
The Association among Hypothalamic Subnits, Gonadotropic and Sex Hormone Plasmas Levels in Alzheimer’s Disease

 , ,
, , 
Abstract
1. Introduction
2. Methods and Materials
2.1. Participants
2.2. Cognitive Status Groups
2.3. Image Acquisition and Preprocessing
2.4. Hypothalamic Segmentation
2.5. Volumetric Analysis
2.6. Hormonal Metrics
2.7. Statistical Analysis
3. Results
3.1. Demographic and Clinical Information
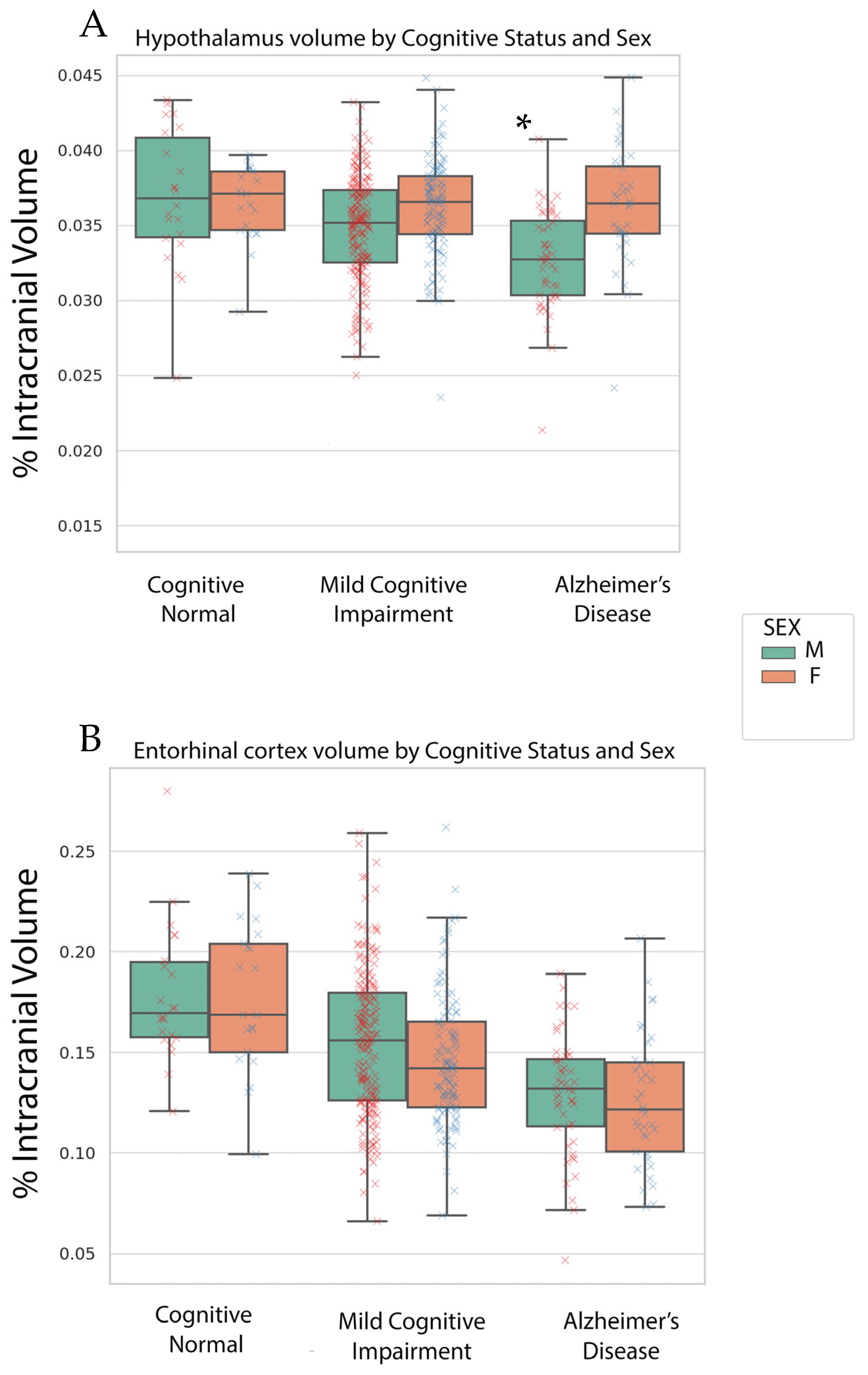
3.2. Volumetric Findings
3.3. Plasma Hormone Regression Analyses Findings
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sisk, C.L.; Zehr, J.L. Pubertal hormones organize the adolescent brain and behavior. Front. Neuroendocr. 2005, 26, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Vest, R.S.; Pike, C.J. Gender, sex steroid hormones, and Alzheimer’s disease. Horm. Behav. 2013, 63, 301–307. [Google Scholar] [CrossRef]
- Li, R.; Singh, M. Sex differences in cognitive impairment and Alzheimer’s disease. Front. Neuroendocr. 2014, 35, 385–403. [Google Scholar] [CrossRef]
- van der Velpen, I.F.; de Feijter, M.; Raina, R.; Özel, F.; Perry, M.; Ikram, M.A.; Vernooij, M.W.; Luik, A.I. Psychosocial health modifies associations between HPA-axis function and brain structure in older age. Psychoneuroendocrinology 2023, 153, 106106. [Google Scholar] [CrossRef]
- Van Drunen, R.; Eckel-Mahan, K. Circadian Rhythms of the Hypothalamus: From Function to Physiology. Clocks Sleep 2021, 3, 189–226. [Google Scholar] [CrossRef]
- Doorduijn, A.S.; de van der Schueren, M.; van de Rest, O.; de Leeuw, F.A.; Hendriksen, H.M.; Teunissen, C.E.; Scheltens, P.; van Der Flier, W.; Visser, M. Energy intake and expenditure in patients with Alzheimer’s disease and mild cognitive impairment: The NUDAD project. Alzheimer’s Dement. 2020, 16, e042429. [Google Scholar] [CrossRef]
- Billot, B.; Bocchetta, M.; Todd, E.; Dalca, A.V.; Rohrer, J.D.; Iglesias, J.E. Automated segmentation of the hypothalamus and associated subunits in brain MRI. NeuroImage 2020, 223, 117287. [Google Scholar] [CrossRef]
- Stewart, C.A.; Finger, E.C. The supraoptic and paraventricular nuclei in healthy aging and neurodegeneration. Handb. Clin. Neurol. 2021, 180, 105–123. [Google Scholar] [CrossRef]
- Verdile, G.; Laws, S.M.; Henley, D.; Ames, D.; Bush, A.I.; Ellis, K.A.; Faux, N.G.; Gupta, V.B.; Li, Q.X.; Masters, C.L.; et al. Associations between gonadotropins, testosterone and β amyloid in men at risk of Alzheimer’s disease. Mol. Psychiatry 2014, 19, 69–75. [Google Scholar] [CrossRef]
- Gurvich, C.; Le, J.; Thomas, N.; Thomas, E.H.X.; Kulkarni, J. Sex hormones and cognition in aging. Vitam. Horm. 2021, 115, 511–533. [Google Scholar] [CrossRef]
- Verdile, G.; Yeap, B.B.; Clarnette, R.M.; Dhaliwal, S.; Burkhardt, M.S.; Chubb, S.A.; De Ruyck, K.; Rodrigues, M.; Mehta, P.D.; Foster, J.K.; et al. Luteinizing hormone levels are positively correlated with plasma amyloid-beta protein levels in elderly men. J. Alzheimer’s Dis. JAD 2008, 14, 201–208. [Google Scholar] [CrossRef]
- Coley, N.; Andrieu, S.; Jaros, M.; Weiner, M.; Cedarbaum, J.; Vellas, B. Suitability of the Clinical Dementia Rating-Sum of Boxes as a single primary endpoint for Alzheimer’s disease trials. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2011, 7, 602–610.e602. [Google Scholar] [CrossRef]
- Morris, J.C. The Clinical Dementia Rating (CDR): Current version and scoring rules. Neurology 1993, 43, 2412–2414. [Google Scholar] [CrossRef]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Bernstein, M.A.; Fox, N.C.; Thompson, P.; Alexander, G.; Harvey, D.; Borowski, B.; Britson, P.J.; LWhitwell, J.; Ward, C.; et al. The Alzheimer’s Disease Neuroimaging Initiative (ADNI): MRI methods. J. Magn. Reson. Imaging 2008, 27, 685–691. [Google Scholar] [CrossRef]
- Ofori, E.; DeKosky, S.T.; Febo, M.; Colon-Perez, L.; Chakrabarty, P.; Duara, R.; Adjouadi, M.; Golde, T.E.; Vaillancourt, D.E. Free-water imaging of the hippocampus is a sensitive marker of Alzheimer’s disease. NeuroImage Clin. 2019, 24, 101985. [Google Scholar] [CrossRef]
- Kim, S.; Swaminathan, S.; Inlow, M.; Risacher, S.L.; Nho, K.; Shen, L.; Foroud, T.M.; Petersen, R.C.; Aisen, P.S.; Soares, H.; et al. Influence of genetic variation on plasma protein levels in older adults using a multi-analyte panel. PLoS ONE 2013, 8, e70269. [Google Scholar] [CrossRef]
- Mu, Y.; Gage, F.H. Adult hippocampal neurogenesis and its role in Alzheimer’s disease. Mol. Neurodegener. 2011, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.T.; Wang, W.E.; Zaborszky, L.; Golde, T.E.; DeKosky, S.; Duara, R.; Loewenstein, D.A.; Adjouadi, M.; Coombes, S.A.; Vaillancourt, D.E. Association of Cognitive Impairment With Free Water in the Nucleus Basalis of Meynert and Locus Coeruleus to Transentorhinal Cortex Tract. Neurology 2022, 98, e700–e710. [Google Scholar] [CrossRef] [PubMed]
- Oyola, M.G.; Handa, R.J. Hypothalamic-pituitary-adrenal and hypothalamic-pituitary-gonadal axes: Sex differences in regulation of stress responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef]
- McEwen, B.S.; Milner, T.A. Understanding the broad influence of sex hormones and sex differences in the brain. J. Neurosci. Res. 2017, 95, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Ebner, N.C.; Kamin, H.; Diaz, V.; Cohen, R.A.; MacDonald, K. Hormones as “difference makers” in cognitive and socioemotional aging processes. Front. Psychol. 2014, 5, 1595. [Google Scholar] [CrossRef]
- Popp, J.; Wolfsgruber, S.; Heuser, I.; Peters, O.; Hüll, M.; Schröder, J.; Möller, H.J.; Lewczuk, P.; Schneider, A.; Jahn, H.; et al. Cerebrospinal fluid cortisol and clinical disease progression in MCI and dementia of Alzheimer’s type. Neurobiol. Aging 2015, 36, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Adedeji, D.O.; Holleman, J.; Juster, R.P.; Udeh-Momoh, C.T.; Kåreholt, I.; Hagman, G.; Aspö, M.; Adagunodo, S.; Håkansson, K.; Kivipelto, M.; et al. Longitudinal study of Alzheimer’s disease biomarkers, allostatic load, and cognition among memory clinic patients. Brain Behav. Immun. Health 2023, 28, 100592. [Google Scholar] [CrossRef] [PubMed]
- Hiller, A.J.; Ishii, M. Disorders of Body Weight, Sleep and Circadian Rhythm as Manifestations of Hypothalamic Dysfunction in Alzheimer’s Disease. Front. Cell Neurosci. 2018, 12, 471. [Google Scholar] [CrossRef] [PubMed]
- Liguori, C. Orexin and Alzheimer’s Disease. Curr. Top. Behav. Neurosci. 2017, 33, 305–322. [Google Scholar] [CrossRef]
- Gegenhuber, B.; Tollkuhn, J. Sex Differences in the Epigenome: A Cause or Consequence of Sexual Differentiation of the Brain? Genes 2019, 10, 432. [Google Scholar] [CrossRef]
- Murta, V.; Ferrari, C. Peripheral Inflammation and Demyelinating Diseases. Adv. Exp. Med. Biol. 2016, 949, 263–285. [Google Scholar] [CrossRef]
- Starrett, J.R.; Moenter, S.M. Hypothalamic kisspeptin neurons as potential mediators of estradiol negative and positive feedback. Peptides 2023, 163, 170963. [Google Scholar] [CrossRef]

{kind=link}
| Group | N | Age (Years) | Sex (% Female) | Education (Years) | CDR-SB | MMSE | RAVLT | eTIV (Liters) |
|---|---|---|---|---|---|---|---|---|
| Cognitive Normal | 54 | 75.3 ± 5.9 | 28/26 (48%) | 15.6 ± 3 | 0.1 ± 0.3 | 29.1 ± 1.2 | 35.6 ± 34.9 | 1.51 ± 0.15 |
| Mild Cognitive Impairment | 343 | 74.9 ± 7.3 | 223/120 (35%) | 15.7 ± 3 | 2.2 ± 1.4 | 26.4 ± 2.9 | 67.6 ± 31.4 | 1.58 ± 0.15 |
| Alzheimer’s Disease | 96 | 75.1 ± 7.9 | 53/43 (45%) | 15.2 ± 3 | 5.6 ± 2.7 | 21.3 ± 4.6 | 85.2 ± 25.6 | 1.57 ± 0.20 |
| Totals & Statistics | - | F = 0.09, p = 0.92 | χ2 = 5.5, p = 0.06 | F = 1.2, p = 0.30 | F = 223.1, p < 0.001 | F = 132.5, p < 0.001 | F = 44.6, p < 0.001 | F = 3.0, p = 0.049 |
| Cognitive Normal | MCI | AD | |
|---|---|---|---|
| Follicle Stimulating Hormone | N/A | Anterior-superior (R2 = 0.05) | Inferior Tuberal (R2 = 0.11) |
| Luteinizing Hormone | Posterior Hypothalamus (R2 = 0.21) | N/A | Inferior Tuberal (R2 = 0.12) |
| Progesterone | Inferior Tuberal (R2 = 0.38) | Superior Tuberal (R2 = 0.08) | N/A |
| Testosterone | Inferior Tuberal (R2 = 0.20) | N/A | N/A |
| CN | MCI | AD | |
|---|---|---|---|
| Follicle Stimulating Hormone | N/A | Inferior Tuberal & Posterior Hypothalamus (R2 = 0.06) | N/A |
| Luteinizing Hormone | Posterior Hypothalamus (R2 = 0.21) | Posterior Hypothalamus (R2 = 0.06) | N/A |
| Progesterone | Inferior Tuberal (R2 = 0.18) | N/A | N/A |
| Testosterone | Superior Tuberal (R2 = 0.21) | Inferior Tuberal (R2 = 0.05) | N/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ofori, E.; Solis, A.; Punjani, N.; on behalf of the Alzheimer’s Disease Neuroimaging Initiative. The Association among Hypothalamic Subnits, Gonadotropic and Sex Hormone Plasmas Levels in Alzheimer’s Disease. Brain Sci. 2024, 14, 276. https://doi.org/10.3390/brainsci14030276
Ofori E, Solis A, Punjani N, on behalf of the Alzheimer’s Disease Neuroimaging Initiative. The Association among Hypothalamic Subnits, Gonadotropic and Sex Hormone Plasmas Levels in Alzheimer’s Disease. Brain Sciences. 2024; 14(3):276. https://doi.org/10.3390/brainsci14030276
Chicago/Turabian StyleOfori, Edward, Anamaria Solis, Nahid Punjani, and on behalf of the Alzheimer’s Disease Neuroimaging Initiative. 2024. "The Association among Hypothalamic Subnits, Gonadotropic and Sex Hormone Plasmas Levels in Alzheimer’s Disease" Brain Sciences 14, no. 3: 276. https://doi.org/10.3390/brainsci14030276
APA StyleOfori, E., Solis, A., Punjani, N., & on behalf of the Alzheimer’s Disease Neuroimaging Initiative. (2024). The Association among Hypothalamic Subnits, Gonadotropic and Sex Hormone Plasmas Levels in Alzheimer’s Disease. Brain Sciences, 14(3), 276. https://doi.org/10.3390/brainsci14030276