Reduced Resting-State EEG Power Spectra and Functional Connectivity after 24 and 36 Hours of Sleep Deprivation

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Procedures
2.3. Data Collection and Processing
2.4. Data Analysis
2.4.1. Power Spectral Analysis
2.4.2. Source Localization Analysis
2.4.3. EEG Source Connectivity Analysis
3. Results
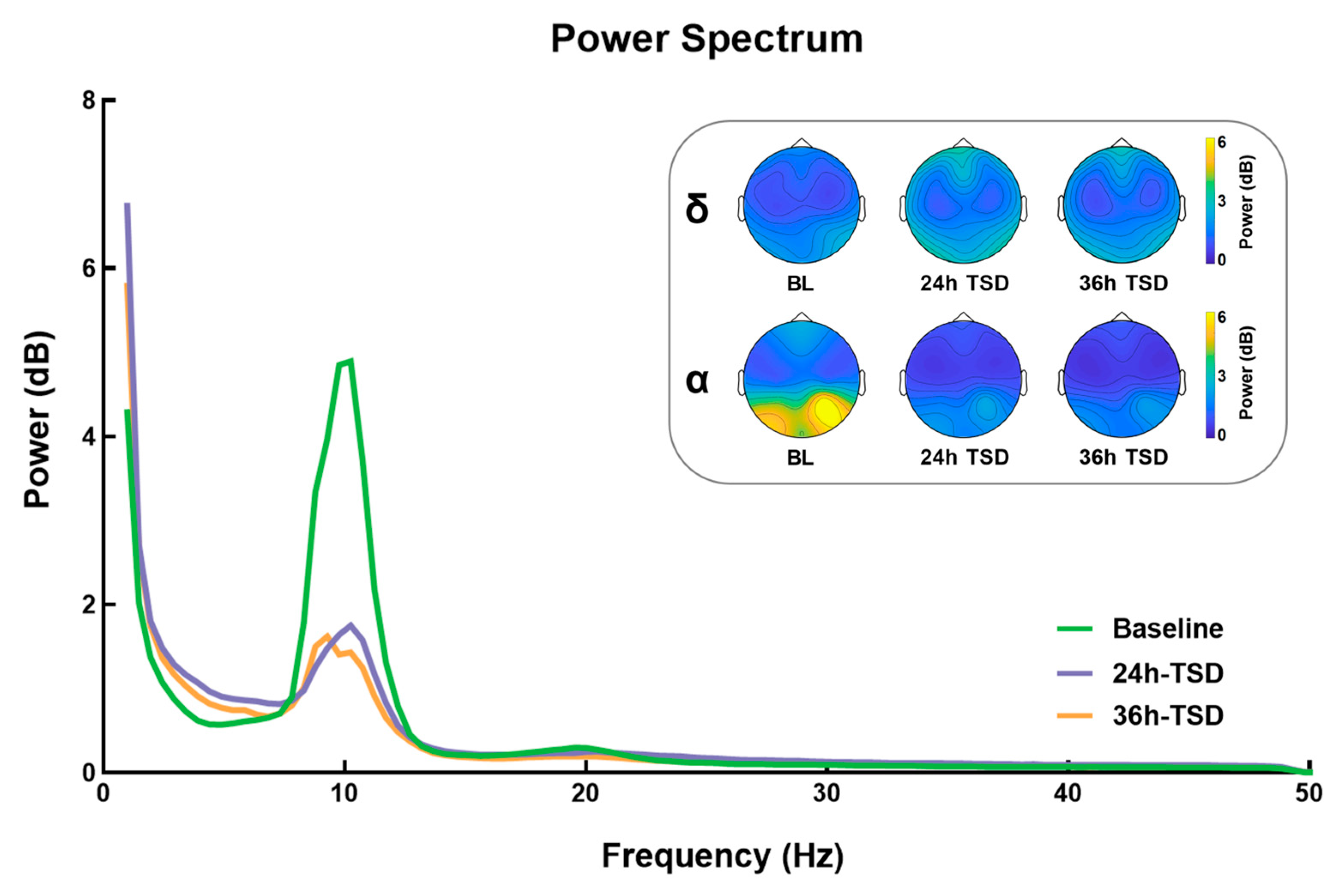
3.1. Power Spectrum Results
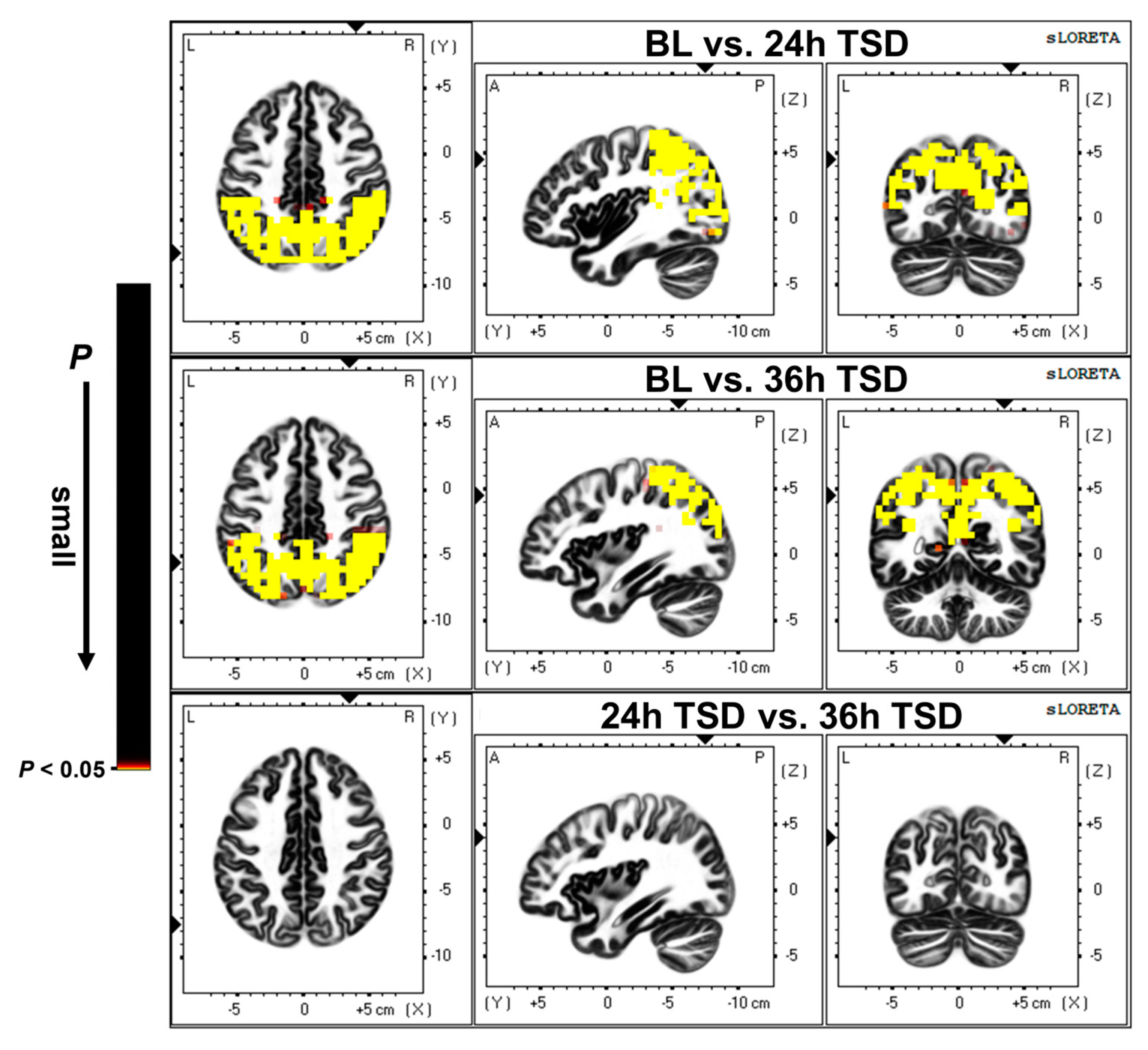
3.2. Source Location Results
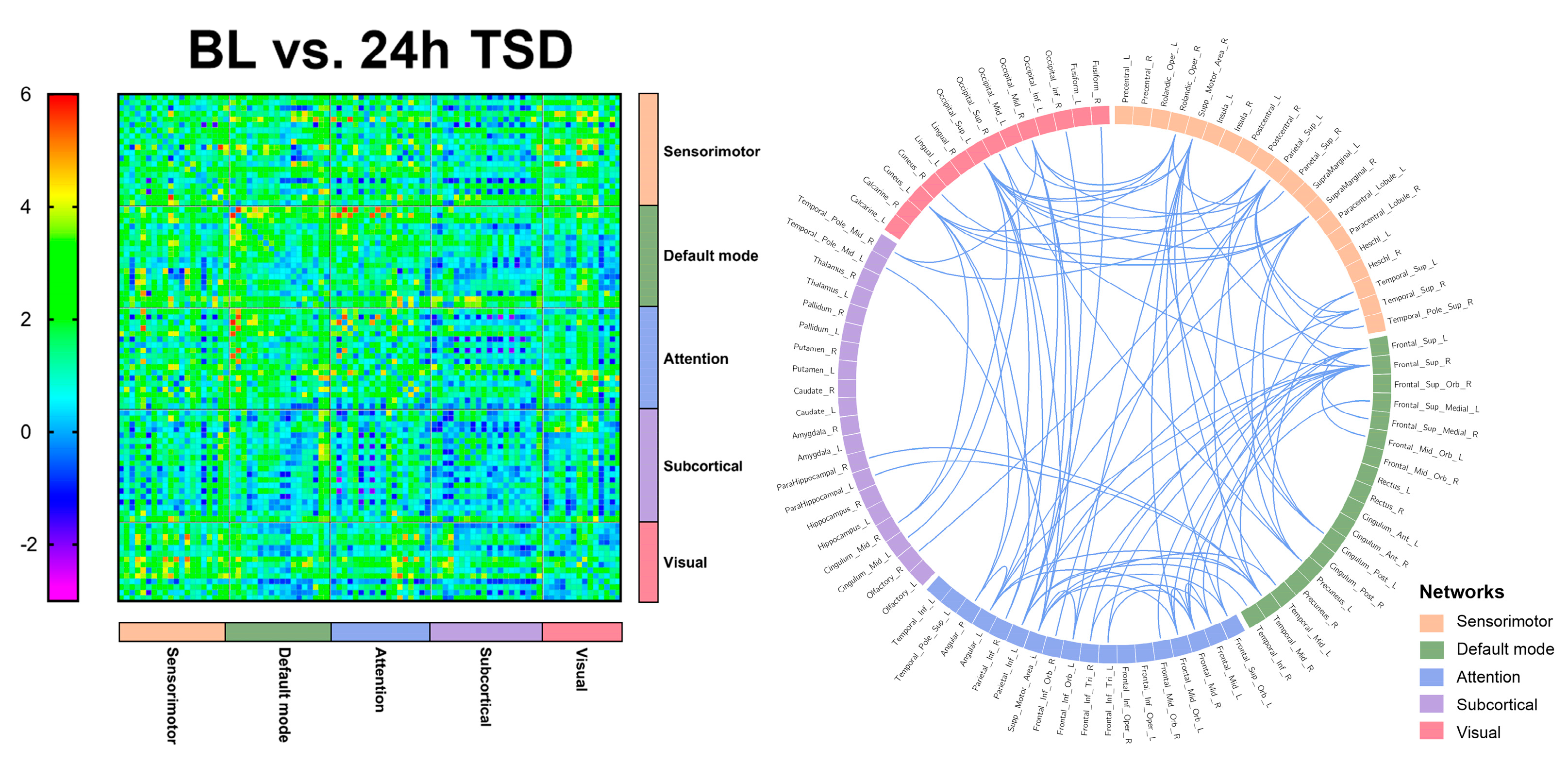
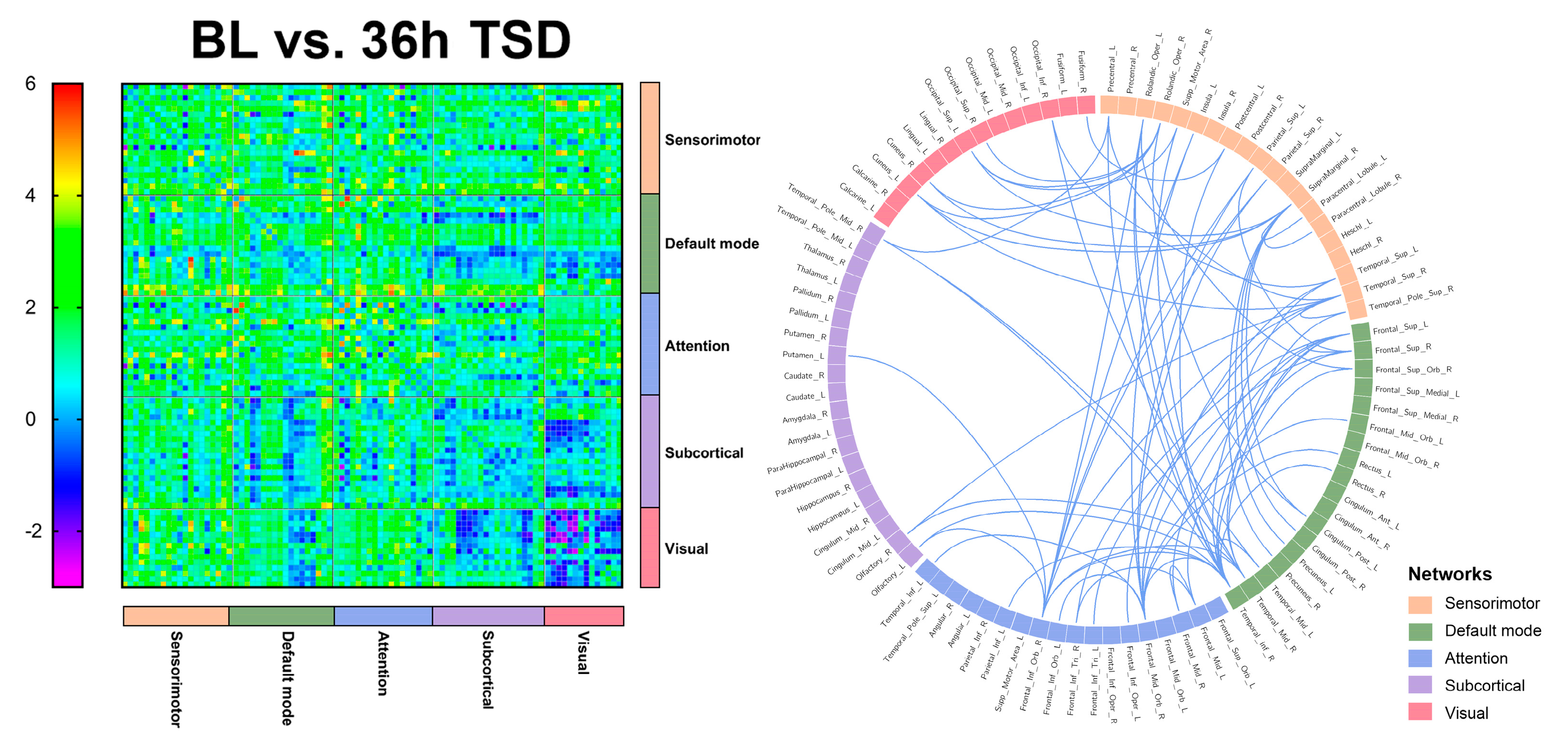
3.3. EEG Source Connectivity Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reynolds, A.C.; Banks, S. Total sleep deprivation, chronic sleep restriction and sleep disruption. Prog. Brain Res. 2010, 185, 91–103. [Google Scholar] [PubMed]
- Hudson, A.N.; Van Dongen, H.P.A.; Honn, K.A. Sleep deprivation, vigilant attention, and brain function: A review. Neuropsychopharmacology 2020, 45, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Arnal, P.J.; Sauvet, F.; Leger, D.; van Beers, P.; Bayon, V.; Bougard, C.; Rabat, A.; Millet, G.Y.; Chennaoui, M. Benefits of Sleep Extension on Sustained Attention and Sleep Pressure Before and During Total Sleep Deprivation and Recovery. Sleep 2015, 38, 1935–1943. [Google Scholar] [CrossRef] [PubMed]
- Krause, A.J.; Simon, E.B.; Mander, B.A.; Greer, S.M.; Saletin, J.M.; Goldstein-Piekarski, A.N.; Walker, M.P. The sleep-deprived human brain. Nat. Rev. Neurosci. 2017, 18, 404–418. [Google Scholar] [CrossRef]
- Kusztor, A.; Raud, L.; Juel, B.E.; Nilsen, A.S.; Storm, J.F.; Huster, R.J. Sleep deprivation differentially affects subcomponents of cognitive control. Sleep 2019, 42, zsz016. [Google Scholar] [CrossRef]
- Tempesta, D.; Socci, V.; De Gennaro, L.; Ferrara, M. Sleep and emotional processing. Sleep Med. Rev. 2018, 40, 183–195. [Google Scholar] [CrossRef]
- Killgore, W.D.; Balkin, T.J.; Wesensten, N.J. Impaired decision making following 49 h of sleep deprivation. J. Sleep Res. 2006, 15, 7–13. [Google Scholar] [CrossRef]
- Grèzes, J.; Erblang, M.; Vilarem, E.; Quiquempoix, M.; Van Beers, P.; Guillard, M.; Sauvet, F.; Mennella, R.; Rabat, A. Impact of total sleep deprivation and related mood changes on approach-avoidance decisions to threat-related facial displays. Sleep 2021, 44, zsab186. [Google Scholar] [CrossRef]
- Chu, C.; Holst, S.C.; Elmenhorst, E.M.; Foerges, A.L.; Li, C.; Lange, D.; Hennecke, E.; Baur, D.M.; Beer, S.; Hoffstaedter, F.; et al. Total Sleep Deprivation Increases Brain Age Prediction Reversibly in Multisite Samples of Young Healthy Adults. J. Neurosci. 2023, 43, 2168–2177. [Google Scholar] [CrossRef]
- Strijkstra, A.M.; Beersma, D.G.; Drayer, B.; Halbesma, N.; Daan, S. Subjective sleepiness correlates negatively with global alpha (8-12 Hz) and positively with central frontal theta (4-8 Hz) frequencies in the human resting awake electroencephalogram. Neurosci. Lett. 2003, 340, 17–20. [Google Scholar] [CrossRef]
- Corsi-Cabrera, M.; Sánchez, A.I.; del-Río-Portilla, Y.; Villanueva, Y.; Pérez-Garci, E. Effect of 38 h of total sleep deprivation on the waking EEG in women: Sex differences. Int. J. Psychophysiol. 2003, 50, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Hayashi, M.; Hori, T. Topographical characteristics and principal component structure of the hypnagogic EEG. Sleep 1997, 20, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.J.; Czeisler, C.A. Contribution of the circadian pacemaker and the sleep homeostat to sleep propensity, sleep structure, electroencephalographic slow waves, and sleep spindle activity in humans. J. Neurosci. 1995, 15, 3526–3538. [Google Scholar] [CrossRef]
- Gorgoni, M.; Ferlazzo, F.; Ferrara, M.; Moroni, F.; D’Atri, A.; Fanelli, S.; Gizzi Torriglia, I.; Lauri, G.; Marzano, C.; Rossini, P.M.; et al. Topographic electroencephalogram changes associated with psychomotor vigilance task performance after sleep deprivation. Sleep Med. 2014, 15, 1132–1139. [Google Scholar] [CrossRef]
- De Gennaro, L.; Marzano, C.; Veniero, D.; Moroni, F.; Fratello, F.; Curcio, G.; Ferrara, M.; Ferlazzo, F.; Novelli, L.; Concetta Pellicciari, M.; et al. Neurophysiological correlates of sleepiness: A combined TMS and EEG study. NeuroImage 2007, 36, 1277–1287. [Google Scholar] [CrossRef]
- Finelli, L.A.; Baumann, H.; Borbély, A.A.; Achermann, P. Dual electroencephalogram markers of human sleep homeostasis: Correlation between theta activity in waking and slow-wave activity in sleep. Neuroscience 2000, 101, 523–529. [Google Scholar] [CrossRef]
- Tinguely, G.; Finelli, L.A.; Landolt, H.P.; Borbély, A.A.; Achermann, P. Functional EEG topography in sleep and waking: State-dependent and state-independent features. NeuroImage 2006, 32, 283–292. [Google Scholar] [CrossRef]
- Ferreira, C.; Deslandes, A.; Moraes, H.; Cagy, M.; Pompeu, F.; Basile, L.F.; Piedade, R.; Ribeiro, P. Electroencephalographic changes after one night of sleep deprivation. Arq. de Neuro-Psiquiatr. 2006, 64, 388–393. [Google Scholar] [CrossRef]
- Miraglia, F.; Tomino, C.; Vecchio, F.; Gorgoni, M.; De Gennaro, L.; Rossini, P.M. The brain network organization during sleep onset after deprivation. Clin. Neurophysiol. 2021, 132, 36–44. [Google Scholar] [CrossRef]
- Knoblauch, V.; Kräuchi, K.; Renz, C.; Wirz-Justice, A.; Cajochen, C. Homeostatic control of slow-wave and spindle frequency activity during human sleep: Effect of differential sleep pressure and brain topography. Cereb. Cortex 2002, 12, 1092–1100. [Google Scholar] [CrossRef]
- Curcio, G.; Ferrara, M.; Pellicciari, M.C.; Cristiani, R.; De Gennaro, L. Effect of total sleep deprivation on the landmarks of stage 2 sleep. Clin. Neurophysiol. 2003, 114, 2279–2285. [Google Scholar] [CrossRef] [PubMed]
- De Gennaro, L.; Ferrara, M. Sleep spindles: An overview. Sleep Med. Rev. 2003, 7, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Verweij, I.M.; Romeijn, N.; Smit, D.J.; Piantoni, G.; Van Someren, E.J.; van der Werf, Y.D. Sleep deprivation leads to a loss of functional connectivity in frontal brain regions. BMC Neurosci. 2014, 15, 88. [Google Scholar] [CrossRef] [PubMed]
- Kar, S.; Routray, A.; Nayak, B.P. Functional network changes associated with sleep deprivation and fatigue during simulated driving: Validation using blood biomarkers. Clin. Neurophysiol. 2011, 122, 966–974. [Google Scholar] [CrossRef]
- Koenis, M.M.; Romeijn, N.; Piantoni, G.; Verweij, I.; Van der Werf, Y.D.; Van Someren, E.J.; Stam, C.J. Does sleep restore the topology of functional brain networks? Hum. Brain Mapp. 2013, 34, 487–500. [Google Scholar] [CrossRef]
- Lim, J.; Tan, J.C.; Parimal, S.; Dinges, D.F.; Chee, M.W. Sleep deprivation impairs object-selective attention: A view from the ventral visual cortex. PLoS ONE. 2010, 5, e9087. [Google Scholar] [CrossRef]
- Yoo, S.S.; Gujar, N.; Hu, P.; Jolesz, F.A.; Walker, M.P. The human emotional brain without sleep—A prefrontal amygdala disconnect. Curr. Biol. 2007, 17, R877–R878. [Google Scholar] [CrossRef]
- Sämann, P.G.; Tully, C.; Spoormaker, V.I.; Wetter, T.C.; Holsboer, F.; Wehrle, R.; Czisch, M. Increased sleep pressure reduces resting state functional connectivity. Magma 2010, 23, 375–389. [Google Scholar] [CrossRef]
- De Havas, J.A.; Parimal, S.; Soon, C.S.; Chee, M.W. Sleep deprivation reduces default mode network connectivity and anti-correlation during rest and task performance. NeuroImage 2012, 59, 1745–1751. [Google Scholar] [CrossRef]
- Liu, H.; Li, H.; Wang, Y.; Lei, X. Enhanced brain small-worldness after sleep deprivation: A compensatory effect. J. Sleep Res. 2014, 23, 554–563. [Google Scholar] [CrossRef]
- Skurvydas, A.; Kazlauskaite, D.; Zlibinaite, L.; Cekanauskaite, A.; Valanciene, D.; Karanauskiene, D.; Zuoziene, I.J.; Majauskiene, D.; Mickeviciene, D.; Satas, A. Effects of two nights of sleep deprivation on executive function and central and peripheral fatigue during maximal voluntary contraction lasting 60s. Physiol. Behav. 2021, 229, 113226. [Google Scholar] [CrossRef]
- Stenson, A.R.; Whitney, P.; Hinson, J.M.; Hansen, D.A.; Lawrence-Sidebottom, D.; Skeiky, L.; Riedy, S.M.; Kurinec, C.A.; Van Dongen, H.P.A. Effects of total sleep deprivation on components of top-down attentional control using a flexible attentional control task. J. Sleep Res. 2023, 32, e13744. [Google Scholar] [CrossRef] [PubMed]
- Chua, E.C.; Fang, E.; Gooley, J.J. Effects of total sleep deprivation on divided attention performance. PLoS ONE 2017, 12, e0187098. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shao, Y.; Jin, X.; Cai, X.; Du, F. Decreased effective connectivity between insula and anterior cingulate cortex during a working memory task after prolonged sleep deprivation. Behav. Brain Res. 2021, 409, 113263. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, A.; De Koninck, J.; Campbell, K.B. Disentangling specific inhibitory versus general decision-making processes during sleep deprivation using a Go/NoGo ERP paradigm. Int. J. Psychophysiol. 2019, 141, 18–27. [Google Scholar] [CrossRef]
- Chee, M.W.; Chuah, L.Y.; Venkatraman, V.; Chan, W.Y.; Philip, P.; Dinges, D.F. Functional imaging of working memory following normal sleep and after 24 and 35 h of sleep deprivation: Correlations of fronto-parietal activation with performance. NeuroImage 2006, 31, 419–428. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H., 3rd; Berman, S.R.; Kupfer, D.J. The Pittsburgh Sleep Quality Index: A new instrument for psychiatric practice and research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Corsi-Cabrera, M.; Galindo-Vilchis, L.; del-Río-Portilla, Y.; Arce, C.; Ramos-Loyo, J. Within-subject reliability and inter-session stability of EEG power and coherent activity in women evaluated monthly over nine months. Clin. Neurophysiol. 2007, 118, 9–21. [Google Scholar] [CrossRef]
- Duan, W.; Chen, X.; Wang, Y.J.; Zhao, W.; Yuan, H.; Lei, X. Reproducibility of power spectrum, functional connectivity and network construction in resting-state EEG. J. Neurosci. Methods 2021, 348, 108985. [Google Scholar] [CrossRef] [PubMed]
- Shenfield, L.; Beanland, V.; Filtness, A.; Apthorp, D. The impact of sleep loss on sustained and transient attention: An EEG study. Peer J. 2020, 8, e8960. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.P.; Makeig, S.; Humphries, C.; Lee, T.W.; McKeown, M.J.; Iragui, V.; Sejnowski, T.J. Removing electroencephalographic artifacts by blind source separation. Psychophysiology 2000, 37, 163–178. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods Find. Exp. Clin. Pharmacol. 2002, 24, 5–12. [Google Scholar]
- Mazziotta, J.; Toga, A.; Evans, A.; Fox, P.; Lancaster, J.; Zilles, K.; Woods, R.; Paus, T.; Simpson, G.; Pike, B.; et al. A probabilistic atlas and reference system for the human brain: International Consortium for Brain Mapping (ICBM). Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2001, 356, 1293–1322. [Google Scholar] [CrossRef]
- Holmes, A.P.; Blair, R.C.; Watson, J.D.; Ford, I. Nonparametric analysis of statistic images from functional mapping experiments. J. Cereb. Blood Flow Metab. 1996, 16, 7–22. [Google Scholar] [CrossRef]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.M. FieldTrip: Open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2011, 2011, 156869. [Google Scholar] [CrossRef]
- Schoffelen, J.M.; Gross, J. Source connectivity analysis with MEG and EEG. Hum. Brain Mapp. 2009, 30, 1857–1865. [Google Scholar] [CrossRef]
- Zhuo, Z.; Mo, X.; Ma, X.; Han, Y.; Li, H. Identifying aMCI with functional connectivity network characteristics based on subtle AAL atlas. Brain Res. 2018, 1696, 81–90. [Google Scholar] [CrossRef]
- van Diessen, E.; Numan, T.; van Dellen, E.; van der Kooi, A.W.; Boersma, M.; Hofman, D.; van Lutterveld, R.; van Dijk, B.W.; van Straaten, E.C.; Hillebrand, A.; et al. Opportunities and methodological challenges in EEG and MEG resting state functional brain network research. Clin. Neurophysiol. 2015, 126, 1468–1481. [Google Scholar] [CrossRef]
- Zalesky, A.; Fornito, A.; Bullmore, E.T. Network-based statistic: Identifying differences in brain networks. NeuroImage 2010, 53, 1197–1207. [Google Scholar] [CrossRef]
- Winkler, A.M.; Ridgway, G.R.; Webster, M.A.; Smith, S.M.; Nichols, T.E. Permutation inference for the general linear model. NeuroImage 2014, 92, 381–397. [Google Scholar] [CrossRef] [PubMed]
- Jespersen, K.V.; Stevner, A.; Fernandes, H.; Sørensen, S.D.; Van Someren, E.; Kringelbach, M.; Vuust, P. Reduced structural connectivity in Insomnia Disorder. J. Sleep Res. 2020, 29, e12901. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Drummond, S.P.; Brown, G.G.; Gillin, J.C.; Stricker, J.L.; Wong, E.C.; Buxton, R.B. Altered brain response to verbal learning following sleep deprivation. Nature 2000, 403, 655–657. [Google Scholar] [CrossRef]
- Robinson, J.L.; Erath, S.A.; Kana, R.K.; El-Sheikh, M. Neurophysiological differences in the adolescent brain following a single night of restricted sleep—A 7T fMRI study. Dev. Cogn. Neurosci. 2018, 31, 1–10. [Google Scholar] [CrossRef]
- Garrity, A.G.; Pearlson, G.D.; McKiernan, K.; Lloyd, D.; Kiehl, K.A.; Calhoun, V.D. Aberrant "default mode" functional connectivity in schizophrenia. Am. J. Psychiatry 2007, 164, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The brain’s default network: Anatomy, function, and relevance to disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef]
- Gusnard, D.A.; Raichle, M.E.; Raichle, M.E. Searching for a baseline: Functional imaging and the resting human brain. Nat. Rev. Neurosci. 2001, 2, 685–694. [Google Scholar] [CrossRef]
- Bai, F.; Zhang, Z.; Yu, H.; Shi, Y.; Yuan, Y.; Zhu, W.; Zhang, X.; Qian, Y. Default-mode network activity distinguishes amnestic type mild cognitive impairment from healthy aging: A combined structural and resting-state functional MRI study. Neurosci. Lett. 2008, 438, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; van Veen, V. Anterior cingulate cortex and conflict detection: An update of theory and data. Cogn. Affect. Behav. Neurosci. 2007, 7, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Weiss, P.H.; Marshall, J.C.; Zilles, K.; Fink, G.R. Are action and perception in near and far space additive or interactive factors? NeuroImage 2003, 18, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Corbetta, M.; Miezin, F.M.; Shulman, G.L.; Petersen, S.E. A PET study of visuospatial attention. J. Neurosci. 1993, 13, 1202–1226. [Google Scholar] [CrossRef]
- Corbetta, M.; Shulman, G.L.; Miezin, F.M.; Petersen, S.E. Superior parietal cortex activation during spatial attention shifts and visual feature conjunction. Science 1995, 270, 802–805. [Google Scholar] [CrossRef]
- Culham, J.C.; Valyear, K.F. Human parietal cortex in action. Curr. Res. Neurobiol. 2006, 16, 205–212. [Google Scholar] [CrossRef]
- Iacoboni, M. Visuo-motor integration and control in the human posterior parietal cortex: Evidence from TMS and fMRI. Neuropsychologia 2006, 44, 2691–2699. [Google Scholar] [CrossRef]
- Federico, G.; Reynaud, E.; Navarro, J.; Lesourd, M.; Gaujoux, V.; Lamberton, F.; Ibarrola, D.; Cavaliere, C.; Alfano, V.; Aiello, M.; et al. The cortical thickness of the area PF of the left inferior parietal cortex mediates technical-reasoning skills. Sci. Rep. 2022, 12, 11840. [Google Scholar] [CrossRef]
- Del Angel, J.; Cortez, J.; Juárez, D.; Guerrero, M.; García, A.; Ramírez, C.; Valdez, P. Effects of sleep reduction on the phonological and visuospatial components of working memory. Sleep Sci. 2015, 8, 68–74. [Google Scholar] [CrossRef]
- Thomas, M.; Sing, H.; Belenky, G.; Holcomb, H.; Mayberg, H.; Dannals, R.; Wagner, H.; Thorne, D.; Popp, K.; Rowland, L.; et al. Neural basis of alertness and cognitive performance impairments during sleepiness. I. Effects of 24 h of sleep deprivation on waking human regional brain activity. J. Sleep Res. 2000, 9, 335–352. [Google Scholar] [CrossRef]
- Adolphs, R. Cognitive neuroscience of human social behaviour. Nature reviews. Neuroscience 2003, 4, 165–178. [Google Scholar]
- Nir, Y.; Andrillon, T.; Marmelshtein, A.; Suthana, N.; Cirelli, C.; Tononi, G.; Fried, I. Selective neuronal lapses precede human cognitive lapses following sleep deprivation. Nat. Med. 2017, 23, 1474–1480. [Google Scholar] [CrossRef]
- Bell-McGinty, S.; Habeck, C.; Hilton, H.J.; Rakitin, B.; Scarmeas, N.; Zarahn, E.; Flynn, J.; DeLaPaz, R.; Basner, R.; Stern, Y. Identification and differential vulnerability of a neural network in sleep deprivation. Cereb. Cortex 2004, 14, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Javaheripour, N.; Shahdipour, N.; Noori, K.; Zarei, M.; Camilleri, J.A.; Laird, A.R.; Fox, P.T.; Eickhoff, S.B.; Eickhoff, C.R.; Rosenzweig, I.; et al. Functional brain alterations in acute sleep deprivation: An activation likelihood estimation meta-analysis. Sleep Med. Rev. 2019, 46, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Federico, G.; Alfano, V.; Garramone, F.; Mele, G.; Salvatore, M.; Aiello, M.; Cavaliere, C. Self-Reported Sleep Quality Across Age Modulates Resting-State Functional Connectivity in Limbic and Fronto-Temporo-Parietal Networks: An Exploratory Cross-Sectional fMRI Study. Front. Aging. Neurosci. 2022, 14, 806374. [Google Scholar] [CrossRef] [PubMed]
- Dinges, D.F.; Pack, F.; Williams, K.; Gillen, K.A.; Powell, J.W.; Ott, G.E.; Aptowicz, C.; Pack, A.I. Cumulative sleepiness, mood disturbance, and psychomotor vigilance performance decrements during a week of sleep restricted to 4-5 hours per night. Sleep 1997, 20, 267–277. [Google Scholar] [PubMed]
- Bonnet, M.H.; Arand, D.L. Clinical effects of sleep fragmentation versus sleep deprivation. Sleep Med. Rev. 2003, 7, 297–310. [Google Scholar] [CrossRef]
- Ben Simon, E.; Maron-Katz, A.; Lahav, N.; Shamir, R.; Hendler, T. Tired and misconnected: A breakdown of brain modularity following sleep deprivation. Hum. Brain Mapp. 2017, 38, 3300–3314. [Google Scholar] [CrossRef]
- Kaufmann, T.; Elvsåshagen, T.; Alnæs, D.; Zak, N.; Pedersen, P.Ø.; Norbom, L.B.; Quraishi, S.H.; Tagliazucchi, E.; Laufs, H.; Bjørnerud, A.; et al. The brain functional connectome is robustly altered by lack of sleep. NeuroImage 2016, 127, 324–332. [Google Scholar] [CrossRef]
- Drummond, S.P.; Paulus, M.P.; Tapert, S.F. Effects of two nights sleep deprivation and two nights recovery sleep on response inhibition. J. Sleep Res. 2006, 15, 261–265. [Google Scholar] [CrossRef]
- Li, C.; Fronczek-Poncelet, J.; Lange, D.; Hennecke, E.; Kroll, T.; Matusch, A.; Aeschbach, D.; Bauer, A.; Elmenhorst, E.M.; Elmenhorst, D. Impact of acute sleep deprivation on dynamic functional connectivity states. Hum. Brain Mapp. 2020, 41, 994–1005. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J.F.; Romero, S.; Mañanas, M.A.; Alcalá, M.; Antonijoan, R.M.; Giménez, S. Acute Sleep Deprivation Induces a Local Brain Transfer Information Increase in the Frontal Cortex in a Widespread Decrease Context. Sensors 2016, 16, 540. [Google Scholar] [CrossRef] [PubMed]
- Mander, B.A.; Santhanam, S.; Saletin, J.M.; Walker, M.P. Wake deterioration and sleep restoration of human learning. Curr. Biol. 2011, 21, R183–R184. [Google Scholar] [CrossRef] [PubMed]
- Mander, B.A.; Rao, V.; Lu, B.; Saletin, J.M.; Ancoli-Israel, S.; Jagust, W.J.; Walker, M.P. Impaired prefrontal sleep spindle regulation of hippocampal-dependent learning in older adults. Cereb. Cortex 2014, 24, 3301–3309. [Google Scholar] [CrossRef]
- Miller, S.L.; Celone, K.; DePeau, K.; Diamond, E.; Dickerson, B.C.; Rentz, D.; Pihlajamäki, M.; Sperling, R.A. Age-related memory impairment associated with loss of parietal deactivation but preserved hippocampal activation. Proc. Natl. Acad. Sci. USA 2008, 105, 2181–2186. [Google Scholar] [CrossRef]
- Sperling, R.A.; Bates, J.F.; Chua, E.F.; Cocchiarella, A.J.; Rentz, D.M.; Rosen, B.R.; Schacter, D.L.; Albert, M.S. fMRI studies of associative encoding in young and elderly controls and mild Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 2003, 74, 44–50. [Google Scholar] [CrossRef]
- Lim, A.S.; Fleischman, D.A.; Dawe, R.J.; Yu, L.; Arfanakis, K.; Buchman, A.S.; Bennett, D.A. Regional Neocortical Gray Matter Structure and Sleep Fragmentation in Older Adults. Sleep 2016, 39, 227–235. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, J.; Xu, L.; Song, T.; Peng, Z.; Zhang, Z.; An, X.; Chen, S.; Zhong, X.; Shao, Y. Reduced Resting-State EEG Power Spectra and Functional Connectivity after 24 and 36 Hours of Sleep Deprivation. Brain Sci. 2023, 13, 949. https://doi.org/10.3390/brainsci13060949
Lian J, Xu L, Song T, Peng Z, Zhang Z, An X, Chen S, Zhong X, Shao Y. Reduced Resting-State EEG Power Spectra and Functional Connectivity after 24 and 36 Hours of Sleep Deprivation. Brain Sciences. 2023; 13(6):949. https://doi.org/10.3390/brainsci13060949
Chicago/Turabian StyleLian, Jie, Lin Xu, Tao Song, Ziyi Peng, Zheyuan Zhang, Xin An, Shufang Chen, Xiao Zhong, and Yongcong Shao. 2023. "Reduced Resting-State EEG Power Spectra and Functional Connectivity after 24 and 36 Hours of Sleep Deprivation" Brain Sciences 13, no. 6: 949. https://doi.org/10.3390/brainsci13060949
APA StyleLian, J., Xu, L., Song, T., Peng, Z., Zhang, Z., An, X., Chen, S., Zhong, X., & Shao, Y. (2023). Reduced Resting-State EEG Power Spectra and Functional Connectivity after 24 and 36 Hours of Sleep Deprivation. Brain Sciences, 13(6), 949. https://doi.org/10.3390/brainsci13060949