
Hand Motions Reveal Attentional Status and Subliminal Semantic Processing: A Mouse-Tracking Technique

 ,
, 
Abstract
1. Introduction
1.1. Background
1.2. Subliminal Semantic Processing and Attention
1.3. Summary of Previous Research
1.4. Purposes of the Current Study
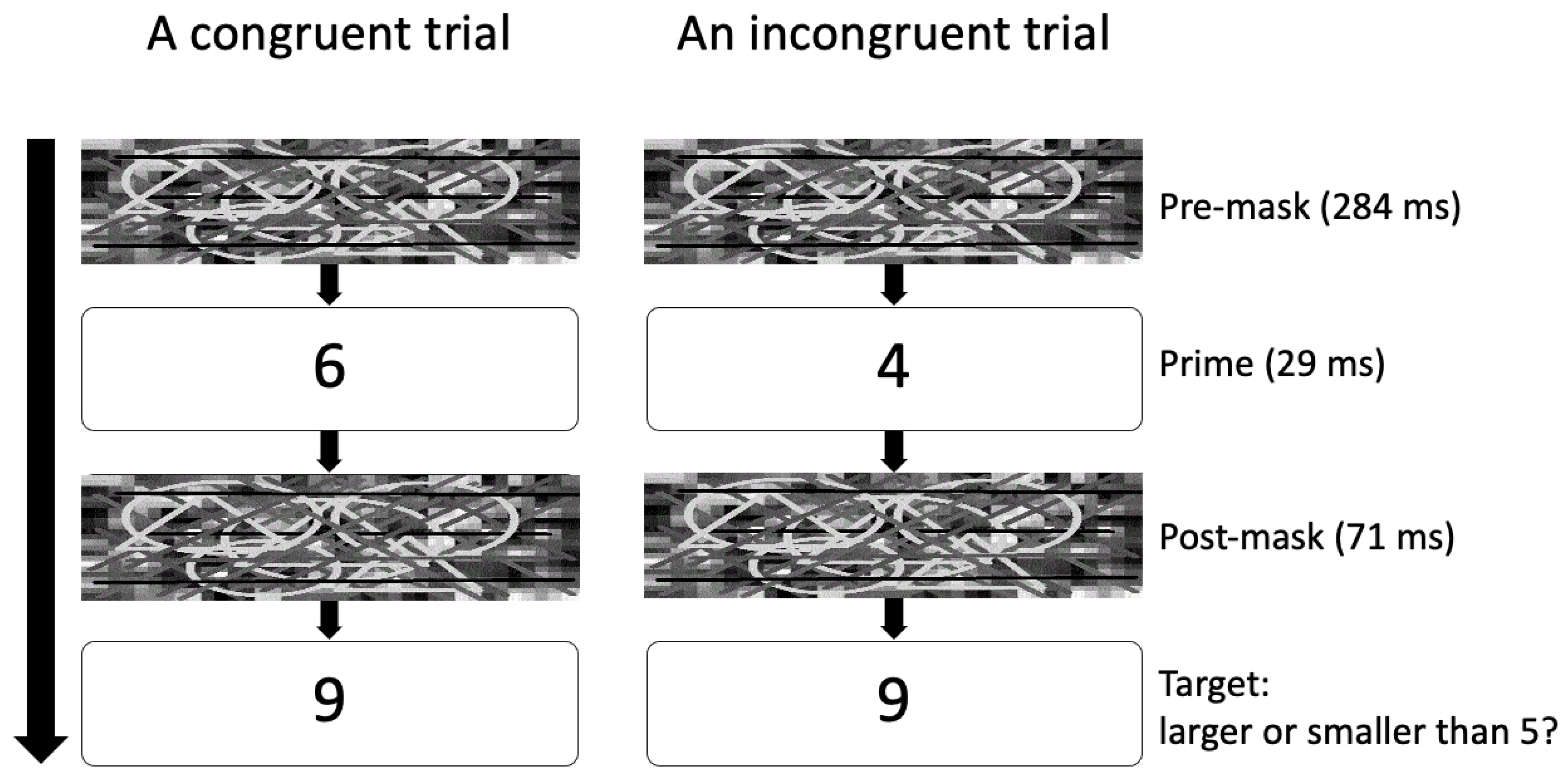
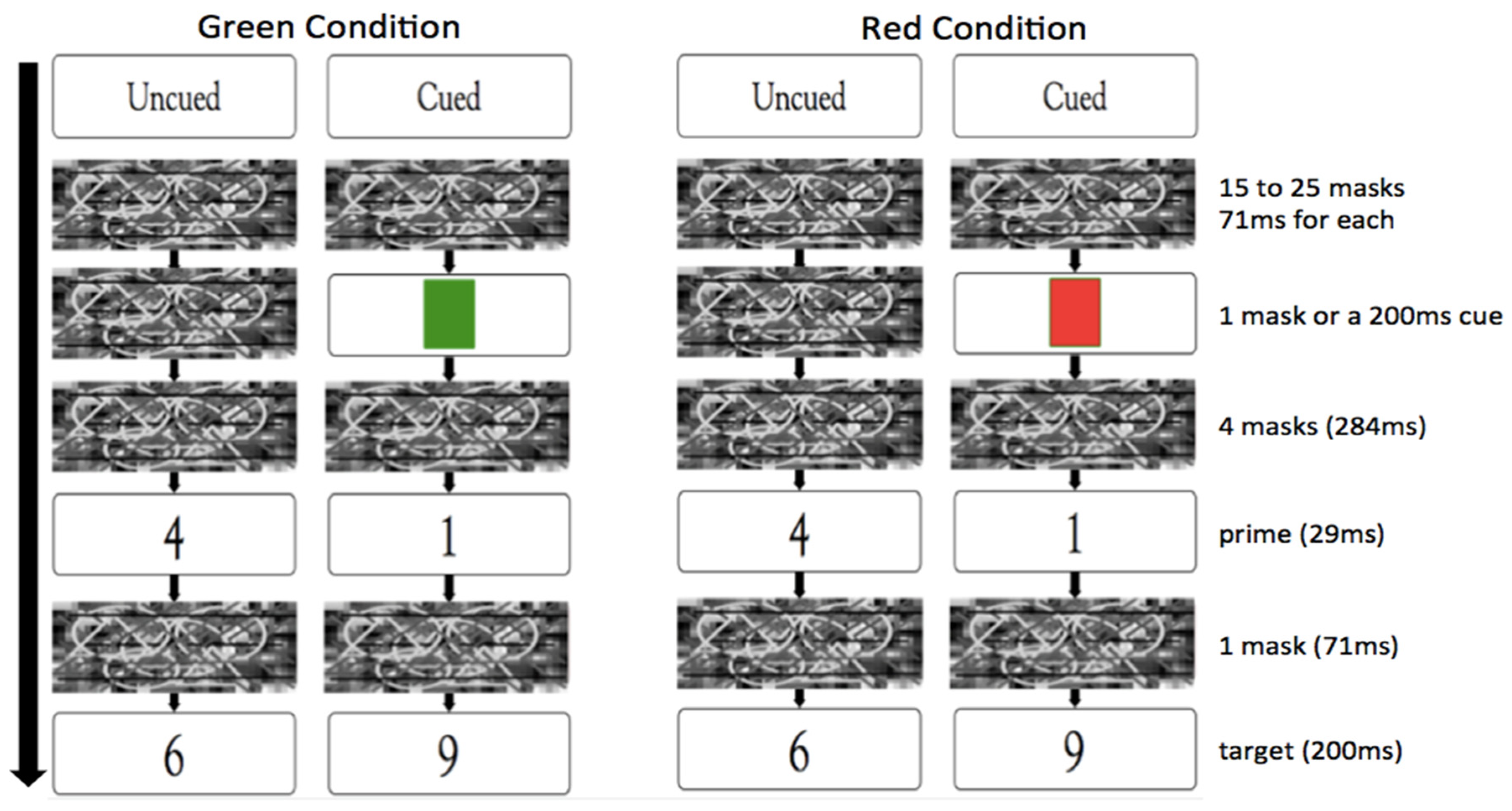
2. Materials and Methods
2.1. Overview of the Experiment
2.2. Participants
2.3. Materials and Apparatus
2.4. Experimental Procedure
3. Results
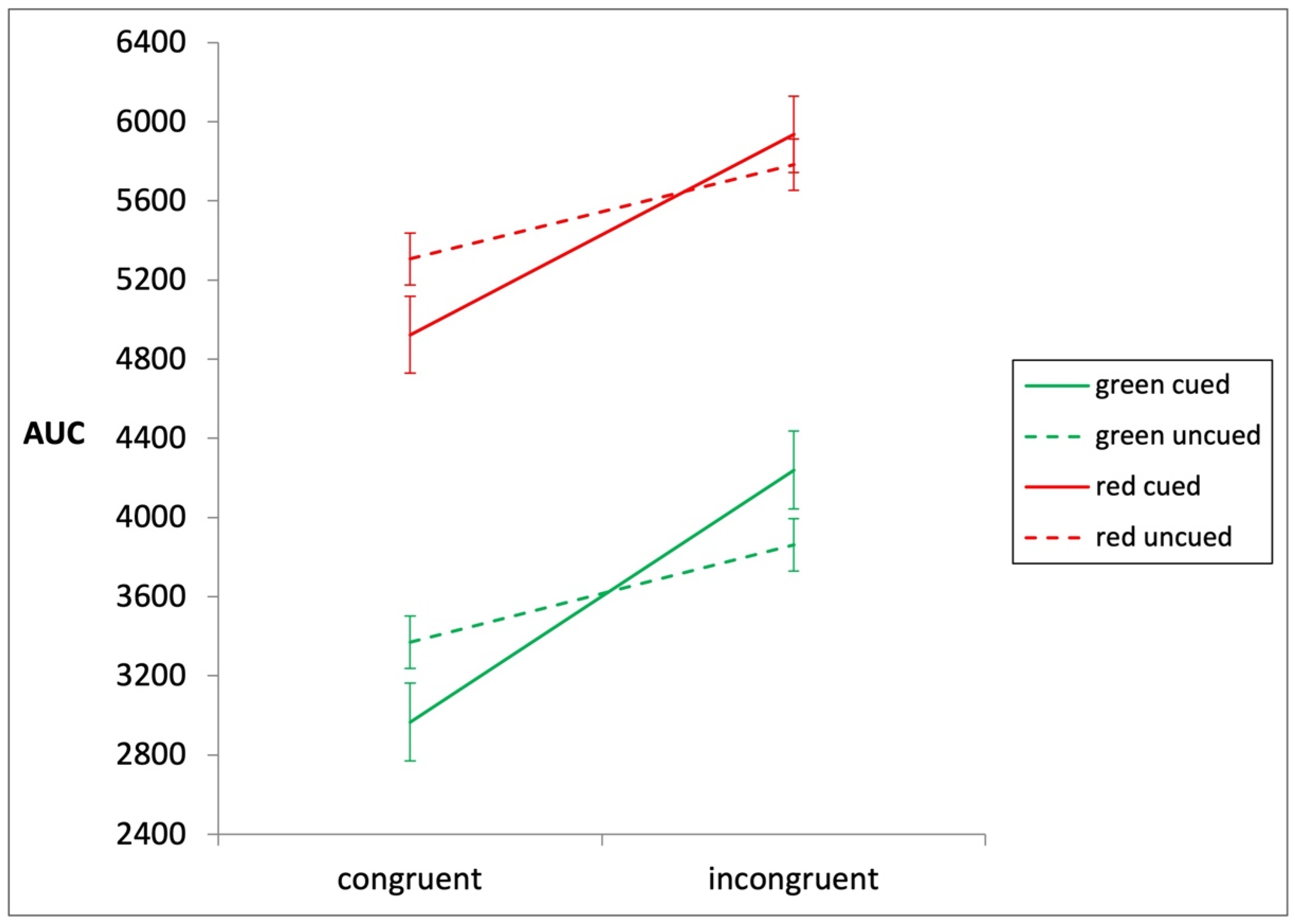
3.1. The Three-Way ANOVA
3.2. The Impact of Presenting an Attentional Cue
3.3. The Impact of Cue Color
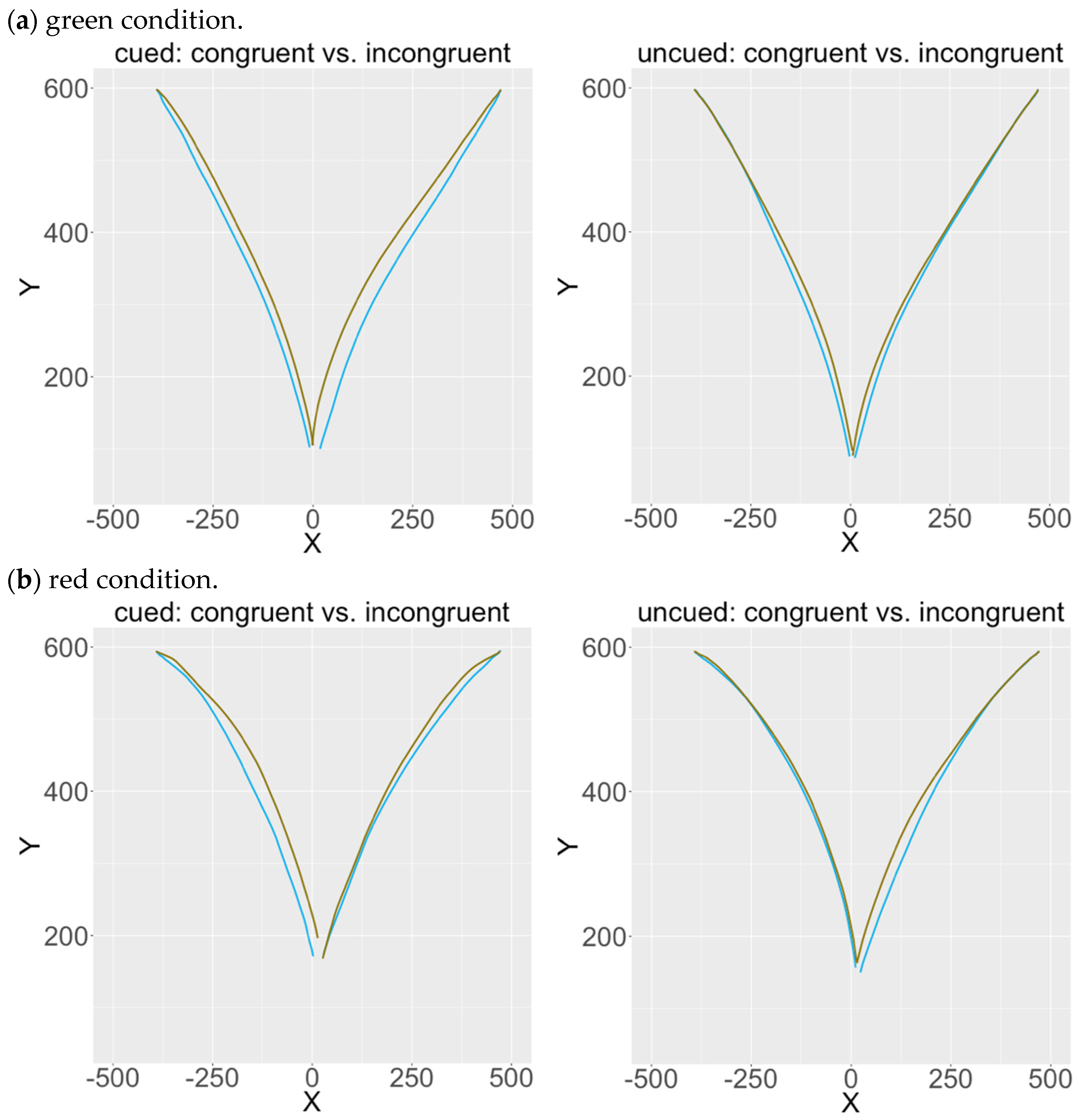
3.4. Characteristic of Average Motion Trajectories: Congruent vs. Incongruent Trials in Green and Red Conditions
3.5. Awareness Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Collins, A.M.; Loftus, E.F. A spreading-activation theory of semantic processing. Psychol. Rev. 1975, 82, 407–428. [Google Scholar] [CrossRef]
- Anderson, J.R. A spreading activation theory of memory. J. Verbal Learn. Verbal Behav. 1983, 22, 261–295. [Google Scholar] [CrossRef]
- Dehaene, S.; Changeux, J.P.; Naccache, L.; Sackur, J.; Sergent, C. Conscious, preconscious, and subliminal processing: A testable taxonomy. Trends Cogn. Sci. 2006, 10, 204–211. [Google Scholar] [CrossRef]
- Koch, C.; Tsuchiya, N. Attention and consciousness: Two distinct brain processes. Trends Cogn. Sci. 2007, 11, 16–22. [Google Scholar] [CrossRef]
- Van den Bussche, E.; Van den Noortgate, W.; Reynvoet, B. Mechanisms of masked priming: A meta-analysis. Psychol. Bull. 2009, 135, 452–477. [Google Scholar] [CrossRef]
- Barsalou, L.W.; Niedenthal, P.M.; Barbey, A.K.; Ruppert, J.A. Social embodiment. In Psychology of Learning and Motivation; Ross, B.H., Ed.; Academic Press: San Diego, CA, USA, 2003; Volume 43, pp. 43–92. [Google Scholar]
- Xiao, K.; Gao, Y.; Imran, S.A.; Chowdhury, S.; Commuri, S.; Jiang, F. Cross-modal motion aftereffects transfer between vision and touch in early deaf adults. Sci. Rep. 2021, 11, 4395. [Google Scholar] [CrossRef]
- Thelen, E.; Spencer, J.P. Postural control during reaching in young infants: A dynamic systems approach. Neurosci. Biobehav. Rev. 1998, 22, 507–514. [Google Scholar] [CrossRef]
- Keysers, C.; Paracampo, R.; Gazzola, V. What neuromodulation and lesion studies tell us about the function of the mirror neuron system and embodied cognition. Curr. Opin. Psychol. 2018, 24, 35–40. [Google Scholar] [CrossRef]
- Coombes, S.A.; Janelle, C.M.; Duley, A.R. Emotion and motor control: Movement attributes following affective picture processing. J. Mot. Behav. 2005, 37, 425–436. [Google Scholar] [CrossRef]
- Harris, C.M.; Wolpert, D.M. Signal-dependent noise determines motor planning. Nature 1998, 394, 780–784. [Google Scholar] [CrossRef]
- Gallivan, J.P.; Bowman, N.A.; Chapman, C.S.; Wolpert, D.M.; Flanagan, J.R. The sequential encoding of competing action goals involves dynamic restructuring of motor plans in working memory. J. Neurophysiol. 2016, 115, 3113–3122. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Song, J.H.; Nakayama, K. Hidden cognitive states revealed in choice reaching tasks. Trends Cogn. Sci. 2009, 13, 360–366. [Google Scholar] [CrossRef]
- Mollo, G.; Pulvermüller, F.; Hauk, O. Movement priming of EEG/MEG brain responses for action-words characterizes the link between language and action. Cortex 2016, 74, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Spivey, M.J.; Dale, R. Continuous dynamics in real-time cognition. Curr. Dir. Psychol. Sci. 2006, 15, 207–211. [Google Scholar] [CrossRef]
- Dale, R.; Kehoe, C.; Spivey, M.J. Graded motor responses in the time course of categorizing atypical exemplars. Mem. Cogn. 2007, 35, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Wolpert, D.M.; Landy, M.S. Motor control is decision-making. Curr. Opin. Neurobiol. 2012, 22, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Dotan, D.; Pinheiro-Chagas, P.; Al Roumi, F.; Dehaene, S. Track it to crack it: Dissecting processing stages with finger tracking. Trends Cogn. Sci. 2019, 23, 1058–1070. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kohn, N.; Yu, N.Y. Tracking mouse movement in feature inference: Category labels are different from feature labels. Mem. Cogn. 2007, 35, 852–863. [Google Scholar] [CrossRef]
- Freeman, J.B.; Pauker, K.; Apfelbaum, E.P.; Ambady, N. Continuous dynamics in the real-time perception of race. J. Exp. Soc. Psychol. 2010, 46, 179–185. [Google Scholar] [CrossRef]
- Yamauchi, T.; Xiao, K. Reading emotion from mouse cursor motions: Affective computing approach. Cogn. Sci. 2018, 42, 771–819. [Google Scholar] [CrossRef]
- Yamauchi, T.; Bowman, C. Mining cursor motions to find the gender, experience and feelings of computer users. In Proceedings of the IEEE International Conference on Data Mining (ICDM 2014): Workshop on Domain Driven Data Mining, Shenzhen, China, 14 December 2014; IEEE Computer Society: Piscataway, NJ, USA; pp. 221–230. [Google Scholar]
- Freihaut, P.; Göritz, A.S.; Rockstroh, C.; Blum, J. Tracking stress via the computer mouse? Promises and challenges of a potential behavioral stress marker. Behav. Res. Methods 2021, 53, 2281–2301. [Google Scholar] [PubMed]
- Freihaut, P.; Göritz, A.S. Using the computer mouse for stress measurement—An empirical investigation and critical review. Int. J. Hum. Comput. Stud. 2021, 145, 102520. [Google Scholar] [CrossRef]
- Leontyev, A.; Yamauchi, T. Discerning Mouse Trajectory Features with the Drift Diffusion Model. Cogn. Sci. 2021, 45, e13046. [Google Scholar] [CrossRef]
- Leontyev, A.; Sun, S.; Wolfe, M.; Yamauchi, T. Augmented Go/No-Go Task: Mouse Cursor Motion Measures Improve ADHD Symptom Assessment in Healthy College Students. Front. Psychol. 2018, 9, 496. [Google Scholar] [CrossRef] [PubMed]
- Leontyev, A.; Yamauchi, T. Mouse movement measures enhance the stop-signal task in adult ADHD assessment. PLoS ONE 2019, 14, e0225437. [Google Scholar] [CrossRef]
- Leontyev, A.; Yamauchi, T.; Razavi, M. Machine Learning Stop Signal Test (ML-SST): ML-based Mouse Tracking Enhances Adult ADHD Diagnosis. In Proceedings of the 2019 8th International Conference on Affective Computing and Intelligent Interaction Workshops and Demos (ACIIW), Cambridge, UK, 3–6 September 2019; IEEE Computer Society: Piscataway, NJ, USA. [Google Scholar]
- Freeman, J.B.; Johnson, K.L. More than meets the eye: Split-second social perception. Trends Cogn. Sci. 2016, 20, 362–374. [Google Scholar] [CrossRef]
- Freeman, J.B. Doing psychological science by hand. Curr. Dir. Psychol. Sci. 2018, 27, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Tafuro, A.; Vallesi, A.; Ambrosini, E. Cognitive brakes in interference resolution: A mouse-tracking and EEG co-registration study. Cortex 2020, 133, 188–200. [Google Scholar] [CrossRef]
- Freeman, J.B.; Ambady, N. Motions of the hand expose the partial and parallel activation of stereotypes. Psychol. Sci. 2009, 20, 1183–1188. [Google Scholar] [CrossRef] [PubMed]
- Wojnowicz, M.T.; Ferguson, M.J.; Dale, R.; Spivey, M.J. The self-organization of explicit attitudes. Psychol. Sci. 2009, 20, 1428–1435. [Google Scholar] [CrossRef] [PubMed]
- Calluso, C.; Committeri, G.; Pezzulo, G.; Lepora, N.; Tosoni, A. Analysis of hand kinematics reveals inter-individual differences in intertemporal decision dynamics. Exp. Brain Res. 2015, 233, 3597–3611. [Google Scholar] [CrossRef]
- Farmer, T.A.; Anderson, S.E.; Spivey, M.J. Gradiency and visual context in syntactic garden-paths. J. Mem. Lang. 2007, 57, 570–595. [Google Scholar] [CrossRef]
- Dehaene, S.; Naccache, L.; Le Clec’H, G.; Koechlin, E.; Mueller, M.; Dehaene-Lambertz, G.; van de Moortele, P.F.; Le Bihan, D. Imaging unconscious semantic priming. Nature 1998, 395, 597–600. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, A.G.; Sean, C.D.; Richard, L.A. Three cognitive markers of unconscious semantic activation. Science 1996, 273, 1699–1702. [Google Scholar] [CrossRef] [PubMed]
- Berkovitch, L.; Dehaene, S. Subliminal syntactic priming. Cogn. Psychol. 2019, 109, 26–46. [Google Scholar] [CrossRef]
- Naccache, L.; Blandin, E.; Dehaene, S. Unconscious masked priming depends on temporal attention. Psychol. Sci. 2002, 13, 416–424. [Google Scholar] [CrossRef]
- Prasad, S.; Mishra, R.K. The nature of unconscious attention to subliminal cues. Vision 2019, 3, 38. [Google Scholar] [CrossRef]
- Mao, X.; Xie, C.; Shi, J.; Huang, Q.; Jiang, R.; Meng, F.; Shen, H.; Miao, L.; Cui, S.; Li, A. The essential role of executive attention in unconscious visuomotor priming. Front. Psychol. 2022, 13, 800781. [Google Scholar] [CrossRef]
- Van Boxtel, J.J.; Tsuchiya, N.; Koch, C. Consciousness and attention: On sufficiency and necessity. Front. Psychol. 2010, 1, 217. [Google Scholar] [CrossRef]
- Cohen, M.A.; Dennett, D.C.; Kanwisher, N. What is the bandwidth of perceptual experience? Trends Cogn. Sci. 2016, 20, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Mashour, G.A.; Roelfsema, P.; Changeux, J.P.; Dehaene, S. Conscious processing and the global neuronal workspace hypothesis. Neuron 2020, 105, 776–798. [Google Scholar] [CrossRef]
- Sim, E.J.; Harpaintner, M.; Kiefer, M. Is subliminal face processing modulated by attentional task sets? Evidence from masked priming effects in a gender decision task. Open Psychol. 2020, 2, 76–89. [Google Scholar] [CrossRef]
- Khalid, S.; Finkbeiner, M.; König, P.; Ansorge, U. Subcortical human face processing? Evidence from masked priming. J. Exp. Psychol. Hum. Percept. Perform. 2013, 39, 989–1002. [Google Scholar] [CrossRef]
- Wang, J.; JIAO, R.; Zhang, M. The mechanism of the effect of task setting on negative compatibility effect: The effect of top-down cognition control on subliminal prime processing. Acta Psychol. Sin. 2016, 48, 1370–1378. [Google Scholar] [CrossRef]
- Kiefer, M.; Trumpp, N.M.; Schaitz, C.; Reuss, H.; Kunde, W. Attentional modulation of masked semantic priming by visible and masked task cues. Cognition 2019, 187, 62–77. [Google Scholar] [CrossRef]
- Block, N. Perceptual consciousness overflows cognitive access. Trends Cogn. Sci. 2011, 15, 567–575. [Google Scholar] [CrossRef]
- Block, N. Rich conscious perception outside focal attention. Trends Cogn. Sci. 2014, 18, 445–447. [Google Scholar] [CrossRef]
- Handschack, J.; Rothkirch, M.; Sterzer, P.; Hesselmann, G. No effect of attentional modulation by spatial cueing in a masked numerical priming paradigm using continuous flash suppression (CFS). PeerJ 2023, 11, e14607. [Google Scholar] [CrossRef] [PubMed]
- Eo, K.; Cha, O.; Chong, S.C.; Kang, M.S. Less is more: Semantic information survives interocular suppression when attention is diverted. J. Neurosci. 2016, 36, 5489–5497. [Google Scholar] [CrossRef][Green Version]
- Rahnev, D.A.; Huang, E.; Lau, H. Subliminal stimuli in the near absence of attention influence top-down cognitive control. Atten. Percept. Psychophys. 2012, 74, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, M.; Palermo, R. The role of spatial attention in nonconscious processing: A comparison of face and nonface stimuli. Psychol. Sci. 2009, 20, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.; Reddy, L.; Koch, C. Face identification in the near-absence of focal attention. Vis. Res. 2006, 46, 2336–2343. [Google Scholar] [CrossRef]
- Reddy, L.; Patrick, W.; Christof, K. Face-gender discrimination is possible in the near-absence of attention. J. Vis. 2004, 4, 4. [Google Scholar] [CrossRef]
- Li, F.F.; VanRullen, R.; Koch, C.; Perona, P. Rapid natural scene categorization in the near absence of attention. Proc. Natl. Acad. Sci. USA 2002, 99, 9596–9601. [Google Scholar] [CrossRef] [PubMed]
- Underwood, G.; Templeman, E.; Lamming, L.; Foulsham, T. Is attention necessary for object identification? Evidence from eye movements during the inspection of real-world scenes. Conscious. Cogn. 2008, 17, 159–170. [Google Scholar] [CrossRef]
- Stillman, P.E.; Shen, X.; Ferguson, M.J. Ferguson. How mouse-tracking can advance social cognitive theory. Trends Cogn. Sci. 2018, 22, 531–543. [Google Scholar] [CrossRef]
- Freeman, J.B.; Ambady, N. MouseTracker: Software for studying real-time mental processing using a computer mouse-tracking method. Behav. Res. Methods 2010, 42, 226–241. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.B.; Ambady, N. A dynamic interactive theory of person construal. Psychol. Rev. 2011, 118, 247–279. [Google Scholar] [CrossRef]
- Xiao, K.; Yamauchi, T. Subliminal semantic priming in near absence of attention: A cursor motion study. Conscious. Cogn. 2015, 38, 88–98. [Google Scholar] [CrossRef]
- Lowry, M.; Trivedi, N.; Boyd, P.; Julian, A.; Treviño, M.; Lama, Y.; Heley, K.; Perna, F. Making decisions about health information on social media: A mouse-tracking study. Cogn. Res. Princ. Implic. 2022, 7, 68. [Google Scholar] [CrossRef]
- Vadillo, M.A.; Konstantinidis, E.; Shanks, D.R. Underpowered samples, false negatives, and unconscious learning. Psychon. Bull. Rev. 2016, 23, 87–102. [Google Scholar] [CrossRef]
- Yang, Y.H.; Tien, Y.H.; Yang, P.L.; Yeh, S.L. Role of consciousness in temporal integration of semantic information. Cogn. Affect. Behav. Neurosci. 2017, 17, 954–972. [Google Scholar] [CrossRef]
- Avneon, M.; Lamy, D. Reexamining unconscious response priming: A liminal-prime paradigm. Conscious. Cogn. 2018, 59, 87–103. [Google Scholar] [CrossRef]
- Schneider, I.K.; van Harreveld, F.; Rotteveel, M.; Topolinski, S.; van der Pligt, J.; Schwarz, N.; Koole, S.L. The path of ambivalence: Tracing the pull of opposing evaluations using mouse trajectories. Front. Psychol. 2015, 6, 996. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Incera, S.; McLennan, C.T. A reverse Stroop task with mouse tracking. Front. Psychol. 2016, 7, 670. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Yamauchi, T. Semantic priming revealed by mouse movement trajectories. Conscious. Cogn. 2014, 27, 42–52. [Google Scholar] [CrossRef]
- Xiao, K.; Yamauchi, T. The role of attention in subliminal semantic processing: A mouse tracking study. PLoS ONE 2017, 12, e0178740. [Google Scholar] [CrossRef]
- Xiao, K.; Yamauchi, T.; Bowman, C. Assessing Masked Semantic Priming: Cursor Trajectory versus Response Time Measures. In Proceedings of the 37th Annual Meeting of the Cognitive Science Society (CogSci 2015), Pasadena, CA, USA, 22–25 July 2015. [Google Scholar]
- Kareklas, I.; Muehling, D.D.; King, S. The effect of color and self-view priming in persuasive communications. J. Bus. Res. 2019, 98, 33–49. [Google Scholar] [CrossRef]
- Argiris, G.; Rumiati, R.I.; Crepaldi, D. No fruits without color: Cross-modal priming and EEG reveal different roles for different features across semantic categories. PLoS ONE 2021, 16, e0234219. [Google Scholar] [CrossRef]
- Schneider, W.; Shiffrin, R.M. Controlled and automatic human information processing: I. Detection, search, and attention. Psychol. Rev. 1977, 84, 1–66. [Google Scholar] [CrossRef]
- Shiffrin, R.M.; Schneider, W. Controlled and automatic human information processing: II. Perceptual learning, automatic attending and a general theory. Psychol. Rev. 1977, 84, 127–190. [Google Scholar] [CrossRef]
- Cohen, M.A.; Cavanagh, P.; Chun, M.M.; Nakayama, K. The attentional requirements of consciousness. Trends Cogn. Sci. 2012, 16, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Dehaene, S.; Kerszberg, M.; Changeux, J.P. A neuronal model of a global workspace in effortful cognitive tasks. Proc. Natl. Acad. Sci. USA 1998, 95, 14529–14534. [Google Scholar] [CrossRef] [PubMed]
- Dehaene, S.; Naccache, L. Towards a cognitive neuroscience of consciousness: Basic evidence and a workspace framework. Cognition 2001, 79, 1–37. [Google Scholar] [CrossRef]
- Baars, B.J.; Franklin, S.; Ramsoy, T.Z. Global workspace dynamics: Cortical “binding and propagation” enables conscious contents. Front. Psychol. 2013, 4, 200. [Google Scholar] [CrossRef]
- Kiefer, M.; Brendel, D. Attentional modulation of unconscious “automatic” processes: Evidence from event-related potentials in a masked priming paradigm. J. Cogn. Neurosci. 2006, 18, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Mudrik, L.; Faivre, N.; Koch, C. Information integration without awareness. Trends Cogn. Sci. 2014, 18, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Mudrik, L.; Breska, A.; Lamy, D.; Deouell, L.Y. Integration without awareness: Expanding the limits of unconscious processing. Psychol. Sci. 2011, 22, 764–770. [Google Scholar] [CrossRef]
- Koch, C.; Massimini, M.; Boly, M.; Tononi, G. Neural correlates of consciousness: Progress and problems. Nat. Rev. Neurosci. 2016, 17, 307–321. [Google Scholar] [CrossRef]
- Gavazzi, G.; Rossi, A.; Orsolini, S.; Diciotti, S.; Giovannelli, F.; Salvadori, E.; Pantoni, L.; Mascalchi, M.; Viggiano, M.P. Impulsivity trait and proactive cognitive control: An fMRI study. Eur. J. Neurosci. 2019, 49, 1171–1179. [Google Scholar] [CrossRef]
- Quétard, B.; Spatola, N.; Parris, B.A.; Ferrand, L.; Augustinova, M. A mouse-tracking study of the composite nature of the Stroop effect at the level of response execution. PLoS ONE 2023, 18, e0279036. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictor | b | SE | 95% CI | t | p | |
|---|---|---|---|---|---|---|
| Cued | (intercept) | 1251.03 | 198.07 | [862.81, 1639.25] | 6.32 *** | <0.001 |
| d’ | 0.17 | 0.16 | [−0.16, 0.50] | 1.02 | 0.313 | |
| Uncued | (intercept) | 479.97 | 133.92 | [217.49, 742.45] | 3.58 ** | 0.001 |
| d’ | 0.13 | 0.17 | [−0.19, 0.46] | 0.80 | 0.429 |
| Predictor | b | SE | 95% CI | t | p | |
|---|---|---|---|---|---|---|
| Cued | (intercept) | 1014.28 | 297.93 | [430.34, 1598.22] | 3.40 ** | 0.002 |
| d’ | −0.01 | 0.17 | [−0.35, 0.34] | −0.03 | 0.973 | |
| Uncued | (intercept) | 474.07 | 132.30 | [214.76, 733.38] | 3.57 ** | 0.001 |
| d’ | 0.04 | 0.16 | [−0.28, 0.36] | 0.25 | 0.807 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, K.; Zhang, A.; Qu, J.; Deng, F.; Guo, C.; Yamauchi, T. Hand Motions Reveal Attentional Status and Subliminal Semantic Processing: A Mouse-Tracking Technique. Brain Sci. 2023, 13, 1267. https://doi.org/10.3390/brainsci13091267
Xiao K, Zhang A, Qu J, Deng F, Guo C, Yamauchi T. Hand Motions Reveal Attentional Status and Subliminal Semantic Processing: A Mouse-Tracking Technique. Brain Sciences. 2023; 13(9):1267. https://doi.org/10.3390/brainsci13091267
Chicago/Turabian StyleXiao, Kunchen, Anqi Zhang, Jingke Qu, Feifei Deng, Chenyan Guo, and Takashi Yamauchi. 2023. "Hand Motions Reveal Attentional Status and Subliminal Semantic Processing: A Mouse-Tracking Technique" Brain Sciences 13, no. 9: 1267. https://doi.org/10.3390/brainsci13091267
APA StyleXiao, K., Zhang, A., Qu, J., Deng, F., Guo, C., & Yamauchi, T. (2023). Hand Motions Reveal Attentional Status and Subliminal Semantic Processing: A Mouse-Tracking Technique. Brain Sciences, 13(9), 1267. https://doi.org/10.3390/brainsci13091267