
Sound-Evoked Neural Activity in Normal-Hearing Tinnitus: Effects of Frequency and Stimulated Ear Side


Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
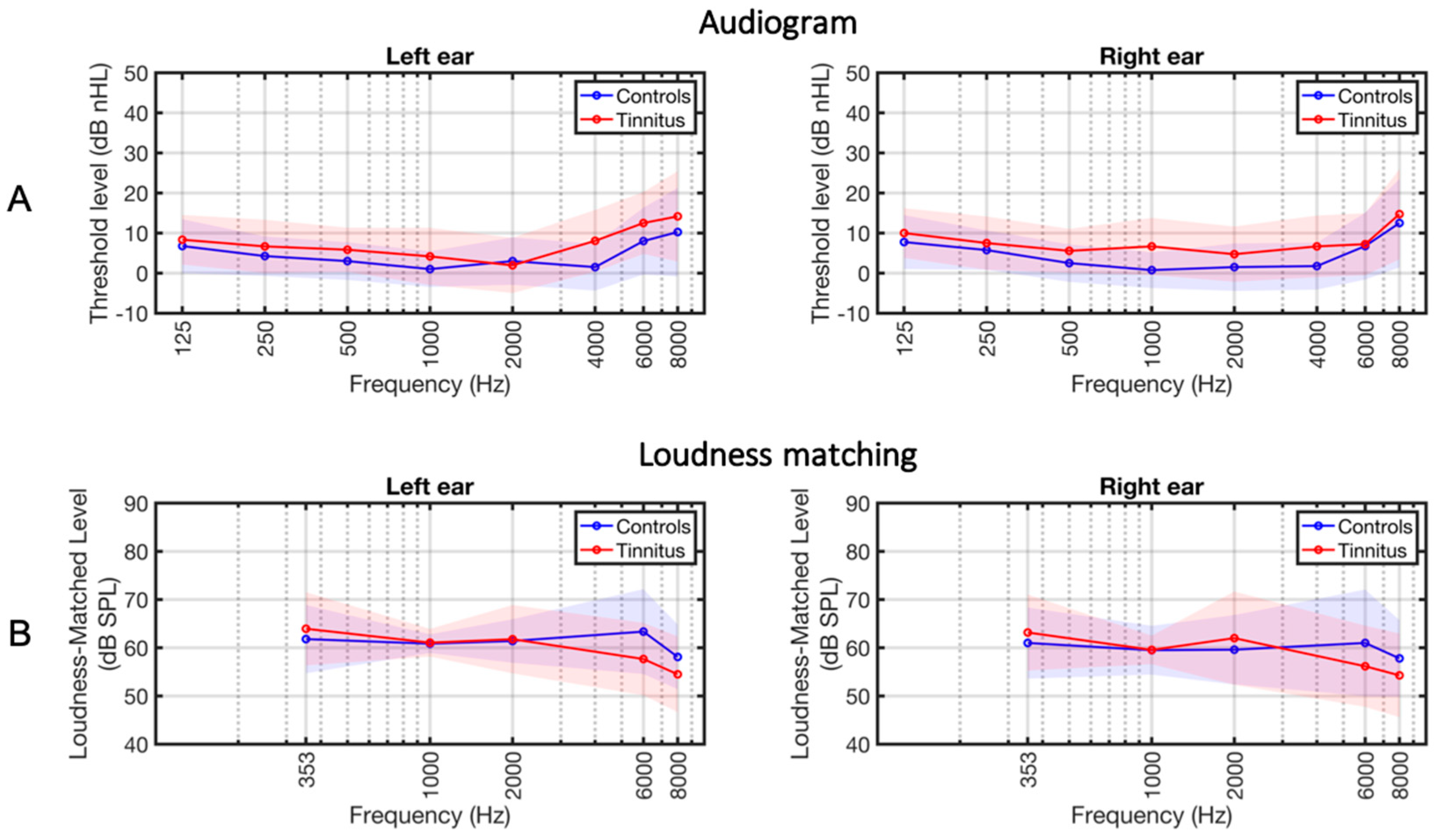
2.2. Acoustic Stimuli
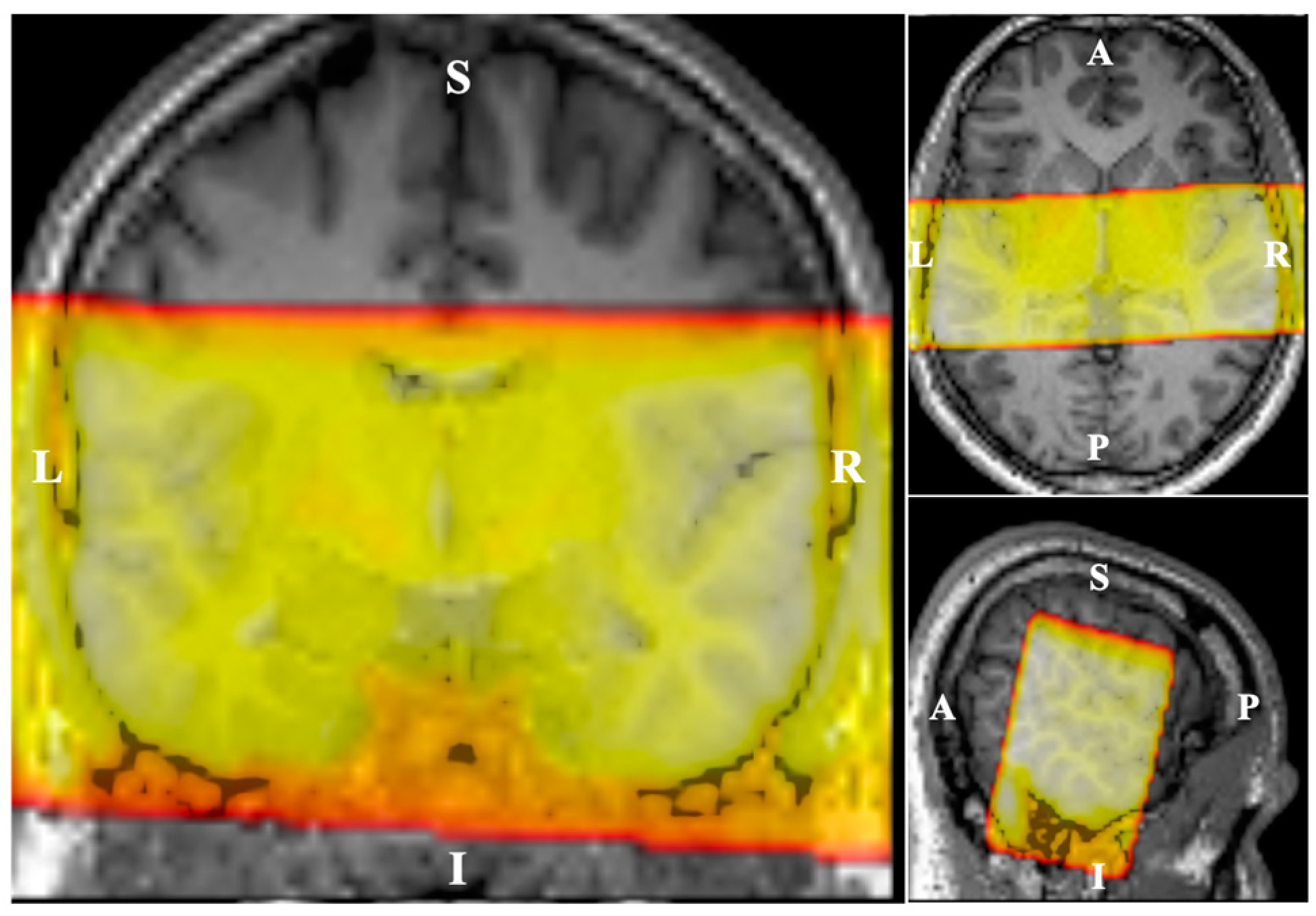
2.3. Imaging Paradigm
2.4. Data Analysis
2.4.1. Pre-Processing
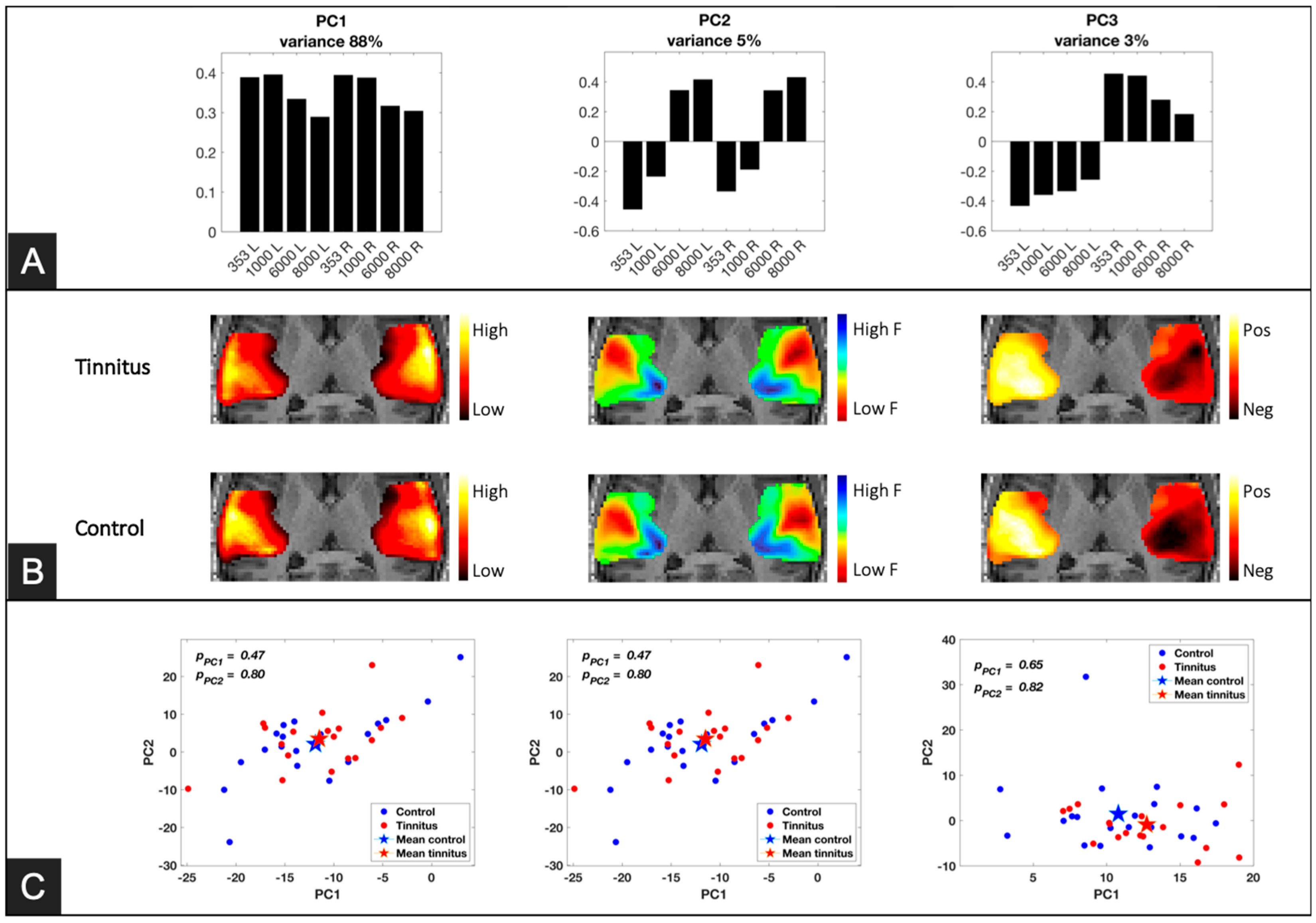
2.4.2. Principal Component Analysis (PCA)
2.4.3. Generalized Linear Model (GLM) Analysis
2.4.4. Effects of Tinnitus Severity and Hyperacusis
3. Results
3.1. Group Descriptions
3.2. Principal Component Analysis (PCA)
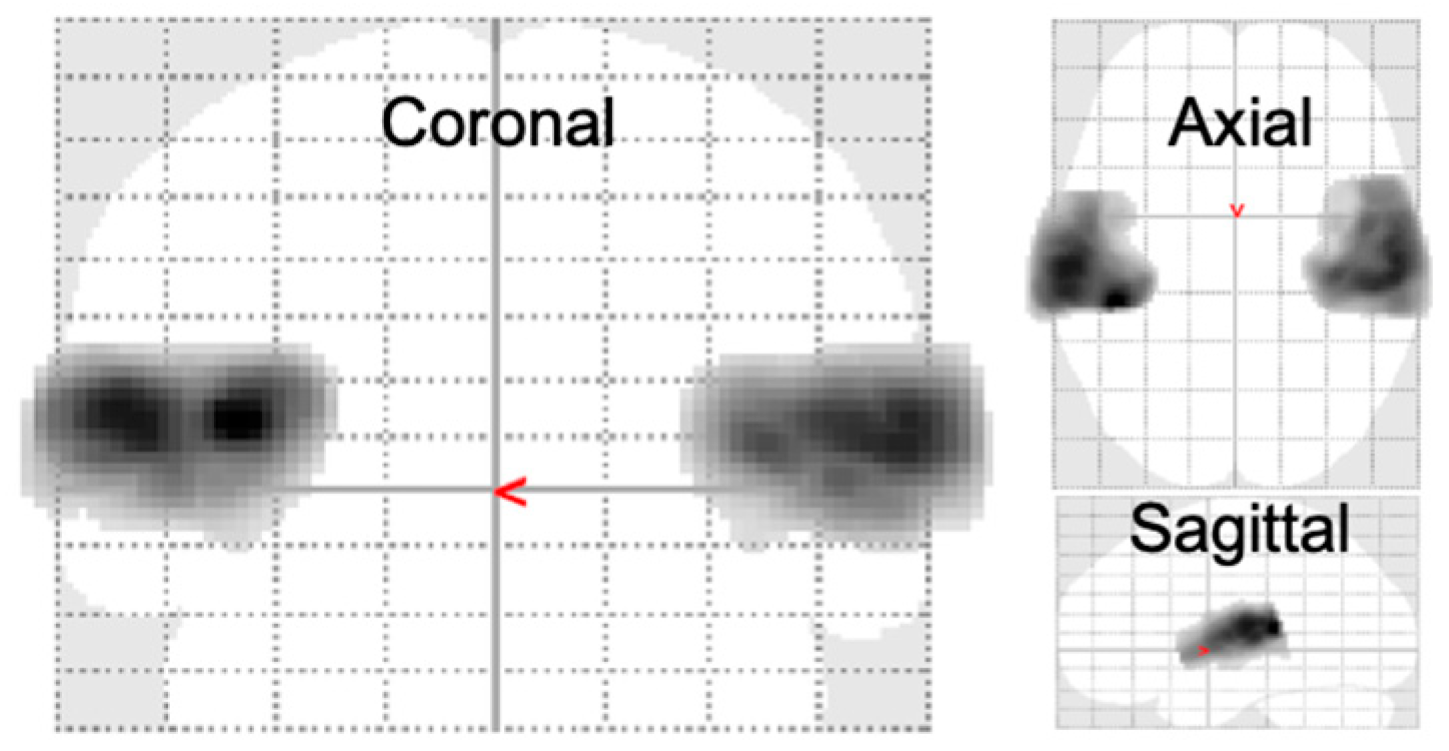
3.3. Full Factorial GLM Analysis
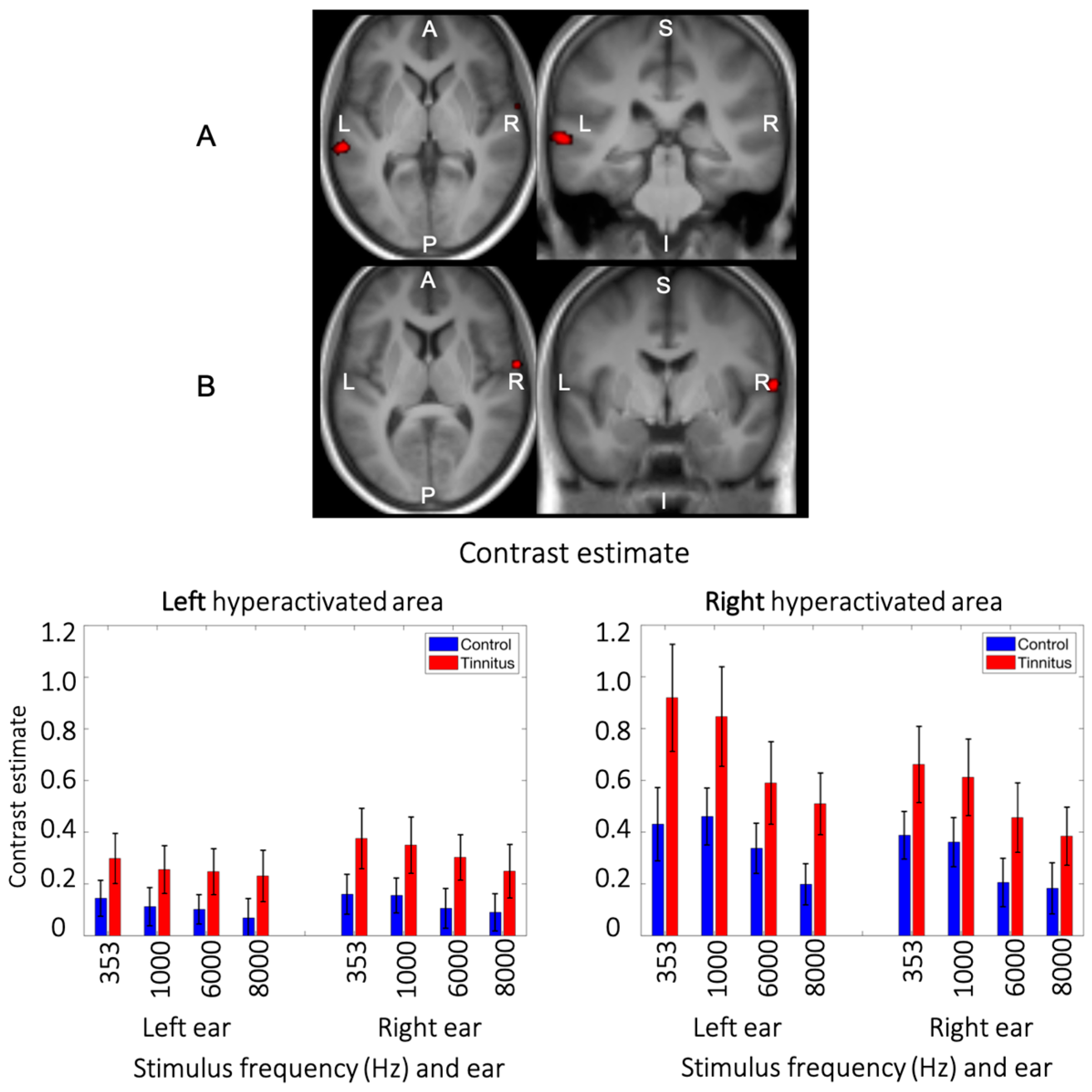
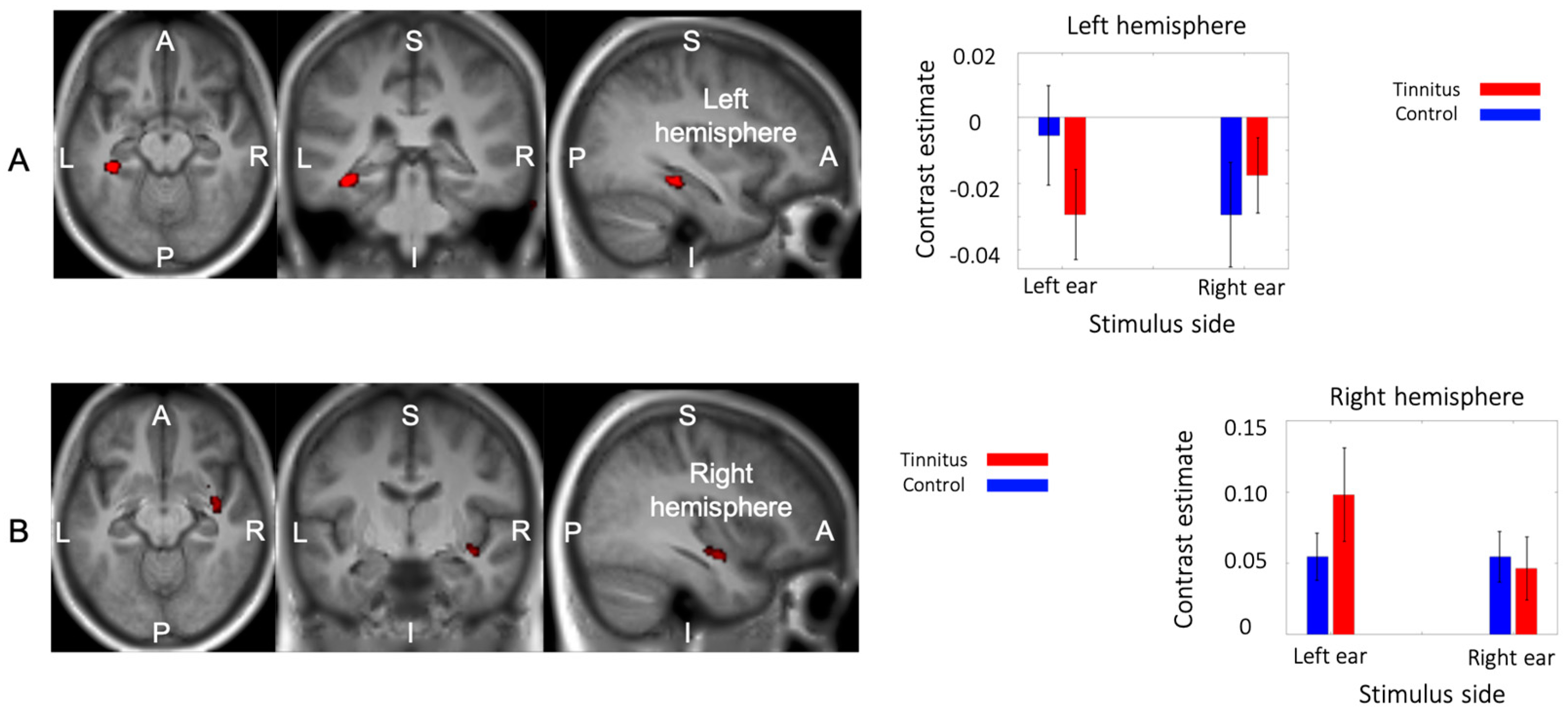
3.4. Main Effect of Group
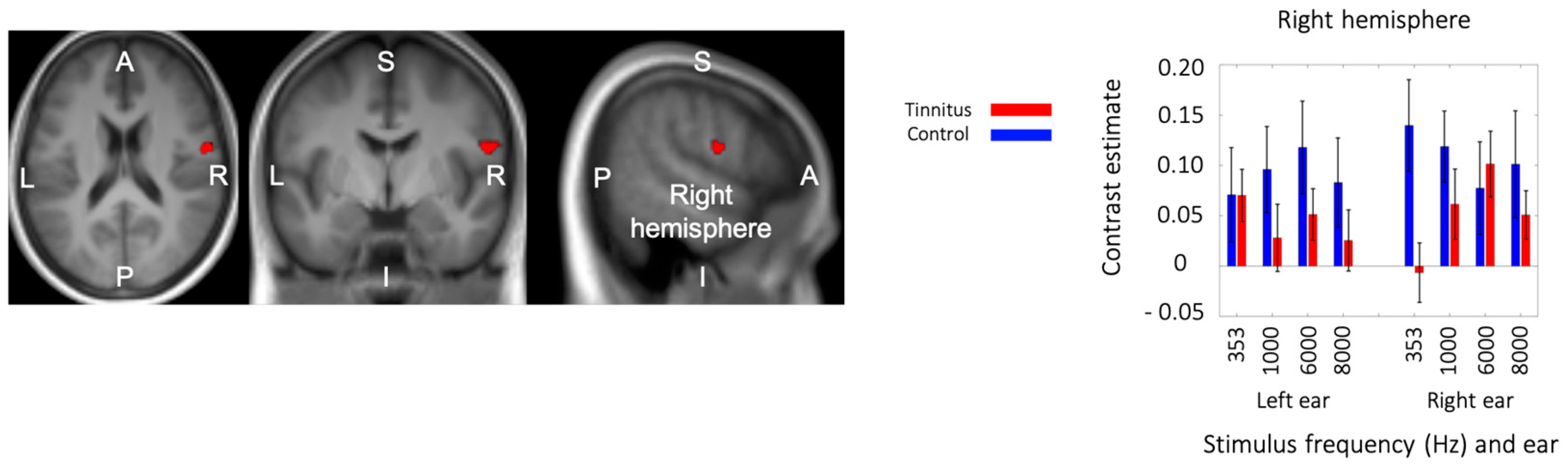
3.5. Interaction between Group and Side of Stimualtion
3.6. Triple Interaction: Group × Stimulation Side × Frequency
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baguley, D.; McFerran, D.; Hall, D. Tinnitus. Lancet 2013, 382, 1600–1607. [Google Scholar] [CrossRef] [PubMed]
- Langguth, B.; de Ridder, D.; Schlee, W.; Kleinjung, T. Tinnitus: Clinical Insights in Its Pathophysiology-A Perspective. J. Assoc. Res. Otolaryngol. 2024, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Husain, F.T.; Medina, R.E.; Davis, C.W.; Szymko-Bennett, Y.; Simonyan, K.; Pajor, N.M.; Horwitz, B. Neuroanatomical changes due to hearing loss and chronic tinnitus: A combined VBM and DTI study. Brain Res. 2011, 1369, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Noreña, A.J.; Eggermont, J.J. Changes in spontaneous neural activity immediately after an acoustic trauma: Implications for neural correlates of tinnitus. Hear. Res. 2003, 183, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Kaltenbach, J.A. Increases in spontaneous activity in the dorsal cochlear nucleus of the rat following exposure to high-intensity sound. Neurosci. Lett. 1998, 250, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Bauer, C.A.; Turner, J.G.; Caspary, D.M.; Myers, K.S.; Brozoski, T.J. Tinnitus and inferior colliculus activity in chinchillas related to three distinct patterns of cochlear trauma. J. Neurosci. Res. 2008, 86, 2564–2578. [Google Scholar] [CrossRef] [PubMed]
- Koops, E.A.; Renken, R.J.; Lanting, C.P.; van Dijk, P. Cortical Tonotopic Map Changes in Humans Are Larger in Hearing Loss Than in Additional Tinnitus. J. Neurosci. 2020, 40, 3178–3185. [Google Scholar] [CrossRef] [PubMed]
- Knipper, M.; Van Dijk, P.; Nunes, I.; Rüttiger, L.; Zimmermann, U. Advances in the neurobiology of hearing disorders: Recent developments regarding the basis of tinnitus and hyperacusis. Prog. Neurobiol. 2013, 111, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Langers, D.R.M.; de Kleine, E.; van Dijk, P. Tinnitus does not require macroscopic tonotopic map reorganization. Front. Syst. Neurosci. 2012, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.W.; Halpin, C.F.; Nam, E.-C.; Levine, R.A.; Melcher, J.R. Tinnitus, Diminished Sound-Level Tolerance, and Elevated Auditory Activity in Humans With Clinically Normal Hearing Sensitivity. J. Neurophysiol. 2010, 104, 3361–3370. [Google Scholar] [CrossRef]
- Melcher, J.R.; Levine, R.A.; Bergevin, C.; Norris, B. The auditory midbrain of people with tinnitus: Abnormal sound-evoked activity revisited. Hear. Res. 2009, 257, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Gutschalk, A.; Steinmann, I. Stimulus dependence of contralateral dominance in human auditory cortex. Hum. Brain Mapp. 2015, 36, 883–896. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Khalfa, S.; Dubal, S.; Veuillet, E.; Perez-Diaz, F.; Jouvent, R.; Collet, L. Psychometric Normalization of a Hyperacusis Questionnaire. ORL 2002, 64, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Zigmond, A.S.; Snaith, R.P. The Hospital Anxiety and Depression Scale. Acta Psychiatr. Scand. 1983, 67, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Newman, C.W.; Jacobson, G.P.; Spitzer, J.B. Development of the Tinnitus Handicap Inventory. Arch. Otolaryngol.-Head Neck Surg. 1996, 122, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Meikle, M.B.; Henry, J.A.; Griest, S.E.; Stewart, B.J.; Abrams, H.B.; McArdle, R.; Myers, P.J.; Newman, C.W.; Sandridge, S.; Turk, D.C.; et al. The Tinnitus Functional Index. Ear Hear. 2012, 33, 153–176. [Google Scholar] [CrossRef]
- Makani, P.; Thioux, M.; Pyott, S.J.; van Dijk, P. A Combined Image- and Coordinate-Based Meta-Analysis of Whole-Brain Voxel-Based Morphometry Studies Investigating Subjective Tinnitus. Brain Sci. 2022, 12, 1192. [Google Scholar] [CrossRef]
- Fuller, T.E.; Haider, H.F.; Kikidis, D.; Lapira, A.; Mazurek, B.; Norena, A.; Rabau, S.; Lardinois, R.; Cederroth, C.R.; Edvall, N.K.; et al. Different Teams, Same Conclusions? A Systematic Review of Existing Clinical Guidelines for the Assessment and Treatment of Tinnitus in Adults. Front. Psychol. 2017, 8, 206. [Google Scholar] [CrossRef]
- Cima, R.F.F.; Mazurek, B.; Haider, H.; Kikidis, D.; Lapira, A.; Noreña, A.; Hoare, D.J. A multidisciplinary European guideline for tinnitus: Diagnostics, assessment, and treatment. HNO 2019, 67, 10–42. [Google Scholar] [CrossRef]
- Hall, D.A.; Haggard, M.P.; Akeroyd, M.A.; Palmer, A.R.; Summerfield, A.Q.; Elliott, M.R.; Gurney, E.M.; Bowtell, R.W. ?sparse? temporal sampling in auditory fMRI. Hum. Brain Mapp. 1999, 7, 213–223. [Google Scholar] [CrossRef]
- Friston, K.J. Statistical Parametric Mapping. In Neuroscience Databases; Springer: Boston, MA, USA, 2003; pp. 237–250. [Google Scholar]
- Andersson, J.L.R.; Skare, S.; Ashburner, J. How to correct susceptibility distortions in spin-echo echo-planar images: Application to diffusion tensor imaging. Neuroimage 2003, 20, 870–888. [Google Scholar] [CrossRef]
- Goodpaster, A.M.; Kennedy, M.A. Quantification and statistical significance analysis of group separation in NMR-based metabonomics studies. Chemom. Intell. Lab. Syst. 2011, 109, 162–170. [Google Scholar] [CrossRef]
- Thirion, B.; Pinel, P.; Mériaux, S.; Roche, A.; Dehaene, S.; Poline, J.-B. Analysis of a large fMRI cohort: Statistical and methodological issues for group analyses. Neuroimage 2007, 35, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Aazh, H.; Moore, B.C.J. Factors related to uncomfortable loudness levels for patients seen in a tinnitus and hyperacusis clinic. Int. J. Audiol. 2017, 56, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Langers, D.R.M.; van Dijk, P. Mapping the Tonotopic Organization in Human Auditory Cortex with Minimally Salient Acoustic Stimulation. Cereb. Cortex 2012, 22, 2024–2038. [Google Scholar] [CrossRef] [PubMed]
- Lanting, C.P.; de Kleine, E.; van Dijk, P. Neural activity underlying tinnitus generation: Results from PET and fMRI. Hear. Res. 2009, 255, 1–13. [Google Scholar] [CrossRef]
- Koops, E.A.; van Dijk, P. Hyperacusis in tinnitus patients relates to enlarged subcortical and cortical responses to sound except at the tinnitus frequency. Hear. Res. 2021, 401, 108158. [Google Scholar] [CrossRef]
- Vanneste, S.; Plazier, M.; van der Loo, E.; Van de Heyning, P.; De Ridder, D. The difference between uni- and bilateral auditory phantom percept. Clin. Neurophysiol. 2011, 122, 578–587. [Google Scholar] [CrossRef]
- Maudoux, A.; Lefebvre, P.; Cabay, J.-E.; Demertzi, A.; Vanhaudenhuyse, A.; Laureys, S.; Soddu, A. Auditory Resting-State Network Connectivity in Tinnitus: A Functional MRI Study. PLoS ONE 2012, 7, e36222. [Google Scholar] [CrossRef]
- Schmidt, S.A.; Akrofi, K.; Carpenter-Thompson, J.R.; Husain, F.T. Default Mode, Dorsal Attention and Auditory Resting State Networks Exhibit Differential Functional Connectivity in Tinnitus and Hearing Loss. PLoS ONE 2013, 8, e76488. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; Vanneste, S.; Weisz, N.; Londero, A.; Schlee, W.; Elgoyhen, A.B.; Langguth, B. An integrative model of auditory phantom perception: Tinnitus as a unified percept of interacting separable subnetworks. Neurosci. Biobehav. Rev. 2014, 44, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Wang, B.; Chen, B.; Galvin, J.J.; Fu, Q.-J. Tinnitus impairs segregation of competing speech in normal-hearing listeners. Sci. Rep. 2020, 10, 19851. [Google Scholar] [CrossRef] [PubMed]
- Grabski, K.; Lamalle, L.; Vilain, C.; Schwartz, J.; Vallée, N.; Tropres, I.; Baciu, M.; Le Bas, J.; Sato, M. Functional MRI assessment of orofacial articulators: Neural correlates of lip, jaw, larynx, and tongue movements. Hum. Brain Mapp. 2012, 33, 2306–2321. [Google Scholar] [CrossRef]
- Vielsmeier, V.; Lehner, A.; Strutz, J.; Steffens, T.; Kreuzer, P.M.; Schecklmann, M.; Landgrebe, M.; Langguth, B.; Kleinjung, T. The Relevance of the High Frequency Audiometry in Tinnitus Patients with Normal Hearing in Conventional Pure-Tone Audiometry. BioMed Res. Int. 2015, 2015, 302515. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tinnitus | Control | t-Value | p-Value | |
|---|---|---|---|---|
| Age (years old) | 40.2 ± 10.5 | 29.5 ± 8.5 | 3.4392 | 0.0015 |
| Sex | 12 M, 6 F | 13 M, 7 F | 0.9139 † | |
| Hearing Threshold dB (nHL) | 7.8 ± 3.7 | 4.8 ± 4.2 | 2.2650 | 0.0295 |
| Handedness | 3 L, 15 R | 5 L, 15 R | 0.5292 † | |
| Anxiwty score | 6.4 ± 3.8 | 4.8 ± 3.3 | 1.3744 | 0.1778 |
| Depression score | 4.5 ± 3.7 | 3.2 ± 2.6 | 1.2786 | 0.209 |
| Hyperacusis score | 21 ± 7.4 | 12.8 ± 7.5 | 3.3385 | 0.002 |
| Tinnitus Handicap Inventory | 36.8 ± 21.7 | - | - | |
| Tinnitus Funcional Index | 34.5 ± 19.7 | - | - | |
| Tinnitus laterlity | 3 L, 1 R, 12 bi | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Safazadeh, S.; Thioux, M.; Renken, R.J.; van Dijk, P. Sound-Evoked Neural Activity in Normal-Hearing Tinnitus: Effects of Frequency and Stimulated Ear Side. Brain Sci. 2024, 14, 544. https://doi.org/10.3390/brainsci14060544
Safazadeh S, Thioux M, Renken RJ, van Dijk P. Sound-Evoked Neural Activity in Normal-Hearing Tinnitus: Effects of Frequency and Stimulated Ear Side. Brain Sciences. 2024; 14(6):544. https://doi.org/10.3390/brainsci14060544
Chicago/Turabian StyleSafazadeh, Shahin, Marc Thioux, Remco J. Renken, and Pim van Dijk. 2024. "Sound-Evoked Neural Activity in Normal-Hearing Tinnitus: Effects of Frequency and Stimulated Ear Side" Brain Sciences 14, no. 6: 544. https://doi.org/10.3390/brainsci14060544
APA StyleSafazadeh, S., Thioux, M., Renken, R. J., & van Dijk, P. (2024). Sound-Evoked Neural Activity in Normal-Hearing Tinnitus: Effects of Frequency and Stimulated Ear Side. Brain Sciences, 14(6), 544. https://doi.org/10.3390/brainsci14060544