
A Potential Biomarker of Brain Activity in Autism Spectrum Disorders: A Pilot fNIRS Study in Female Preschoolers

 , , ,
, , ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Apparatus and Experimental Design
2.3. Signal Processing
2.4. Statistical Analysis
3. Results
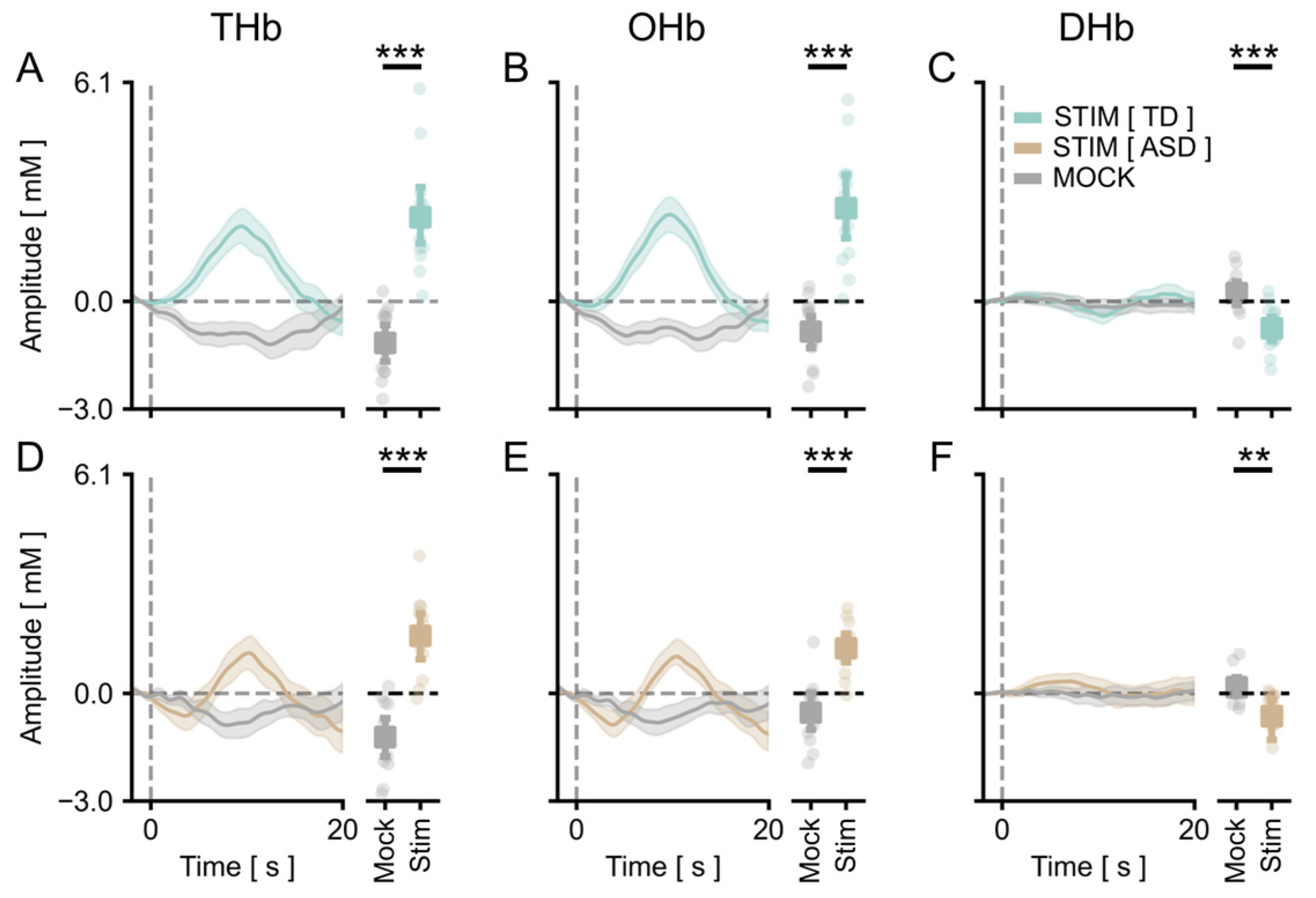
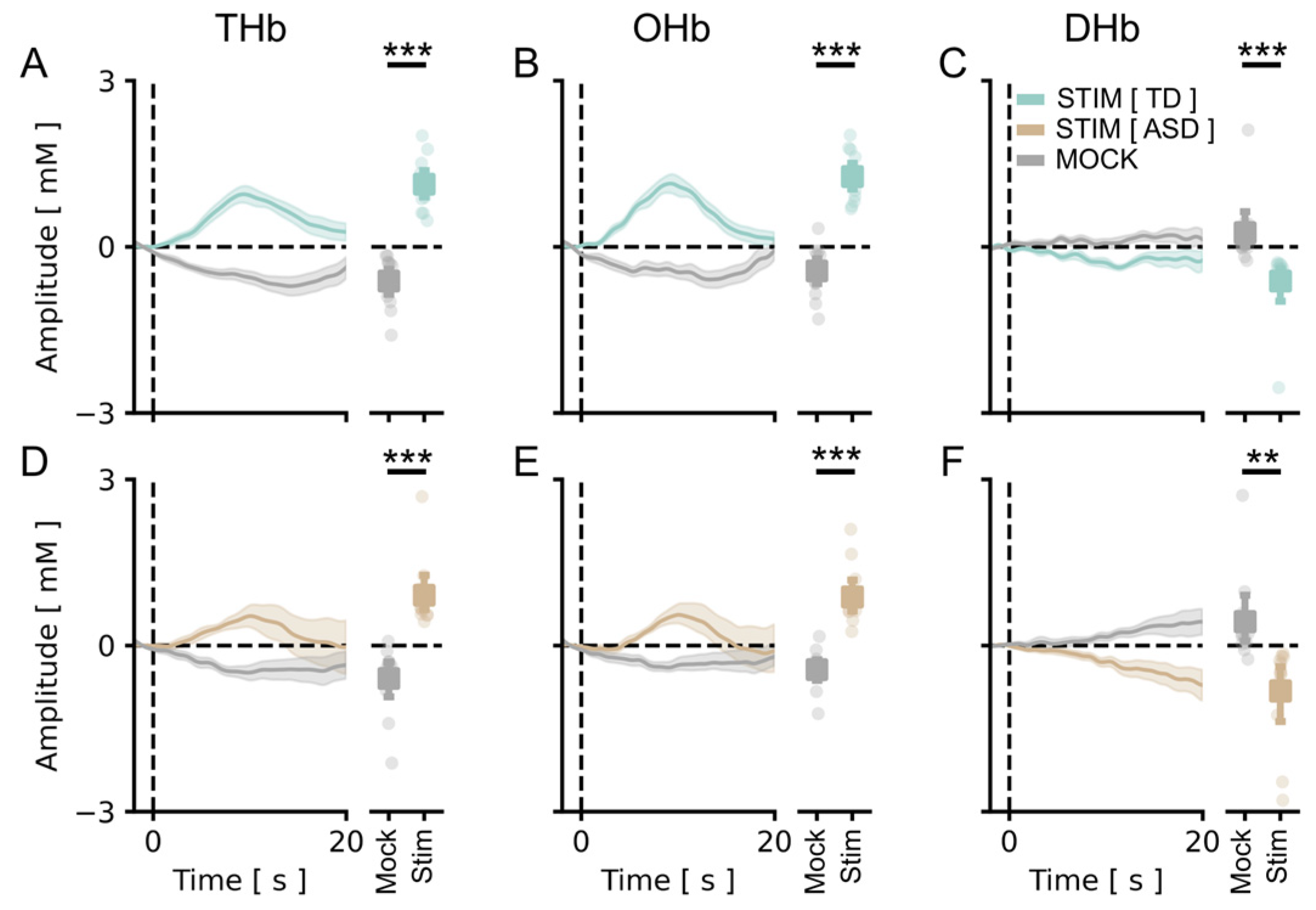
3.1. A Cartoon-Based Stimulus Is Able to Evoke Reliable Responses in the Visual Cortex of fASD Children
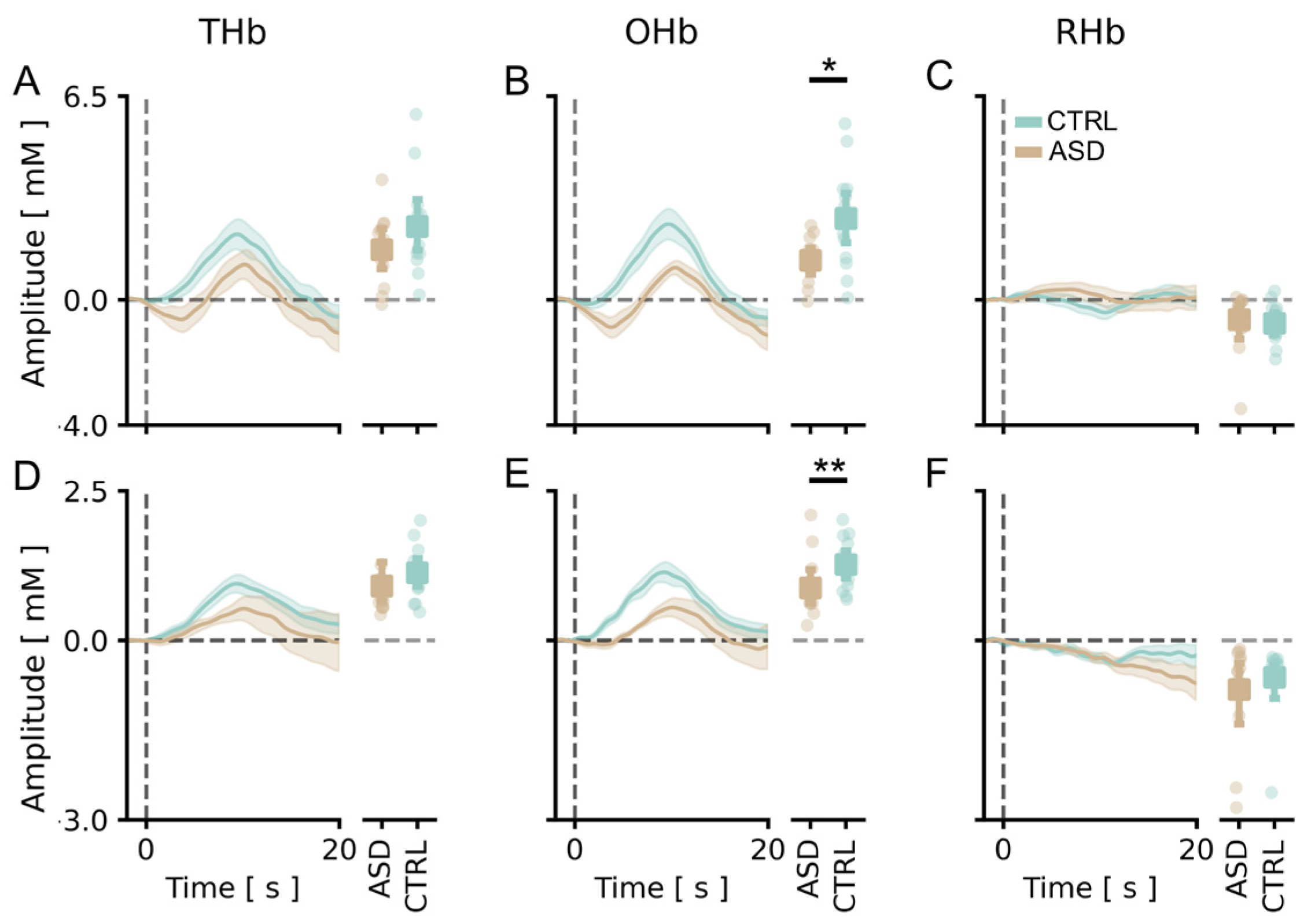
3.2. The Amplitude of Visual Cortical Responses Is Significantly Lower in the ASD Cohort
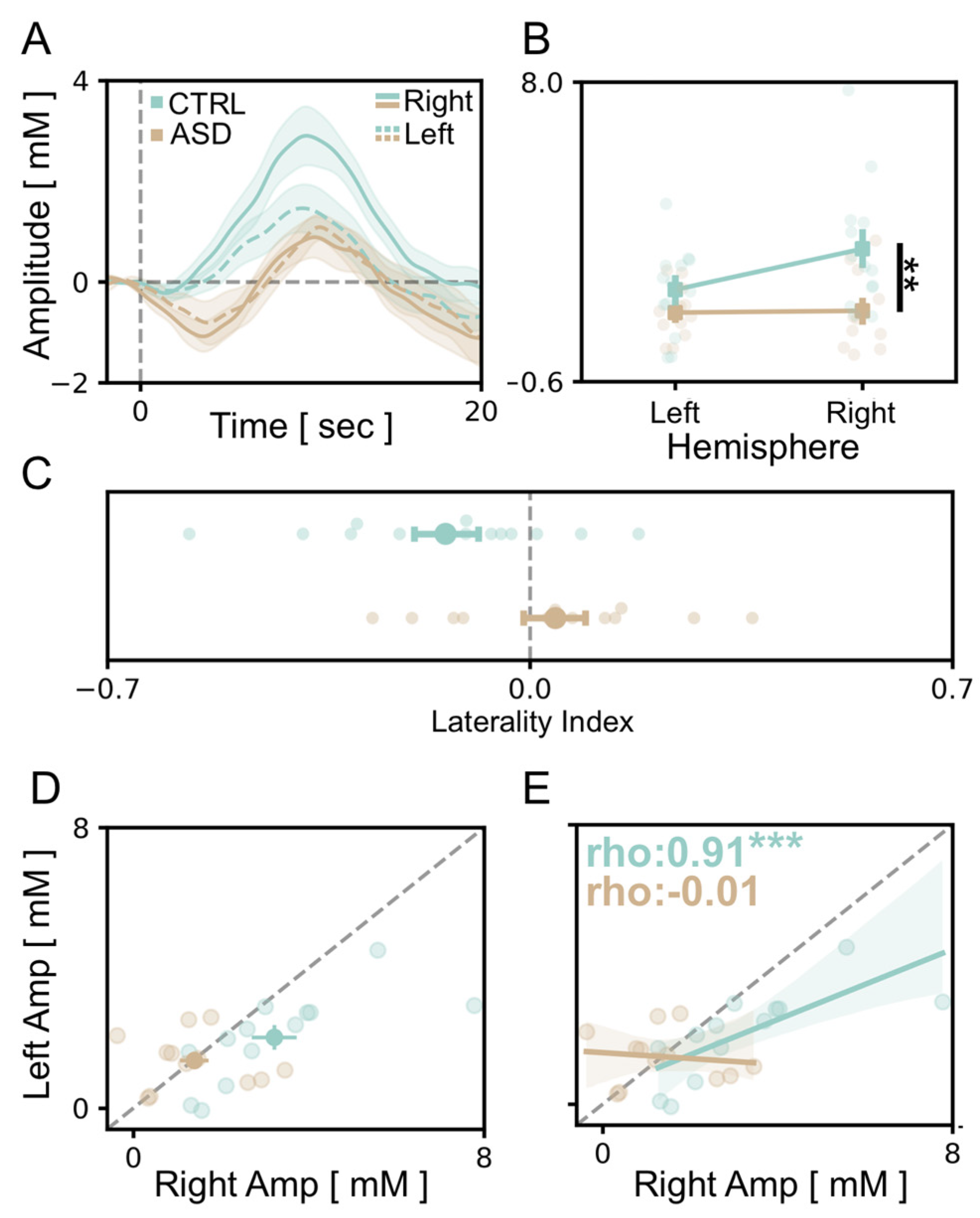
3.3. Atypical Lateralization of Visual Responses in the ASD Cohort
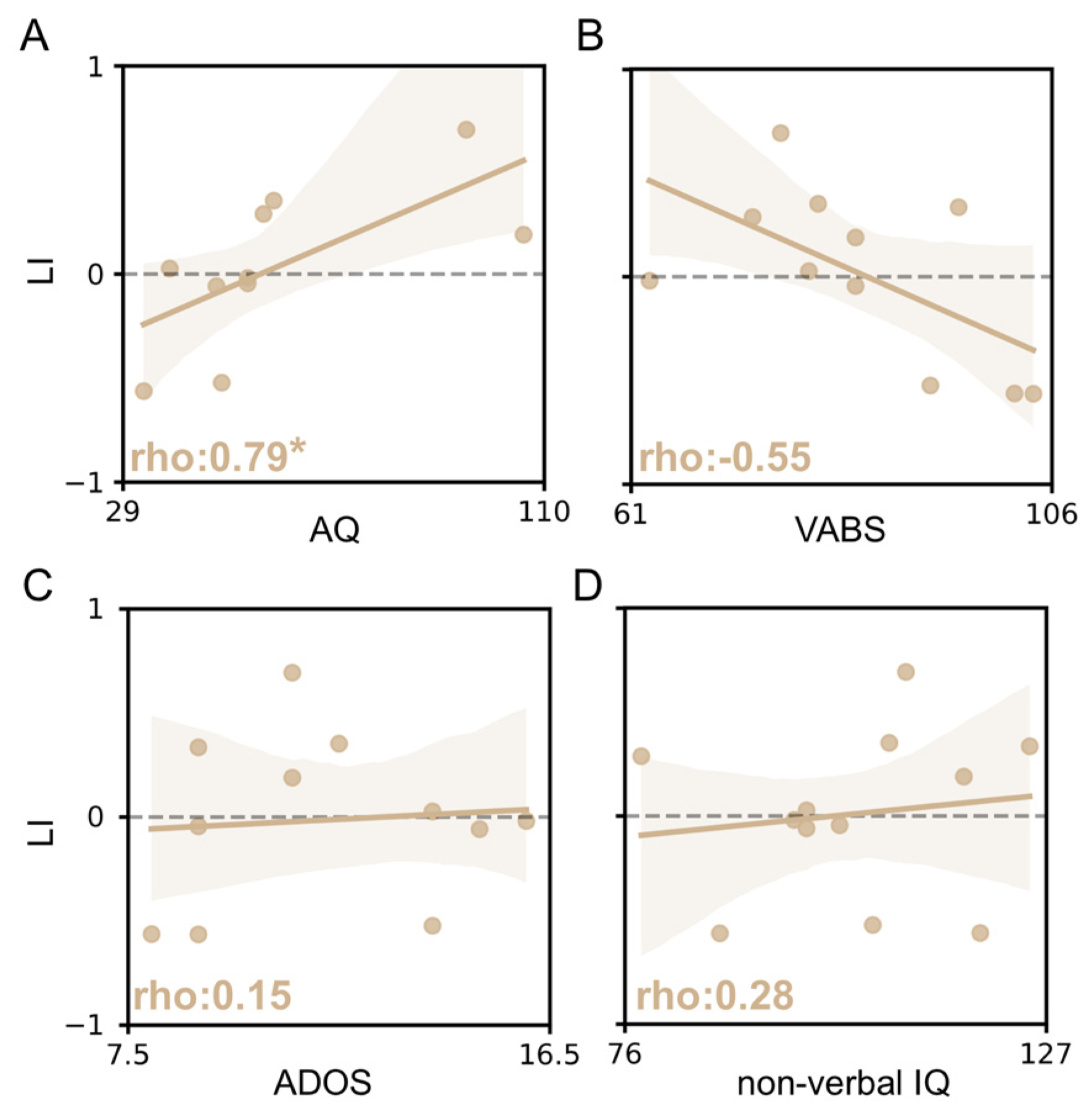
3.4. vHDR Might Be Predictive of Symptom Severity in fASD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mangun, G.R.; Luck, S.J.; Plager, R.; Loftus, W.; Hillyard, S.A.; Handy, T.; Clark, V.P.; Gazzaniga, M.S. Monitoring the Visual World: Hemispheric Asymmetries and Subcortical Processes in Attention. J. Cogn. Neurosci. 1994, 6, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Zeidan, J.; Fombonne, E.; Scorah, J.; Ibrahim, A.; Durkin, M.S.; Saxena, S.; Yusuf, A.; Shih, A.; Elsabbagh, M. Global Prevalence of Autism: A Systematic Review Update. Autism Res. 2022, 15, 778–790. [Google Scholar] [CrossRef] [PubMed]
- Narzisi, A.; Posada, M.; Barbieri, F.; Chericoni, N.; Ciuffolini, D.; Pinzino, M.; Romano, R.; Scattoni, M.L.; Tancredi, R.; Calderoni, S.; et al. Prevalence of Autism Spectrum Disorder in a Large Italian Catchment Area: A School-Based Population Study within the ASDEU Project. Epidemiol. Psychiatr. Sci. 2020, 29, e5. [Google Scholar] [CrossRef]
- Dawson, G.; Rogers, S.; Munson, J.; Smith, M.; Winter, J.; Greenson, J.; Donaldson, A.; Varley, J. Randomized, Controlled Trial of an Intervention for Toddlers with Autism: The Early Start Denver Model. Pediatrics 2010, 125, e17–e23. [Google Scholar] [CrossRef] [PubMed]
- Hyman, S.L.; Levy, S.E.; Myers, S.M.; Council on Children with Disabilities, Section on Developmental and Behavioral Pediatrics. Identification, Evaluation, and Management of Children with Autism Spectrum Disorder. Pediatrics 2020, 145, e20193447. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.E.; Dai, Y.G.; Fein, D.A.; Robins, D.L. Characteristics of Toddlers with Early versus Later Diagnosis of Autism Spectrum Disorder. Autism 2021, 25, 416–428. [Google Scholar] [CrossRef]
- Landa, R.J. Diagnosis of Autism Spectrum Disorders in the First 3 Years of Life. Nat. Clin. Pract. Neurol. 2008, 4, 138–147. [Google Scholar] [CrossRef]
- Wolfers, T.; Floris, D.L.; Dinga, R.; van Rooij, D.; Isakoglou, C.; Kia, S.M.; Zabihi, M.; Llera, A.; Chowdanayaka, R.; Kumar, V.J.; et al. From Pattern Classification to Stratification: Towards Conceptualizing the Heterogeneity of Autism Spectrum Disorder. Neurosci. Biobehav. Rev. 2019, 104, 240–254. [Google Scholar] [CrossRef]
- Shulman, C.; Esler, A.; Morrier, M.J.; Rice, C.E. Diagnosis of Autism Spectrum Disorder Across the Lifespan. Psychiatr. Clin. N. Am. 2020, 43, 583–603. [Google Scholar] [CrossRef]
- Shen, L.; Zhao, Y.; Zhang, H.; Feng, C.; Gao, Y.; Zhao, D.; Xia, S.; Hong, Q.; Iqbal, J.; Liu, X.K.; et al. Advances in Biomarker Studies in Autism Spectrum Disorders. Adv. Exp. Med. Biol. 2019, 1118, 207–233. [Google Scholar]
- Dworzynski, K.; Ronald, A.; Bolton, P.; Happé, F. How Different Are Girls and Boys above and below the Diagnostic Threshold for Autism Spectrum Disorders? J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Van Wijngaarden-Cremers, P.J.M.; van Eeten, E.; Groen, W.B.; Van Deurzen, P.A.; Oosterling, I.J.; Van der Gaag, R.J. Gender and Age Differences in the Core Triad of Impairments in Autism Spectrum Disorders: A Systematic Review and Meta-Analysis. J. Autism Dev. Disord. 2014, 44, 627–635. [Google Scholar] [CrossRef]
- Lai, M.-C.; Baron-Cohen, S.; Buxbaum, J.D. Understanding Autism in the Light of Sex/gender. Mol. Autism 2015, 6, 24. [Google Scholar] [CrossRef]
- Dean, M.; Harwood, R.; Kasari, C. The Art of Camouflage: Gender Differences in the Social Behaviors of Girls and Boys with Autism Spectrum Disorder. Autism 2017, 21, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Green, R.M.; Travers, A.M.; Howe, Y.; McDougle, C.J. Women and Autism Spectrum Disorder: Diagnosis and Implications for Treatment of Adolescents and Adults. Curr. Psychiatry Rep. 2019, 21, 22. [Google Scholar] [CrossRef] [PubMed]
- Calderoni, S. Sex/gender Differences in Children with Autism Spectrum Disorder: A Brief Overview on Epidemiology, Symptom Profile, and Neuroanatomy. J. Neurosci. Res. 2022, 101, 739–750. [Google Scholar] [CrossRef]
- Billeci, L.; Callara, A.L.; Guiducci, L.; Prosperi, M.; Morales, M.A.; Calderoni, S.; Muratori, F.; Santocchi, E. A Randomized Controlled Trial into the Effects of Probiotics on Electroencephalography in Preschoolers with Autism. Autism 2023, 27, 117–132. [Google Scholar] [CrossRef]
- Frye, R.E.; Vassall, S.; Kaur, G.; Lewis, C.; Karim, M.; Rossignol, D. Emerging Biomarkers in Autism Spectrum Disorder: A Systematic Review. Ann. Transl. Med. 2019, 7, 792. [Google Scholar] [CrossRef]
- Ayoub, M.J.; Keegan, L.; Tager-Flusberg, H.; Gill, S.V. Neuroimaging Techniques as Descriptive and Diagnostic Tools for Infants at Risk for Autism Spectrum Disorder: A Systematic Review. Brain Sci. 2022, 12, 602. [Google Scholar] [CrossRef]
- Kikuchi, M.; Yoshimura, Y.; Mutou, K.; Minabe, Y. Magnetoencephalography in the Study of Children with Autism Spectrum Disorder. Psychiatry Clin. Neurosci. 2016, 70, 74–88. [Google Scholar] [CrossRef]
- Amaral, D.G.; Schumann, C.M.; Nordahl, C.W. Neuroanatomy of Autism. Trends Neurosci. 2008, 31, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E.; Campbell, K.; Solso, S. Brain Growth across the Life Span in Autism: Age-Specific Changes in Anatomical Pathology. Brain Res. 2011, 1380, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Bosl, W.; Tierney, A.; Tager-Flusberg, H.; Nelson, C. EEG Complexity as a Biomarker for Autism Spectrum Disorder Risk. BMC Med. 2011, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Billeci, L.; Sicca, F.; Maharatna, K.; Apicella, F.; Narzisi, A.; Campatelli, G.; Calderoni, S.; Pioggia, G.; Muratori, F. On the Application of Quantitative EEG for Characterizing Autistic Brain: A Systematic Review. Front. Hum. Neurosci. 2013, 7, 442. [Google Scholar] [CrossRef]
- O’Reilly, C.; Lewis, J.D.; Elsabbagh, M. Is Functional Brain Connectivity Atypical in Autism? A Systematic Review of EEG and MEG Studies. PLoS ONE 2017, 12, e0175870. [Google Scholar] [CrossRef] [PubMed]
- Pagnozzi, A.M.; Conti, E.; Calderoni, S.; Fripp, J.; Rose, S.E. A Systematic Review of Structural MRI Biomarkers in Autism Spectrum Disorder: A Machine Learning Perspective. Int. J. Dev. Neurosci. 2018, 71, 68–82. [Google Scholar] [CrossRef]
- van de Rijt, L.P.; van Wanrooij, M.M.; Snik, A.F.; Mylanus, E.A.; van Opstal, A.J.; Roye, A. Measuring cortical activity during auditory processing with functional near-infrared spectroscopy. J. Hear. Sci. 2018, 8, 9–18. [Google Scholar] [CrossRef]
- Gervain, J.; Mehler, J.; Werker, J.F.; Nelson, C.A.; Csibra, G.; Lloyd-Fox, S.; Shukla, M.; Aslin, R.N. Near-Infrared Spectroscopy: A Report from the McDonnell Infant Methodology Consortium. Dev. Cogn. Neurosci. 2011, 1, 22–46. [Google Scholar] [CrossRef]
- Vanderwert, R.E.; Nelson, C.A. The Use of near-Infrared Spectroscopy in the Study of Typical and Atypical Development. Neuroimage 2014, 85 Pt 1, 264–271. [Google Scholar] [CrossRef]
- Doi, H.; Shinohara, K. fNIRS Studies on Hemispheric Asymmetry in Atypical Neural Function in Developmental Disorders. Front. Hum. Neurosci. 2017, 11, 137. [Google Scholar] [CrossRef]
- Zhang, F.; Roeyers, H. Exploring Brain Functions in Autism Spectrum Disorder: A Systematic Review on Functional near-Infrared Spectroscopy (fNIRS) Studies. Int. J. Psychophysiol. 2019, 137, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Conti, E.; Scaffei, E.; Bosetti, C.; Marchi, V.; Costanzo, V.; Dell’Oste, V.; Mazziotti, R.; Dell’Osso, L.; Carmassi, C.; Muratori, F.; et al. Looking for “fNIRS Signature” in Autism Spectrum: A Systematic Review Starting from Preschoolers. Front. Neurosci. 2022, 16, 785993. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Fox, S.; Blasi, A.; Elwell, C.E.; Charman, T.; Murphy, D.; Johnson, M.H. Reduced Neural Sensitivity to Social Stimuli in Infants at Risk for Autism. Proc. Biol. Sci. 2013, 280, 20123026. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Fox, S.; Blasi, A.; Pasco, G.; Gliga, T.; Jones, E.J.H.; Murphy, D.G.M.; Elwell, C.E.; Charman, T.; Johnson, M.H. BASIS Team Cortical Responses before 6 Months of Life Associate with Later Autism. Eur. J. Neurosci. 2018, 47, 736–749. [Google Scholar] [CrossRef]
- Edwards, L.A.; Wagner, J.B.; Tager-Flusberg, H.; Nelson, C.A. Differences in Neural Correlates of Speech Perception in 3 Month Olds at High and Low Risk for Autism Spectrum Disorder. J. Autism Dev. Disord. 2017, 47, 3125–3138. [Google Scholar] [CrossRef]
- Li, Y.; Jia, H.; Yu, D. Novel Analysis of fNIRS Acquired Dynamic Hemoglobin Concentrations: Application in Young Children with Autism Spectrum Disorder. Biomed. Opt. Express 2018, 9, 3694–3710. [Google Scholar] [CrossRef]
- Jia, H.; Li, Y.; Yu, D. Attenuation of Long-Range Temporal Correlations of Neuronal Oscillations in Young Children with Autism Spectrum Disorder. Neuroimage Clin. 2018, 20, 424–432. [Google Scholar] [CrossRef]
- Robertson, C.E.; Baron-Cohen, S. Sensory Perception in Autism. Nat. Rev. Neurosci. 2017, 18, 671–684. [Google Scholar] [CrossRef]
- Girault, J.B.; Donovan, K.; Hawks, Z.; Talovic, M.; Forsen, E.; Elison, J.T.; Shen, M.D.; Swanson, M.R.; Wolff, J.J.; Kim, S.H.; et al. Infant Visual Brain Development and Inherited Genetic Liability in Autism. Am. J. Psychiatry 2022, 179, 573–585. [Google Scholar] [CrossRef]
- Durand, S.; Patrizi, A.; Quast, K.B.; Hachigian, L.; Pavlyuk, R.; Saxena, A.; Carninci, P.; Hensch, T.K.; Fagiolini, M. NMDA Receptor Regulation Prevents Regression of Visual Cortical Function in the Absence of Mecp2. Neuron 2012, 76, 1078–1090. [Google Scholar] [CrossRef]
- de Freitas Dotto, P.; Cavascan, N.N.; Berezovsky, A.; Sacai, P.Y.; Rocha, D.M.; Pereira, J.M.; Salomão, S.R. Sweep Visually Evoked Potentials and Visual Findings in Children with West Syndrome. Eur. J. Paediatr. Neurol. 2014, 18, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Mazziotti, R.; Lupori, L.; Sagona, G.; Gennaro, M.; Della Sala, G.; Putignano, E.; Pizzorusso, T. Searching for Biomarkers of CDKL5 Disorder: Early-Onset Visual Impairment in CDKL5 Mutant Mice. Hum. Mol. Genet. 2017, 26, 2290–2298. [Google Scholar] [CrossRef] [PubMed]
- Felgerolle, C.; Hébert, B.; Ardourel, M.; Meyer-Dilhet, G.; Menuet, A.; Pinto-Morais, K.; Bizot, J.-C.; Pichon, J.; Briault, S.; Perche, O. Visual Behavior Impairments as an Aberrant Sensory Processing in the Mouse Model of Fragile X Syndrome. Front. Behav. Neurosci. 2019, 13, 228. [Google Scholar] [CrossRef] [PubMed]
- Mazziotti, R.; Cacciante, F.; Sagona, G.; Lupori, L.; Gennaro, M.; Putignano, E.; Alessandrì, M.G.; Ferrari, A.; Battini, R.; Cioni, G.; et al. Novel Translational Phenotypes and Biomarkers for Creatine Transporter Deficiency. Brain Commun. 2020, 2, fcaa089. [Google Scholar] [CrossRef] [PubMed]
- Mazziotti, R.; Scaffei, E.; Conti, E.; Marchi, V.; Rizzi, R.; Cioni, G.; Battini, R.; Baroncelli, L. The Amplitude of fNIRS Hemodynamic Response in the Visual Cortex Unmasks Autistic Traits in Typically Developing Children. Transl. Psychiatry 2022, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.; Rutter, M.; DiLavore, P.C.; Risi, S. Autism Diagnostic Observation Schedule–Generic; PsycTESTS Dataset: Lawrence, KS, USA, 2016. [Google Scholar]
- Ruta, L.; Mazzone, D.; Mazzone, L.; Wheelwright, S.; Baron-Cohen, S. The Autism-Spectrum Quotient—Italian Version: A Cross-Cultural Confirmation of the Broader Autism Phenotype. J. Autism Dev. Disord. 2012, 42, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S. Wechsler Preschool and Primary Scale of Intelligence; Springer: Cham, Switzerland, 2013. [Google Scholar]
- Sparrow, S.S.; Cicchetti, D.; Balla, D.A. Vineland Adaptive Behavior Scales, 2nd ed.; PsycTESTS Dataset: Lawrence, KS, USA, 2012. [Google Scholar]
- Peirce, J.; Gray, J.R.; Simpson, S.; MacAskill, M.; Höchenberger, R.; Sogo, H.; Kastman, E.; Lindeløv, J.K. PsychoPy2: Experiments in Behavior Made Easy. Behav. Res. Methods 2019, 51, 195–203. [Google Scholar] [CrossRef]
- Gollapudi, S. OpenCV with Python. In Learn Computer Vision Using OpenCV; Apress: Berkeley, CA, USA, 2019; pp. 31–50. [Google Scholar]
- Conti, E.; Calderoni, S.; Gaglianese, A.; Pannek, K.; Mazzotti, S.; Rose, S.; Scelfo, D.; Tosetti, M.; Muratori, F.; Cioni, G.; et al. Lateralization of Brain Networks and Clinical Severity in Toddlers with Autism Spectrum Disorder: A HARDI Diffusion MRI Study. Autism Res. 2016, 9, 382–392. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Mundorf, A.; Peterburs, J.; Ocklenburg, S. Asymmetry in the Central Nervous System: A Clinical Neuroscience Perspective. Front. Syst. Neurosci. 2021, 15, 733898. [Google Scholar] [CrossRef]
- Floris, D.L.; Wolfers, T.; Zabihi, M.; Holz, N.E.; Zwiers, M.P.; Charman, T.; Tillmann, J.; Ecker, C.; Dell’Acqua, F.; Banaschewski, T.; et al. Atypical Brain Asymmetry in Autism-A Candidate for Clinically Meaningful Stratification. Biol Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Postema, M.C.; van Rooij, D.; Anagnostou, E.; Arango, C.; Auzias, G.; Behrmann, M.; Filho, G.B.; Calderoni, S.; Calvo, R.; Daly, E.; et al. Altered Structural Brain Asymmetry in Autism Spectrum Disorder in a Study of 54 Datasets. Nat. Commun. 2019, 10, 4958. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-C.; Sandmann, P.; Thorne, J.D.; Herrmann, C.S.; Debener, S. Association of Concurrent fNIRS and EEG Signatures in Response to Auditory and Visual Stimuli. Brain Topogr. 2015, 28, 710–725. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Fox, S.; Blasi, A.; Elwell, C.E. Illuminating the Developing Brain: The Past, Present and Future of Functional near Infrared Spectroscopy. Neurosci. Biobehav. Rev. 2010, 34, 269–284. [Google Scholar] [CrossRef]
- Fox, S.E.; Wagner, J.B.; Shrock, C.L.; Tager-Flusberg, H.; Nelson, C.A. Neural Processing of Facial Identity and Emotion in Infants at High-Risk for Autism Spectrum Disorders. Front. Hum. Neurosci. 2013, 7, 89. [Google Scholar] [CrossRef]
- Bhat, A.N.; McDonald, N.M.; Eilbott, J.E.; Pelphrey, K.A. Exploring Cortical Activation and Connectivity in Infants with and without Familial Risk for Autism during Naturalistic Social Interactions: A Preliminary Study. Infant Behav. Dev. 2019, 57, 101337. [Google Scholar] [CrossRef]
- Kita, Y.; Gunji, A.; Inoue, Y.; Goto, T.; Sakihara, K.; Kaga, M.; Inagaki, M.; Hosokawa, T. Self-Face Recognition in Children with Autism Spectrum Disorders: A near-Infrared Spectroscopy Study. Brain Dev. 2011, 33, 494–503. [Google Scholar] [CrossRef]
- Kovarski, K.; Mennella, R.; Wong, S.M.; Dunkley, B.T.; Taylor, M.J.; Batty, M. Enhanced Early Visual Responses During Implicit Emotional Faces Processing in Autism Spectrum Disorder. J. Autism Dev. Disord. 2019, 49, 871–886. [Google Scholar] [CrossRef]
- Lewis, J.D.; Evans, A.C.; Pruett, J.R., Jr.; Botteron, K.N.; McKinstry, R.C.; Zwaigenbaum, L.; Estes, A.M.; Collins, D.L.; Kostopoulos, P.; Gerig, G.; et al. The Emergence of Network Inefficiencies in Infants With Autism Spectrum Disorder. Biol. Psychiatry 2017, 82, 176–185. [Google Scholar] [CrossRef]
- Lockwood Estrin, G.; Milner, V.; Spain, D.; Happé, F.; Colvert, E. Barriers to Autism Spectrum Disorder Diagnosis for Young Women and Girls: A Systematic Review. Rev. J. Autism Dev. Disord. 2021, 8, 454–470. [Google Scholar] [CrossRef]
- D’Mello, A.M.; Frosch, I.R.; Li, C.E.; Cardinaux, A.L.; Gabrieli, J.D.E. Exclusion of Females in Autism Research: Empirical Evidence for a “Leaky” Recruitment-to-Research Pipeline. Autism Res. 2022, 15, 1929–1940. [Google Scholar] [CrossRef]
- Peng, C.; Hou, X. Applications of Functional near-Infrared Spectroscopy (fNIRS) in Neonates. Neurosci. Res. 2021, 170, 18–23. [Google Scholar] [CrossRef]
- Van der Hallen, R.; Evers, K.; Brewaeys, K.; Van den Noortgate, W.; Wagemans, J. Global Processing Takes Time: A Meta-Analysis on Local-Global Visual Processing in ASD. Psychol. Bull. 2015, 141, 549–573. [Google Scholar] [CrossRef]
- Apicella, F.; Costanzo, V.; Purpura, G. Are Early Visual Behavior Impairments Involved in the Onset of Autism Spectrum Disorders? Insights for Early Diagnosis and Intervention. Eur. J. Pediatr. 2020, 179, 225–234. [Google Scholar] [CrossRef]
- Spiteri, S.; Crewther, D. Neural Mechanisms of Visual Motion Anomalies in Autism: A Two-Decade Update and Novel Aetiology. Front. Neurosci. 2021, 15, 756841. [Google Scholar] [CrossRef]
- Jung, M.; Tu, Y.; Lang, C.A.; Ortiz, A.; Park, J.; Jorgenson, K.; Kong, X.-J.; Kong, J. Decreased Structural Connectivity and Resting-State Brain Activity in the Lateral Occipital Cortex Is Associated with Social Communication Deficits in Boys with Autism Spectrum Disorder. Neuroimage 2019, 190, 205–212. [Google Scholar] [CrossRef]
- Corballis, P.M. Visuospatial Processing and the Right-Hemisphere Interpreter. Brain Cogn. 2003, 53, 171–176. [Google Scholar] [CrossRef]
- Jonas, J.; Frismand, S.; Vignal, J.-P.; Colnat-Coulbois, S.; Koessler, L.; Vespignani, H.; Rossion, B.; Maillard, L. Right Hemispheric Dominance of Visual Phenomena Evoked by Intracerebral Stimulation of the Human Visual Cortex. Hum. Brain Mapp. 2014, 35, 3360–3371. [Google Scholar] [CrossRef]
- Hougaard, A.; Jensen, B.H.; Amin, F.M.; Rostrup, E.; Hoffmann, M.B.; Ashina, M. Cerebral Asymmetry of fMRI-BOLD Responses to Visual Stimulation. PLoS ONE 2015, 10, e0126477. [Google Scholar] [CrossRef]
- Robertson, C.E.; Kravitz, D.J.; Freyberg, J.; Baron-Cohen, S.; Baker, C.I. Slower Rate of Binocular Rivalry in Autism. J. Neurosci. 2013, 33, 16983–16991. [Google Scholar] [CrossRef]
- Robertson, C.E.; Ratai, E.-M.; Kanwisher, N. Reduced GABAergic Action in the Autistic Brain. Curr. Biol. 2016, 26, 80–85. [Google Scholar] [CrossRef]
- Genç, E.; Bergmann, J.; Tong, F.; Blake, R.; Singer, W.; Kohler, A. Callosal Connections of Primary Visual Cortex Predict the Spatial Spreading of Binocular Rivalry across the Visual Hemifields. Front. Hum. Neurosci. 2011, 5, 161. [Google Scholar] [CrossRef]
- Reynolds, J.H.; Chelazzi, L. Attentional Modulation of Visual Processing. Annu. Rev. Neurosci. 2004, 27, 611–647. [Google Scholar] [CrossRef]
- Speed, A.; Del Rosario, J.; Mikail, N.; Haider, B. Spatial Attention Enhances Network, Cellular and Subthreshold Responses in Mouse Visual Cortex. Nat. Commun. 2020, 11, 505. [Google Scholar] [CrossRef]
- Bartolomeo, P.; Seidel Malkinson, T. Hemispheric Lateralization of Attention Processes in the Human Brain. Curr. Opin. Psychol. 2019, 29, 90–96. [Google Scholar] [CrossRef]
- Gotham, K.; Pickles, A.; Lord, C. Standardizing ADOS Scores for a Measure of Severity in Autism Spectrum Disorders. J. Autism Dev. Disord. 2009, 39, 693–705. [Google Scholar] [CrossRef]
- Alvares, G.A.; Bebbington, K.; Cleary, D.; Evans, K.; Glasson, E.J.; Maybery, M.T.; Pillar, S.; Uljarević, M.; Varcin, K.; Wray, J.; et al. The Misnomer of “High Functioning Autism”: Intelligence Is an Imprecise Predictor of Functional Abilities at Diagnosis. Autism 2020, 24, 221–232. [Google Scholar] [CrossRef]
- Saulnier, C.A.; Klaiman, C.; McQueen, E. Adaptive Behavior Profiles in Autism Spectrum Disorder. Curr. Psychiatry Rep. 2022, 24, 749–756. [Google Scholar] [CrossRef]
- Corbett, B.A.; Schwartzman, J.M.; Libsack, E.J.; Muscatello, R.A.; Lerner, M.D.; Simmons, G.L.; White, S.W. Camouflaging in Autism: Examining Sex-Based and Compensatory Models in Social Cognition and Communication. Autism Res. 2021, 14, 127–142. [Google Scholar] [CrossRef]
- Neuhaus, E.; Lowry, S.J.; Santhosh, M.; Kresse, A.; Edwards, L.A.; Keller, J.; Libsack, E.J.; Kang, V.Y.; Naples, A.; Jack, A.; et al. Resting State EEG in Youth with ASD: Age, Sex, and Relation to Phenotype. J. Neurodev. Disord. 2021, 13, 33. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Age | Gender | Head | Cap Size | Cartoon Selected |
|---|---|---|---|---|---|
| A1 | 4 | F | 49 | 52 | Frozen Walt Disney Pictures, 2013 |
| A2 | 5 | F | 50.5 | 52 | The Little Mermaid Walt Disney Pictures, 1989 |
| A3 | 6 | F | 53 | 54 | The Chipmunks, S7E04 “Bye, George” MWS and DIC Entertainment |
| A4 | 5 | F | 50.5 | 52 | The Lion King Walt Disney Pictures,1994 |
| A5 | 4 | F | 50.5 | 52 | The Chipmunks, S7E04 “Bye, George” MWS and DIC Entertainment |
| A6 | 6 | F | 53 | 54 | SpongeBob, S12E01 “FarmerBob” Nickelodeon Animation |
| A7 | 3 | F | 51 | 52 | The Little Mermaid Walt Disney Pictures, 1989 |
| A8 | 4 | F | 49.5 | 52 | Frozen Walt Disney Pictures, 2013 |
| A9 | 6 | F | 53 | 54 | Inside out Disney-Pixar, 2015 |
| A10 | 6 | F | 53 | 54 | The Little Mermaid Walt Disney Pictures, 1989 |
| A11 | 4 | F | 41.5 | 54 | Frozen Walt Disney Pictures, 2013 |
| A12 | 6 | F | 50.5 | 54 | Frozen Walt Disney Pictures, 2013 |
| A13 | 3 | F | 50 | 52 | The Aristocats Walt Disney Productions, 1970 |
| ID | Age | Gender | Head | Cap Size | Cartoon | ADOS_TOT | ADOS_comp | AQ_tot | nv_IQ | Cognitive Scale | VABS_tot |
|---|---|---|---|---|---|---|---|---|---|---|---|
| B1 | 4 | F | 51 | 52 | Frozen Walt Disney Pictures, 2013 | 16 | 7 | 53 | 96 | GMDS III | 63 |
| B2 | 5 | F | 50.5 | 52 | Frozen Walt Disney Pictures, 2013 | n.a. | n.a. | 56 | 78 | Leiter III | 74 |
| B3 | 6 | F | 53 | 54 | Bing, S1E06 “Smoothie” Tandem Films e Digitales Studios, 2014 | 14 | 5 | 38 | 98 | WPPSI III | 80 |
| B4 | 4 | F | 51 | 52 | Frozen Walt Disney Pictures, 2013 | 9 | 5 | n.a. | 125 | Leiter III | 96 |
| B5 | 3 | F | 48.5 | 52 | Bing, S1E06 “Smoothie” Tandem Films e Digitales Studios, 2014 | 14 | 5 | 48 | 106 | WPPSI III | 93 |
| B6 | 6 | F | 55 | 56 | Uncle Grandpa, S3E04 “Uncle Easter” Cartoon Network Studios, 2016 | 12 | 7 | 58 | 108 | WISC IV | 81 |
| B7 | 5 | F | 50 | 52 | The Chipmunks, S7E04 “Bye, George” MWS and DIC Entertainment | 9 | 6 | 96 | 87 | GMDS III | 87 |
| B8 | 6 | F | 52 | 52 | Frozen Walt Disney Pictures, 2013 | 11 | 6 | 106 | 117 | WPPSI III | 85 |
| B9 | 5 | F | 51.5 | 52 | Bing, S1E06 “Smoothie” Tandem Films e Digitales Studios, 2014 | 9 | 5 | 53 | 102 | WPPSI III | 85 |
| B10 | 4 | F | 52 | 52 | Frozen Walt Disney Pictures, 2013 | 15 | 5 | 47 | 98 | GMDS III | n.a. |
| B11 | 3 | F | 53 | 52 | Curious George S1E03 “Zeros to Donuts”, Universal Animation Studios, 2006 | 8 | 4 | 33 | 119 | WPPSI III | 102 |
| B12 | 3 | F | 48.5 | 52 | Masha and the Bear S1E26 “Home” Improvement, Animaccord Animation Studio | 11 | 5 | 95 | 110 | Leiter III | 77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scaffei, E.; Mazziotti, R.; Conti, E.; Costanzo, V.; Calderoni, S.; Stoccoro, A.; Carmassi, C.; Tancredi, R.; Baroncelli, L.; Battini, R. A Potential Biomarker of Brain Activity in Autism Spectrum Disorders: A Pilot fNIRS Study in Female Preschoolers. Brain Sci. 2023, 13, 951. https://doi.org/10.3390/brainsci13060951
Scaffei E, Mazziotti R, Conti E, Costanzo V, Calderoni S, Stoccoro A, Carmassi C, Tancredi R, Baroncelli L, Battini R. A Potential Biomarker of Brain Activity in Autism Spectrum Disorders: A Pilot fNIRS Study in Female Preschoolers. Brain Sciences. 2023; 13(6):951. https://doi.org/10.3390/brainsci13060951
Chicago/Turabian StyleScaffei, Elena, Raffaele Mazziotti, Eugenia Conti, Valeria Costanzo, Sara Calderoni, Andrea Stoccoro, Claudia Carmassi, Raffaella Tancredi, Laura Baroncelli, and Roberta Battini. 2023. "A Potential Biomarker of Brain Activity in Autism Spectrum Disorders: A Pilot fNIRS Study in Female Preschoolers" Brain Sciences 13, no. 6: 951. https://doi.org/10.3390/brainsci13060951
APA StyleScaffei, E., Mazziotti, R., Conti, E., Costanzo, V., Calderoni, S., Stoccoro, A., Carmassi, C., Tancredi, R., Baroncelli, L., & Battini, R. (2023). A Potential Biomarker of Brain Activity in Autism Spectrum Disorders: A Pilot fNIRS Study in Female Preschoolers. Brain Sciences, 13(6), 951. https://doi.org/10.3390/brainsci13060951