
Resting Electroencephalography Microstates and Alpha Power Modulation in Preschool-Aged Children with Autism Spectrum Disorder



Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure
2.3. EEG Data Acquisition and Preprocessing
2.4. EEG Microstate Analysis
2.5. Characterization of Alpha Activities
2.6. Statistical Analysis
3. Results
3.1. Abnormal Aperiodic-Adjusted Alpha Activity and Microstate Parameters in ASD
3.1.1. Aperiodic-Adjusted Alpha Activity
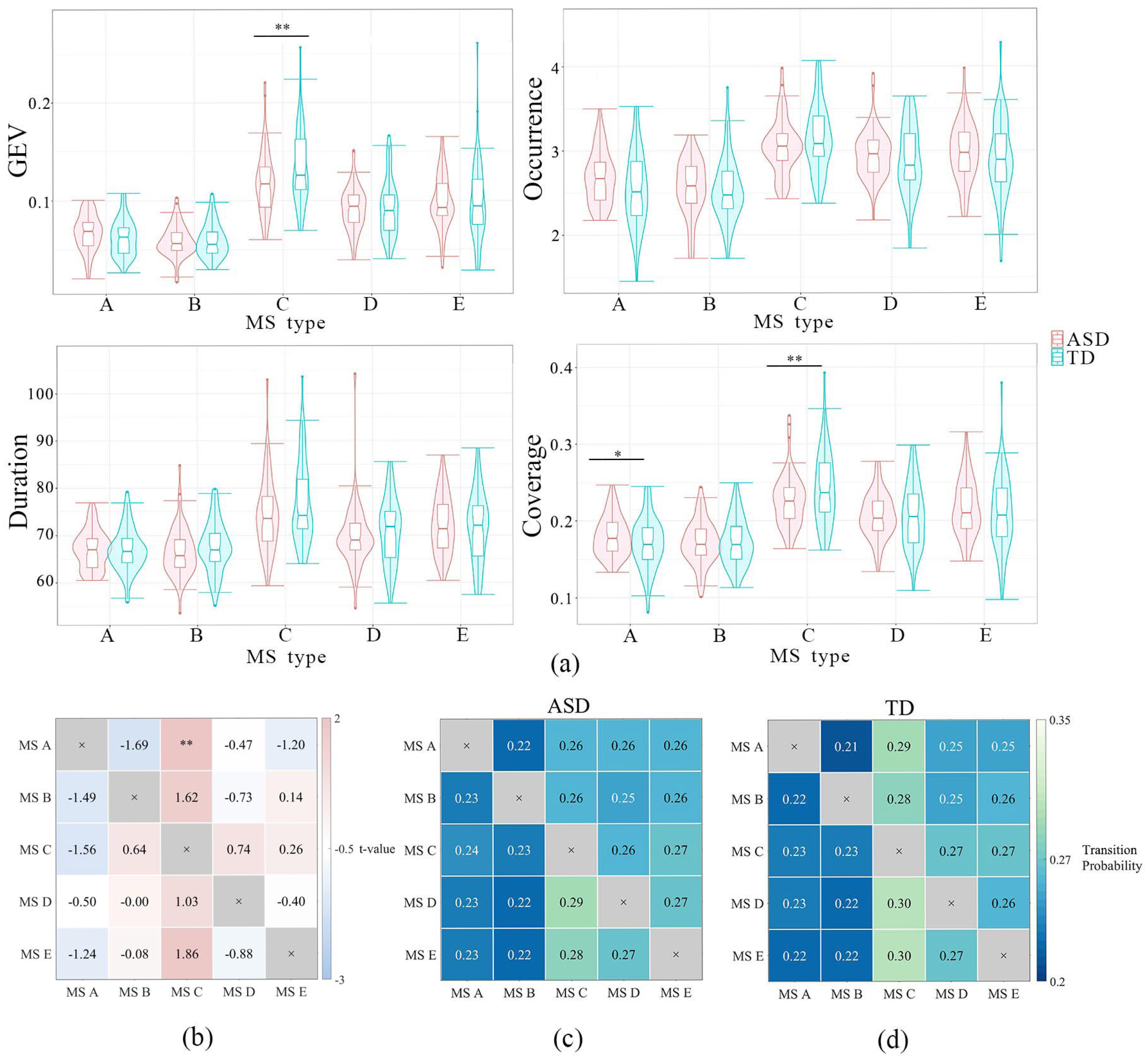
3.1.2. Microstate (MS) Analysis
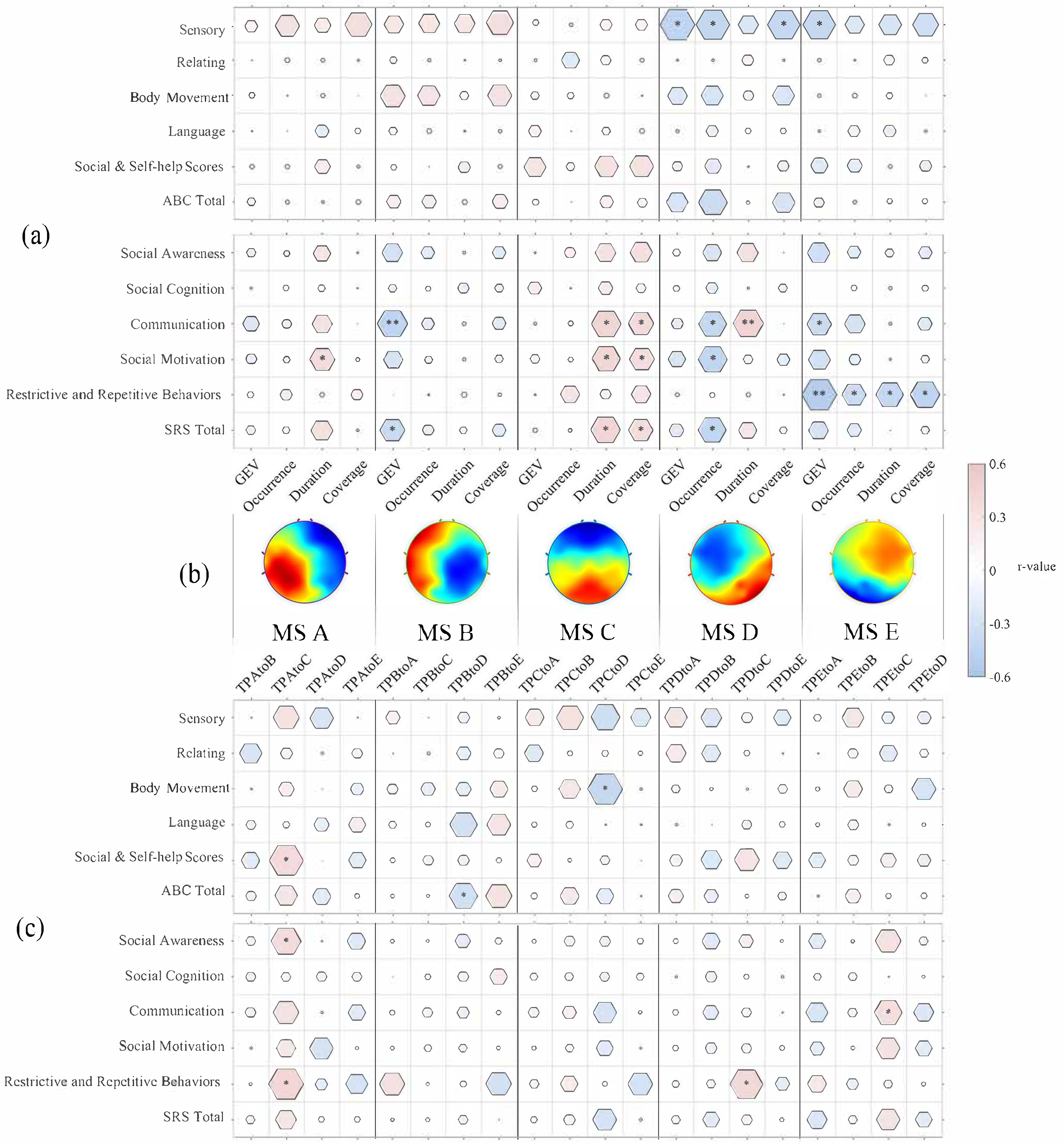
3.2. Aberrant Temporal Dynamics of Microstates Related to ASD Symptoms and Social Performance
3.2.1. Microstate A
3.2.2. Microstate B
3.2.3. Microstate C
3.2.4. Microstate D
3.2.5. Microstate E
3.3. Independence of the Effect of Microstates on ASD Symptoms
3.3.1. Alpha Activity Modulated Microstates
3.3.2. Semi-Partial Correlation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ASD | Autism spectrum disorder |
| EEG | Electroencephalography |
| ABC | Autism Behavior Checklist |
| SRS | Social Responsiveness Scale |
| MS | Microstate |
| TP | Transition probability |
| GEV | Global explained variance |
| GFP | Global field power |
References
- Sarmiento, C.; Lau, C. Diagnostic and Statistical Manual of Mental Disorders. In The Wiley Encyclopedia of Personality and Individual Differences, 5th ed.; Carducci, B.J., Nave, C.S., Fabio, A., Saklofske, D.H., Stough, C., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 125–129. ISBN 978-1-119-05753-6. [Google Scholar]
- Di Martino, A.; Ross, K.; Uddin, L.Q.; Sklar, A.B.; Castellanos, F.X.; Milham, M.P. Functional Brain Correlates of Social and Nonsocial Processes in Autism Spectrum Disorders: An Activation Likelihood Estimation Meta-Analysis. Biol. Psychiatry 2009, 65, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, M.V.; Pierce, K.; Eyler, L.T.; Barnes, C.C.; Ahrens-Barbeau, C.; Solso, S.; Campbell, K.; Courchesne, E. Different Functional Neural Substrates for Good and Poor Language Outcome in Autism. Neuron 2015, 86, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, E.; Lowry, S.J.; Santhosh, M.; Kresse, A.; Edwards, L.A.; Keller, J.; Libsack, E.J.; Kang, V.Y.; Naples, A.; Jack, A.; et al. Resting State EEG in Youth with ASD: Age, Sex, and Relation to Phenotype. J. Neurodev. Disord. 2021, 13, 33. [Google Scholar] [CrossRef]
- Kana, R.K.; Maximo, J.O.; Williams, D.L.; Keller, T.A.; Schipul, S.E.; Cherkassky, V.L.; Minshew, N.J.; Just, M.A. Aberrant Functioning of the Theory-of-Mind Network in Children and Adolescents with Autism. Mol. Autism 2015, 6, 59. [Google Scholar] [CrossRef]
- Khalil, R.; Tindle, R.; Boraud, T.; Moustafa, A.A.; Karim, A.A. Social Decision Making in Autism: On the Impact of Mirror Neurons, Motor Control, and Imitative Behaviors. CNS Neurosci. Ther. 2018, 24, 669–676. [Google Scholar] [CrossRef]
- Abbott, A.E.; Nair, A.; Keown, C.L.; Datko, M.; Jahedi, A.; Fishman, I.; Müller, R.-A. Patterns of Atypical Functional Connectivity and Behavioral Links in Autism Differ Between Default, Salience, and Executive Networks. Cereb. Cortex 2016, 26, 4034–4045. [Google Scholar] [CrossRef]
- He, C.; Chen, Y.; Jian, T.; Chen, H.; Guo, X.; Wang, J.; Wu, L.; Chen, H.; Duan, X. Dynamic Functional Connectivity Analysis Reveals Decreased Variability of the Default-mode Network in Developing Autistic Brain. Autism Res. 2018, 11, 1479–1493. [Google Scholar] [CrossRef]
- Itahashi, T.; Yamada, T.; Watanabe, H.; Nakamura, M.; Ohta, H.; Kanai, C.; Iwanami, A.; Kato, N.; Hashimoto, R. Alterations of Local Spontaneous Brain Activity and Connectivity in Adults with High-Functioning Autism Spectrum Disorder. Mol. Autism 2015, 6, 30. [Google Scholar] [CrossRef]
- Nijhof, A.D.; Bardi, L.; Brass, M.; Wiersema, J.R. Brain Activity for Spontaneous and Explicit Mentalizing in Adults with Autism Spectrum Disorder: An fMRI Study. NeuroImage Clin. 2018, 18, 475–484. [Google Scholar] [CrossRef]
- Michel, C.M.; Koenig, T. EEG Microstates as a Tool for Studying the Temporal Dynamics of Whole-Brain Neuronal Networks: A Review. NeuroImage 2018, 180, 577–593. [Google Scholar] [CrossRef]
- Mishra, A.; Englitz, B.; Cohen, M.X. EEG Microstates as a Continuous Phenomenon. NeuroImage 2020, 208, 116454. [Google Scholar] [CrossRef] [PubMed]
- Britz, J.; Van De Ville, D.; Michel, C.M. BOLD Correlates of EEG Topography Reveal Rapid Resting-State Network Dynamics. NeuroImage 2010, 52, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Koenig, T.; Lehmann, D.; Merlo, M.C.G.; Kochi, K.; Hell, D.; Koukkou, M. A Deviant EEG Brain Microstate in Acute, Neuroleptic-Naive Schizophrenics at Rest. Eur. Arch. Psychiatry Clin. Neurosci. 1999, 249, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, D.; Faber, P.L.; Galderisi, S.; Herrmann, W.M.; Kinoshita, T.; Koukkou, M.; Mucci, A.; Pascual-Marqui, R.D.; Saito, N.; Wackermann, J.; et al. EEG Microstate Duration and Syntax in Acute, Medication-Naïve, First-Episode Schizophrenia: A Multi-Center Study. Psychiatry Res. Neuroimaging 2005, 138, 141–156. [Google Scholar] [CrossRef]
- Poulsen, A.T.; Pedroni, A.; Langer, N.; Hansen, L.K. Microstate EEGlab Toolbox: An Introductory Guide. BioRxiv 2018, 289850. [Google Scholar] [CrossRef]
- Custo, A.; Van De Ville, D.; Wells, W.M.; Tomescu, M.I.; Brunet, D.; Michel, C.M. Electroencephalographic Resting-State Networks: Source Localization of Microstates. Brain Connect. 2017, 7, 671–682. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D.; Biscay, R.J.; Bosch-Bayard, J.; Lehmann, D.; Kochi, K.; Kinoshita, T.; Yamada, N.; Sadato, N. Assessing Direct Paths of Intracortical Causal Information Flow of Oscillatory Activity with the Isolated Effective Coherence (iCoh). Front. Hum. Neurosci. 2014, 8, 448. [Google Scholar] [CrossRef]
- Seitzman, B.A.; Abell, M.; Bartley, S.C.; Erickson, M.A.; Bolbecker, A.R.; Hetrick, W.P. Cognitive Manipulation of Brain Electric Microstates. NeuroImage 2017, 146, 533–543. [Google Scholar] [CrossRef]
- Milz, P.; Faber, P.L.; Lehmann, D.; Koenig, T.; Kochi, K.; Pascual-Marqui, R.D. The Functional Significance of EEG Microstates—Associations with Modalities of Thinking. NeuroImage 2016, 125, 643–656. [Google Scholar] [CrossRef]
- Koenig, T.; Prichep, L.; Lehmann, D.; Sosa, P.V.; Braeker, E.; Kleinlogel, H.; Isenhart, R.; John, E.R. Millisecond by Millisecond, Year by Year: Normative EEG Microstates and Developmental Stages. NeuroImage 2002, 16, 41–48. [Google Scholar] [CrossRef]
- Bochet, A.; Sperdin, H.F.; Rihs, T.A.; Kojovic, N.; Franchini, M.; Jan, R.K.; Michel, C.M.; Schaer, M. Early Alterations of Large-Scale Brain Networks Temporal Dynamics in Young Children with Autism. Commun. Biol. 2021, 4, 968. [Google Scholar] [CrossRef] [PubMed]
- Zanesco, A.P. Normative Temporal Dynamics of Resting EEG Microstates. Brain Topogr. 2024, 37, 243–264. [Google Scholar] [CrossRef]
- Das, S.; Zomorrodi, R.; Enticott, P.G.; Kirkovski, M.; Blumberger, D.M.; Rajji, T.K.; Desarkar, P. Resting State Electroencephalography Microstates in Autism Spectrum Disorder: A Mini-Review. Front. Psychiatry 2022, 13, 988939. [Google Scholar] [CrossRef]
- Takarae, Y.; Zanesco, A.; Keehn, B.; Chukoskie, L.; Müller, R.; Townsend, J. EEG Microstates Suggest Atypical Resting-State Network Activity in High-Functioning Children and Adolescents with Autism Spectrum Development. Dev. Sci. 2022, 25, e13231. [Google Scholar] [CrossRef]
- Antonova, E.; Holding, M.; Suen, H.C.; Sumich, A.; Maex, R.; Nehaniv, C. EEG Microstates: Functional Significance and Short-Term Test-Retest Reliability. Neuroimage Rep. 2022, 2, 100089. [Google Scholar] [CrossRef]
- Spadone, S.; Croce, P.; Zappasodi, F.; Capotosto, P. Pre-Stimulus EEG Microstates Correlate With Anticipatory Alpha Desynchronization. Front. Hum. Neurosci. 2020, 14, 182. [Google Scholar] [CrossRef]
- Von Wegner, F.; Bauer, S.; Rosenow, F.; Triesch, J.; Laufs, H. EEG Microstate Periodicity Explained by Rotating Phase Patterns of Resting-State Alpha Oscillations. NeuroImage 2021, 224, 117372. [Google Scholar] [CrossRef]
- Dickinson, A.; DiStefano, C.; Senturk, D.; Jeste, S.S. Peak Alpha Frequency Is a Neural Marker of Cognitive Function across the Autism Spectrum. Eur. J. Neurosci. 2018, 47, 643–651. [Google Scholar] [CrossRef]
- Lefebvre, A.; Delorme, R.; Delanoë, C.; Amsellem, F.; Beggiato, A.; Germanaud, D.; Bourgeron, T.; Toro, R.; Dumas, G. Alpha Waves as a Neuromarker of Autism Spectrum Disorder: The Challenge of Reproducibility and Heterogeneity. Front. Neurosci. 2018, 12, 662. [Google Scholar] [CrossRef]
- Adelhöfer, N.; Paulus, T.; Mückschel, M.; Bäumer, T.; Bluschke, A.; Takacs, A.; Tóth-Fáber, E.; Tárnok, Z.; Roessner, V.; Weissbach, A.; et al. Increased Scale-Free and Aperiodic Neural Activity during Sensorimotor Integration—A Novel Facet in Tourette Syndrome. Brain Commun. 2021, 3, fcab250. [Google Scholar] [CrossRef]
- Donoghue, T.; Haller, M.; Peterson, E.J.; Varma, P.; Sebastian, P.; Gao, R.; Noto, T.; Lara, A.H.; Wallis, J.D.; Knight, R.T.; et al. Parameterizing Neural Power Spectra into Periodic and Aperiodic Components. Nat. Neurosci. 2020, 23, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.T.; Clark, G.M.; Bigelow, F.J.; Lum, J.A.G.; Enticott, P.G. Periodic and Aperiodic Neural Activity Displays Age-Dependent Changes across Early-to-Middle Childhood. Dev. Cogn. Neurosci. 2022, 54, 101076. [Google Scholar] [CrossRef] [PubMed]
- Merkin, A.; Sghirripa, S.; Graetz, L.; Smith, A.E.; Hordacre, B.; Harris, R.; Pitcher, J.; Semmler, J.; Rogasch, N.C.; Goldsworthy, M. Do Age-Related Differences in Aperiodic Neural Activity Explain Differences in Resting EEG Alpha? Neurobiol. Aging 2023, 121, 78–87. [Google Scholar] [CrossRef]
- Rico-Picó, J.; Moyano, S.; Conejero, Á.; Hoyo, Á.; Ballesteros-Duperón, M.Á.; Rueda, M.R. Early Development of Electrophysiological Activity: Contribution of Periodic and Aperiodic Components of the EEG Signal. Psychophysiology 2023, 60, e14360. [Google Scholar] [CrossRef]
- Hill, A.T.; Bailey, N.W.; Zomorrodi, R.; Hadas, I.; Kirkovski, M.; Das, S.; Lum, J.A.G.; Enticott, P.G. EEG Microstates in Early-to-middle Childhood Show Associations with Age, Biological Sex, and Alpha Power. Hum. Brain Mapp. 2023, 44, 6484–6498. [Google Scholar] [CrossRef]
- Durka, P.J.; Klekowicz, H.; Blinowska, K.J.; Szelenberger, W.; Niemcewicz, S. A Simple System for Detection of EEG Artifacts in Polysomnographic Recordings. IEEE Trans. Biomed. Eng. 2003, 50, 526–528. [Google Scholar] [CrossRef]
- Kindler, J.; Hubl, D.; Strik, W.K.; Dierks, T.; Koenig, T. Resting-State EEG in Schizophrenia: Auditory Verbal Hallucinations Are Related to Shortening of Specific Microstates. Clin. Neurophysiol. 2011, 122, 1179–1182. [Google Scholar] [CrossRef]
- Tomescu, M.I.; Papasteri, C.C.; Sofonea, A.; Boldasu, R.; Kebets, V.; Pistol, C.A.D.; Poalelungi, C.; Benescu, V.; Podina, I.R.; Nedelcea, C.I.; et al. Spontaneous Thought and Microstate Activity Modulation by Social Imitation. NeuroImage 2022, 249, 118878. [Google Scholar] [CrossRef]
- Faber, P.L.; Travis, F.; Milz, P.; Parim, N. EEG Microstates during Different Phases of Transcendental Meditation Practice. Cogn. Process. 2017, 18, 307–314. [Google Scholar] [CrossRef]
- Katayama, H.; Gianotti, L.R.R.; Isotani, T.; Faber, P.L.; Sasada, K.; Kinoshita, T.; Lehmann, D. Classes of Multichannel EEG Microstates in Light and Deep Hypnotic Conditions. Brain Topogr. 2007, 20, 7–14. [Google Scholar] [CrossRef]
- Jia, H.; Yu, D. Aberrant Intrinsic Brain Activity in Patients with Autism Spectrum Disorder: Insights from EEG Microstates. Brain Topogr. 2019, 32, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, D.; Ozaki, H.; Pal, I. EEG Alpha Map Series: Brain Micro-States by Space-Oriented Adaptive Segmentation. Electroencephalogr. Clin. Neurophysiol. 1987, 67, 271–288. [Google Scholar] [CrossRef]
- Lombardo, M.V.; Eyler, L.; Moore, A.; Datko, M.; Carter Barnes, C.; Cha, D.; Courchesne, E.; Pierce, K. Default Mode-Visual Network Hypoconnectivity in an Autism Subtype with Pronounced Social Visual Engagement Difficulties. eLife 2019, 8, e47427. [Google Scholar] [CrossRef]
- Gui, A.; Bussu, G.; Tye, C.; Elsabbagh, M.; Pasco, G.; Charman, T.; Johnson, M.H.; Jones, E.J.H. Attentive Brain States in Infants with and without Later Autism. Transl. Psychiatry 2021, 11, 196. [Google Scholar] [CrossRef]
- Miller, L.J.; Anzalone, M.E.; Lane, S.J.; Cermak, S.A.; Osten, E.T. Concept Evolution in Sensory Integration: A Proposed Nosology for Diagnosis. Am. J. Occup. Ther. 2007, 61, 135–140. [Google Scholar] [CrossRef]
- Baranek, G.T.; Boyd, B.A.; Poe, M.D.; David, F.J.; Watson, L.R. Hyperresponsive Sensory Patterns in Young Children With Autism, Developmental Delay, and Typical Development. Am. J. Ment. Retard. 2007, 112, 233. [Google Scholar] [CrossRef]
- Leekam, S.R.; Nieto, C.; Libby, S.J.; Wing, L.; Gould, J. Describing the Sensory Abnormalities of Children and Adults with Autism. J. Autism Dev. Disord. 2007, 37, 894–910. [Google Scholar] [CrossRef]
- Hilton, C.L.; Harper, J.D.; Kueker, R.H.; Lang, A.R.; Abbacchi, A.M.; Todorov, A.; LaVesser, P.D. Sensory Responsiveness as a Predictor of Social Severity in Children with High Functioning Autism Spectrum Disorders. J. Autism Dev. Disord. 2010, 40, 937–945. [Google Scholar] [CrossRef]
- Baranek, G.T.; Foster, L.G.; Berkson, G. Tactile Defensiveness and Stereotyped Behaviors. Am. J. Occup. Ther. 1997, 51, 91–95. [Google Scholar] [CrossRef]
- Keehn, B.; Shih, P.; Brenner, L.A.; Townsend, J.; Müller, R.-A. Functional Connectivity for an “Island of Sparing” in Autism Spectrum Disorder: An fMRI Study of Visual Search: FCMRI for Visual Search in Autism. Hum. Brain Mapp. 2013, 34, 2524–2537. [Google Scholar] [CrossRef]
- Cooper, N.R.; Burgess, A.P.; Croft, R.J.; Gruzelier, J.H. Investigating Evoked and Induced Electroencephalogram Activity in Task-Related Alpha Power Increases during an Internally Directed Attention Task. NeuroReport 2006, 17, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Pani, S.M.; Saba, L.; Fraschini, M. Clinical Applications of EEG Power Spectra Aperiodic Component Analysis: A Mini-Review. Clin. Neurophysiol. 2022, 143, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ostlund, B.D.; Alperin, B.R.; Drew, T.; Karalunas, S.L. Behavioral and Cognitive Correlates of the Aperiodic (1/f-like) Exponent of the EEG Power Spectrum in Adolescents with and without ADHD. Dev. Cogn. Neurosci. 2021, 48, 100931. [Google Scholar] [CrossRef] [PubMed]
- Shuffrey, L.C.; Pini, N.; Potter, M.; Springer, P.; Lucchini, M.; Rayport, Y.; Sania, A.; Firestein, M.; Brink, L.; Isler, J.R.; et al. Aperiodic Electrophysiological Activity in Preterm Infants Is Linked to Subsequent Autism Risk. Dev. Psychobiol. 2022, 64, e22271. [Google Scholar] [CrossRef]
- Farokhzadi, F.; Mohamadi, M.R.; Khajevand Khosli, A.; Akbarfahim, M.; Ali Beigi, N.; Torabi, P. Comparing the Effectiveness of the Transcranial Alternating Current Stimulation (TACS) and Ritalin on Symptoms of Attention Deficit Hyperactivity Disorder in 7-14-Year-Old Children. Acta Med. Iran. 2021, 58, 638–648. [Google Scholar] [CrossRef]
- Jones, K.T.; Johnson, E.L.; Tauxe, Z.S.; Rojas, D.C. Modulation of Auditory Gamma-Band Responses Using Transcranial Electrical Stimulation. J. Neurophysiol. 2020, 123, 2504–2514. [Google Scholar] [CrossRef]
- Kayarian, F.B.; Jannati, A.; Rotenberg, A.; Santarnecchi, E. Targeting Gamma-Related Pathophysiology in Autism Spectrum Disorder Using Transcranial Electrical Stimulation: Opportunities and Challenges. Autism Res. 2020, 13, 1051–1071. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Characteristic | ASD | TD |
|---|---|---|
| Age (M ± SD) | 4.59 ± 1.00 | 4.64 ± 0.97 |
| Gender (male/female) | 47/12 | 47/12 |
| ABC score (M ± SD) | 51.13 ± 16.39 | – |
| SRS score (M ± SD) | 100.61 ± 22.26 | – |
| MS Type | MS Parameters | Correlation | Clinical Scores | |||||
|---|---|---|---|---|---|---|---|---|
| Sensory | Relating | Body Movement | Language | Social and Self-Help | ABC Total Score | |||
| A | Occurrence | r | 0.29 | −0.06 | 0.02 | −0.01 | −0.06 | 0.26 |
| semi-r | 0.32 | −0.03 | 0.05 | 0.01 | −0.03 | 0.09 | ||
| Duration | r | 0.2 | 0.05 | −0.06 | −0.17 | 0.19 | −0.02 | |
| semi-r | 0.18 | 0.02 | −0.09 | −0.19 | 0.17 | 0 | ||
| C | Occurrence | r | 0.05 | −0.22 | 0.09 | −0.01 | 0.11 | 0.07 |
| semi-r | 0.04 | −0.23 | 0.08 | −0.02 | 0.1 | −0.02 | ||
| Duration | r | 0.15 | 0.13 | −0.07 | 0.11 | 0.29 | −0.09 | |
| semi-r | 0.18 | 0.18 | −0.02 | 0.13 | 0.33 | 0.22 | ||
| D | Occurrence | r | −0.41 | −0.04 | −0.27 | −0.15 | −0.2 | −0.29 |
| semi-r | −0.4 | 0.01 | −0.24 | −0.13 | −0.18 | −0.31 | ||
| Duration | r | −0.25 | 0.15 | −0.13 | 0.08 | 0.02 | −0.08 | |
| semi-r | −0.28 | 0.11 | −0.17 | 0.06 | −0.01 | −0.09 | ||
| E | Occurrence | r | −0.24 | 0.02 | −0.06 | 0.15 | −0.18 | 0.02 |
| semi-r | −0.28 | −0.03 | −0.12 | 0.13 | −0.23 | −0.11 | ||
| Duration | r | −0.28 | 0.14 | 0.08 | −0.16 | −0.05 | −0.14 | |
| semi-r | −0.34 | 0.07 | 0.01 | −0.21 | −0.11 | −0.16 | ||
| Social Awareness | Social Cognition | Communication | Social Motivation | Restrictive and Repetitive Behaviors | SRS Total Score | |||
| A | Occurrence | r | −0.09 | −0.09 | −0.14 | −0.1 | 0.17 | 0.04 |
| semi-r | −0.08 | 0.01 | −0.1 | −0.03 | 0.12 | −0.05 | ||
| Duration | r | 0.25 | 0.11 | 0.29 | 0.36 * | 0.07 | 0.21 | |
| semi-r | 0.25 | 0.09 | 0.28 | 0.35 * | 0.08 | 0.31 | ||
| C | Occurrence | r | 0.16 | −0.02 | 0.08 | 0.08 | 0.27 | 0.16 |
| semi-r | 0.16 | −0.05 | 0.07 | 0.06 | 0.29 | 0.05 | ||
| Duration | r | 0.29 | 0.21 | 0.43 * | 0.4 * | 0.16 | 0.40 * | |
| semi-r | 0.3 | 0.24 | 0.44 * | 0.43 * | 0.14 | 0.44 * | ||
| D | Occurrence | r | −0.11 | −0.09 | −0.17 | −0.25 | −0.06 | −0.23 |
| semi-r | −0.11 | −0.1 | −0.18 | −0.26 | −0.05 | −0.21 | ||
| Duration | r | −0.27 | −0.18 | −0.4 * | −0.42 * | −0.08 | −0.40 * | |
| semi-r | −0.26 | −0.14 | −0.38 * | −0.39 * | −0.11 | −0.39 * | ||
| E | Occurrence | r | −0.19 | 0.09 | −0.29 | −0.17 | −0.34 * | −0.19 |
| semi-r | −0.24 | 0 | −0.38 * | −0.27 | −0.32 | −0.3 | ||
| Duration | r | −0.13 | 0.05 | −0.05 | −0.03 | −0.39 * | −0.15 | |
| semi-r | −0.15 | −0.02 | −0.09 | −0.09 | −0.37 * | −0.04 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, M.; Yang, Z.; Wang, L.; Lu, S.; Han, J.; Li, X. Resting Electroencephalography Microstates and Alpha Power Modulation in Preschool-Aged Children with Autism Spectrum Disorder. Brain Sci. 2025, 15, 544. https://doi.org/10.3390/brainsci15060544
Ma M, Yang Z, Wang L, Lu S, Han J, Li X. Resting Electroencephalography Microstates and Alpha Power Modulation in Preschool-Aged Children with Autism Spectrum Disorder. Brain Sciences. 2025; 15(6):544. https://doi.org/10.3390/brainsci15060544
Chicago/Turabian StyleMa, Mingxuan, Ziying Yang, Leiyan Wang, Shan Lu, Junxia Han, and Xiaoli Li. 2025. "Resting Electroencephalography Microstates and Alpha Power Modulation in Preschool-Aged Children with Autism Spectrum Disorder" Brain Sciences 15, no. 6: 544. https://doi.org/10.3390/brainsci15060544
APA StyleMa, M., Yang, Z., Wang, L., Lu, S., Han, J., & Li, X. (2025). Resting Electroencephalography Microstates and Alpha Power Modulation in Preschool-Aged Children with Autism Spectrum Disorder. Brain Sciences, 15(6), 544. https://doi.org/10.3390/brainsci15060544