Abstract
Alzheimer’s disease (AD) represents a significant challenge among neurodegenerative disorders, as effective treatments and therapies remain largely undeveloped. Despite extensive research efforts employing various methodologies and diverse genetic models focused on amyloid-β (Aβ) pathology, the research for effective therapeutic strategies remains inconclusive. The key pathological features of AD include Aβ senile plaques, neurofibrillary tangles (NFTs), and the activation of neuroinflammatory pathways. Presently, investigations into AD and assessing potential treatments predominantly utilize Aβ transgenic models. Conversely, non-transgenic models may provide valuable insights into the multifaceted pathological states associated with AD. Thus, these models may serve as practical complementary tools for evaluating therapeutic and intervention strategies, since the primary AD risk factors are most frequently modeled. This review aims to critically assess the existing literature on AD non-transgenic models induced by streptozotocin, scopolamine, aging, mechanical stress, metals, and dietary patterns to enhance their application in AD research.
1. Introduction
Alzheimer’s disease (AD) is the most common type of dementia worldwide and accounts for up to 70% of all dementia diagnoses [1]. Its incidence escalates significantly with age, and AD is classically classified into two primary forms: sporadic AD (SAD) (or late-onset AD) and familial AD (FAD) (or early-onset AD) [2]. SAD constitutes the majority of AD cases (over 90%), and it typically manifests after the age of 65. SAD represents a multifactorial disorder in which both environmental factors and genetic predisposition could contribute to the genesis of the disease [2]. Many risk factors have been linked to the development of SAD, including genetic predisposition, aging, exposure to metals, head injury, diet, mitochondrial dysfunction, vascular disease, immune system dysfunction, infectious disease, and genetic predisposition [3].
In comparison, FAD accounts for less than 10% of AD cases, and it presents before the age of 65, often in the fourth or fifth decade of life. It is caused by autosomal dominant genetic mutations in the Amyloid β Precursor Protein (APP), Presenilin 1 (PSEN1), or Presenilin 2 (PSEN2) genes, and it exhibits a clear inheritance pattern [4].
Great efforts have been invested in understanding AD physiopathology, with early animal experiments playing a crucial role. The evolution of animal models reflects advancements in AD genetics and pathology. Initially, research focused on observing cognitive decline in aging animals. Before the modern era of genetic engineering, researchers relied on naturally aging animals, such as dogs and primates, to study age-related cognitive changes [5,6]. These studies provided initial insights into the behavioral and neuropathological aspects of brain aging. The development of transgenic rodent models marked a significant turning point [7]. Researchers began creating mice that expressed human genes linked to AD, allowing the study of specific pathological features like amyloid plaques and tau protein pathology [8].
In this sense, murine animal models of AD have been established to resemble key histopathological and cognitive impairments [9]. Although many transgenic AD models have been developed to study pathological changes and novel innovative treatments, they are based mainly on FAD. Transgenic models express a mutated form of human APP in addition to a series of mutations in the enzymes that process APP [10]. Thus, the modifications mentioned result in the release of different isoforms of amyloid β (Aβ) peptide [11]. Therefore, the genetic backgrounds of transgenic mice may confound crucial aspects of AD that might not relate to sporadic forms of the disease.
Although FAD shares histopathological characteristics with SAD, including the increase in the deposition of Aβ peptides and the hyperphosphorylation of tau protein in the brains of AD patients [12], most AD cases belong to SAD.
The mechanisms driving the establishment of SAD are more complex and varied than the dominant monogenic mutations that cause FAD [13]. For this reason, non-transgenic models related to SAD pathological changes, which mimic the risk factors, and the pathways involved in the pathogenesis of SAD, could increase the comprehension of the disease and the rate of success during the development and evaluation of novel compounds. In this review, we discuss the key characteristics of established non-transgenic murine models of AD concerning the primary risk factors. We also explore how pharmacologically induced rat models can help address the challenges faced in AD research and drug development.
2. Methods
A systematic review was conducted on published studies focused on creating non-transgenic animal models of AD related to the main risk factors. This review covered the pathological changes associated with AD and factors related to SAD. It provided a detailed explanation of non-transgenic animal models of AD, including models induced by streptozotocin, scopolamine, aging, mechanical stress, metals, and dietary patterns. Finally, the advantages and disadvantages of transgenic and non-transgenic AD models are discussed.
3. Results
3.1. Evolution of AD Definition
The initial definition of AD was based on clinical–pathological criteria [9]. The diagnosis in the living patient was considered probable AD and then confirmed at autopsy when characteristic neuropathological changes were found [14]. Over time, the concept became confusing, as two nosological entities of a different nature were described: a prototypical clinical syndrome without neuropathological verification and, on the other hand, the histopathological findings that confirmed the diagnosis. In the search for a more integrative definition, the National Institute on Aging and the Alzheimer’s Association proposed the following: “AD refers to deposits of pathological Aβ and tau plaques, defined in vivo by abnormal biomarkers of Aβ and pathological tau (both criteria are necessary)”. This definition includes the underlying pathological processes confirmed by post-mortem examination and the in vivo clinical manifestation evaluated by biomarkers [15]. This definition allows a greater understanding of the mechanisms and develops interventions that prevent or delay the initial onset of symptoms of the preclinical phase of AD.
3.2. Pathological Changes in AD
AD is characterized by alterations in learning and memory that are correlated with degeneration of several brain areas, including the hippocampus and cerebral cortex. Both FAD and SAD share histopathological characteristics, such as senile plaques composed principally of Aβ, neurofibrillary tangles (NFTs), and neuronal and synaptic loss [16].
3.2.1. Aβ and Its Pathological Forms
Aβ is a 4-kDa peptide produced by the proteolytic processing of APP [17]. APP is a transmembrane glycoprotein processed by the action of several proteases, following two processes that compete for the same part of the protein [18]. In the most common pathway, a protease known as α-secretase cleaves APP, releasing a soluble, extracellular fragment of about 695 amino acid residues. Then, the rest of the protein which is integrated into the membrane is processed by the γ-secretase, resulting in the release of the carboxyl terminal, which is transported into lysosomal vesicles for further degradation. This process occurs mainly in the healthy brain and is known as the non-amyloidogenic pathway because the action of α-secretase prevents the formation of the Aβ peptide (Figure 1) [19]. However, in the brains of AD patients, most of the APP is processed differently, following the amyloidogenic pathway. In this pathway, the APP is cleaved firstly by β-secretase (BACE1), resulting in the release of a longer carboxylic fragment of 99 amino acids, which is subsequently processed by the γ-secretase, resulting in the release of Aβ peptide (Figure 1). Once Aβ is released by the sequential action of β and γ secretases, it interacts with other monomers, resulting in the formation of soluble oligomers (Aβos) and insoluble fibrils (Aβfs) [20].
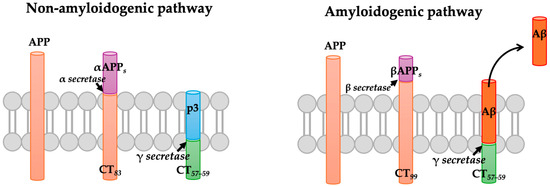
Figure 1.
The amyloid pathway. The amyloid precursor protein (APP) can be cleaved through two pathways. In the non-amyloidogenic pathway, APP is first cleaved by α-secretase and then by γ-secretase, resulting in the formation of a non-toxic fragment. In the amyloidogenic pathway, APP is cleaved by β-secretase, followed by γ-secretase, which ultimately leads to the formation of a neurotoxic Aβ peptide that tends to aggregate.
Senile plaques are composed principally of a central core of insoluble Aβf surrounded by abnormally formed neurites. Additionally, it is possible to find microglial cells and, less frequently, reactive astrocytes in the periphery of senile plaque.
In addition, Aβ can be found as focal diffuse deposits in diverse brain areas, including the cerebral cortex. Interestingly, these deposits are not accompanied by dystrophic neurites or surrounding activated glial cells. Diffuse plaques are not exclusive to AD; in elderly individuals, they can be seen in relatively large numbers of diffuse plaques in the brains of patients without cognitive impairment [21]. In addition, Aβ deposits can be detected in the walls of arterial vessels in the meninges and the cerebral cortex. When this kind of Aβ deposit is minimal, it does not impair the function of the vessels; however, when the deposits severely affect the vessels, there is a higher risk for spontaneous vascular rupture, resulting in localized blood accumulation in brain tissue.
In contrast, oAβ is generally considered the most neurotoxic and pathogenic form of Aβ [22]. It has been demonstrated that oAβ accumulation occurs early in AD development, including before plaque formation [23]. The consequence of increased oAβ production in the brain has been widely demonstrated in behavioral, neuropathological, and cell-biological studies [24].
Recent studies have shown that oAβ is capable of binding to various surface receptors, including advanced glycation product receptor (RAGE), α7-nicotinic acetylcholine receptor (α7 nAChR), N-methyl-d-aspartate receptor (NMDAR), and cellular prion protein (PrPc), among others, which are highly expressed in neurons and glial cells [25]. oAβ binding to RAGE promotes the development of neuronal and microglial toxicity by several pathways, including the development of oxidative stress, synaptic dysfunction, and, ultimately, neuronal cell death [26]. Indeed, it is shown that RAGE expression is significantly increased in the brain of AD patients, especially in the blood vessels [25]. In addition, oAβ interacts with NMDARs at the postsynaptic terminal, and their activation induces Ca2+ dysregulation, neuronal death, and synaptic dysfunction. Nevertheless, it remains uncertain whether oAβ directly interacts with NMDAR subunits [27]. The α7 nAChR is a potential receptor that binds to fAβ with high affinity. This receptor mediates the phosphorylation of tau proteins induced by Aβ through the activation of ERK and JNK signaling pathways. Additionally, Aβ can directly interact with cell membranes, forming ion channels or pores that lead to membrane disruption and, consequently, neuronal damage [28].
In contrast, fAβ induces neurotoxicity and dysfunction of synapses in AD brains by generating reactive oxygen species (ROS). ROS generation increases Aβ levels and promotes its deposit [29]. Interaction of fAβ with with Cu2+ ions triggers the production of ROS, which are favored by the high concentration of Cu2+ ions in the amyloid plaque deposits in the brains of individuals with AD. Once bound to fAβ, Cu2+ ions can directly convert physiological H2O2 to the hydroxyl radical (·OH). Several experiments proved that when the Cu2+ was reduced to Cu1+ by biological lowering agents such as vitamin C, cholesterol, and catecholamines, the Cu1+-Aβ complex generated ROS [30].
3.2.2. Phosphorylated Tau and NFTs
Tau is a microtubule-associated protein mainly distributed in the axon of the neurons of the healthy brain [31]. Tau regulates the enzymatic activity of phosphatases and kinases through signal transduction mechanisms [32]. Nevertheless, mutations in tau protein result in changes in its affinity to microtubules, changes in its conformation, alterations in posttranslational modifications, and increased aggregation propensity [33]. The pathologic tau changes in AD brains, including phosphorylation and increased aggregating ability, are related to brain damage [34]. In AD, hyperphosphorylated tau is associated with the formation of NFTs, which is related to conformational changes in the structure of tau [35]. Due to modifications in their structure, tau protein is redistributed in both the soma and dendrites of neurons [36]. Recent studies have also shown that tau is present in glial cells. The dynamics of tau and other microtubule-binding proteins, which attach to microtubules, are strictly regulated by various factors to ensure proper function [37]. When tau detaches from microtubules, it accumulates in neurites and neuronal cell bodies, leading to the formation of NFTs, which are significant pathological features of AD. The accumulation of paired helical filaments and NFTs disrupts normal physiological functions, resulting in apoptosis and neuronal loss [38]. This deterioration manifests as cognitive impairment and disease progression, ultimately leading to death. Additionally, mutations in the tau gene are linked to frontotemporal dementia and parkinsonism, indicating that tau dysfunction may play a role in neurodegeneration [39].
3.3. Factors Related to SAD
Research increasingly indicates that in SAD, various risk factors stemming from the interactions between genes and environmental influences contribute to the activation and progression of the disease (Figure 2), particularly in the context of an aging brain [40]. Many researchers in the field of AD suggest that these environmental risk factors play a significant role during the preclinical phase of the illness, which can occur decades before the onset of clinical dementia. Key modifiable risk factors for SAD primarily include cardiovascular issues such as diabetes and obesity, as well as lifestyle habits like smoking, physical activity, diet, and alcohol consumption.

Figure 2.
Risk factors involved in the development of SAD.
3.3.1. Aging
Aging is the primary and undisputed risk factor for AD [41]. Several critical processes related to aging—such as oxidative stress, mitochondrial dysfunction, impaired blood–brain barrier (BBB) function, and arteriolosclerosis, which can lead to hypoxia—have all been linked to an increased risk of developing AD. Notably, many of these processes are associated with enhanced amyloidogenic processing of APP and the aggregation of Aβ [42].
Research involving AD transgenic mice has shown that cerebral hypoxia increases the activity of BACE1. This enzyme’s activity is regulated by the hypoxia-inducible factor 1-alpha (HIF-1α). Consequently, the increase in BACE1 activity leads to elevated production of Aβ.
3.3.2. Metal Exposure
There is substantial evidence linking metal exposure to the development of AD. An increased metal concentration in the brain is associated with Aβ deposition in senile plaques, leading to a state of oxidative stress that results in neuronal damage [43]. Aβ catalyzes the reduction of Cu2+ and Fe3+, and in the absence of sufficient antioxidant mechanisms, this process can produce toxic ROS that may contribute to the pathogenesis of AD. The generation of ROS accelerates the aggregation of Aβ by creating species that are less effectively eliminated through the BBB and are more prone to aggregation [44].
AD has been associated with exposure to certain metals, including aluminum (Al3+). This metal can enter the body through occupational exposure or by consuming food or water contaminated with traces of aluminum, mainly when using aluminum cookware [45]. Once in the body, aluminum can disrupt the BBB and accumulate in the brain, altering mitochondrial function, depleting adenosine triphosphate (ATP), and producing ROS. This process can lead to cell death through apoptotic or necrotic mechanisms [46]. Additionally, aluminum can inhibit the activity of antioxidant enzymes, disrupting brain neurochemistry and causing oxidative damage to brain DNA.
3.3.3. Brain Injury
Vascular brain injury that leads to reduced blood flow, known as hypoperfusion, can initiate and promote the development of amyloid pathology in the brain. Multiple pathological processes connect traumatic brain injury (TBI) with neurodegeneration and dementia [47]. The term chronic traumatic encephalopathy (CTE) refers to the neurological changes that can occur later in life in individuals who have experienced repeated concussive head injuries. This condition can present symptoms and pathological features similar to those of other neurological diseases, including AD, Parkinson’s disease (PD), frontotemporal dementia, and amyotrophic lateral sclerosis [48]. Postmortem examinations of brains affected by TBI and CTE have revealed pathological features typically associated with AD [49], such as increases in hyperphosphorylated tau (P-tau) and, in certain cases, Aβ deposits. Recent consensus criteria classify CTE as a tauopathy [50].
3.3.4. Proinflammatory Cytokines
In AD patients, the brain exhibits a significant inflammatory response primarily driven by activated microglia and astrocytes [51]. Both species, oAβ and fAβ, stimulate the activation of microglia in the AD brain, leading to a proinflammatory response mediated by cytokines such as interleukin-1 (IL-1), interleukin-6 (IL-6), and tumor necrosis factor-alpha (TNF-α), in addition to the complement system and chemokines. This inflammatory response plays a critical role in the neurodegeneration associated with AD [52,53,54].
Furthermore, elevated levels of circulating proinflammatory cytokines have consistently been reported in individuals with AD [55]. Research has shown that higher serum levels of IL-6, TNF-α, and IL-1 are linked to an increased risk of developing AD. Studies have also identified communication pathways between the peripheral immune and central nervous system, indicating that elevated levels of peripheral proinflammatory cytokines can lead to increased cytokine production in the central nervous system [56].
In AD pathogenesis, central proinflammatory cytokines have been shown to promote the phosphorylation of tau protein and the production of amyloid beta, thereby exacerbating the neurodegenerative process [57].
3.3.5. Type 2 Diabetes Mellitus
Large-scale, population-based prospective studies have demonstrated that type 2 diabetes increases the risk of developing AD [58]. While there is ongoing debate between epidemiological findings and neuropathological observations regarding the relationship between type 2 diabetes and AD, experimental research has linked insulin and insulin resistance to the pathogenesis of AD in several ways. In the brain, insulin and insulin-like growth factor signaling regulate a wide range of functions, including glucose and energy metabolism, as well as the growth, differentiation, migration, survival, and plasticity of neurons [59].
Specifically, in terms of Aβ metabolism, insulin aids in the trafficking of APP and Aβ from the trans-Golgi network to the plasma membrane, which facilitates the extracellular release of Aβ. Additionally, insulin resistance in mice—induced by a high-fat diet—results in increased activity of BACE1 and γ-secretase, contributing to the accumulation of Aβ [60].
3.3.6. Obesity
The human brain contains a significant amount of cholesterol, primarily located in myelin and the plasma membranes of glial cells and neurons [61]. Within cell membranes, cholesterol is associated with specialized lipid microdomains known as lipid rafts. These lipid rafts play a crucial role in the amyloidogenic processing of APP and the oligomerization of Aβ peptide, suggesting that cholesterol may influence these processes [62].
Research involving animal models has indicated that a cholesterol-rich diet tends to increase Aβ accumulation in the brain. This phenomenon can be explained by previous studies demonstrating that cholesterol affects the processing of APP. In cultures of rat hippocampal neurons, lowering cellular cholesterol levels—either by inhibiting de novo synthesis with statins or using the cholesterol-extracting agent methyl-β-cyclodextrin—resulted in a significant decrease in Aβ production and secretion. Furthermore, cholesterol depletion led to a marked reduction in Aβ levels. In contrast, the secretion of the APP ectodomain (produced through the non-amyloidogenic α-secretase pathway) was observed to increase [63].
3.3.7. Smoking
Observational studies indicate a link between current smoking and an increased risk of dementia, SAD, and cognitive decline. It is estimated that nearly 14% of AD cases may be potentially attributable to smoking due to its high prevalence [64]. Smoking may raise the risk of AD through several mechanisms, primarily related to oxidative stress and inflammatory responses [65]. Furthermore, the risk increases significantly with greater cumulative cigarette exposure.
Research has shown that mice exposed to daily cigarette smoke for 15 weeks experience oxidative stress in brain tissue, as evidenced by elevated levels of malondialdehyde, which is a marker of lipoperoxidation [66]. Another study found that rats exposed to daily cigarette smoke for approximately 8 weeks exhibited significantly increased levels of 8-hydroxydeoxyguanosine, a marker of oxidative damage to RNA and DNA nucleosides, particularly in the dentate gyrus and CA3 subfields of the hippocampus [67].
Additionally, Khanna et al. reported that rats exposed to cigarette smoke five days a week for six weeks showed significantly elevated levels of proinflammatory cytokines, including IFN-γ and TNF-α, in the brain. Multiple cytokine genes, such as TNF-α, IL-1α, IL-1β, and Th17, were also upregulated. These changes contribute to the development of neuroinflammation and brain damage [68].
3.3.8. Diet
Diet has been shown to influence the immune system, and various nutrients and bioactive components can affect neuroinflammatory processes in animals [69]. For example, polyphenols, unsaturated fats, and antioxidant vitamins help reduce oxidative stress and neuroinflammation, whereas saturated fats promote inflammation, particularly in the hypothalamus [70].
However, it remains unclear whether the effects of diet on neurocognition are directly mediated by neuroinflammatory processes or by other immune mechanisms in vivo. An increasing amount of evidence suggests that peripheral inflammation and changes in the gut microbiome can enhance neuroinflammation and accelerate neurodegeneration, and these external factors can also be influenced by diet [71].
3.3.9. Alcohol Consumption
Research indicates a connection between the development of cognitive impairment, dementia, and harmful alcohol use [72]. However, the relationship between alcohol consumption and cognitive outcomes is still debated. Observational studies suggest that heavy alcohol consumption is linked to alcoholic dementia and can result in a decline in mental and executive functioning. On the other hand, moderate alcohol intake may offer a protective effect. This relationship between alcohol consumption and cognitive decline is often described as U-shaped [73].
3.3.10. Genetics
FAD is typically characterized by a rapid rate of progression and follows a Mendelian pattern of inheritance. Three main genes—APP, PSEN1, and PSEN2—encode proteins that are involved in the breakdown of APP and the production of Aβ, and they have been strongly linked to the pathophysiology of FAD. In contrast, the genes associated with SAD increase the risk of the disease in a non-Mendelian manner. First-degree relatives of individuals with SAD have a lifetime risk of developing the disease that is twice as high as that of individuals without an affected first-degree relative. Furthermore, SAD is more common in monozygotic (identical) twins than in dizygotic (fraternal) twins, indicating a significant genetic contribution, estimated to be between 60% and 80% [74].
For over a decade, the only well-established genetic risk factor for SAD has been the APOEe4 allele, which is located on chromosome 19q13. This allele is a part of the APOE gene, which encodes a lipid-binding protein and has three common isoforms: APOEe2, e3, and e4. Having a single APOE-e4 allele is associated with a 2- to 3-fold increased risk of developing AD, while having two copies can increase the risk by 5-fold or more [74]. Additionally, APOEe4 is linked to lower cognitive performance, particularly in the memory domain, and is associated with mild cognitive impairment (MCI) and the progression from MCI to dementia [75].
In addition, genome-wide association studies have identified polymorphisms in or near several genes that are associated with AD risk, including ABCA7, CLU, SORL1, BIN1, PICALM, and TREM2 [76].
3.4. Non-Transgenic Models
Considering the many factors related to SAD, non-transgenic mouse models have been developed and used to evaluate new treatments, as shown in Figure 3. However, these models are less well-known and employed than transgenic models. As Figure 3 shows, these models are developed by mimicking the risk factors for SAD.
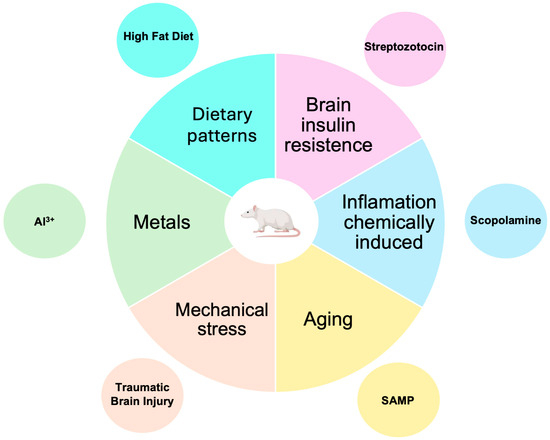
Figure 3.
Non-transgenic murine models for AD research. Although these models employ mechanisms related to risk factors for developing SAD (which accounts for 95% of AD cases), they are less commonly used than transgenic models.
3.4.1. Models Induced by the Administration of Streptozotocin (STZ)
Streptozotocin (STZ) is a bacterial toxin that selectively damages pancreatic β cells. It is widely used to induce diabetes in animal models. Administration of STZ in rodents via the intracerebroventricular (iv) route reduces cognition and promotes the formation of Aβ aggregates and phosphorylation of tau protein (Table 1) [77]. Due to its chemical nature, it can generate intracellular free radicals, nitric oxide (NO), hydrogen peroxide (H2O2), and mitochondrial damage. In addition, this compound causes neurodegeneration, oxidative and nitrative stress, glucose hypometabolism, a cerebral state of insulin resistance, progressive cholinergic deficiency [78], and neuroinflammation [79]. Similar biochemical, physiological, and anatomical effects are observed in patients with AD, making STZ a valuable means of obtaining animal models of AD. Models can be generated employing rats or mice. The rat model is obtained by administering 3 mg/kg of STZ under anesthesia; the injection is performed with the aid of a stereotaxic apparatus using the coordinates 0.9 mm posterior to the bregma point, ±1.8 mm lateral, and 3.8 mm vertical. The STZ is dissolved in 2 µL of artificial cerebrospinal fluid or sodium citrate buffer solution and is applied with a microsyringe. Administration can be performed on one or two occasions. The evaluation of the model is carried out three weeks after the injection [78,80].
In a mouse model, 3 mg/kg of STZ is administered unilaterally or bilaterally under anesthesia using a stereotaxic device with the coordinates −0.3 mm posterior, −1 mm lateral, and −2.5 mm vertical. As in the previous case, the STZ is dissolved and applied with a microsyringe. The model evaluation tests are carried out three weeks after the injection [81,82,83].
STZ-induced AD models have been evaluated and characterized for over 20 years, but their study has not yet been completed. Among the advantages that these models have over the transgenic animal model are that genetic manipulation does not begin at birth; pathological alterations can start at any age of the animal; the onset, development, and progression of cognitive impairments, as well as biochemical and structural changes, can be followed at different time points, and these models are suitable for studying potential drugs for prevention and treatment in AD [78].

Table 1.
Characteristics of non-transgenic animal models of AD.
Table 1.
Characteristics of non-transgenic animal models of AD.
| Model | Animal | Aβ | P-Tau | Neuro-Inflammation | Other Changes | Time | Selected References |
|---|---|---|---|---|---|---|---|
| Stretptozotocyn | Mouse Rat | ✔ | ✔ | ✔ | Oxidative and nitrative stress, glucose hypometabolism cerebral state of insulin resistance, progressive cholinergic deficiency, increased exploratory activity, impaired learning and spatial memory. | 3 weeks | [81] |
| Scopolamine | Mouse Rat | ✔ | ✔ | ✔ | Neuronal apoptosis, changes in the expression of choline acetyltransferase (ChAT), acetylcholinesterase (AChE), and acetylcholine (ACh) receptors. Changes in memory evaluated by MWM, passive avoidance. | 1 to 6 weeks | [84] |
| Aging | Mouse | ✔ | ✔ | ✔ | Oxidative stress, cognitive deficit, especially in spatial memory and learning tasks. | 7–8 months | [85] |
| Mechanical stress | Mouse Rat | ✔ | ✔ | ✔ | Cognitive dysfunction, including impairments in memory and object recognition, along with behavioral disturbances such as anxiety and symptoms resembling depression. | 1 month | [86] |
| Aluminum | Rat | ✔ | ✔ | Lipoperoxidation increased malondialdehyde levels, lower superoxide dismutase activity, and catalase activity. | 4–6 weeks | [87] | |
| Dietary patterns High-fat diet | Rat Mice | ✔ | ✔ | ✔ | Dysregulations in autophagy, apoptosis. Oxidative stress and decreases in long-term potentiation. | 8–30 weeks | [88] |
Both rat and mouse models have been used to study AD pathology and evaluate new pharmacological treatments. Assessed drugs include inhibitors of the enzyme choline acetylcholinesterase, NMDA receptor antagonists, antioxidants, anti-inflammatory drugs, antidiabetic compounds, glycogen synthase kinase 3β (GSK3β) inhibitors, and antihypertensives [78].
3.4.2. Models Induced by the Administration of Scopolamine
The pathogenic mechanisms induced by scopolamine administration to mice or rats correlate with altered cholinergic function, increased oxidative stress, the induction of the amyloid cascade, and the expression of inflammatory mediators [89]. The mentioned changes contribute to the loss of cholinergic neurons and deficits in attention and memory (Table 1) [90]. In addition, scopolamine increased phosphorylated tau protein levels by >2-fold [91], thus highlighting the importance of post-translational phosphorylation modification in exacerbating tau pathology—increased hyperphosphorylated tau results as a consequence of the upregulation of GSK3β [91]. GSK3β, a tau kinase, may also be involved in AD pathogenesis through the upregulation of Aβ production and suppression of ACh synthesis [92].
Models can be generated employing rats or mice. Male 8–10-week-old mice weighing 25–30 g are used. After acclimation, animals are administered scopolamine at 0.75 mg/kg intraperitoneally (ip) once daily for seven consecutive days to induce memory impairment. The assessment of pathological changes occurs after treatment ends [93]. The Morris water maze test (MWM) evaluates the memory impairment produced by scopolamine administration in mice. Histological studies have demonstrated that in the CA1 region of the hippocampus, using the Nissl test, inflamed and vacuolated neurons show nuclear and apoptotic pyknosis, along with decreased neurons in the scopolamine model [94].
Additionally, changes in the expression of choline acetyltransferase (ChAT), acetylcholinesterase (AChE), and ACh receptors in the brains of rats that receive scopolamine have been reported [70]. A second model in mice involves administering scopolamine (4 mg/kg) via ip daily for 6 days. In this study, memory changes were assessed through the MWM test, and the levels of ACh and AChE were confirmed [95]. In addition, Wistar albino rats weighing 250 to 300 g can induce the scopolamine model. Scopolamine 1 mg/kg/ip is administered for 14 to 45 days. Once the treatment is finished, the evaluation tests are carried out [96]. The model is assessed by the passive avoidance memory test consisting of a dark/light two-compartment shuttle box separated by a door, with a conductor available on the floor of the dark compartment for electric shocks [97]. The object recognition test investigates short-term recognition memory functions [98]. Biochemical tests for reduced glutathione (GSH), malondialdehyde (MDA), and myeloperoxidase (MPO) are used to determine levels of oxidative stress [99].
3.4.3. Models Induced by Aging
Aging is the most significant risk factor for SAD. The mechanisms underlying aging-related susceptibility to SAD are not well known. There are transgenic and non-transgenic rodent aging models in which animals are observed throughout their lives to study aging-related changes and underlying disease mechanisms and to test possible treatments [100]. One of the non-transgenic rodent models includes the following:
SAMP8 mice were developed from senescence-accelerated mice (SAM), which are senescence-accelerated strains that developed spontaneously from breeding pairs of the AKR/J series [85,101]. SAM are classified into two groups: senescence-accelerated prone mice (SAMP1, 2, 3, 6, 7, 8, 9, 10, and 11) and senescence-accelerated resistant mice (SAMR1, 4, and 5) [102]. SAMR strains show typical aging features, whereas SAMP strains show senescence acceleration and age-related pathological phenotypes similar to aging disorders observed in humans. SAMP mice display various clinical features of aging disorders as early as 7 or 8 months of age, which is approximately equivalent to middle age for mice compared to normal-aged mice (>20 months) [103]. Although they are not exact models of human aging, they exhibit aging-related physiological and, particularly, behavioral changes more rapidly than other mouse strains. They have been widely used in several studies of age-related neurodegeneration, including SAD [102,104].
The most important features of this model are as follows:
- Premature aging with an early onset of age-related markers [102,103].
- Memory and learning problems. One of the most studied traits of SAMP8 mice is their cognitive deficit, especially in spatial memory and learning tasks as they age [103,104,105].
- Histopathological brain features similar to SAD in humans. As SAMP8 mice age, they accumulate Aβ plaques and neurofibrillary tangles in the brain, two hallmark histopathological AD findings in humans [104,105].
- Oxidative stress. SAMP8 mice have been shown to have increased levels of oxidative stress in the brain, which contributes to cellular damage and may be related to their accelerated aging and the onset of neurodegenerative diseases [85,106].
These characteristics of SAMP8 mice make this model one of the leading models for studying the mechanisms of AD and testing potential treatments and therapies that may delay or mitigate the effects of aging and dementia (Table 1).
3.4.4. Models Induced by Mechanical Stress
Many factors can alter the brain cells. Among the stressors that cause damage to the brain are electromagnetic radiation, temperature, and atmospheric pressure. These factors cause an essential change in the structural conformation of neuroglial proteins. Brain cells are sensitive to physical and mechanical stress. Indeed, various studies associate the development of neurodegenerative pathologies with exposure to injuries caused by traumas related to physical or work activity, as is the case of impact sports athletes, such as boxers and football players, where these patients present pathologies similar to those existing in AD [107]. The use of animal models enables one to illustrate that traumatic brain injuries can trigger amyloidogenic processing of APP and accumulation of Aβ in rodents. Tau pathology can be observed in rodent models subjected to mild repetitive trauma (Table 1). Although they may be nonspecific, neuroinflammation and neuronal loss can also be induced in these models through methods such as controlled cortical impact, weight drop impact, and repetitive mild trauma. Additionally, cognitive dysfunction—including memory impairment, object recognition issues, behavioral disturbances (such as anxiety and depression-like symptoms), and sensorimotor deficits—has been reported in models of TBI [108].
In patients with traumatic brain injuries, there is often an increase in the amount of Aβ at the injury site shortly after the trauma occurs. Similarly, studies on TBI conducted in mice have also revealed the presence of Aβ at the site of the injury [109]. Research has shown that specific proteins can move between interconnected cells via axons in different brain areas. This movement may play a crucial role in the development of amyloid pathology in AD brains [110]. Adult male BALB/c mice of approximately 6 weeks are commonly used to induce the model, weighing 19 and 24 g. Each mouse is exposed to repetitive sub-concussive impacts with stainless steel masses (marbles) of 13.5 or 20 g dropped at a height of 20 or 40 cm through a hollow tube to obtain impact forces on the dorsal face of the skull between 0.03 and 0.04 at 20 cm and between 0.04 and 0.08 J at 40 cm; the impact frequencies were one, two, or up to four per day at specific times for each group of study mice. The mice did not present neurological problems after the impacts; only those in which a force of 0.08 J was used presented some lethargy and some social isolation; the measurement of Aβ and tau protein was performed by confirmation with immunoassays; significant values were found in the studies carried out at higher altitudes, and at lower altitude, these values were lower. It is concluded that the best results were obtained with greater exposure to the processes and that this procedure is the most appropriate compared to other studies [111].
3.4.5. Models Induced by Metals
In living organisms, metal ions are essential in trace amounts but can be toxic at high levels. Increased metal concentration in the brain has been linked to generating ROS, which can affect DNA and proteins and constitute part of the etiology of neurodegenerative diseases in aging. The accumulation of free metal ions such as iron (Fe2+), copper (Cu2+), zinc (Zn2+) [112,113], aluminum (Al3+) [114], and manganese (Mn2+), among others, has been correlated with AD, PD, amyotrophic lateral sclerosis, Wilson’s disease, hereditary ferritinopathy, and stroke [115].
Al3+ is not an essential element for living organisms, but it plays a role in several important processes. These include axonal transport, neurotransmitter synthesis, synaptic transmission, protein phosphorylation and dephosphorylation, protein degradation, gene expression, and the inflammatory response. However, high concentrations of aluminum in the brain can lead to spatial memory deficiencies, affect emotional reactions, and impair various brain functions related to memory and learning. [116]. The toxicity mechanism of Al3+ is not fully known. However, it has been described that due to its chemical nature, it has an affinity for negatively charged molecules such as phosphate, carboxyl, and hydroxyl groups of biomolecules, with which it can establish covalent bonds and affect their topology; when binding to DNA, it modifies gene expression [117]. When binding to ATP, it influences energy metabolism; this ion also inhibits the function of protein phosphatases and protein kinases, as well as ion-activated and exchanger proteins [116], inhibits dephosphorylation of the tau protein, and increases its aggregation [118]; in addition, it promotes the oxidation of Fe, increases the formation of ROS, and causes an inflammatory response in different tissues [114]. In the past decade, it was reported that rats treated with chronic intake of Al3+ showed a significant increase in the expression of the APP gene in the cerebral cortex and hippocampus, suggesting that one of the mechanisms contributing to the neuropathological picture of Al3+ in AD is through the activation of the cascade of formation of the Aβ peptide [119]. In recent decades, AlCl3 has been used to generate rat models that reproduce (albeit partially) the symptoms and pathophysiological alterations of AD.
According to bibliographic findings, there are two processes for generating rat models with AD through the administration of AlCl3. The first consists of randomly selecting Wistar rats weighing 230 ± 20 g and administering 4.2–50 mg/kg of AlCl3 dissolved in water ip for 28–42 days (depending on the experimental design); 24 h later, behavioral assessment tests, including the MWM, are performed, and 24 h later, biochemical tests of MDA levels, superoxide dismutase activity, and catalase activity) [120,121] are carried out to screen individuals that mimic the symptomatology and neuropathology of AD. The second consists of randomly selecting Wistar rats weighing 100–200 g and administering a combination of AlCl3 and d-galactose (d-gal), in the amounts of 150 mg/kg and 300 mg/kg, respectively, dissolved in saline solution (0.9%), for one week. Twenty-four hours later, the behavior is evaluated employing MWM, and 24 h later, they are sacrificed. To assess the model, the corresponding biochemical and neurochemical tests are performed: lipoperoxidation, MDA levels, superoxide dismutase activity, and catalase activity) [122,123]. Both processes give rise to animals that reproduce the symptoms of AD and are suitable for studies of the mechanisms involved in it. The main advantage of the two methods is that the latter requires less experimental time and a lower cost [124]. Rat models of induced AD have been used in the evaluation of novel neuroprotective compounds for the prevention and future treatments of AD in humans [125].
3.4.6. Dietary Patterns
Dietary patterns have been examined as modifiable risk factors for cognitive decline, dementia, and related conditions [126]. Clinical studies have explored various dietary patterns, including the Mediterranean diet, ketogenic diet, and DASH diet, and their effects on AD pathology. These studies suggest that a diet low in sugar and high in omega-3 polyunsaturated fatty acids and antioxidant compounds may be beneficial in preventing cognitive decline [127]. Conversely, adhering to an unhealthy diet can increase the risk of AD. In a cross-sectional study, a preference for junk food significantly predicted cerebral Aβ deposition [128].
Additionally, the consumption of a high-fat diet (HFD) is a major contributor to overweightness and obesity. This can lead to elevated levels of circulating free fatty acids, resulting in low-grade inflammation due to the increased release of proinflammatory cytokines. These inflammatory processes activate signaling pathways, including serine kinases, IKKβ, and JNK1, which inhibit insulin receptor signaling and enhance insulin resistance. This situation contributes to cognitive impairment and neurodegeneration [129]. Furthermore, high fat and high fructose consumption are associated with diabetes and hyperinsulinemia, both of which are risk factors for developing SAD. A link between obesity and AD has been suggested, where insulin resistance plays a role in regulating inflammatory processes, adipokine dysregulation, oxidative stress, and mitochondrial dysfunction [130].
Non-transgenic animal models have proven useful for studying various dietary patterns’ molecular and cellular basis and their relationship with SAD [131]. For instance, Busquets et al. reported the formation of Aβ in C57BL/6J mice fed an HFD over a long period. These mice also exhibited dysregulations in autophagy and apoptosis alongside exacerbated inflammatory processes [132]. The HFD induces brain insulin resistance by reducing the tyrosine phosphorylation of the insulin receptor and increasing the serine phosphorylation of insulin receptor substrate 1, which can disrupt the proteins involved in the insulin signaling pathway [131]. Additionally, an HFD exacerbates neuroinflammation and oxidative stress while diminishing long-term potentiation in the hippocampus [132].
However, this model has limitations related to the varying compositions of diets reported and the different time points used to evaluate molecular pathways. For example, studies involving an 8-week HFD consumption period have indicated BBB damage associated with microglial activation, increased oxidative stress, elevated proinflammatory cytokines, and memory impairment [133]. In contrast, consuming an HFD for 30 weeks increases protein expression of phosphorylated tau, APP, and BACE1 cleaving enzymes, leading to the formation of Aβ plaques and neurofibrillary tangles [134]. Conversely, short-term consumption periods of an HFD (3, 7, or 10 days) affect several markers of inflammation, endoplasmic reticulum stress, and apoptosis in the hippocampus of young mice [135]. Therefore, the molecular and cellular changes over time in this model remain incompletely understood and may offer temporally relevant biological insights.
4. Discussion
Animal models of AD have played a crucial role in defining critical disease-related mechanisms and are essential for evaluating novel therapeutic approaches. Many treatments currently being tested in clinical trials originate from studies conducted on rodent models. The current transgenic models of AD are derived from familial FAD and are histopathologically indistinguishable from SAD [136].
However, SAD’s mechanisms are more complex and varied than the monogenic dominant mutations responsible for FAD, making them less understood and less extensively modeled [137]. Transgenic models that overproduce mutant APP lead to amyloid accumulation. Due to the complexity of the disease, none of the transgenic lines have been able to replicate all aspects of AD pathology. This highlights the limitations of using transgenic systems to reproduce the human disease process fully.
Doubly mutated transgenic models with increased Aβ accumulation develop lesions earlier [138]. In contrast, neurofibrillary tangles are primarily found in transgenic mice that overexpress mutated tau [139]. While mixed transgenic models that incorporate both Aβ and tau accumulation mimic the histological changes observed in AD, the genetic modifications required to express these pathological features differ from those found in familial FAD. Additionally, many transgenic AD models fail to replicate the human condition’s neuronal and synaptic loss, characteristic of the disease.
Furthermore, the inflammation present in these models does not accurately reflect the inflammation seen in humans with AD, as there are notable differences in the nature and severity of the inflammation between humans and AD transgenic mice [140]. The complexity of the disease suggests that no single transgenic line can replicate all aspects of AD pathology, highlighting the limitations of using transgenic systems to model human disease processes.
Another concern with transgenic AD models is that the overproduction of Aβ drives the pathological process in line with the amyloid cascade hypothesis. These models effectively reproduce FAD, while the majority of individuals experience SAD. In SAD, the disease is linked to an imbalance between the production and clearance of Aβ, leading to its accumulation over time. However, the mechanisms behind this imbalance remain poorly understood [140].
From a practical perspective, there is often a discrepancy between preclinical animal models and human clinical trial results. While transgenic models enhance our understanding of AD pathogenesis, their limitations highlight the need to explore new approaches in AD animal modeling.
In contrast, non-transgenic experimental models of AD serve as a valuable and complementary preclinical strategy alongside transgenic animal models. These non-transgenic approaches demonstrate the ability to resemble the key mechanisms underlying neurodegeneration in AD [141], including Aβ deposition, tau hyperphosphorylation and neuroinflammation (Table 1). Furthermore, it is imperative to study the combination of various non-transgenic models to evaluate changes in Aβ deposition, tau protein phosphorylation, neuroinflammation, and subsequent cognitive impairment. This is because patients with AD generally present several pathophysiological alterations that converge toward the development of the disease [3]. This would facilitate understanding of the changes that promote its development, in addition to providing a model that allows for the evaluation of known and novel therapeutics, particularly those that have shown favorable outcomes in this type of model. This standardization should involve using a single animal model and assessing the same therapeutic strategy across different models. Utilizing non-transgenic animal models facilitates the investigation of risk factors and the identification of molecular mechanisms contributing to SAD. This research domain is currently underexplored but holds the potential for advancing the early identification of individuals at risk and the development of innovative therapeutic strategies. While the present landscape of preclinical research examining therapeutic interventions in non-transgenic animal models of SAD remains limited and incomplete, we advocate that these models may enhance drug development. They offer a cost-effective, accessible, and efficient research platform, allowing for a shift in the focus of drug development from traditional amyloid-targeted strategies to novel pathological approaches within the context of AD.
5. Conclusions
AD is a devastating condition that significantly impacts society. Despite decades of research and billions of dollars invested, effective treatments have not yet reached the market. Most drug candidates that have failed in clinical trials showed anti-AD activity in various transgenic animal models. While transgenic mouse models have been the primary method used in drug development research to evaluate potential therapies, it is crucial to recognize the need for animal models that better reflect the key pathological changes associated with similar risk factors.
These models could play an increasingly important role in current drug development efforts for AD. Therefore, there is an urgent need for a model that accurately represents the changes seen in sporadic SAD, the most common form of AD, which could facilitate successful drug development in the future.
Author Contributions
Conceptualization, writing—original draft preparation M.H.-R., E.M.J., M.I.N.-V., J.M.V.L. and M.M.-R.; literature search and writing—review and editing M.H.-R., E.M.J. and M.I.N.-V.; supervision M.H.-R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCYT), grant number 320862; Secretaría de Educación, Ciencia, Tecnología e Innovación de la Ciudad de México, grant number SECTEI/129/2023; and Instituto Politécnico Nacional, grant number 20241531.
Acknowledgments
The authors acknowledge the following for their financial support for this research: Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCYT) through grant number 320862; Secretaría de Educación, Ciencia, Tecnología e Innovación de la Ciudad de México through grant number SECTEI/129/2023; and Instituto Politécnico Nacional through grant number 20241531.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| α7 nAChR | α7-nicotinic acetylcholine receptor |
| ACh | acetylcholine |
| AChE | acetylcholinesterase |
| AD | Alzheimer’s disease |
| ChAT | cholineacetyltransferase |
| ATP | adenosine triphosphate |
| Al3+ | aluminum |
| AD | Alzheimer’s disease |
| APP | amyloid β precursor protein |
| Aβ | amyloid-β |
| fAβ | amyloid-β fibril |
| oAβ | amyloid-β oligomer |
| ALS | amyotrophic lateral sclerosis |
| BACE1 | β secretase |
| PrPc | cellular prion protein |
| CNS | central nervous system |
| CTE | chronic traumatic encephalopathy |
| Cu2+ | copper |
| d-gal | d-galactose |
| FAD | familiar form of AD |
| RAGE | glycation product receptor |
| GSK3β | glycogen synthase kinase 3β |
| H2O2 | hydrogen peroxide |
| HFD | high-fat diet |
| ·OH | hydroxyl radical |
| P-Tau | hyperphosphorylated tau |
| HIF-1α | hypoxia-inducible factor 1-alpha |
| IGF | insulin-like growth factor |
| icv | intracerebroventricular |
| ip | intraperitoneal |
| Fe2+ | iron |
| MDA | malondialdehyde |
| MCI | mild cognitive impairment |
| Mn2+ | manganese |
| MWM | Morris water maze |
| MPO | myeloperoxidase |
| NMDA | N-methyl-d-aspartate |
| NFTs | neurofibrillary tangles |
| NO | nitric oxide |
| PD | Parkinson’s disease |
| PSEN1 | Presenilin 1 |
| PSEN2 | Presenilin 2 |
| P-tau | hyperphosphorylated tau |
| ROS | reactive oxygen species |
| GSH | reduced glutathione |
| SAM | senescence-accelerated mouse |
| SAD | sporadic form of AD |
| STZ | streptozotocin |
| TBI | traumatic brain injury |
| WD | Wilson’s disease |
| Zn2+ | zinc |
References
- Gao, S.; Burney, H.N.; Callahan, C.M.; Purnell, C.E.; Hendrie, H.C. Incidence of Dementia and Alzheimer Disease Over Time: A Meta-Analysis. J. Am. Geriatr. Soc. 2019, 67, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Dorszewska, J.; Prendecki, M.; Oczkowska, A.; Dezor, M.; Kozubski, W. Molecular Basis of Familial and Sporadic Alzheimer’s Disease. Curr. Alzheimer Res. 2016, 13, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.A. What causes Alzheimer’s disease? Folia. Neuropathol. 2013, 51, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Sims, R.; van der Lee, S.J.; Naj, A.C.; Bellenguez, C.; Jun, G.; Logsdon, B.A.; Williams, J. Genetic mechanisms of Alzheimer’s disease. Nat. Neurosci. 2020, 23, 304–316. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Zhang, Y. Animal models of Alzheimer’s disease: Applications, evaluation, and perspectives. Zool. Res. 2022, 43, 1026–1040. [Google Scholar] [CrossRef]
- Van Dam, D.; De Deyn, P.P. Non human primate models for Alzheimer’s disease-related research and drug discovery. Expert. Opin. Drug Discov. 2016, 12, 187–200. [Google Scholar] [CrossRef]
- Drummond, E.; Wisniewski, T. Transgenic mouse models of Alzheimer’s disease: Progress and challenges. J. Alzheimer’s Dis. 2017, 57, 341–363. [Google Scholar]
- Zhong, M.Z.; Peng, T.; Duarte, M.L.; Wang, M.; Cai, D. Updates on mouse models of Alzheimer’s disease. Mol. Neurodegener. 2024, 19, 23. [Google Scholar] [CrossRef]
- Wenk, G.L. Animal models of Alzheimer’s disease: Are they valid and useful? Acta Neurobiol. Exp. 1990, 50, 219–223. [Google Scholar]
- Ameen-Ali, K.E.; Wharton, S.B.; Simpson, J.E.; Heath, P.R.; Sharp, P.; Berwick, J. Review: Neuropathology and behavioural features of transgenic murine models of Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 2017, 43, 553–570. [Google Scholar] [CrossRef]
- Sarasa, M.; Pesini, P. Natural non-trasgenic animal models for research in Alzheimer’s disease. Curr. Alzheimer Res. 2009, 6, 171–178. [Google Scholar]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Perl, D.P. Neuropathology of Alzheimer’s disease. Mt. Sinai J. Med. 2010, 77, 32–42. [Google Scholar] [PubMed]
- Alzheimer, A. Alzheimer’s disease (AD) is an irreversible, progressive neurodegenerative disorder that occurs gradually and results in memory loss, unusual behavior, personality changes, and a decline in thinking abilities. AD is named after Uber eine eijenartige Erkrankung der Hirnride. Allg. Z. Psychiatr. 1907, 64, 146–148. [Google Scholar]
- Jack, C.R.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Re-search Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer’s Dement. 2018, 14, 535–562. [Google Scholar]
- Gilbert, B.J. The role of amyloid β in the pathogenesis of Alzheimer’s disease. J. Clin. Pathol. 2013, 66, 362–366. [Google Scholar] [CrossRef]
- Dickson, D.W. The pathogenesis of senile plaques. J. Neuropathol. Exp. Neurol. 1997, 56, 321–339. [Google Scholar] [CrossRef]
- Tiwari, S.; Atluri, V.; Kaushik, A.; Yndart, A.; Nair, M. Alzheimer’s disease: Pathogenesis, diagnostics, and therapeutics. Int. J. Nanomed. 2019, 19, 5541–5554. [Google Scholar]
- Tiana, G.; Simona, F.; Broglia, R.A.; Colombo, G. Thermodynamics of beta-amyloid fibril formation. J. Chem. Phys. 2004, 120, 8307–8317. [Google Scholar] [CrossRef]
- Walsh, D.M.; Klyubin, I.; Fadeeva, J.V.; Rowan, M.J.; Selkoe, D.J. Amyloid-beta oligomers: Their production, toxicity and therapeutic inhibition. Biochem. Soc. Trans. 2002, 30, 552–557. [Google Scholar] [CrossRef]
- Morris, J.C.; Storandt, M.; McKeel, D.W., Jr.; Rubin, E.H.; Price, J.L.; Grant, E.A.; Berg, L. Cerebral amyloid deposition and diffuse plaques in “normal” aging: Evidence for presymptomatic and very mild Alzheimer’s disease. Neurology 1996, 46, 707–719. [Google Scholar] [PubMed]
- Hayden, E.Y.; Teplow, D.B. Amyloid β-protein oligomers and Alzheimer’s disease. Alzheimers Res. Ther. 2013, 5, 60. [Google Scholar]
- Gyure, K.A.; Durham, R.; Stewart, W.F.; Smialek, J.E.; Troncoso, J.C. Intraneuronal abeta-amyloid precedes development of amyloid plaques in Down syndrome. Arch. Pathol. Lab. Med. 2001, 125, 489–492. [Google Scholar] [PubMed]
- Walsh, D.M.; Klyubin, I.; Shankar, G.M.; Townsend, M.; Fadeeva, J.V.; Betts, V.; Podlisny, M.B.; Cleary, J.P.; Ashe, K.H.; Rowan, M.J.; et al. The role of cell-derived oligomers of Abeta in Alzheimer’s disease and avenues for therapeutic intervention. Biochem. Soc. Trans. 2005, 33, 1087–1090. [Google Scholar]
- Yan, S.S.; Chen, D.; Yan, S.; Guo, L.; Du, H.; Chen, J.X. RAGE is a key cellular target for Abeta-induced perturbation in Alzheimer’s disease. Front. Biosci. 2012, 4, 240–250. [Google Scholar]
- Koerich, S.; Parreira, G.M.; de Almeida, D.L.; Vieira, R.P.; de Oliveira, A.C.P. Receptors for Advanced Glycation End Products (RAGE): Promising Targets Aiming at the Treatment of Neurodegenerative Conditions. Curr. Neuropharmacol. 2023, 21, 219–234. [Google Scholar]
- Wang, Z.C.; Zhao, J.; Li, S. Dysregulation of synaptic and extrasynaptic N-methyl-D-aspartate receptors induced by amyloid-β. Neurosci. Bull. 2013, 29, 752–760. [Google Scholar]
- Ni, R.; Marutle, A.; Nordberg, A. Modulation of α7 nicotinic acetylcholine receptor and fibrillar amyloid-β interactions in Alzheimer’s disease brain. J. Alzheimers Dis. 2013, 33, 841–851. [Google Scholar]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 87–95. [Google Scholar]
- Opazo, C.; Huang, X.; Cherny, R.A.; Moir, R.D.; Roher, A.E.; White, A.R.; Cappai, R.; Masters, C.L.; Tanzi, R.E.; Inestrosa, N.C.; et al. Metalloenzyme-like activity of Alzheimer’s disease beta-amyloid. Cu-dependent catalytic conversion of dopamine, cholesterol, and biological reducing agents to neurotoxic H(2)O(2). J. Biol. Chem. 2002, 277, 40302–40308. [Google Scholar]
- Guo, T.; Noble, W.; Hanger, D.P. Roles of tau protein in health and disease. Acta Neuropathol. 2017, 133, 665–704. [Google Scholar] [CrossRef] [PubMed]
- Mietelska-Porowska, A.; Wasik, U.; Goras, M.; Filipek, A.; Niewiadomska, G. Tau protein modifications and interactions: Their role in function and dysfunction. Int. J. Mol. Sci. 2014, 15, 4671–4713. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.T.; Patel, T.; Kang, S.G.; Yarahmady, A.; Srinivasan, M.; Julien, O.; Heras, J.; Mok, S.A. Disease-Associated Mutations in Tau Encode for Changes in Aggregate Structure Conformation. ACS Chem. Neurosci. 2023, 14, 4282–4297. [Google Scholar] [CrossRef] [PubMed]
- Rawat, P.; Sehar, U.; Bisht, J.; Selman, A.; Culberson, J.; Reddy, P.H. Phosphorylated Tau in Alzheimer’s Disease and Other Tauopathies. Int. J. Mol. Sci. 2022, 23, 12841. [Google Scholar] [CrossRef]
- Sinsky, J.; Pichlerova, K.; Hanes, J. Tau Protein Interaction Partners and Their Roles in Alzheimer’s Disease and Other Tauopathies. Int. J. Mol. Sci. 2021, 22, 9207. [Google Scholar] [CrossRef]
- Goedert, M.; Jakes, R. Mutations causing neurodegenerative tauopathies. Biochim. Biophys. Acta 2005, 1739, 240–250. [Google Scholar]
- Ramkumar, A.; Jong, B.Y.; Ori-McKenney, K.M. ReMAPping the microtubule landscape: How phosphorylation dictates the activities of microtubule-associated proteins. Dev. Dyn. 2018, 247, 138–155. [Google Scholar]
- Reddy, P.H.; Reddy, T.P. Mitochondria as a therapeutic target for aging and neurodegenerative diseases. Curr. Alzheimer Res. 2011, 8, 393–409. [Google Scholar]
- Tolnay, M.; Probst, A. The neuropathological spectrum of neurodegenerative tauopathies. IUBMB Life 2003, 55, 299–305. [Google Scholar]
- Reitz, C.; Brayne, C.; Mayeux, R. Epidemiology of Alzheimer disease. Nat. Rev. Neurol. 2011, 7, 137–152. [Google Scholar]
- Zetterberg, H.; Mattsson, N. Understanding the cause of sporadic Alzheimer’s disease. Expert. Rev. Neurother. 2014, 14, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Guglielmotto, M.; Aragno, M.; Autelli, R.; Giliberto, L.; Novo, E.; Colombatto, S.; Danni, O.; Parola, M.; Smith, M.A.; Perry, G.; et al. The up-regulation of BACE1 mediated by hypoxia and ischemic injury: Role of oxidative stress and HIF1alpha. J. Neurochem. 2009, 108, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Atwood, C.S.; Hartshorn, M.A.; Multhaup, G.; Goldstein, L.E.; Scarpa, R.C.; Cuajungco, M.P.; Gray, D.N.; Lim, J.; Moir, R.D.; et al. The A beta peptide of Alzheimer’s disease directly produces hydrogen peroxide through metal ion reduction. Biochemistry 1999, 38, 7609–7616. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R.; Oliver, C.N. Metal-catalyzed oxidation of proteins. Physiological consequences. J. Biol. Chem. 1991, 266, 2005–2008. [Google Scholar] [CrossRef]
- Islam, F.; Shohag, S.; Akhter, S.; Islam, M.R.; Sultana, S.; Mitra, S.; Chandran, D.; Khandaker, M.U.; Ashraf, G.M.; Idris, A.M.; et al. Exposure of metal toxicity in Alzheimer’s disease: An extensive review. Front. Pharmacol. 2022, 13, 903099. [Google Scholar] [CrossRef]
- Kawahara, M.; Kato-Negishi, M.; Tanaka, K.I. Dietary Trace Elements and the Pathogenesis of Neurodegenerative Diseases. Nutrients 2023, 15, 2067. [Google Scholar] [CrossRef]
- Ramos-Cejudo, J.; Wisniewski, T.; Marmar, C.; Zetterberg, H.; Blennow, K.; de Leon, M.J.; Fossati, S. Traumatic Brain Injury and Alzheimer’s Disease: The Cerebrovascular Link. EBioMedicine 2018, 28, 21–30. [Google Scholar] [CrossRef]
- McKee, A.C.; Stern, R.A.; Nowinski, C.J.; Stein, T.D.; Alvarez, V.E.; Daneshvar, D.H.; Lee, H.S.; Wojtowicz, S.M.; Hall, G.; Baugh, C.M.; et al. The spectrum of disease in chronic traumatic encephalopathy. Brain 2013, 136 Pt 1, 43–64. [Google Scholar] [CrossRef]
- Washington, P.M.; Morffy, N.; Parsadanian, M.; Zapple, D.N.; Burns, M.P. Experimental traumatic brain injury induces rapid aggregation and oligomerization of amyloid-beta in an Alzheimer’s disease mouse model. J. Neurotrauma 2014, 31, 125–134. [Google Scholar] [CrossRef]
- McKee, A.C.; Cairns, N.J.; Dickson, D.W.; Folkerth, R.D.; Keene, C.D.; Litvan, I.; Perl, D.P.; Stein, T.D.; Vonsattel, J.P.; Stewart, W.; et al. TBI/CTE group. The first NINDS/NIBIB consensus meeting to define neuropathological criteria for the diagnosis of chronic traumatic encephalopathy. Acta Neuropathol. 2016, 131, 75–86. [Google Scholar] [CrossRef]
- Zotova, E.; Nicoll, J.A.; Kalaria, R.; Holmes, C.; Boche, D. Inflammation in Alzheimer’s disease: Relevance to pathogenesis and therapy. Alzheimers Res. Ther. 2010, 2, 1. [Google Scholar] [PubMed]
- Smith, J.A.; Das, A.; Ray, S.K.; Banik, N.L. Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Res. Bull. 2012, 87, 10–20. [Google Scholar] [PubMed]
- Weisman, D.; Hakimian, E.; Ho, G.J. Interleukins, inflammation, and mechanisms of Alzheimer’s disease. Vitam. Horm. 2006, 74, 505–530. [Google Scholar] [PubMed]
- Rubio-Perez, J.M.; Morillas-Ruiz, J.M. A review: Inflammatory process in Alzheimer’s disease, role of cytokines. Sci. World J. 2012, 2012, 756357. [Google Scholar]
- Mrak, R.E.; Griffin, W.S. Potential inflammatory biomarkers in Alzheimer’s disease. J. Alzheimers Dis. 2005, 8, 369–375. [Google Scholar]
- Banks, W.A.; Farr, S.A.; Morley, J.E. Entry of blood-borne cytokines into the central nervous system: Effects on cognitive processes. Neuroimmunomodulation 2003, 10, 319–327. [Google Scholar]
- Shaftel, S.S.; Griffin, W.S.; O’Banion, M.K. The role of interleukin-1 in neuroinflammation and Alzheimer disease: An evolving perspective. J. Neuroinflammation 2008, 5, 7. [Google Scholar] [CrossRef]
- Ott, A.; Stolk, R.P.; van Harskamp, F.; Pols, H.A.; Hofman, A.; Breteler, M.M. Diabetes mellitus and the risk of dementia: The Rotterdam Study. Neurology 1999, 53, 1937–1942. [Google Scholar]
- Neumann, K.F.; Rojo, L.; Navarrete, L.P.; Farías, G.; Reyes, P.; Maccioni, R.B. Insulin resistance and Alzheimer’s disease: Molecular links & clinical implications. Curr. Alzheimer Res. 2008, 5, 438–447. [Google Scholar]
- Son, S.M.; Song, H.; Byun, J.; Park, K.S.; Jang, H.C.; Park, Y.J.; Mook-Jung, I. Altered APP processing in insulin-resistant conditions is mediated by autophagosome accumulation via the inhibition of mammalian target of rapamycin pathway. Diabetes 2012, 61, 3126–3138. [Google Scholar]
- Bjorkhem, I.; Meaney, S. Brain cholesterol: Long secret life behind a barrier. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 806–815. [Google Scholar] [PubMed]
- Dietschy, J.M.; Turley, S.D. Cholesterol metabolism in the brain. Curr. Opin. Lipidol. 2001, 12, 105–112. [Google Scholar] [PubMed]
- Simons, M.; Keller, P.; Dichgans, J.; Schulz, J.B. Cholesterol and Alzheimer’s disease: Is there a link? Neurology 2001, 57, 1089–1093. [Google Scholar] [PubMed]
- Barnes, D.E.; Yaffe, K. The projected effect of risk factor reduction on Alzheimer’s disease prevalence. Lancet Neurol. 2011, 10, 819–828. [Google Scholar]
- Durazzo, T.C.; Mattsson, N.; Weiner, M.W. Smoking and increased Alzheimer’s disease risk: A review of potential mechanisms. Alzheimers Dement. 2014, 10, S122–S145. [Google Scholar]
- Rueff-Barroso, C.R.; Trajano, E.T.; Alves, J.N.; Paiva, R.O.; Lanzetti, M.; Pires, K.M.; Bezerra, F.S.; Pinho, R.A.; Valenca, S.S.; Porto, L.C. Organ-related cigarette smoke-induced oxidative stress is strain-dependent. Med. Sci. Monit. 2010, 16, BR218–BR226. [Google Scholar]
- Ho, Y.S.; Yang, X.; Yeung, S.C.; Chiu, K.; Lau, C.F.; Tsang, A.W.; Mak, J.C.; Chang, R.C. Cigarette smoking accelerated brain aging and induced pre-Alzheimer-like neuropathology in rats. PLoS ONE 2012, 7, e36752. [Google Scholar]
- Khanna, A.; Guo, M.; Mehra, M.; Royal, W., 3rd. Inflammation and oxidative stress induced by cigarette smoke in Lewis rat brains. J. Neuroimmunol. 2013, 254, 69–75. [Google Scholar]
- Calder, P.C.; Bosco, N.; Bourdet-Sicard, R.; Capuron, L.; Delzenne, N.; Doré, J.; Franceschi, C.; Lehtinen, M.J.; Recker, T.; Salvioli, S.; et al. Health relevance of the modification of low grade inflammation in ageing (inflammageing) and the role of nutrition. Ageing Res. Rev. 2017, 40, 95–119. [Google Scholar]
- Miquel, S.; Champ, C.; Day, J.; Aarts, E.; Bahr, B.A.; Bakker, M.; Bánáti, D.; Calabrese, V.; Cederholm, T.; Cryan, J.; et al. Poor cognitive ageing: Vulnerabilities, mechanisms and the impact of nutritional interventions. Ageing Res. Rev. 2018, 42, 40–55. [Google Scholar]
- McGrattan, A.M.; McGuinness, B.; McKinley, M.C.; Kee, F.; Passmore, P.; Woodside, J.V.; McEvoy, C.T. Diet and Inflammation in Cognitive Ageing and Alzheimer’s Disease. Curr. Nutr. Rep. 2019, 8, 53–65. [Google Scholar] [PubMed]
- Peng, B.; Yang, Q.B.; Joshi, R.; Liu, Y.; Akbar, M.; Song, B.J.; Zhou, S.; Wang, X. Role of Alcohol Drinking in Alzheimer’s Disease, Parkinson’s Disease, and Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2020, 21, 2316. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Risk Reduction of Cognitive Decline and Dementia: WHO Guidelines; World Health Organization: Geneva, Switzerland, 2019; Available online: https://www.who.int/publications/i/item/9789241550543 (accessed on 20 January 2024).
- Kuusisto, J.; Koivisto, K.; Kervinen, K.; Mykkänen, L.; Helkala, E.L.; Vanhanen, M.; Hänninen, T.; Pyörälä, K.; Kesäniemi, Y.A.; Riekkinen, P.; et al. Association of apolipoprotein E phenotypes with late onset Alzheimer’s disease: Population based study. BMJ 1994, 309, 636–638. [Google Scholar] [PubMed]
- Barabash, A.; Marcos, A.; Ancín, I.; Vázquez-Alvarez, B.; de Ugarte, C.; Gil, P.; Fernández, C.; Encinas, M.; López-Ibor, J.J.; Cabranes, J.A. APOE, ACT and CHRNA7 genes in the conversion from amnestic mild cognitive impairment to Alzheimer’s disease. Neurobiol. Aging 2009, 30, 1254–1264. [Google Scholar] [CrossRef]
- Karch, C.M.; Goate, A.M. Alzheimer’s disease risk genes and mechanisms of disease pathogenesis. Biol. Psychiatry 2015, 77, 43–51. [Google Scholar]
- Kamat, P.K. Streptozotocin induced Alzheimer’s disease like changes and the underlying neural degeneration and regeneration mechanism. Neural Regen. Res. 2015, 10, 1050–1052. [Google Scholar]
- Salkovic-Petrisic, M.; Knezovic, A.; Hoyer, S.; Riederer, P. What have we learned from the streptozotocin-induced animal model of sporadic Alzheimer’s disease, about the therapeutic strategies in Alzheimer’s research. J. Neural Transm. 2013, 120, 233–252. [Google Scholar]
- Rai, S.; Kamat, P.K.; Nath, C.; Shukla, R. Glial activation and post-synaptic neurotoxicity: The key events in Streptozotocin (ICV) induced memory impairment in rats. Pharmacol. Biochem. Behav. 2014, 117, 104–117. [Google Scholar]
- Labak, M.; Foniok, T.; Kirk, D.; Rushforth, D.; Tomanek, B.; Jasiński, A.; Grieb, P. Metabolic changes in rat brain following intracerebroventricular injections of streptozotocin: A model of sporadic Alzheimer’s disease. Acta Neurochir. Suppl. 2010, 106, 177–181. [Google Scholar]
- Chen, Y.; Liang, Z.; Blanchard, J.; Dai, C.L.; Sun, S.; Lee, M.H.; Grundke-Iqbal, I.; Iqbal, K.; Liu, F.; Gong, C.X. A non-transgenic mouse model (icv-STZ mouse) of Alzheimer’s disease: Similarities to and differences from the transgenic model (3xTg-AD mouse). Mol. Neurobiol. 2013, 47, 711–725. [Google Scholar]
- Chen, Y.; Liang, Z.; Tian, Z.; Blanchard, J.; Dai, C.L.; Chalbot, S.; Iqbal, K.; Liu, F.; Gong, C.X. Intracerebroventricular streptozotocin exacerbates Alzheimer-like changes of 3xTg-AD mice. Mol. Neurobiol. 2014, 49, 547–562. [Google Scholar]
- Grieb, P. Intracerebroventricular Streptozotocin Injections as a Model of Alzheimer’s Disease: In Search of a Relevant Mechanism. Mol. Neurobiol. 2016, 53, 1741–1752. [Google Scholar] [CrossRef]
- Tang, K.S. The cellular and molecular processes associated with scopolamine-induced memory deficit: A model of Alzheimer’s biomarkers. Life Sci. 2019, 233, 116695. [Google Scholar]
- Morley, J.E.; Armbrecht, H.J.; Farr, S.A.; Kumar, V.B. The senescence accelerated mouse (SAMP8) as a model for oxidative stress and Alzheimer’s disease. Biochim. Biophys. Acta 2012, 1822, 650–656. [Google Scholar]
- Levy Nogueira, M.; Epelbaum, S.; Steyaert, J.M.; Dubois, B.; Schwartz, L. Mechanical stress models of Alzheimer’s disease pathology. Alzheimers Dement. 2016, 12, 324–333. [Google Scholar]
- Song, J. Animal Model of Aluminum-Induced Alzheimer’s Disease. Adv. Exp. Med. Biol. 2018, 1091, 113–127. [Google Scholar]
- Valentin-Escalera, J.; Leclerc, M.; Calon, F. High-Fat Diets in Animal Models of Alzheimer’s Disease: How Can Eating Too Much Fat Increase Alzheimer’s Disease Risk? J. Alzheimers. Dis. 2024, 97, 977–1005. [Google Scholar]
- Bagheri, M.; Joghataei, M.T.; Mohseni, S.; Roghani, M. Genistein ameliorates learning and memory deficits in amyloid β(1–40) rat model of Alzheimer’s disease. Neurobiol. Learn Mem. 2011, 95, 270–276. [Google Scholar]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the Cholinergic System. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef]
- Safar, M.M.; Arab, H.H.; Rizk, S.M.; El-Maraghy, S.A. Bone Marrow-Derived Endothelial Progenitor Cells Protect Against Scopolamine-Induced Alzheimer-Like Pathological Aberrations. Mol. Neurobiol. 2016, 53, 1403–1418. [Google Scholar] [CrossRef]
- Llorens-Martín, M.; Jurado, J.; Hernández, F.; Avila, J. GSK-3β, a pivotal kinase in Alzheimer disease. Front. Mol. Neurosci. 2014, 7, 46. [Google Scholar]
- Lu, C.; Wang, Y.; Xu, T.; Li, Q.; Wang, D.; Zhang, L.; Fan, B.; Wang, F.; Liu, X. Genistein Ameliorates Scopolamine-Induced Amnesia in Mice Through the Regulation of the Cholinergic Neurotransmission, Antioxidant System and the ERK/CREB/BDNF Signaling. Front. Pharmacol. 2018, 9, 1153. [Google Scholar]
- Sun, D.; Unnithan, R.R.; French, C. Scopolamine Impairs Spatial Information Recorded With “Miniscope” Calcium Imaging in Hippocampal Place Cells. Front. Neurosci. 2021, 15, 640350. [Google Scholar]
- Shen, H.; Zheng, Y.; Chen, R.; Huang, X.; Shi, G. Neuroprotective effects of quercetin 3-O-sophoroside from Hibiscus rosa-sinensis Linn. on scopolamine-induced amnesia in mice. J. Funct. Foods 2021, 76, 104291. [Google Scholar] [CrossRef]
- Ghasemi, S.; Moradzadeh, M.; Hosseini, M.; Beheshti, F.; Sadeghnia, H.R. Beneficial effects of Urtica dioica on scopolamine-induced memory impairment in rats: Protection against acetylcholinesterase activity and neuronal oxidative damage. Drug Chem. Toxicol. 2019, 42, 167–175. [Google Scholar]
- Kameyama, T.; Nabeshima, T.; Kozawa, T. Step-down-type passive avoidance- and escape-learning method. Suitability for experimental amnesia models. J. Pharmacol. Methods 1986, 16, 39–52. [Google Scholar]
- Utkan, T.; Gocmez, S.S.; Ozer, C.; Gacar, N.; Aricioglu, F. Selective and nonselective neuronal NOS inhibitors impair cognitive function in the three panel runway and passive avoidance tasks in rats. Pharmacol. Biochem. Behav. 2012, 101, 515–519. [Google Scholar] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzym. 1978, 52, 302–310. [Google Scholar]
- Sanchez-Varo, R.; Mejias-Ortega, M.; Fernandez-Valenzuela, J.J.; Nu√±ez-Diaz, C.; Caceres-Palomo, L.; Vegas-Gomez, L.; Sanchez-Mejias, E.; Trujillo-Estrada, L.; Garcia-Leon, J.A.; Moreno-Gonzalez, I.; et al. Transgenic Mouse Models of Alzheimer’s Disease: An Integrative Analysis. Int. J. Mol. Sci. 2022, 23, 5404. [Google Scholar] [CrossRef]
- Morley, J.E.; Farr, S.A.; Kumar, V.B.; Armbrecht, H.J. The SAMP8 mouse: A model to develop therapeutic interventions for Alzheimer’s disease. Curr. Pharm. Des. 2012, 18, 1123–1130. [Google Scholar]
- Kurokawa, T.; Ozaki, N.; Ishibashi, S. Difference between senescence-accelerated prone and resistant mice in response to insulin in the heart. Mech. Ageing Dev. 1998, 102, 25–32. [Google Scholar] [CrossRef]
- Pallas, M.; Camins, A.; Smith, M.A.; Perry, G.; Lee, H.G.; Casadesus, G. From aging to Alzheimer’s disease: Unveiling “the switch” with the senescence-accelerated mouse model (SAMP8). J. Alzheimers Dis. 2008, 15, 615–624. [Google Scholar] [CrossRef]
- Takeda, T. Senescence-accelerated mouse (SAM) with special references to neurodegeneration models, SAMP8 and SAMP10 mice. Neurochem. Res. 2009, 34, 639–659. [Google Scholar] [CrossRef]
- Cheng, X.R.; Zhou, W.X.; Zhang, Y.X. The behavioral, pathological and therapeutic features of the senescence-accelerated mouse prone 8 strain as an Alzheimer’s disease animal model. Ageing Res. Rev. 2014, 13, 13–37. [Google Scholar] [CrossRef]
- Rebrin, I.; Zicker, S.; Wedekind, K.J.; Paetau-Robinson, I.; Packer, L.; Sohal, R.S. Effect of antioxidant-enriched diets on glutathione redox status in tissue homogenates and mitochondria of the senescence-accelerated mouse. Free Radic. Biol. Med. 2005, 39, 549–557. [Google Scholar] [CrossRef]
- Prieto, R.; Gutiérrez-González, R.; Pascual, J.M.; Roda, J.M.; Cerdán, S.; Matias-Guiu, J.; Barcia, J.A. Modelos experimentales de traumatismo craneoencefálico. Experimental models of traumatic brain injury. Neurocirugia 2009, 20, 225–244. [Google Scholar] [CrossRef]
- Ojo, J.O.; Mouzon, B.; Greenberg, M.B.; Bachmeier, C.; Mullan, M.; Crawford, F. Repetitive mild traumatic brain injury augments tau pathology and glial activation in aged hTau mice. J. Neuropathol. Exp. Neurol. 2013, 72, 137–151. [Google Scholar] [CrossRef]
- Breunig, J.J.; Guillot-Sestier, M.V.; Town, T. Brain injury, neuroinflammation and Alzheimer’s disease. Front. Aging Neurosci. 2013, 5, 26. [Google Scholar] [CrossRef]
- Payno, M.; Villanueva, C.; Matías-Guiu, J. Modelos experimentales de la enfermedad de Alzheimer. Experimental models in Alzheimer’s disease. Neurologia 2009, 24, 255–262. [Google Scholar]
- Levy Nogueira, M.; Hamraz, M.; Abolhassani, M.; Bigan, E.; Lafitte, O.; Steyaert, J.M.; Dubois, B.; Schwartz, L. Mechanical stress increases brain amyloid β, tau, and α-synuclein concentrations in wild-type mice. Alzheimers Dement. 2018, 14, 444–453. [Google Scholar] [CrossRef]
- Lovell, M.A.; Robertson, J.D.; Teesdale, W.J.; Campbell, J.L.; Markesbery, W.R. Copper, iron and zinc in Alzheimer’s disease senile plaques. J. Neurol. Sci. 1998, 158, 47–52. [Google Scholar] [CrossRef]
- Huang, X.; Atwood, C.S.; Moir, R.D.; Hartshorn, M.A.; Vonsattel, J.P.; Tanzi, R.E.; Bush, A.I. Zinc-induced Alzheimer’s Abeta1-40 aggregation is mediated by conformational factors. J. Biol. Chem. 1997, 272, 26464–26470. [Google Scholar] [CrossRef]
- Campbell, A. The potential role of aluminium in Alzheimer’s disease. Nephrol. Dial. Transplant. 2002, 17 (Suppl. S2), 17–20. [Google Scholar] [CrossRef]
- Mitra, J.; Guerrero, E.N.; Hegde, P.M.; Wang, H.; Boldogh, I.; Rao, K.S.; Mitra, S.; Hegde, M.L. New perspectives on oxidized genome damage and repair inhibition by pro-oxidant metals in neurological diseases. Biomolecules 2014, 4, 678–703. [Google Scholar] [CrossRef]
- Kawahara, M.; Kato-Negishi, M. Link between Aluminum and the Pathogenesis of Alzheimer’s Disease: The Integration of the Aluminum and Amyloid Cascade Hypotheses. Int. J. Alzheimers Dis. 2011, 2011, 276393. [Google Scholar] [CrossRef]
- Lukiw, W.J.; LeBlanc, H.J.; Carver, L.A.; McLachlan, D.R.; Bazan, N.G. Run-on gene transcription in human neocortical nuclei. Inhibition by nanomolar aluminum and implications for neurodegenerative disease. J. Mol. Neurosci. 1998, 11, 67–78. [Google Scholar] [CrossRef]
- Scott, C.W.; Fieles, A.; Sygowski, L.A.; Caputo, C.B. Aggregation of tau protein by aluminum. Brain Res. 1993, 628, 77–84. [Google Scholar] [CrossRef]
- Walton, J.R.; Wang, M.X. APP expression, distribution and accumulation are altered by aluminum in a rodent model for Alzheimer’s disease. J. Inorg. Biochem. 2009, 103, 1548–1554. [Google Scholar] [CrossRef]
- Bitra, V.R.; Rapaka, D.; Mathala, N.; Akula, A. Effect of wheat grass powder on aluminum induced Alzheimer’s disease in Wistar rats. Asian Pac. J. Trop. Med. 2014, 7, S278–S281. [Google Scholar] [CrossRef]
- Bazzari, F.H.; Abdallah, D.M.; El-Abhar, H.S. Chenodeoxycholic Acid Ameliorates AlCl3-Induced Alzheimer’s Disease Neurotoxicity and Cognitive Deterioration via Enhanced Insulin Signaling in Rats. Molecules 2019, 24, 1992. [Google Scholar] [CrossRef]
- Liaquat, L.; Ahmad, S.; Sadir, S.; Batool, Z.; Khaliq, S.; Tabassum, S.; Emad, S.; Madiha, S.; Shahzad, S.; Haider, S. Development of AD like symptoms following co-administration of AlCl3 and d-gal in rats: A neurochemical, biochemical and behavioural study. Pak. J. Pharm. Sci. 2017, 30 (Suppl. S2), 647–653. [Google Scholar]
- Haider, S.; Liaquat, L.; Ahmad, S.; Batool, Z.; Siddiqui, R.A.; Tabassum, S.; Shahzad, S.; Rafiq, S.; Naz, N. Naringenin protects AlCl3/d-galactose induced neurotoxicity in rat model of AD via attenuation of acetylcholinesterase levels and inhibition of oxidative stress. PLoS ONE 2020, 15, e0227631. [Google Scholar]
- Livingston, G.; Sommerlad, A.; Orgeta, V.; Costafreda, S.G.; Huntley, J.; Ames, D.; Ballard, C.; Banerjee, S.; Burns, A.; Cohen-Mansfield, J.; et al. Dementia prevention, intervention, and care. Lancet 2017, 390, 2673–2734. [Google Scholar]
- Samadi, M.; Moradi, S.; Moradinazar, M.; Mostafai, R.; Pasdar, Y. Dietary pattern in relation to the risk of Alzheimer’s disease: A systematic review. Neurol. Sci. 2019, 40, 2031–2043. [Google Scholar] [CrossRef]
- Hill, E.; Clifton, P.; Goodwill, A.M.; Dennerstein, L.; Campbell, S.; Szoeke, C. Dietary patterns and β-amyloid deposition in aging Australian women. Alzheimers Dement. 2018, 4, 535–541. [Google Scholar]
- Pugazhenthi, S.; Qin, L.; Reddy, P.H. Common neurodegenerative pathways in obesity, diabetes, and Alzheimer’s disease. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1037–1045. [Google Scholar]
- Nuzzo, D.; Picone, P.; Baldassano, S.; Caruana, L.; Messina, E.; Gammazza, A.; Cappello, F.; Mulè, F.; Carlo, M. Insulin resistance as common molecular denominator linking obesity to Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 723–735. [Google Scholar]
- Baranowski, B.J.; Bott, K.N.; MacPherson, R.E.K. Evaluation of neuropathological effects of a high-fat high-sucrose diet in middle-aged male C57BL6/J mice. Physiol. Rep. 2018, 6, e13729. [Google Scholar] [CrossRef]
- Busquets, O.; Ettcheto, M.; Pallàs, M.; Beas-Zarate, C.; Verdaguer, E.; Auladell, C.; Folch, J.; Camins, A. Long-term exposition to a high fat diet favors the appearance of β-amyloid depositions in the brain of C57BL/6J mice. A potential model of sporadic Alzheimer’s disease. Mech. Ageing Dev. 2017, 162, 38–45. [Google Scholar]
- Niu, L.; Han, D.W.; Xu, R.L.; Han, B.; Zhou, X.; Wu, H.W.; Li, S.H.; Qu, C.X.; Liu, M. A high-sugar high-fat diet induced metabolic syndrome shows some symptoms of Alzheimer’s disease in rats. J. Nutr. Heal. Aging 2016, 20, 509–513. [Google Scholar]
- Salas, I.H.; Weerasekera, A.; Ahmed, T.; Callaerts-Vegh, Z.; Himmelreich, U.; D’Hooge, R.; Balschun, D.; Saido, T.C.; De Strooper, B.; Dotti, C.G. High fat diet treatment impairs hippocampal long-term potentiation without alterations of the core neuropathological features of Alzheimer disease. Neurobiol. Dis. 2018, 13, 82–96. [Google Scholar]
- Tucsek, Z.; Toth, P.; Sosnowska, D.; Gautam, T.; Mitschelen, M.; Koller, A.; Szalai, G.; Sonntag, W.E.; Ungvari, Z.; Csiszar, A. Obesity in aging exacerbates blood-brain barrier disruption, neuroinflammation, and oxidative stress in the mouse hippocampus: Effects on expression of genes involved in beta-amyloid generation and Alzheimer’s disease. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 1212–1226. [Google Scholar] [PubMed]
- Chang, W.; Huang, D.; Lo, Y.M.; Tee, Q.; Kuo, P.; Wu, J.S.; Huang, W.; Shen, S. Protective effect of caffeic acid against Alzheimer’s disease pathogenesis via modulating cerebral insulin signaling, β-amyloid accumulation, and synaptic plasticity in hyperinsulinemic rats. J. Agric. Food Chem. 2019, 67, 7684–7693. [Google Scholar] [CrossRef] [PubMed]
- Nakandakari, S.C.B.R.; Muñoz, V.R.; Kuga, G.K.; Gaspar, R.C.; Sant’Ana, M.R.; Pavan, I.C.B.; da Silva, L.G.S.; Morelli, A.P.; Simabuco, F.M.; da Silva, A.S.R.; et al. Short-term high-fat diet modulates several inflammatory, ER stress, and apoptosis markers in the hippocampus of young mice. Brain Behav. Immun. 2019, 79, 284–293. [Google Scholar]
- Saraceno, C.; Musardo, S.; Marcello, E.; Pelucchi, S.; Di Luca, M. Modeling Alzheimer’s disease: From past to future. Front. Pharmacol. 2013, 4, 77. [Google Scholar]
- Guo, T.; Zhang, D.; Zeng, Y.; Huang, T.Y.; Xu, H.; Zhao, Y. Molecular and cellular mechanisms underlying the pathogenesis of Alzheimer’s disease. Mol. Neurodegener. 2020, 15, 40. [Google Scholar] [CrossRef]
- Tag, S.H.; Kim, B.; Bae, J.; Chang, K.A.; Im, H.I. Neuropathological and behavioral features of an APP/PS1/MAPT (6xTg) transgenic model of Alzheimer’s disease. Mol. Brain 2022, 15, 51. [Google Scholar] [CrossRef]
- Lewis, J.; Dickson, D.W.; Lin, W.L.; Chisholm, L.; Corral, A.; Jones, G.; Yen, S.H.; Sahara, N.; Skipper, L.; Yager, D.; et al. Enhanced neurofibrillary degeneration in transgenic mice expressing mutant tau and APP. Science 2001, 293, 1487–1491. [Google Scholar] [CrossRef]
- Trejo-Lopez, J.A.; Yachnis, A.T.; Prokop, S. Neuropathology of Alzheimer’s Disease. Neurotherapeutics 2022, 19, 173–185. [Google Scholar] [CrossRef]
- Moya-Alvarado, G.; Gershoni-Emek, N.; Perlson, E.; Bronfman, F.C. Neurodegeneration and Alzheimer’s disease (AD). What Can Proteomics Tell Us About the Alzheimer’s Brain? Mol. Cell Proteom. 2016, 15, 409–425. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).