
Cross-Modal Tinnitus Remediation: A Tentative Theoretical Framework



Abstract
1. Introduction
1.1. Background
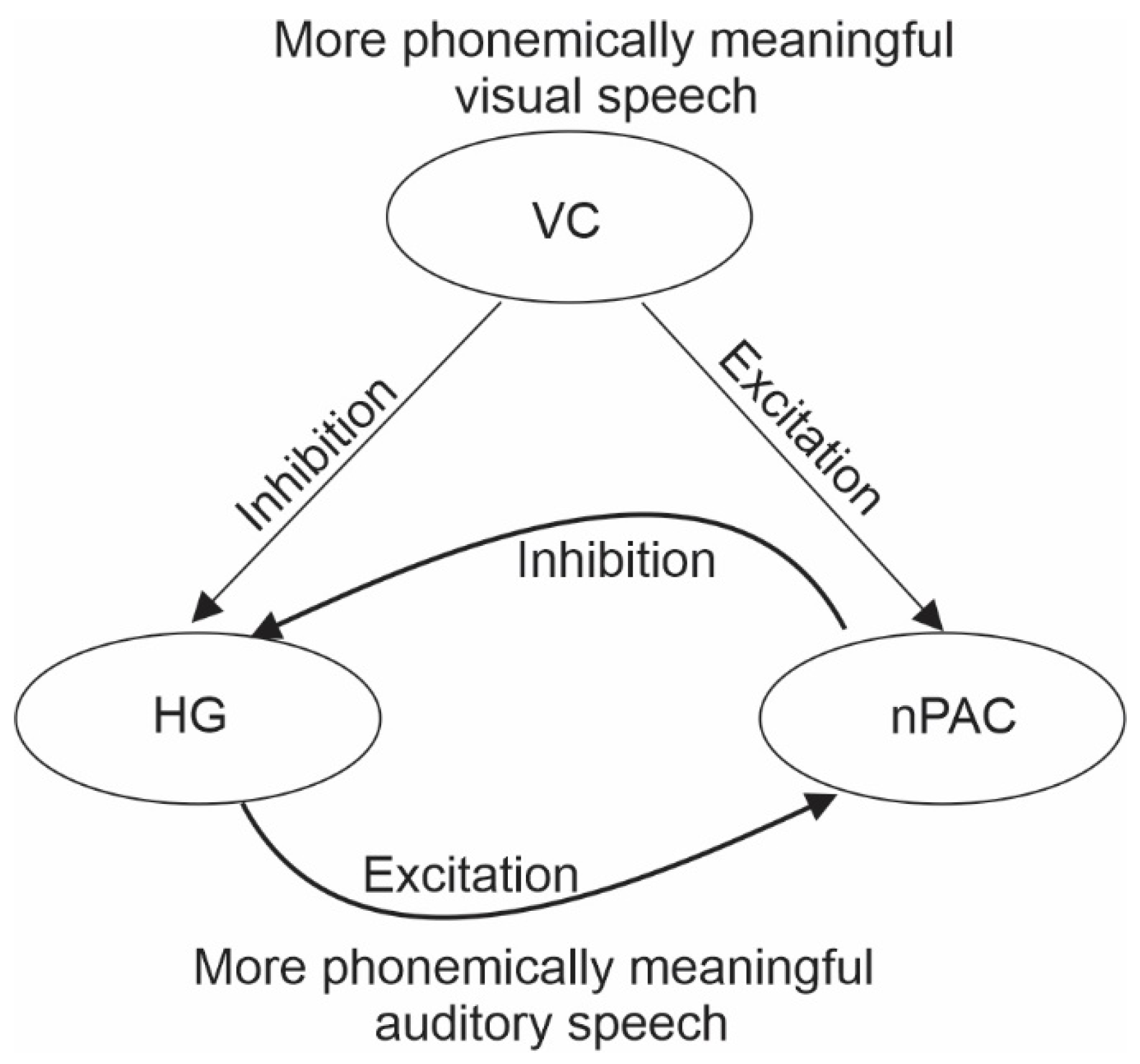
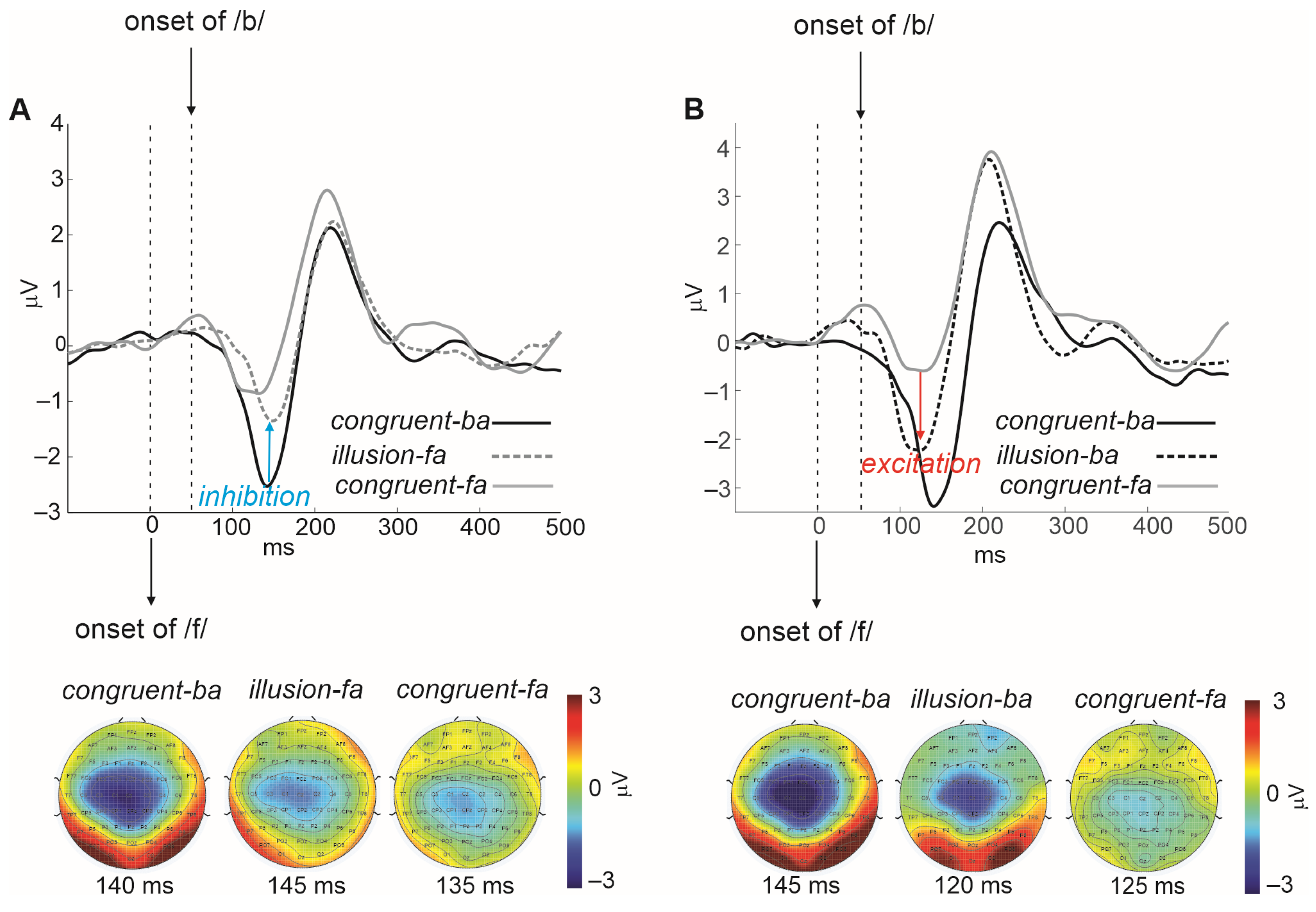
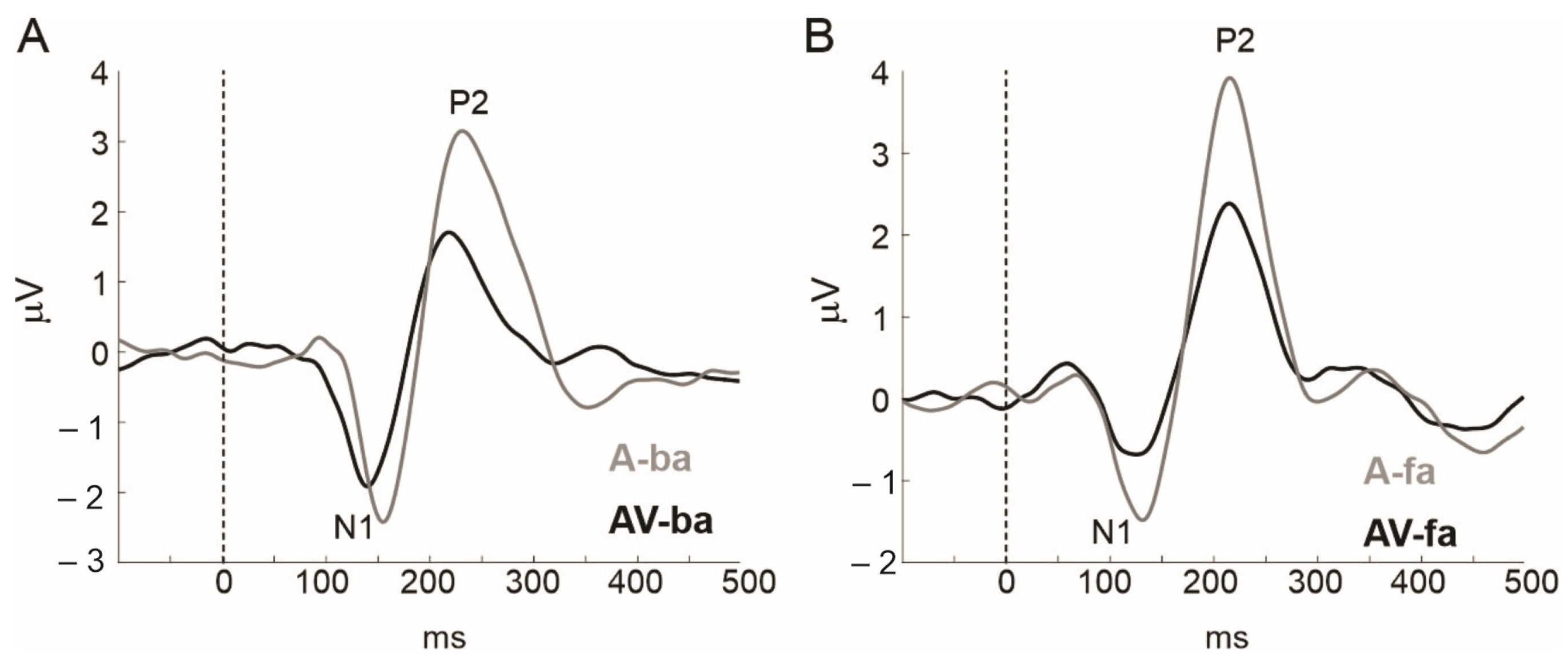
1.2. The Primary Visual Influence on the Auditory Cortex Is Inhibitory
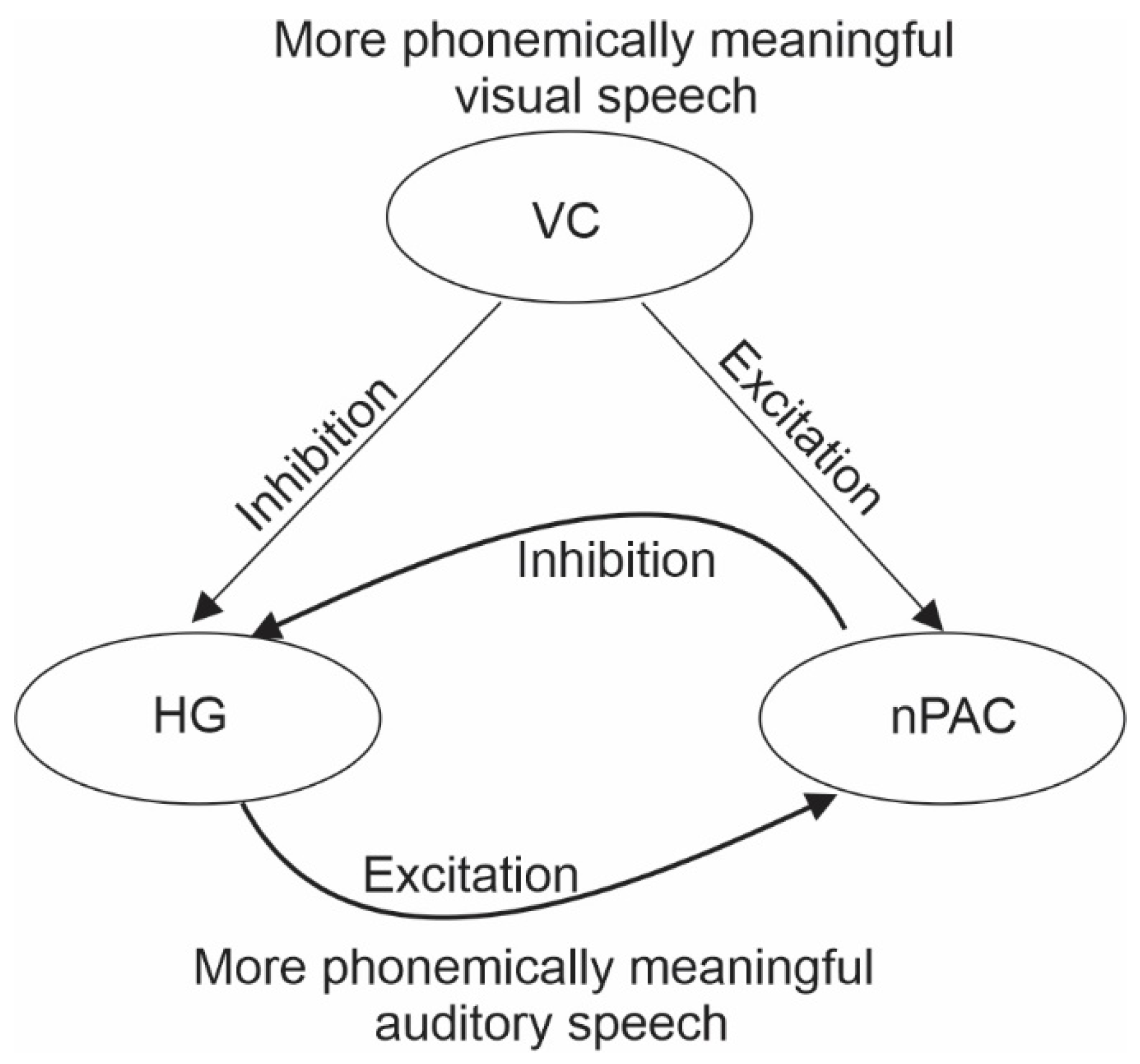
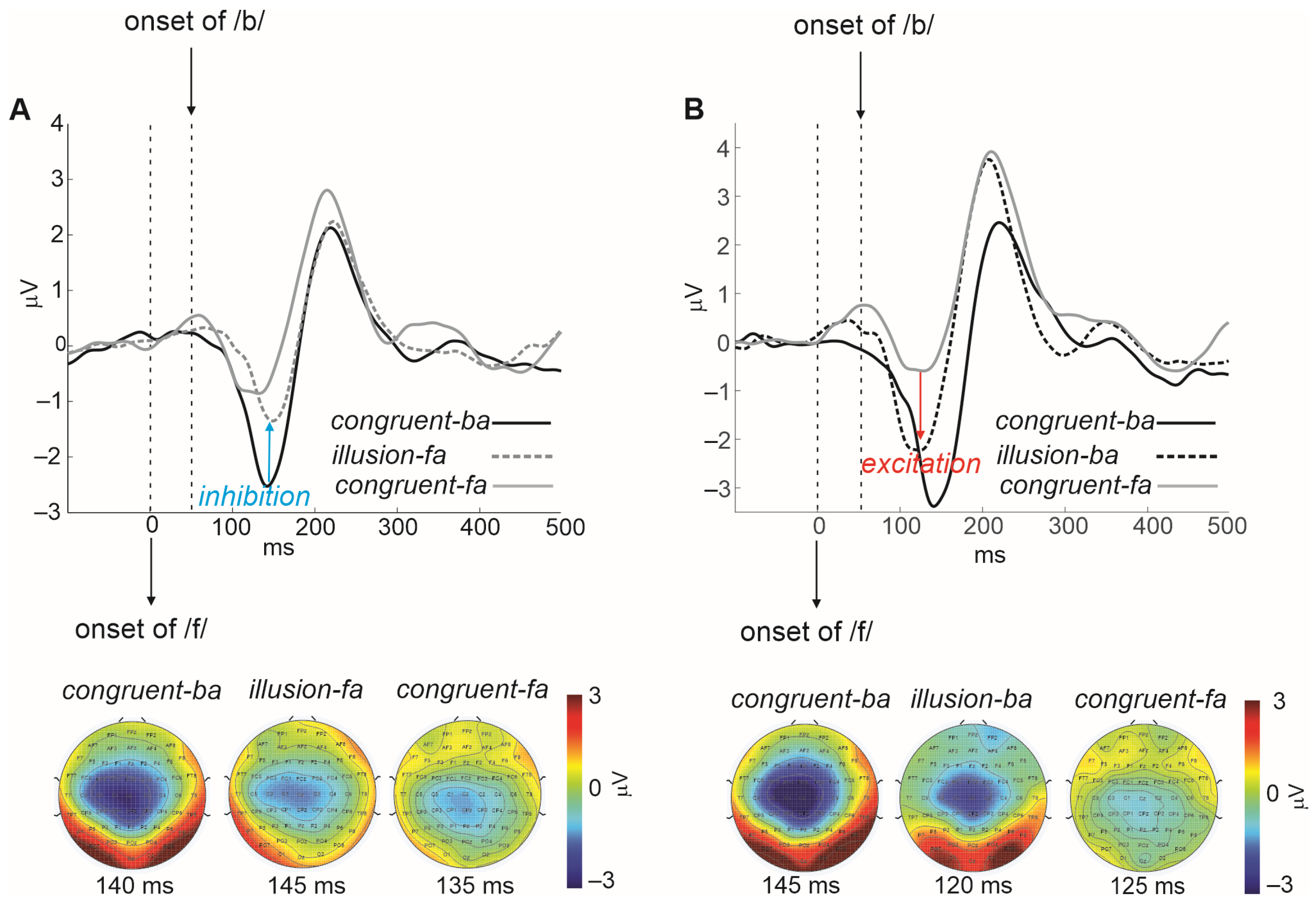
1.3. The Secondary Visual Influence on Auditory Representations Is Modulatory
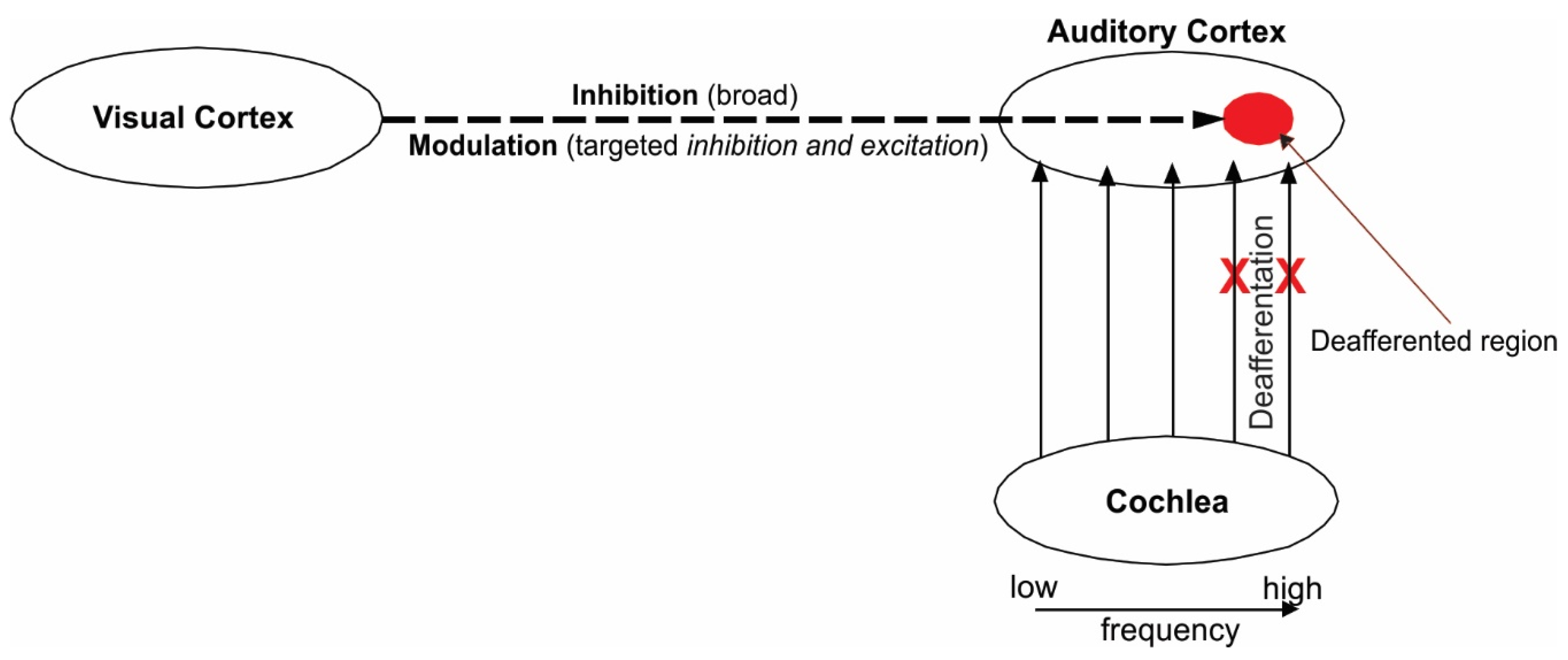
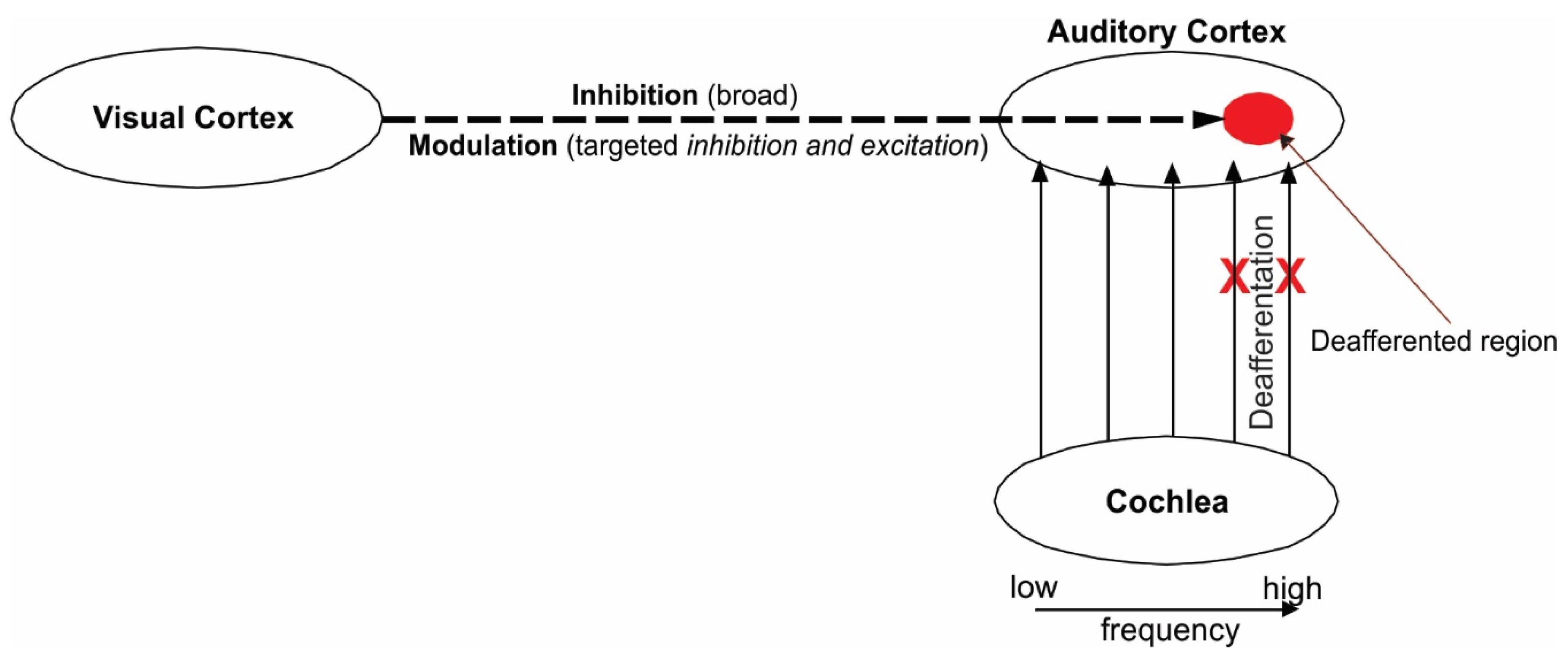
2. Theoretical Framework
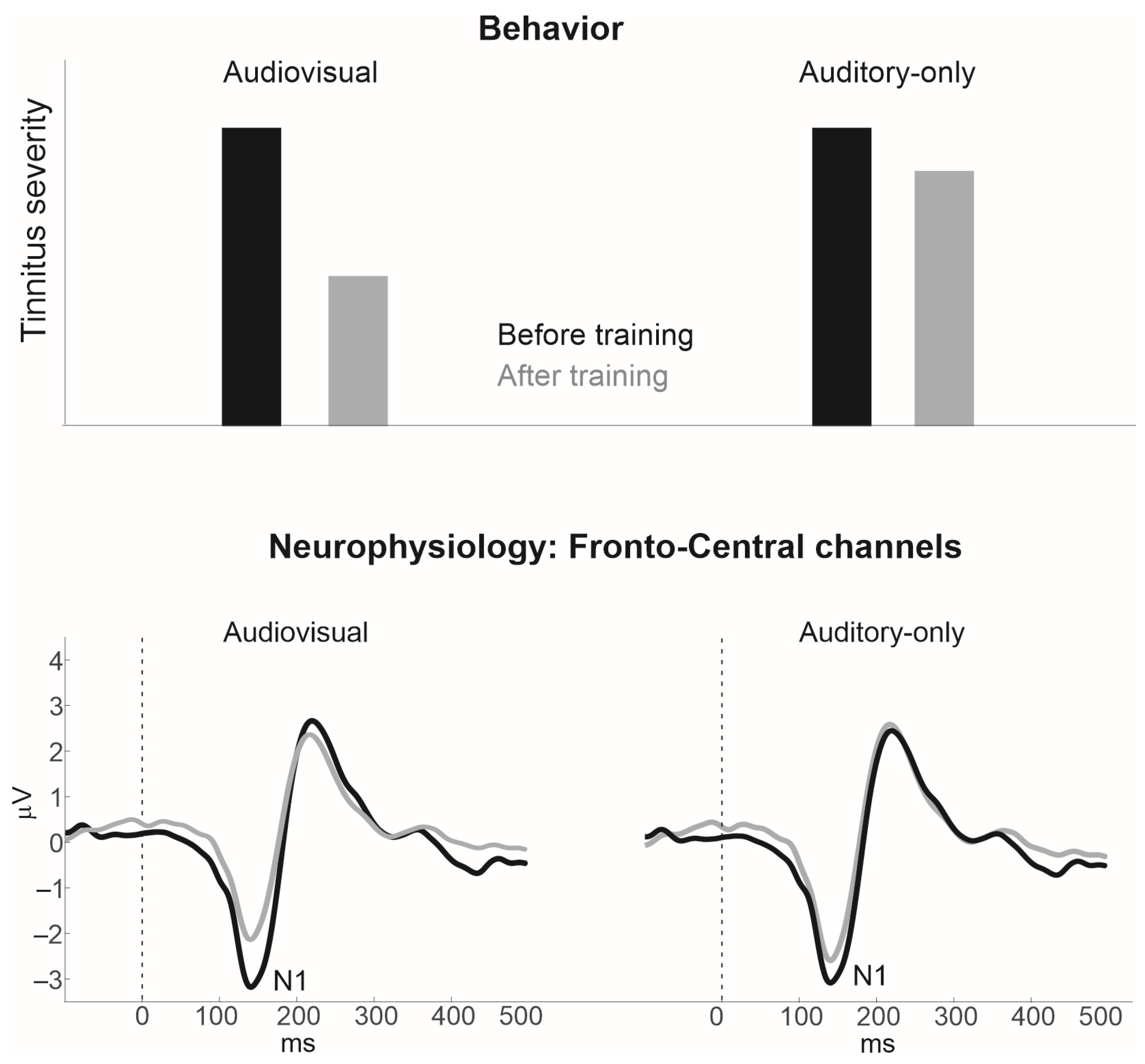
3. Audiovisual Training as a Means to Remedy Tinnitus
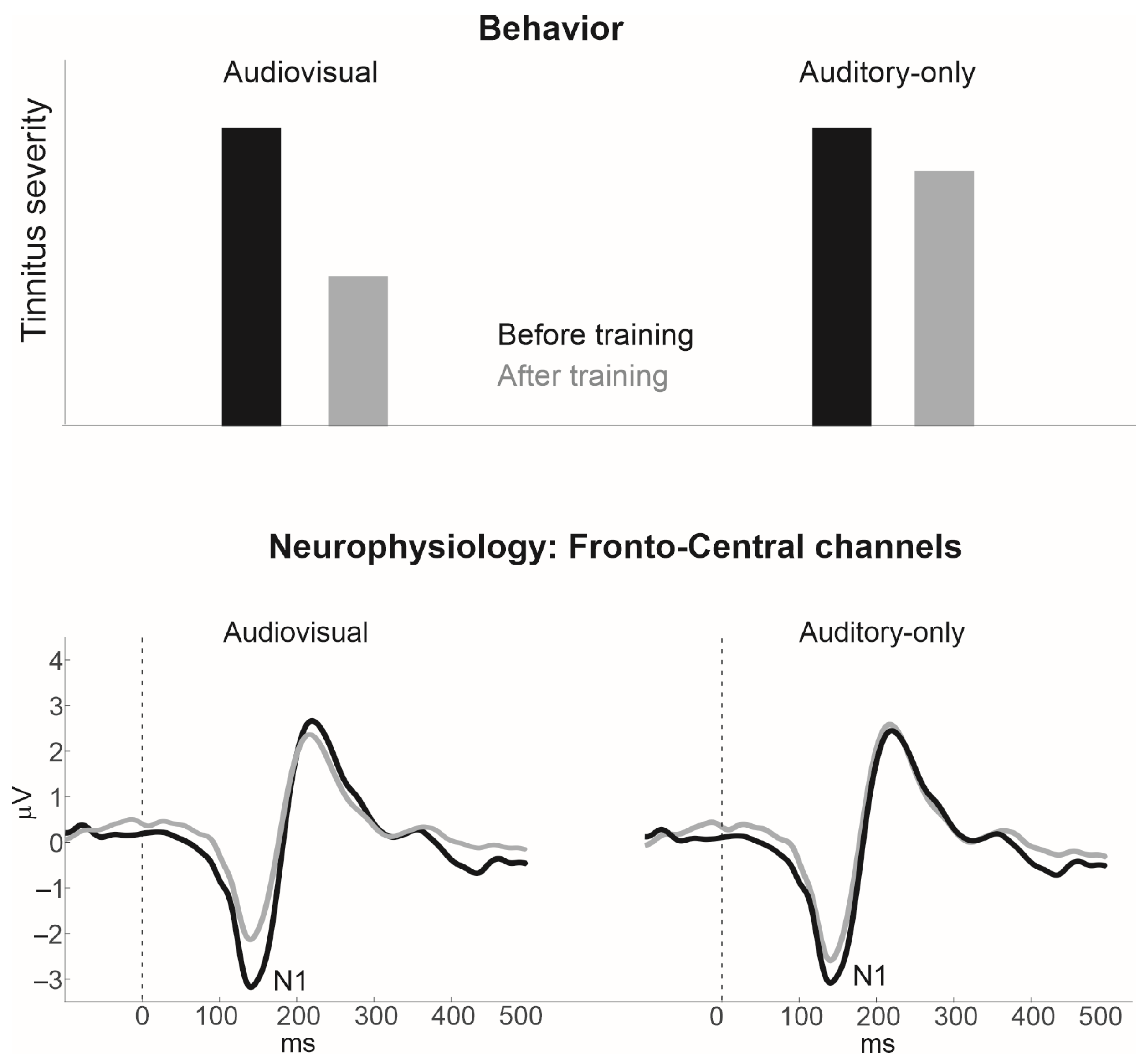
3.1. Audiovisual Filling-In of Notched Speech
3.2. Potential Limitations
3.3. Alternate Approaches, Moving beyond the N1 AEP
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Galazyuk, A.V.; Wenstrup, J.J.; Hamid, M.A. Tinnitus and underlying brain mechanisms. Curr. Opin. Otolaryngol. Head Neck Surg. 2012, 20, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Malouff, J.M.; Schutte, N.S.; Zucker, L.A. Tinnitus-related distress: A review of recent findings. Curr. Psychiatry Rep. 2011, 13, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Henry, J.A.; Roberts, L.E.; Caspary, D.M.; Theodoroff, S.M.; Salvi, R.J. Underlying Mechanisms of Tinnitus: Review and Clinical Implications. J. Am. Acad. Audiol. 2014, 25, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Langguth, B.; Kreuzer, P.M.; Kleinjung, T.; De Ridder, D. Tinnitus: Causes and clinical management. Lancet Neurol. 2013, 12, 920–930. [Google Scholar] [CrossRef]
- Tinnitus. Available online: https://www.nidcd.nih.gov/health/tinnitus#2 (accessed on 10 November 2023).
- Cederroth, C.R.; Gallus, S.; Hall, D.A.; Kleinjung, T.; Langguth, B.; Maruotti, A.; Meyer, M.; Norena, A.; Probst, T.; Pryss, R.; et al. Editorial: Towards an Understanding of Tinnitus Heterogeneity. Front. Aging Neurosci. 2019, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, A.; Ringdahl, A. Tinnitus—A study of its prevalence and characteristics. Br. J. Audiol. 1989, 23, 53–62. [Google Scholar] [CrossRef]
- Davis, A.; El Refaie, A. Epidemiology of tinnitus. In Tinnitus Handbook; Tyler, R., Ed.; Singular (Thomson Learning): San Diego, CA, USA, 2000; pp. 1–23. [Google Scholar]
- Henry, J.L.; Kangas, M.; Wilson, P.H. Development of the psychological impact of tinnitus interview: A clinician-administered measure of tinnitus-related distress. Int. Tinnitus J. 2001, 7, 20–26. [Google Scholar]
- Roberts, L.E.; Eggermont, J.J.; Caspary, D.M.; Shore, S.E.; Melcher, J.R.; Kaltenbach, J.A. Ringing ears: The neuroscience of tinnitus. J. Neurosci. 2010, 30, 14972–14979. [Google Scholar] [CrossRef]
- Han, B.I.; Lee, H.W.; Kim, T.Y.; Lim, J.S.; Shin, K.S. Tinnitus: Characteristics, Causes, Mechanisms, and Treatments. J. Clin. Neurol. 2009, 5, 11–19. [Google Scholar] [CrossRef]
- Reed, G.F. An audiometric study of two hundred cases of subjective tinnitus. AMA Arch. Otolaryngol. 1960, 71, 84–94. [Google Scholar] [CrossRef]
- Eggermont, J.J.; Roberts, L.E. The neuroscience of tinnitus. Trends Neurosci. 2004, 27, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Jastreboff, P.J.; Brennan, J.F.; Coleman, J.K.; Sasaki, C.T. Phantom auditory sensation in rats: An animal model for tinnitus. Behav. Neurosci. 1988, 102, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Longenecker, R.J.; Galazyuk, A.V. Development of tinnitus in CBA/CaJ mice following sound exposure. J. Assoc. Res. Otolaryngol. 2011, 12, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.J.; Beck, J.M.; Latham, P.E.; Pouget, A. Bayesian inference with probabilistic population codes. Nat. Neurosci. 2006, 9, 1432–1438. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, N.F.; Licari, F.G.; Klapchar, M.; Elkin, R.L.; Gao, Y.; Chen, G.; Kaltenbach, J.A. Noise-induced hyperactivity in the inferior colliculus: Its relationship with hyperactivity in the dorsal cochlear nucleus. J. Neurophysiol. 2012, 108, 976–988. [Google Scholar] [CrossRef]
- Mulders, W.H.A.M.; Robertson, D. Hyperactivity in the auditory midbrain after acoustic trauma: Dependence on cochlear activity. Neuroscience 2009, 164, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Mulders, W.H.A.M.; Seluakumaran, K.; Robertson, D. Efferent Pathways Modulate Hyperactivity in Inferior Colliculus. J. Neurosci. 2010, 30, 9578–9587. [Google Scholar] [CrossRef]
- Marschall, T.M.; Brederoo, S.G.; Ćurčić-Blake, B.; Sommer, I.E. Deafferentation as a cause of hallucinations. Curr. Opin. Psychiatry 2020, 33, 206–211. [Google Scholar] [CrossRef]
- Brozoski, T.J.; Bauer, C.A.; Caspary, D.M. Elevated Fusiform Cell Activity in the Dorsal Cochlear Nucleus of Chinchillas with Psychophysical Evidence of Tinnitus. J. Neurosci. 2002, 22, 2383–2390. [Google Scholar] [CrossRef]
- Chambers, A.R.; Resnik, J.; Yuan, Y.; Whitton, J.P.; Edge, A.S.; Liberman, M.C.; Polley, D.B. Central Gain Restores Auditory Processing following Near-Complete Cochlear Denervation. Neuron 2016, 89, 867–879. [Google Scholar] [CrossRef]
- Rajan, R. Receptor organ damage causes loss of cortical surround inhibition without topographic map plasticity. Nat. Neurosci. 1998, 1, 138–143. [Google Scholar] [CrossRef]
- Thai-Van, H.; Micheyl, C.; Moore, B.C.J.; Collet, L. Enhanced frequency discrimination near the hearing loss cut-off: A consequence of central auditory plasticity induced by cochlear damage? Brain 2003, 126, 2235–2245. [Google Scholar] [CrossRef]
- Wu, P.; Liu, H.; Li, X.; Fan, T.; Zhang, X. A Novel Lip Descriptor for Audio-Visual Keyword Spotting Based on Adaptive Decision Fusion. IEEE Trans. Multimed. 2016, 18, 326–338. [Google Scholar] [CrossRef]
- Eggermont, J.J.; Komiya, H. Moderate noise trauma in juvenile cats results in profound cortical topographic map changes in adulthood. Hear. Res. 2000, 142, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Rajan, R.; Irvine, D.R. Neuronal responses across cortical field A1 in plasticity induced by peripheral auditory organ damage. Audiol. Neurootol. 1998, 3, 123–144. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, V.; Nieschalk, M.; Stoll, W.; Rajan, R.; Pantev, C. Cortical reorganization in patients with high frequency cochlear hearing loss. Hear. Res. 2001, 158, 95–101. [Google Scholar] [CrossRef]
- Hoke, M.; Feldmann, H.; Pantev, C.; Lütkenhöner, B.; Lehnertz, K. Objective evidence of tinnitus in auditory evoked magnetic fields. Hear. Res. 1989, 37, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, G.P.; Calder, J.A.; Newman, C.W.; Peterson, E.L.; Wharton, J.A.; Ahmad, B. Electrophysiological indices of selective auditory attention in subjects with and without tinnitus. Hear. Res. 1996, 97, 66–74. [Google Scholar] [CrossRef]
- Guiraud, J.; Besle, J.; Arnold, L.; Boyle, P.; Giard, M.-H.; Bertrand, O.; Norena, A.; Truy, E.; Collet, L. Evidence of a tonotopic organization of the auditory cortex in cochlear implant users. J. Neurosci. 2007, 27, 7838–7846. [Google Scholar] [CrossRef]
- Pantev, C.; Roberts, L.; Elbert, T.; Roβ, B.; Wienbruch, C. Tonotopic organization of the sources of human auditory steady-state responses. Hear. Res. 1996, 101, 62–74. [Google Scholar] [CrossRef]
- Pantev, C.; Hoke, M.; Lütkenhöner, B.; Lehnertz, K. Tonotopic organization of the auditory cortex: Pitch versus frequency representation. Science 1989, 246, 486–488. [Google Scholar] [CrossRef] [PubMed]
- de Azevedo, A.A.; Figueiredo, R.R.; Penido, N.D.O. Tinnitus and event related potentials: A systematic review. Braz. J. Otorhinolaryngol. 2020, 86, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Sereda, M.; Adjamian, P.; Edmondson-Jones, M.; Palmer, A.R.; Hall, D.A. Auditory evoked magnetic fields in individuals with tinnitus. Hear. Res. 2013, 302, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Scherg, M.; Vajsar, J.; Picton, T.W. A source analysis of the late human auditory evoked potentials. J. Cogn. Neurosci. 1989, 1, 336–355. [Google Scholar] [CrossRef]
- Brozoski, T.J.; Ciobanu, L.; Bauer, C.A. Central neural activity in rats with tinnitus evaluated with manganese-enhanced magnetic resonance imaging (MEMRI). Hear. Res. 2007, 228, 168–179. [Google Scholar] [CrossRef]
- Wang, H.; Brozoski, T.J.; Caspary, D.M. Inhibitory neurotransmission in animal models of tinnitus: Maladaptive plasticity. Hear. Res. 2011, 279, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Stracke, H.; Stoll, W.; Pantev, C. Listening to tailor-made notched music reduces tinnitus loudness and tinnitus-related auditory cortex activity. Proc. Natl. Acad. Sci. USA 2010, 107, 1207–1210. [Google Scholar] [CrossRef] [PubMed]
- Pantev, C.; Okamoto, H.; Teismann, H. Music-induced cortical plasticity and lateral inhibition in the human auditory cortex as foundations for tonal tinnitus treatment. Front. Syst. Neurosci. 2012, 6, 50. [Google Scholar] [CrossRef]
- Marks, K.L.; Martel, D.T.; Wu, C.; Basura, G.J.; Roberts, L.E.; Schvartz-Leyzac, K.C.; Shore, S.E. Auditory-somatosensory bimodal stimulation desynchronizes brain circuitry to reduce tinnitus in guinea pigs and humans. Sci. Transl. Med. 2018, 10, eaal3175. [Google Scholar] [CrossRef]
- Spiegel, D.P.; Linford, T.; Thompson, B.; Petoe, M.A.; Kobayashi, K.; Stinear, C.M.; Searchfield, G.D. Multisensory attention training for treatment of tinnitus. Sci. Rep. 2015, 5, 10802. [Google Scholar] [CrossRef]
- Baart, M.; Vroomen, J.; Shaw, K.; Bortfeld, H. Degrading phonetic information affects matching of audiovisual speech in adults, but not in infants. Cognition 2014, 130, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Besle, J.; Fort, A.; Delpuech, C.; Giard, M. Bimodal speech: Early suppressive visual effects in human auditory cortex. Eur. J. Neurosci. 2004, 20, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Karas, P.J.; Magnotti, J.F.; Metzger, B.A.; Zhu, L.L.; Smith, K.B.; Yoshor, D.; Beauchamp, M.S. The visual speech head start improves perception and reduces superior temporal cortex responses to auditory speech. eLife 2019, 8, e48116. [Google Scholar] [CrossRef] [PubMed]
- Pilling, M. Auditory event-related potentials (ERPs) in audio-visual speech perception. J. Speech Lang. Hear. Res. 2009, 52, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Stekelenburg, J.J.; Vroomen, J. Neural correlates of multisensory integration of ecologically valid audiovisual events. J. Cogn. Neurosci. 2007, 19, 1964–1973. [Google Scholar] [CrossRef] [PubMed]
- van Wassenhove, V.; Grant, K.W.; Poeppel, D. Visual speech speeds up the neural processing of auditory speech. Biol. Sci. 2005, 102, 1181–1186. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.; Miller, L.M.; Pitt, M.A.; Shahin, A.J. Putative mechanisms mediating tolerance for audiovisual stimulus onset asynchrony. J. Neurophysiol. 2015, 113, 1437–1450. [Google Scholar] [CrossRef]
- Shahin, A.J.; Backer, K.C.; Rosenblum, L.D.; Kerlin, J.R. Neural mechanisms underlying cross-modal phonetic encoding. J. Neurosci. 2018, 38, 1835–1849. [Google Scholar] [CrossRef]
- Shatzer, H.; Shen, S.; Kerlin, J.R.; Pitt, M.A.; Shahin, A.J. Neurophysiology underlying influence of stimulus reliability on audiovisual integration. Eur. J. Neurosci. 2018, 48, 2836–2848. [Google Scholar] [CrossRef]
- Stekelenburg, J.J.; Vroomen, J. Electrophysiological evidence for a multisensory speech-specific mode of perception. Neuropsychologia 2012, 50, 1425–1431. [Google Scholar] [CrossRef]
- Fujisaki, W.; Shimojo, S.; Kashino, M.; Nishida, S. Recalibration of Audiovisual Simultaneity. Nat. Neurosci. 2004, 7, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Harrar, V.; Harris, L.R. The effect of exposure to asynchronous audio, visual, and tactile stimulus combinations on the perception of simultaneity. Exp. Brain Res. 2008, 186, 517–524. [Google Scholar] [CrossRef]
- Navarra, J.; Vatakis, A.; Zampini, M.; Soto-Faraco, S.; Humphreys, W.; Spence, C. Exposure to asynchronous audiovisual speech extends the temporal window for audiovisual integration. Cogn. Brain Res. 2005, 25, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.G.; Tang, Q.; Dimitrijevic, A.; Starr, A.; Larky, J.; Blevins, N.H. Tinnitus suppression by low-rate electric stimulation and its electrophysiological mechanisms. Hear. Res. 2011, 277, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, B.; Hornickel, J.; Skoe, E.; Nicol, T.; Kraus, N. Context-dependent encoding in the human auditory brainstem relates to hearing speech in noise: Implications for developmental dyslexia. Neuron 2009, 64, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.L.; Savariaux, C. No, there is no 150 ms lead of visual speech on auditory speech, but a range of audiovisual asynchronies varying from small audio lead to large audio lag. PLoS Comput. Biol. 2014, 10, e1003743. [Google Scholar] [CrossRef]
- Abbott, N.M.; Shahin, A.J. Cross-modal phonetic encoding facilitates the McGurk illusion and phonemic restoration. J. Neurophysiol. 2018, 120, 2988–3000. [Google Scholar] [CrossRef]
- Warren, R.M.; Obusek, C.J. Speech perception and phonemic restorations. Percept. Psychophys. 1971, 9, 358–362. [Google Scholar] [CrossRef]
- Samuel, A.G. Phonemic restoration: Insights from a new methodology. J. Exp. Psychol. Gen. 1981, 110, 474–494. [Google Scholar] [CrossRef]
- Shahin, A.J.; Miller, L.M. Multisensory integration enhances phonemic restoration. J. Acoust. Soc. Am. 2009, 125, 1744–1750. [Google Scholar] [CrossRef]
- Shahin, A.J.; Bishop, C.W.; Miller, L.M. Neural mechanisms for illusory filling-in of degraded speech. Neuroimage 2009, 44, 1133–1143. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.; Avery, T.; Turcios, J.; Brancazio, L.; Cook, B.; Landi, N. Electrophysiological Indices of Audiovisual Speech Perception in the Broader Autism Phenotype. Brain Sci. 2017, 7, 60. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.; Avery, T.; Kleinman, D.; Landi, N. Audiovisual Speech Perception in Children with Autism Spectrum Disorders: Evidence from Visual Phonemic Restoration. J. Autism Dev. Disord. 2022, 52, 28–37. [Google Scholar] [CrossRef]
- Jensen, O.; Gelfand, J.; Kounios, J.; Lisman, J.E. Oscillations in the alpha band (9–12 Hz) increase with memory load during retention in a short-term memory task. Cereb. Cortex 2002, 12, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Jensen, O.; Mazaheri, A. Shaping functional architecture by oscillatory alpha activity: Gating by inhibition. Front. Hum. Neurosci. 2010, 4, 186. [Google Scholar] [CrossRef]
- Obleser, J.; Weisz, N. Suppressed alpha oscillations predict intelligibility of speech and its acoustic details. Cereb. Cortex 2012, 22, 2466–2477. [Google Scholar] [CrossRef]
- Santoyo, A.E.; Gonzales, M.G.; Iqbal, Z.J.; Backer, K.C.; Balasubramaniam, R.; Bortfeld, H.; Shahin, A.J. Neurophysiological time course of timbre-induced music-like perception. J. Neurophysiol. 2023, 130, 291–302. [Google Scholar] [CrossRef]
- Weisz, N.; Hartmann, T.; Müller, N.; Lorenz, I.; Obleser, J. Alpha rhythms in audition: Cognitive and clinical perspectives. Front. Psychol. 2011, 2, 73. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Research Method | Main Findings |
|---|---|---|
| [21] | Vigabatrin (a neural inhibitory substance) was administered to rats exhibiting neurophysiological evidence of tinnitus to reduce hyperactivity in the auditory pathway. | Brainstem activity, which is increased in rats with neurophysiological evidence of tinnitus, was lowered by the application of Vigabatrin. |
| [41] |
|
|
| [39] | Music listening therapy, whereby the music was notched around the tinnitus pitch of each individual. Tinnitus sensation was measured at 6 and 12 months after the start of music therapy. | A decrease in tinnitus sensation was observed at 6 months, and an evenlarger reduction was seen at 12-month. Accompanying the decreased tinnitus sensation was a decrease in N1 auditory evoked field to a 500 Hz tone. |
| [40] |
|
|
| [42] | Multisensory tasks (auditory, visual, and tactile) were administered at 20 min daily for 20 days to a population with unilateral tinnitus (n = 18). In one group (integration group), the stimuli were administered on the tinnitus-affected side, while in another group (attention diversion group), the stimuli were administered on the opposite side. Tinnitus levels were reported before and after the training. | After 20 days, both groups showed a significant reduction in tinnitus. There was no significant difference between the two groups. |
| [56] | A low-rate (<100 Hz) electrical stimulation was applied to the apical portion of the cochlea of a person with a unilateral cochlear implant. Tinnitus levels were measured, as well as N1 auditory evoked potential and alpha power. | During stimulation, tinnitus sensation was reduced, N1 auditory evoked potential was reduced, and alpha power generated from the auditory cortex was enhanced. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahin, A.J.; Gonzales, M.G.; Dimitrijevic, A. Cross-Modal Tinnitus Remediation: A Tentative Theoretical Framework. Brain Sci. 2024, 14, 95. https://doi.org/10.3390/brainsci14010095
Shahin AJ, Gonzales MG, Dimitrijevic A. Cross-Modal Tinnitus Remediation: A Tentative Theoretical Framework. Brain Sciences. 2024; 14(1):95. https://doi.org/10.3390/brainsci14010095
Chicago/Turabian StyleShahin, Antoine J., Mariel G. Gonzales, and Andrew Dimitrijevic. 2024. "Cross-Modal Tinnitus Remediation: A Tentative Theoretical Framework" Brain Sciences 14, no. 1: 95. https://doi.org/10.3390/brainsci14010095
APA StyleShahin, A. J., Gonzales, M. G., & Dimitrijevic, A. (2024). Cross-Modal Tinnitus Remediation: A Tentative Theoretical Framework. Brain Sciences, 14(1), 95. https://doi.org/10.3390/brainsci14010095