
Neural Pathways Linking Autonomous Exercise Motivation and Exercise-Induced Unhealthy Eating: A Resting-State fMRI Study


Abstract
1. Introduction
2. Methods
2.1. Participants and Procedures
2.2. Measures
2.2.1. Body Mass Index (BMI)
2.2.2. Autonomous Exercise Motivation
2.2.3. Exercise-Induced Unhealthy Foods Licensing (EUFL)
2.2.4. Image Acquisition
2.2.5. Image Data Preprocessing
2.3. Data Analysis
2.3.1. Neural–Behavior Correlation Analyses
2.3.2. Seed-to-Voxel rsFC Analysis
2.3.3. Mediation Model Analysis
3. Results
3.1. The Correlations between Behavioral Variables
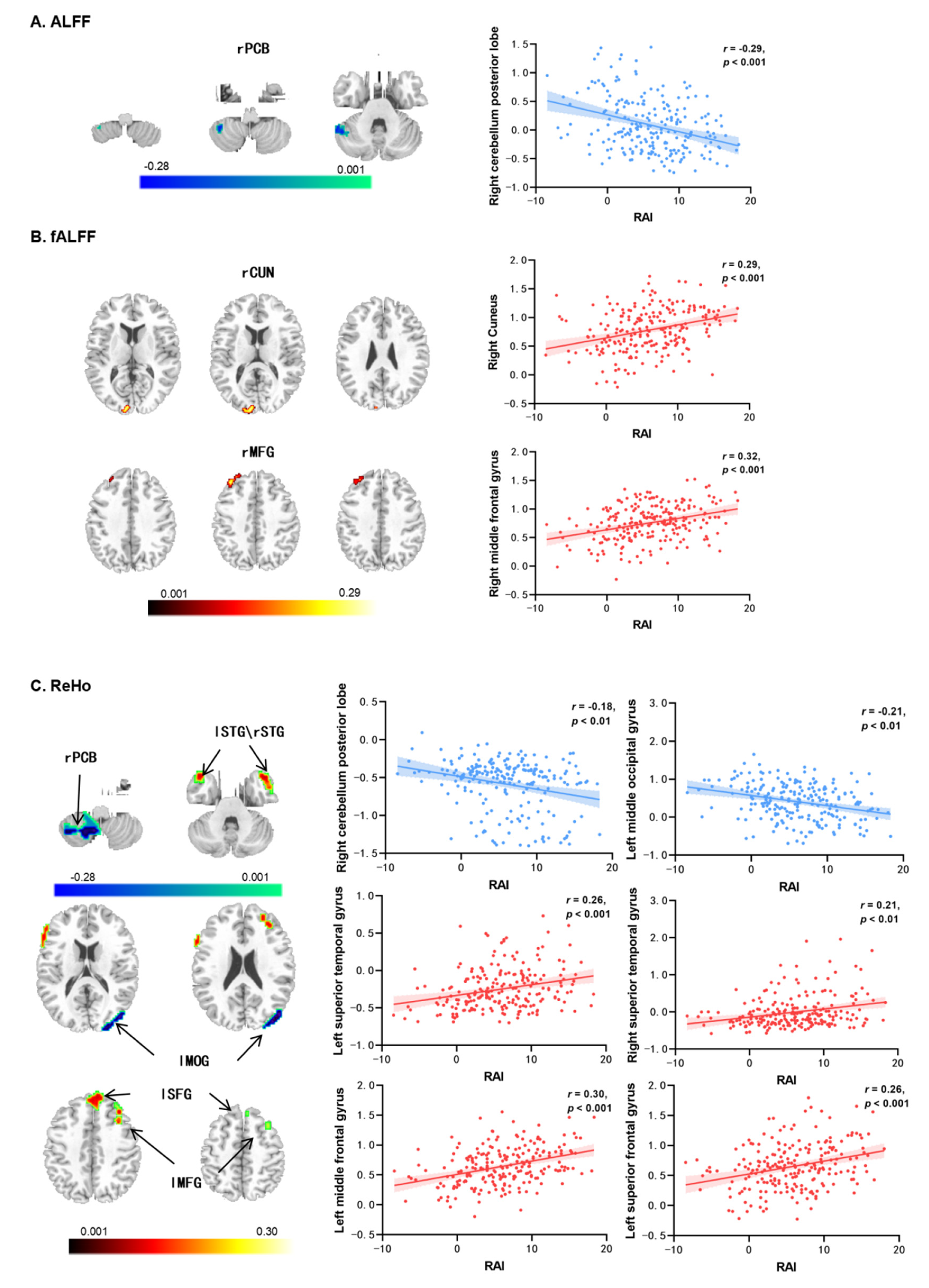
3.2. Autonomous Exercise Motivation-Related Resting-State Brain Activity
3.3. Autonomous Exercise Motivation-Related Functional Connectivity
3.4. Mediation Model Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Finlayson, G.; Caudwell, P.; Gibbons, C.; Hopkins, M.; King, N.; Blundell, J. Low Fat Loss Response after Medium-Term Supervised Exercise in Obese Is Associated with Exercise-Induced Increase in Food Reward. J. Obes. 2011, 2011, 615624. [Google Scholar] [CrossRef] [PubMed]
- Thivel, D.; Doucet, E.; Julian, V.; Cardenoux, C.; Boirie, Y.; Duclos, M. Nutritional compensation to exercise- vs. diet-induced acute energy deficit in adolescents with obesity. Physiol. Behav. 2017, 176, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, C.R.; Rakhmatullaeva, N.; Beckford, S.E.; Ammachathram, A.; Cristobal, A.; Koehler, K. Exercise and the timing of snack choice: Healthy snack choice is reduced in the post-exercise state. Nutrients 2018, 10, 1941. [Google Scholar] [CrossRef]
- Thomas, D.M.; Bouchard, C.; Church, T.; Slentz, C.; Kraus, W.E.; Redman, L.M.; Martin, C.K.; Silva, A.M.; Vossen, M.; Westerterp, K.; et al. Why do individuals not lose more weight from an exercise intervention at a defined dose? An energy balance analysis. Obes. Rev. 2012, 13, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Melanson, E.L.; Keadle, S.K.; Donnelly, J.E.; Donnelly, J.F.; Braun, B.; Braun, B.; King, N.A. Resistance to Exercise-Induced Weight Loss: Compensatory Behavioral Adaptations. Med. Sci. Sports Exerc. 2013, 45, 1600–1609. [Google Scholar] [CrossRef]
- Dimmock, J.A.; Guelfi, K.J.; West, J.S.; Masih, T.; Jackson, B. Does Motivation for Exercise Influence Post-Exercise Snacking Behavior? Nutrients 2015, 7, 4804–4816. [Google Scholar] [CrossRef] [PubMed]
- King, N.A.; Caudwell, P.; Hopkins, M.; Byrne, N.M.; Colley, R.; Hills, A.P.; Stubbs, J.R.; Blundell, J.E. Metabolic and behavioral compensatory responses to exercise interventions: Barriers to weight loss. Obesity 2007, 15, 1373–1383. [Google Scholar] [CrossRef] [PubMed]
- Standage, M.; Ryan, R.M. Self-Determination Theory in Sport and Exercise. In Handbook of Sport Psychology; Wiley: Hoboken, NJ, USA, 2020; pp. 37–56. [Google Scholar] [CrossRef]
- McCaig, D.C.; Hawkins, L.A.; Rogers, P.J. Licence to eat: Information on energy expended during exercise affects subsequent energy intake. Appetite 2016, 107, 323–329. [Google Scholar] [CrossRef]
- Miquelon, P.; Knäuper, B.; Vallerand, R.J. Motivation and goal attainment. The role of compensatory beliefs. Appetite 2012, 58, 608–615. [Google Scholar] [CrossRef]
- Beer, N.J.; Dimmock, J.A.; Jackson, B.; Guelfi, K.J. Exercise-related factors that influence post-exercise energy intake: A psychological perspective. J. Sci. Med. Sport 2020, 23, 1068–1073. [Google Scholar] [CrossRef]
- Deci, E.L.; Ryan, R.M. Self-Determination. In The Corsini Encyclopedia of Psychology; Wiley: Hoboken, NJ, USA, 2010; pp. 1–2. [Google Scholar] [CrossRef]
- Sweeney, A.M.; Freitas, A.L. Construing action abstractly and experiencing autonomy: Implications for physical activity and diet. Motiv. Emot. 2018, 42, 161–177. [Google Scholar] [CrossRef]
- Tylka, T.L.; Homan, K.J. Exercise motives and positive body image in physically active college women and men: Exploring an expanded acceptance model of intuitive eating. Body Image 2015, 15, 90–97. [Google Scholar] [CrossRef]
- Panão, I.; Carraça, E.V. Effects of exercise motivations on body image and eating habits/behaviours: A systematic review. Nutr. Diet. 2020, 77, 41–59. [Google Scholar] [CrossRef] [PubMed]
- West, J.; Guelfi, K.J.; Dimmock, J.A.; Jackson, B. “I deserve a treat”: Exercise motivation as a predictor of post-exercise dietary licensing beliefs and implicit associations toward unhealthy snacks. Psychol. Sport Exerc. 2017, 32, 93–101. [Google Scholar] [CrossRef]
- Beer, N.J.; Jackson, B.; Dimmock, J.A.; Guelfi, K.J. Attenuation of Post-Exercise Energy Intake Following 12 Weeks of Sprint Interval Training in Men and Women with Overweight. Nutrients 2022, 14, 1362. [Google Scholar] [CrossRef]
- Beer, N.J.; Dimmock, J.A.; Jackson, B.; Guelfi, K.J. Interactions of sprint interval exercise and psychological need-support on subsequent food intake among physically inactive men and women. Appl. Physiol. Nutr. Metab. 2020, 45, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Beer, N.J.; Dimmock, J.A.; Jackson, B.; Guelfi, K.J. Providing Choice in Exercise Influences Food Intake at the Subsequent Meal. Med. Sci. Sports Exerc. 2017, 49, 2110–2118. [Google Scholar] [CrossRef] [PubMed]
- Fenzl, N.; Bartsch, K.; Koenigstorfer, J. Labeling exercise fat-burning increases post-exercise food consumption in self-imposed exercisers. Appetite 2014, 81, 1–7. [Google Scholar] [CrossRef]
- Mata, J.; Silva, M.N.; Vieira, P.N.; Carraça, E.V.; Andrade, A.M.; Coutinho, S.R.; Sardinha, L.B.; Teixeira, P.J. Motivational “spill-over” during weight control: Increased self-determination and exercise intrinsic motivation predict eating self-regulation. Health Psychol. 2009, 28, 709. [Google Scholar] [CrossRef]
- Lowe, C.J.; Kolev, D.; Hall, P.A. An exploration of exercise-induced cognitive enhancement and transfer effects to dietary self-control. Brain Cogn. 2016, 110, 102–111. [Google Scholar] [CrossRef]
- Stein, A.T.; Greathouse, L.J.; Otto, M.W. Eating in response to exercise cues: Role of self-control fatigue, exercise habits, and eating restraint. Appetite 2016, 96, 56–61. [Google Scholar] [CrossRef]
- Legault, L.; Inzlicht, M. Self-determination, self-regulation, and the brain: Autonomy improves performance by enhancing neuroaffective responsiveness to self-regulation failure. J. Pers. Soc. Psychol. 2013, 105, 123. [Google Scholar] [CrossRef]
- Di Domenico, S.I.; Ryan, R.M. The Emerging Neuroscience of Intrinsic Motivation: A New Frontier in Self-Determination Research. Front. Hum. Neurosci. 2017, 11, 145. [Google Scholar] [CrossRef] [PubMed]
- Ng, B. The Neuroscience of Growth Mindset and Intrinsic Motivation. Brain Sci. 2018, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Good, D.J.; Li, M.; Deater-Deckard, K. A Genetic Basis for Motivated Exercise. Exerc. Sport Sci. Rev. 2015, 43, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Cheval, B.; Radel, R.; Neva, J.L.; Boyd, L.A.; Swinnen, S.P.; Sander, D.; Boisgontier, M.P. Behavioral and Neural Evidence of the Rewarding Value of Exercise Behaviors: A Systematic Review. Sports Med. 2018, 48, 1389–1404. [Google Scholar] [CrossRef] [PubMed]
- Sarah, D.; Sarah, M.S.; Kaitlyn, B.; Jenna, C.; Katie, P.; Abigail, L.; Colin, D.; Brady, T.; Daniel, C.; Oluwole, O.A.; et al. Evaluating the neural underpinnings of motivation for walking exercise. bioRxiv 2023, 2023, pzad159. [Google Scholar] [CrossRef]
- Simonsson, E.; Stiernman, L.J.; Lundquist, A.; Rosendahl, E.; Hedlund, M.; Lindelöf, N.; Boraxbekk, C.-J. Dopamine D2/3-receptor availability and its association with autonomous motivation to exercise in older adults: An exploratory [11C]-raclopride study. Front. Hum. Neurosci. 2022, 16, 997131. [Google Scholar] [CrossRef] [PubMed]
- Biswal, B.B.; Kylen, J.V.; Hyde, J.S. Simultaneous assessment of flow and BOLD signals in resting-state functional connectivity maps. NMR Biomed. 1997, 10, 165–170. [Google Scholar] [CrossRef]
- Biswal, B.B. Resting state fMRI: A personal history. NeuroImage 2012, 62, 938–944. [Google Scholar] [CrossRef]
- Smitha, K.A.; Akhil Raja, K.; Arun, K.M.; Rajesh, P.G.; Thomas, B.; Kapilamoorthy, T.R.; Kesavadas, C. Resting state fMRI: A review on methods in resting state connectivity analysis and resting state networks. Neuroradiol. J. 2017, 30, 305–317. [Google Scholar] [CrossRef]
- Markland, D.; Tobin, V. A modification to the behavioural regulation in exercise questionnaire to include an assessment of amotivation. J. Sport Exerc. Psychol. 2004, 26, 191–196. [Google Scholar] [CrossRef]
- Liu, J.D.; Chung, P.-K.; Zhang, C.-Q.; Si, G. Chinese-translated Behavioral Regulation in Exercise Questionnaire-2: Evidence from university students in the Mainland and Hong Kong of China. J. Sport Health Sci. 2015, 4, 228–234. [Google Scholar] [CrossRef]
- Chung, P.K.; Liu, J.D. Examination of the Psychometric Properties of the Chinese Translated Behavioral Regulation in Exercise Questionnaire-2. Meas. Phys. Educ. Exerc. 2012, 16, 300–315. [Google Scholar] [CrossRef]
- Goodboy, A.K.; Martin, M.M. Omega over alpha for reliability estimation of unidimensional communication measures. Ann. Int. Commun. Assoc. 2020, 44, 422–439. [Google Scholar] [CrossRef]
- Ling, Y.; Zheng, A.; Li, Q.; Huang, M.; Song, S.; Chen, H. Examination of an extended sociocultural model of lifestyle physical activity among men and women. Curr. Psychol. 2023, 42, 23531–23540. [Google Scholar] [CrossRef]
- West, J.S.; Guelfi, K.J.; Dimmock, J.A.; Jackson, B. Preliminary Validation of the Exercise-Snacking Licensing Scale: Rewarding Exercise with Unhealthy Snack Foods and Drinks. Nutrients 2018, 10, 1866. [Google Scholar] [CrossRef]
- Muschelli, J.; Nebel, M.B.; Caffo, B.S.; Barber, A.D.; Pekar, J.J.; Mostofsky, S.H. Reduction of motion-related artifacts in resting state fMRI using aCompCor. Neuroimage 2014, 96, 22–35. [Google Scholar] [CrossRef]
- Behzadi, Y.; Restom, K.; Liau, J.; Liu, T.T. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. Neuroimage 2007, 37, 90–101. [Google Scholar] [CrossRef]
- Satterthwaite, T.D.; Elliott, M.A.; Gerraty, R.T.; Ruparel, K.; Loughead, J.; Calkins, M.E.; Eickhoff, S.B.; Hakonarson, H.; Gur, R.C.; Gur, R.E. An improved framework for confound regression and filtering for control of motion artifact in the preprocessing of resting-state functional connectivity data. Neuroimage 2013, 64, 240–256. [Google Scholar] [CrossRef]
- Friston, K.J.; Williams, S.; Howard, R.; Frackowiak, R.S.J.; Turner, R. Movement-related effects in fMRI time-series. Magn. Reson. Med. 1996, 35, 346–355. [Google Scholar] [CrossRef]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 2012, 59, 2142–2154. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.-G.; Wang, X.-D.; Zuo, X.-N.; Zang, Y.-F. DPABI: Data processing & analysis for (resting-state) brain imaging. Neuroinformatics 2016, 14, 339–351. [Google Scholar] [PubMed]
- Zou, Q.-H.; Zhu, C.-Z.; Yang, Y.; Zuo, X.-N.; Long, X.-Y.; Cao, Q.-J.; Wang, Y.-F.; Zang, Y.-F. An improved approach to detection of amplitude of low-frequency fluctuation (ALFF) for resting-state fMRI: Fractional ALFF. J. Neurosci. Methods 2008, 172, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Jiang, T.; Lu, Y.; He, Y.; Tian, L. Regional homogeneity approach to fMRI data analysis. NeuroImage 2004, 22, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Song, X.-W.; Dong, Z.-Y.; Long, X.-Y.; Li, S.-F.; Zuo, X.-N.; Zhu, C.-Z.; He, Y.; Yan, C.-G.; Zang, Y.-F. REST: A toolkit for resting-state functional magnetic resonance imaging data processing. PLoS ONE 2011, 6, e25031. [Google Scholar] [CrossRef] [PubMed]
- Worsley, K.J.; Evans, A.C.; Marrett, S.; Neelin, P. A three-dimensional statistical analysis for CBF activation studies in human brain. J. Cereb. Blood Flow Metab. 1992, 12, 900–918. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.F. Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-Based Approach; Guilford Publications: New York, NY, USA, 2017. [Google Scholar]
- Pierce, J.E.; Thomasson, M.; Voruz, P.; Selosse, G.; Péron, J. Explicit and Implicit Emotion Processing in the Cerebellum: A Meta-analysis and Systematic Review. Cerebellum 2023, 22, 852–864. [Google Scholar] [CrossRef] [PubMed]
- Eekers, D.B.P.; in ‘t Ven, L.; Deprez, S.; Jacobi, L.; Roelofs, E.; Hoeben, A.; Lambin, P.; de Ruysscher, D.; Troost, E.G.C. The posterior cerebellum, a new organ at risk? Clin. Transl. Radiat. Oncol. 2018, 8, 22–26. [Google Scholar] [CrossRef]
- Belkhiria, C.; Driss, T.; Habas, C.; Jaafar, H.; Guillevin, R.; de Marco, G. Exploration and Identification of Cortico-Cerebellar-Brainstem Closed Loop During a Motivational-Motor Task: An fMRI Study. Cerebellum 2017, 16, 326–339. [Google Scholar] [CrossRef]
- Crémers, J.; Dessoullières, A.; Garraux, G. Hemispheric specialization during mental imagery of brisk walking. Hum. Brain Mapp. 2012, 33, 873–882. [Google Scholar] [CrossRef]
- Murayama, K.; Matsumoto, M.; Izuma, K.; Sugiura, A.; Ryan, R.M.; Deci, E.L.; Matsumoto, K. How Self-Determined Choice Facilitates Performance: A Key Role of the Ventromedial Prefrontal Cortex. Cereb. Cortex 2015, 25, 1241–1251. [Google Scholar] [CrossRef]
- Jörg, B.; Esther, A.; D’Esposito, M. Influence of Motivation on Control Hierarchy in the Human Frontal Cortex. J. Neurosci. 2015, 35, 3207. [Google Scholar] [CrossRef]
- Swart, J.C.; Frank, M.J.; Määttä, J.I.; Jensen, O.; Cools, R.; den Ouden, H.E.M. Frontal network dynamics reflect neurocomputational mechanisms for reducing maladaptive biases in motivated action. PLoS Biol. 2018, 16, e2005979. [Google Scholar] [CrossRef]
- Jimura, K.; Locke, H.S.; Braver, T.S. Prefrontal cortex mediation of cognitive enhancement in rewarding motivational contexts. Proc. Natl. Acad. Sci. USA 2010, 107, 8871–8876. [Google Scholar] [CrossRef]
- Rehman, A.; Al Khalili, Y. Neuroanatomy, Occipital Lobe; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Tu, S.; Qiu, J.; Martens, U.; Zhang, Q. Category-selective attention modulates unconscious processes in the middle occipital gyrus. Consci. Cogn. 2013, 22, 479–485. [Google Scholar] [CrossRef]
- Vanni, S.; Tanskanen, T.; Seppä, M.; Uutela, K.; Hari, R. Coinciding early activation of the human primary visual cortex and anteromedial cuneus. Proc. Natl. Acad. Sci. USA 2001, 98, 2776–2780. [Google Scholar] [CrossRef]
- Gottlieb, J.; Lopes, M.; Oudeyer, P.-Y. Motivated cognition: Neural and computational mechanisms of curiosity, attention, and intrinsic motivation. In Recent Developments in Neuroscience Research on Human Motivation; Emerald Group Publishing Limited: Bingley, UK, 2016; Volume 19, pp. 149–172. [Google Scholar] [CrossRef]
- Yao, L.; Li, W.; Dai, Z.; Dong, C. Eating behavior associated with gray matter volume alternations: A voxel based morphometry study. Appetite 2016, 96, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Chae, Y.; Lee, I.-S. Central Regulation of Eating Behaviors in Humans: Evidence from Functional Neuroimaging Studies. Nutrients 2023, 15, 3010. [Google Scholar] [CrossRef] [PubMed]
- Iosif, C.I.; Bashir, Z.I.; Apps, R.; Pickford, J. Cerebellar prediction and feeding behaviour. Cerebellum 2023, 22, 1002–1019. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Indices | Mean | SD | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|---|---|
| 1. Sex | - | - | - | |||||
| 2. Age | 18.85 | 0.94 | −0.01 | - | ||||
| 3. BMI | 21.13 | 3.30 | −0.16 * | 0.07 | - | |||
| 4. FD | 0.09 | 0.04 | −0.10 | −0.04 | 0.10 | - | ||
| 5. RAI | 5.99 | 5.28 | −0.23 ** | 0.02 | −0.01 | −0.01 | - | |
| 6. EUFL | 11.16 | 4.05 | 0.19 ** | −0.11 | −0.05 | −0.05 | −0.15 * | - |
| 7. EUDL | 11.50 | 4.39 | 0.11 | −0.14 * | −0.05 | −0.01 | −0.08 | 0.88 *** |
| Brain Regions | H | Peak MNI Coordinates | Peak T | Voxel Size | ||
|---|---|---|---|---|---|---|
| X | Y | Z | ||||
| Correlations with ALFF | ||||||
| Cerebellum posterior lobe (PCB) | R | 54 | −50 | −36 | −4.27 | 268 |
| Correlation with fALFF | ||||||
| Cuneus (CUN) | R | 10 | −96 | 12 | 3.95 | 167 |
| Middle frontal gyrus (MFG) | R | 38 | 34 | 44 | 4.43 | 82 |
| Correlation with ReHo | ||||||
| Cerebellum posterior lobe (PCB) | R | 9 | −60 | −54 | −3.79 | 141 |
| Superior temporal gyrus (STG) | L | −42 | 12 | −24 | 4.43 | 185 |
| Superior temporal gyrus (STG) | R | 42 | 18 | −36 | 3.95 | 320 |
| Middle occipital gyrus (MOG) | L | −45 | −81 | 0 | −4.27 | 218 |
| Middle frontal gyrus (MFG) | L | −36 | 18 | 45 | 4.76 | 114 |
| Superior frontal gyrus (SFG) | L | −6 | 48 | 36 | 4.60 | 99 |
| Seed-based Functional Connectivity | ||||||
| Seed: PCB_R (ALFF) | ||||||
| Middle occipital gyrus (MOG) | R | 22 | −88 | −16 | 3.76 | 813 |
| Seed: PCB_R (ReHo) | ||||||
| Inferior frontal gyrus (IFG) | R | 54 | 6 | 26 | 3.63 | 181 |
| Supramarginal Gyrus (SMG) | L | −58 | −56 | 30 | −4.07 | 199 |
| Postcentral gyrus (PoG) | R | −62 | −14 | 36 | 3.85 | 195 |
| Superior frontal gyrus (SFG) | R | 20 | 18 | 68 | −4.17 | 183 |
| Seed: MOG_L (ReHo) | ||||||
| Cuneus (CUN) | R | 2 | −86 | 36 | 5.36 | 3813 |
| Seed: MFG_L (ReHo) | ||||||
| Cerebellum posterior lobe (PCB) | L | −4 | −52 | −52 | −4.85 | 282 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, Y.; Han, J.; Cui, Y.; Li, W.; Chen, H. Neural Pathways Linking Autonomous Exercise Motivation and Exercise-Induced Unhealthy Eating: A Resting-State fMRI Study. Brain Sci. 2024, 14, 221. https://doi.org/10.3390/brainsci14030221
Ling Y, Han J, Cui Y, Li W, Chen H. Neural Pathways Linking Autonomous Exercise Motivation and Exercise-Induced Unhealthy Eating: A Resting-State fMRI Study. Brain Sciences. 2024; 14(3):221. https://doi.org/10.3390/brainsci14030221
Chicago/Turabian StyleLing, Ying, Jinfeng Han, Yicen Cui, Wei Li, and Hong Chen. 2024. "Neural Pathways Linking Autonomous Exercise Motivation and Exercise-Induced Unhealthy Eating: A Resting-State fMRI Study" Brain Sciences 14, no. 3: 221. https://doi.org/10.3390/brainsci14030221
APA StyleLing, Y., Han, J., Cui, Y., Li, W., & Chen, H. (2024). Neural Pathways Linking Autonomous Exercise Motivation and Exercise-Induced Unhealthy Eating: A Resting-State fMRI Study. Brain Sciences, 14(3), 221. https://doi.org/10.3390/brainsci14030221