Rheum. 2014, 44, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Flodin, P.; Martinsen, S.; Lofgren, M.; Bileviciute-Ljungar, I.; Kosek, E.; Fransson, P. Fibromyalgia is associated with decreased connectivity between pain- and sensorimotor brain areas. Brain Connect. 2014, 4, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.B.; Loitoile, R.; Kosek, E.; Petzke, F.; Carville, S.; Fransson, P.; Marcus, H.; Williams, S.C.; Choy, E.; Mainguy, Y.; et al. Patients with Fibromyalgia Display Less Functional Connectivity in the Brain’s Pain Inhibitory Network. Mol. Pain 2012, 8, 32. [Google Scholar] [CrossRef]
- Gracely, R.H.; Ambrose, K.R. Neuroimaging of fibromyalgia. Best Pract. Res. Clin. Rheumatol. 2011, 25, 271–284. [Google Scholar] [CrossRef]
- Burgmer, M.; Pogatzki-Zahn, E.; Gaubitz, M.; Stuber, C.; Wessoleck, E.; Heuft, G.; Pfleiderer, B. Fibromyalgia unique temporal brain activation during experimental pain: A controlled fMRI Study. J. Neural Transm. 2010, 117, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Burgmer, M.; Pogatzki-Zahn, E.; Gaubitz, M.; Wessoleck, E.; Heuft, G.; Pfleiderer, B. Altered brain activity during pain processing in fibromyalgia. Neuroimage 2009, 44, 502–508. [Google Scholar] [CrossRef]
- Pujol, J.; Lopez-Sola, M.; Ortiz, H.; Vilanova, J.C.; Harrison, B.J.; Yucel, M.; Soriano-Mas, C.; Cardoner, N.; Deus, J. Mapping brain response to pain in fibromyalgia patients using temporal analysis of FMRI. PLoS ONE 2009, 4, e5224. [Google Scholar] [CrossRef]
- Staud, R.; Craggs, J.G.; Perlstein, W.M.; Robinson, M.E.; Price, D.D. Brain activity associated with slow temporal summation of C-fiber evoked pain in fibromyalgia patients and healthy controls. Eur. J. Pain 2008, 12, 1078–1089. [Google Scholar] [CrossRef]
- Cook, D.B.; Lange, G.; Ciccone, D.S.; Liu, W.C.; Steffener, J.; Natelson, B.H. Functional imaging of pain in patients with primary fibromyalgia. J. Rheumatol. 2004, 31, 364–378. [Google Scholar]
- Gracely, R.H.; Petzke, F.; Wolf, J.M.; Clauw, D.J. Functional magnetic resonance imaging evidence of augmented pain processing in fibromyalgia. Arthritis Rheum. 2002, 46, 1333–1343. [Google Scholar] [CrossRef]
- Warren, H.J.M.; Ioachim, G.; Powers, J.M.; Staud, R.; Pukall, C.; Stroman, P.W. Using Structural Equation Modeling to Investigate the Neural Basis of Altered Pain Processing in Fibromyalgia with Functional Magnetic Resonance Imaging. Med. Res. Arch. 2024, 12. [Google Scholar] [CrossRef]
- Stroman, P.W.; Staud, R.; Pukall, C.F. Evidence of a persistent altered neural state in people with fibromyalgia syndrome during functional MRI studies and its relationship with pain and anxiety. PLoS ONE 2025, 20, e0316672. [Google Scholar] [CrossRef] [PubMed]
- Stahl, S.M. Fibromyalgia—Pathways and neurotransmitters. Hum. Psychopharmacol. 2009, 24 (Suppl. S1), S11–S17. [Google Scholar] [CrossRef]
- Caumo, W.; Deitos, A.; Carvalho, S.; Leite, J.; Carvalho, F.; Dussán-Sarria, J.A.; Lopes Tarragó, M.d.G.; Souza, A.; Torres, I.L.d.S.; Fregni, F. Motor Cortex Excitability and BDNF Levels in Chronic Musculoskeletal Pain According to Structural Pathology. Front. Hum. Neurosci. 2016, 10, 357. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, N.; Wei, M.; Pan, Q.; Cheng, C.; Lu, K.E.; Mo, J.; Chen, Y. Methylation factors as biomarkers of fibromyalgia. Ann. Transl. Med. 2023, 11, 169. [Google Scholar] [CrossRef]
- Albrecht, P.J.; Rice, F.L. Fibromyalgia syndrome pathology and environmental influences on afflictions with medically unexplained symptoms. Rev. Environ. Health 2016, 31, 281–294. [Google Scholar] [CrossRef]
- Marino, Y.; Inferrera, F.; D’Amico, R.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Gugliandolo, E.; Fusco, R.; Cuzzocrea, S.; Di Paola, R. Role of mitochondrial dysfunction and biogenesis in fibromyalgia syndrome: Molecular mechanism in central nervous system. Biochim. Biophys. Acta (BBA) Mol. Basis Disease 2024, 1870, 167301. [Google Scholar] [CrossRef] [PubMed]
- Backryd, E.; Tanum, L.; Lind, A.L.; Larsson, A.; Gordh, T. Evidence of both systemic inflammation and neuroinflammation in fibromyalgia patients, as assessed by a multiplex protein panel applied to the cerebrospinal fluid and to plasma. J. Pain Res. 2017, 10, 515–525. [Google Scholar] [CrossRef]
- Stroman, P.W.; Powers, J.M.; Ioachim, G. Proof-of-concept of a novel structural equation modelling approach for the analysis of functional magnetic resonance imaging data applied to investigate individual differences in human pain responses. Hum. Brain Mapp. 2023, 44, 2523–2542. [Google Scholar] [CrossRef]
- Stroman, P.W.; Umraw, M.; Keast, B.; Algitami, H.; Hassanpour, S.; Merletti, J. Structural and Physiological Modeling (SAPM) for the Analysis of Functional MRI Data Applied to a Study of Human Nociceptive Processing. Brain Sci. 2023, 13, 1568. [Google Scholar] [CrossRef]
- Naidich, T.P.; Duvernoy, H.M.; Delman, B.N.; Sorensen, A.G.; Kollias, S.S.; Haacke, E.M. Internal Architecture of the Brain Stem with Key Axial Sections. In Duvernoy’s Atlas of the Human Brain Stem and Cerebellum; Springer: New York, NY, USA; Wien, Austria, 2009; pp. 79–82. [Google Scholar]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A functional connectivity toolbox for correlated and anticorrelated brain networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef]
- Millan, M.J. Descending control of pain. Prog. Neurobiol. 2002, 66, 355–474. [Google Scholar] [PubMed]
- De Leener, B.; Levy, S.; Dupont, S.M.; Fonov, V.S.; Stikov, N.; Louis Collins, D.; Callot, V.; Cohen-Adad, J. SCT: Spinal Cord Toolbox, an open-source software for processing spinal cord MRI data. Neuroimage 2017, 145, 24–43. [Google Scholar] [CrossRef]
- Lang, J.; Bartram, C.T. Fila radicularia of the ventral and dorsal radices of the human spinal cord. Gegenbaurs Morphol. Jahrb. 1982, 128, 417–462. [Google Scholar]
- Liebe, T.; Kaufmann, J.; Li, M.; Skalej, M.; Wagner, G.; Walter, M. In vivo anatomical mapping of human locus coeruleus functional connectivity at 3 T MRI. Hum. Brain Mapp. 2020, 41, 2136–2151. [Google Scholar] [CrossRef]
- Chiang, M.C.; Bowen, A.; Schier, L.A.; Tupone, D.; Uddin, O.; Heinricher, M.M. Parabrachial Complex: A Hub for Pain and Aversion. J. Neurosci. 2019, 39, 8225–8230. [Google Scholar] [CrossRef] [PubMed]
- Talairach, J.; Tournoux, P. Co-Planar Stereotaxic Atlas of the Human Brain; Thieme Medical Publishers, Inc.: New York, NY, USA, 1988. [Google Scholar]
- Leijnse, J.N.; D’Herde, K. Revisiting the segmental organization of the human spinal cord. J. Anat. 2016, 229, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Pauli, W.M.; Nili, A.N.; Tyszka, J.M. A high-resolution probabilistic in vivo atlas of human subcortical brain nuclei. Sci. Data 2018, 5, 180063. [Google Scholar] [CrossRef]
- Keren, N.I.; Lozar, C.T.; Harris, K.C.; Morgan, P.S.; Eckert, M.A. In vivo mapping of the human locus coeruleus. Neuroimage 2009, 47, 1261–1267. [Google Scholar] [CrossRef]
- Stroman, P.W.; Warren, H.J.M.; Ioachim, G.; Powers, J.M.; McNeil, K. A comparison of the effectiveness of functional MRI analysis methods for pain research: The new normal. PLoS ONE 2020, 15, e0243723. [Google Scholar] [CrossRef]
- Ciuffini, R.; Cofini, V.; Muselli, M.; Necozione, S.; Piroli, A.; Marrelli, A. Emotional arousal and valence in patients with fibromyalgia: A pilot study. Front. Pain Res. 2023, 4, 1075722. [Google Scholar] [CrossRef] [PubMed]
- Frumer, L.; Marom Harel, H.; Horesh, D. Why Does It Hurt So Much? Emotion Regulation Mediates the Association between Fibromyalgia Symptoms and Psychological Distress. Healthcare 2023, 11, 1368. [Google Scholar] [CrossRef]
- Montoro, C.I.; Galvez-Sanchez, C.M. The Mediating Role of Depression and Pain Catastrophizing in the Relationship between Functional Capacity and Pain Intensity in Patients with Fibromyalgia. Behav. Neurol. 2022, 2022, 9770047. [Google Scholar] [CrossRef]
- Carrillo-de-la-Pena, M.T.; Trinanes, Y.; Gonzalez-Villar, A.; Gomez-Perretta, C.; Garcia-Larrea, L. Filtering out repetitive auditory stimuli in fibromyalgia: A study of P50 sensory gating. Eur. J. Pain 2015, 19, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Staud, R.; Godfrey, M.M.; Robinson, M.E. Fibromyalgia Patients Are Not Only Hypersensitive to Painful Stimuli But Also to Acoustic Stimuli. J. Pain Off. J. Am. Pain Society 2021, 22, 914–925. [Google Scholar] [CrossRef]
- Samartin-Veiga, N.; Gonzalez-Villar, A.J.; Trinanes, Y.; Gomez-Perretta, C.; Carrillo-de-la-Pena, M.T. Effects of intensity, attention and medication on auditory-evoked potentials in patients with fibromyalgia. Sci. Rep. 2020, 10, 21904. [Google Scholar] [CrossRef]
- Ozsoy-Unubol, T.; Kullakci, H.; Ilhan, I.; Yilmaz, F. Evaluation of Olfactory and Gustatory Functions in Patients with Fibromyalgia Syndrome: Its Relationship with Anxiety, Depression, and Alexithymia. Arch. Rheumatol. 2020, 35, 584–591. [Google Scholar] [CrossRef]
- Nhu, N.T.; Chen, D.Y.; Kang, J.H. Functional Connectivity and Structural Signatures of the Visual Cortical System in Fibromyalgia: A Magnetic Resonance Imaging Study. J. Rheumatol. 2023, 50, 1063–1070. [Google Scholar] [CrossRef]
- Pomares, F.B.; Funck, T.; Feier, N.A.; Roy, S.; Daigle-Martel, A.; Ceko, M.; Narayanan, S.; Araujo, D.; Thiel, A.; Stikov, N.; et al. Histological Underpinnings of Grey Matter Changes in Fibromyalgia Investigated Using Multimodal Brain Imaging. J. Neurosci. 2017, 37, 1090–1101. [Google Scholar] [CrossRef]
- Inferrera, F.; Marino, Y.; D’Amico, R.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Gugliandolo, E.; Fusco, R.; Cuzzocrea, S.; Di Paola, R. Impaired mitochondrial quality control in fibromyalgia: Mechanisms involved in skeletal muscle alteration. Arch. Biochem. Biophys. 2024, 758, 110083. [Google Scholar] [CrossRef]
- Martinez-Lara, A.; Moreno-Fernandez, A.M.; Jimenez-Guerrero, M.; Diaz-Lopez, C.; De-Miguel, M.; Cotan, D.; Sanchez-Alcazar, J.A. Mitochondrial Imbalance as a New Approach to the Study of Fibromyalgia. Open Access Rheumatol. 2020, 12, 175–185. [Google Scholar] [CrossRef]
- Israel, L.; Furer, V.; Levin-Zaidman, S.; Dezorella, N.; Brontvein, O.; Ablin, J.N.; Gross, A. Mitochondrial structural alterations in fibromyalgia: A pilot electron microscopy study. Clin. Exp. Rheumatol. 2024, 42, 1215–1223. [Google Scholar] [CrossRef]
- Fanton, S.; Menezes, J.; Krock, E.; Sandström, A.; Tour, J.; Sandor, K.; Jurczak, A.; Hunt, M.; Baharpoor, A.; Kadetoff, D.; et al. Anti-satellite glia cell IgG antibodies in fibromyalgia patients are related to symptom severity and to metabolite concentrations in thalamus and rostral anterior cingulate cortex. Brain Behav. Immun. 2023, 114, 371–382. [Google Scholar] [CrossRef]
- Krock, E.; Morado-Urbina, C.E.; Menezes, J.; Hunt, M.A.; Sandström, A.; Kadetoff, D.; Tour, J.; Verma, V.; Kultima, K.; Haglund, L.; et al. Fibromyalgia patients with elevated levels of anti–satellite glia cell immunoglobulin G antibodies present with more severe symptoms. Pain 2023, 164, 1828–1840. [Google Scholar] [CrossRef]
- Tsamou, M.; Kremers, F.A.C.; Samaritakis, K.A.; Roggen, E.L. Identifying microRNAs Possibly Implicated in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome and Fibromyalgia: A Review. Int. J. Mol. Sci. 2024, 25, 9551. [Google Scholar] [CrossRef]
- Kosek, E.; Martinsen, S.; Gerdle, B.; Mannerkorpi, K.; Löfgren, M.; Bileviciute-Ljungar, I.; Fransson, P.; Schalling, M.; Ingvar, M.; Ernberg, M.; et al. The translocator protein gene is associated with symptom severity and cerebral pain processing in fibromyalgia. Brain Behav. Immun. 2016, 58, 218–227. [Google Scholar] [CrossRef]
- Katar, M.; Deveci, H.; Deveci, K. Evaluation of clinical relationship of serum niacin and dopamine levels in patients with fibromyalgia syndrome. Turk. J. Phys. Med. Rehabil. 2022, 68, 84–90. [Google Scholar] [CrossRef]
- Wood, P.B.; Glabus, M.F.; Simpson, R.; Patterson, J.C. Changes in Gray Matter Density in Fibromyalgia: Correlation with Dopamine Metabolism. J. Pain 2009, 10, 609–618. [Google Scholar] [CrossRef]
- Filograna, R.; Lee, S.; Tiklova, K.; Mennuni, M.; Jonsson, V.; Ringner, M.; Gillberg, L.; Sopova, E.; Shupliakov, O.; Koolmeister, C.; et al. Mitochondrial dysfunction in adult midbrain dopamine neurons triggers an early immune response. PLoS Genet. 2021, 17, e1009822. [Google Scholar] [CrossRef]
- Gibbons, C.H. Chapter 27—Basics of Autonomic Nervous System Function. In Handbook of Clinical Neurology; Levin, K.H., Chauvel, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 160, pp. 407–418. [Google Scholar]
- Alves, R.L.; Zortea, M.; Serrano, P.V.; Brugnera Tomedi, R.; Pereira de Almeida, R.; Torres, I.L.S.; Fregni, F.; Caumo, W. High-beta oscillations at EEG resting state and hyperconnectivity of pain circuitry in fibromyalgia: An exploratory cross-sectional study. Front. Neurosci. 2023, 17, 1233979. [Google Scholar] [CrossRef]
- Craig, A.D. Interoception: The sense of the physiological condition of the body. Curr. Opin. Neurobiol. 2003, 13, 500–505. [Google Scholar] [CrossRef]
- Bergeron, D.; Obaid, S.; Fournier-Gosselin, M.P.; Bouthillier, A.; Nguyen, D.K. Deep Brain Stimulation of the Posterior Insula in Chronic Pain: A Theoretical Framework. Brain Sci. 2021, 11, 639. [Google Scholar] [CrossRef]
- Hirao, A.; Hojo, Y.; Murakami, G.; Ito, R.; Hashizume, M.; Murakoshi, T.; Uozumi, N. Effects of systemic inflammation on the network oscillation in the anterior cingulate cortex and cognitive behavior. PLoS ONE 2024, 19, e0302470. [Google Scholar] [CrossRef]
- Dong, D.; Li, C.; Zhong, X.; Gao, Y.; Cheng, C.; Sun, X.; Xiong, G.; Ming, Q.; Zhang, X.; Wang, X.; et al. Neuroticism modulates neural activities of posterior cingulate cortex and thalamus during psychosocial stress processing. J. Affect. Disord. 2020, 262, 223–228. [Google Scholar] [CrossRef]
- Smith, J.A.; Ji, Y.; Lorsung, R.; Breault, M.S.; Koenig, J.; Cramer, N.; Masri, R.; Keller, A. Parabrachial Nucleus Activity in Nociception and Pain in Awake Mice. J. Neurosci. 2023, 43, 5656–5667. [Google Scholar] [CrossRef]
- Chu, H.; Sun, J.; Xu, H.; Niu, Z.; Xu, M. Effect of periaqueductal gray melanocortin 4 receptor in pain facilitation and glial activation in rat model of chronic constriction injury. Neurol. Res. 2012, 34, 871–888. [Google Scholar] [CrossRef]
- Liu, X.; He, J.; Jiang, W.; Wen, S.; Xiao, Z. The Roles of Periaqueductal Gray and Dorsal Raphe Nucleus Dopaminergic Systems in the Mechanisms of Thermal Hypersensitivity and Depression in Mice. J. Pain 2023, 24, 1213–1228. [Google Scholar] [CrossRef]
- Yu, R.; Gollub, R.L.; Spaeth, R.; Napadow, V.; Wasan, A.; Kong, J. Disrupted functional connectivity of the periaqueductal gray in chronic low back pain. NeuroImage Clin. 2014, 6, 100–108. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Region | FM Stim | FM Rest | HC Stim | HC Rest |
|---|---|---|---|---|
| IC | 0.35 ± 0.30 | 0.22 ± 0.26 | 0.18 ± 0.24 | 0.20 ± 0.25 |
| PCC | 0.34 ± 0.26 | 0.17 ± 0.30 | 0.14 ± 0.26 | 0.19 ± 0.26 |
| FOrb | 0.27 ± 0.25 | 0.19 ± 0.27 | 0.16 ± 0.28 | 0.14 ± 0.25 |
| ACC | 0.34 ± 0.32 | 0.19 ± 0.31 | 0.15 ± 0.26 | 0.16 ± 0.28 |
| PAG | 0.48 ± 0.49 | 0.24 ± 0.38 | 0.29 ± 0.47 | 0.33 ± 0.43 |
| Hypothalamus | 0.29 ± 0.36 | 0.15 ± 0.45 | 0.18 ± 0.33 | 0.22 ± 0.33 |
| Thalamus | 0.33 ± 0.32 | 0.20 ± 0.27 | 0.16 ± 0.26 | 0.23 ± 0.29 |
| PBN | 0.40 ± 0.35 | 0.21 ± 0.39 | 0.29 ± 0.32 | 0.29 ± 0.33 |
| Amygdala | 0.30 ± 0.37 | 0.18 ± 0.48 | 0.14 ± 0.32 | 0.22 ± 0.34 |
| Brain Region | FM Rest (df = 161) | HC Stim (df = 145) | HC Rest (df = 148) | |||
|---|---|---|---|---|---|---|
| t | p | t | p | t | p | |
| IC | 3.06 | 1.31 × 10−3 | 3.63 | 1.95 × 10−4 | 3.38 | 4.65 × 10−4 |
| PC | 4.00 | 4.80 × 10−5 | 4.65 | 3.67 × 10−6 | 3.65 | 1.80 × 10−4 |
| FOrb | 1.96 | 2.61 × 10−2 | 2.48 | 7.19 × 10−3 | 3.28 | 6.57 × 10−4 |
| AC | 3.04 | 1.39 × 10−3 | 3.96 | 5.79 × 10−5 | 3.74 | 1.32 × 10−4 |
| PAG | 3.39 | 4.43 × 10−4 | 2.26 | 1.25 × 10−2 | 1.93 | 2.75 × 10−2 |
| Hypothalamus | 2.26 | 1.25 × 10−2 | 1.92 | 2.87 × 10−2 | 1.23 | 1.10 × 10−1 |
| Thalamus | 2.65 | 4.42 × 10−3 | 3.34 | 5.35 × 10−4 | 1.9 | 2.97 × 10−2 |
| PBN | 3.31 | 5.82 × 10−4 | 2.04 | 2.18 × 10−2 | 2.06 | 2.03 × 10−2 |
| Amygdala | 1.69 | 4.69 × 10−2 | 2.56 | 5.78 × 10−3 | 1.25 | 1.06 × 10−1 |
| Brain Region | HC Stim (df = 140) | HC Rest (df = 143) | ||
|---|---|---|---|---|
| t | p | t | p | |
| IC | 0.73 | 0.23 | 0.45 | 0.33 |
| PC | 0.57 | 0.28 | −0.41 | 0.66 |
| FOrb | 0.61 | 0.27 | 1.25 | 0.11 |
| AC | 0.9 | 0.19 | 0.7 | 0.24 |
| PAG | −0.74 | 0.77 | −1.29 | 0.90 |
| Hypothalamus | −0.49 | 0.69 | −1.11 | 0.87 |
| Thalamus | 0.94 | 0.17 | −0.58 | 0.72 |
| PBN | −1.27 | 0.90 | −1.25 | 0.89 |
| Amygdala | 0.53 | 0.30 | −0.55 | 0.71 |
| Brain Region | HC Rest (df = 127) | |
|---|---|---|
| t | p | |
| IC | −0.28 | 0.61 |
| PC | −1.01 | 0.84 |
| FOrb | 0.56 | 0.29 |
| AC | −0.19 | 0.58 |
| PAG | −0.43 | 0.66 |
| Hypothalamus | −0.71 | 0.76 |
| Thalamus | −1.42 | 0.92 |
| PBN | 0.03 | 0.49 |
| Amygdala | −1.30 | 0.90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Will, M.; Stroman, P.W. Exploring Neural Signaling Patterns and Their Physiological Origins in Fibromyalgia by Means of Functional MRI Guided by a Review of the Literature. Brain Sci. 2025, 15, 603. https://doi.org/10.3390/brainsci15060603
Will M, Stroman PW. Exploring Neural Signaling Patterns and Their Physiological Origins in Fibromyalgia by Means of Functional MRI Guided by a Review of the Literature. Brain Sciences. 2025; 15(6):603. https://doi.org/10.3390/brainsci15060603
Chicago/Turabian StyleWill, Mara, and Patrick W. Stroman. 2025. "Exploring Neural Signaling Patterns and Their Physiological Origins in Fibromyalgia by Means of Functional MRI Guided by a Review of the Literature" Brain Sciences 15, no. 6: 603. https://doi.org/10.3390/brainsci15060603
APA StyleWill, M., & Stroman, P. W. (2025). Exploring Neural Signaling Patterns and Their Physiological Origins in Fibromyalgia by Means of Functional MRI Guided by a Review of the Literature. Brain Sciences, 15(6), 603. https://doi.org/10.3390/brainsci15060603