
Motor Cortical Correlates of Paired Associative Stimulation Induced Plasticity: A TMS-EEG Study

 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Methods and Materials
2.1. Participants
2.2. Electromyography (EMG)
2.3. Electroencephalography (EEG)
2.4. Electrical Nerve Stimulation
2.5. TMS
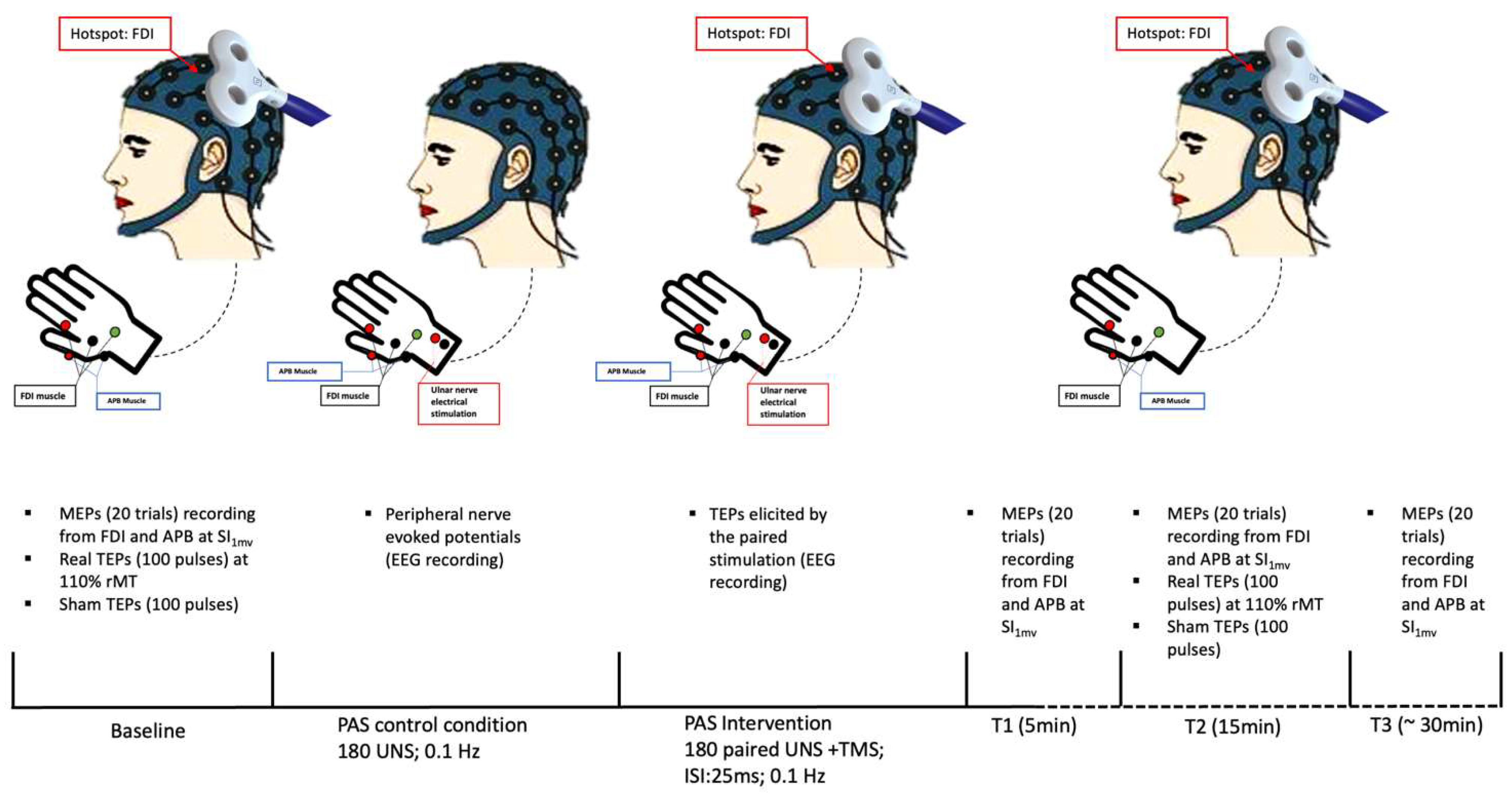
2.6. Experimental Paradigm
2.7. Data Analysis
2.8. Statistical Analysis
3. Results
3.1. Effect of PAS on MEPs
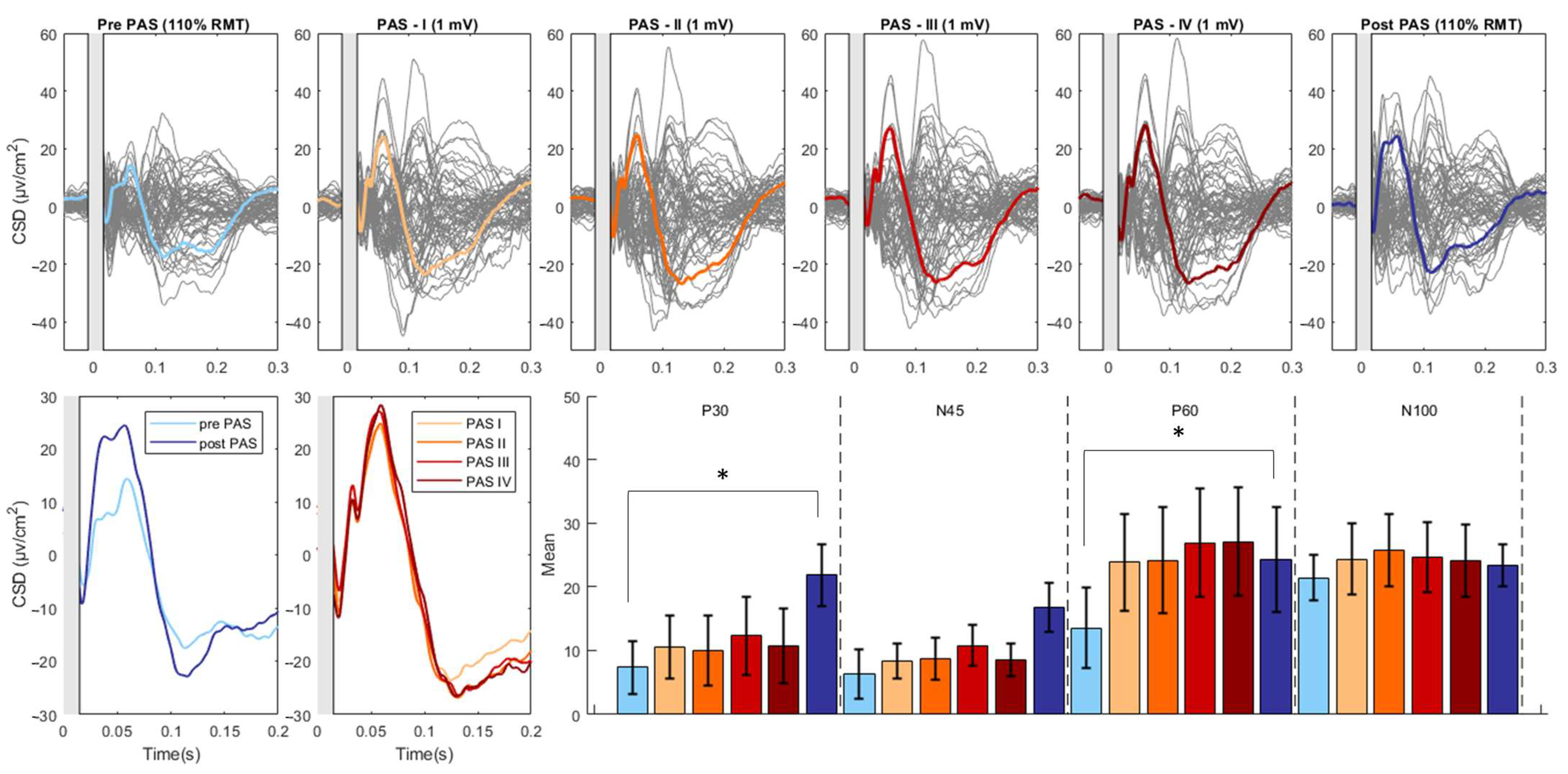
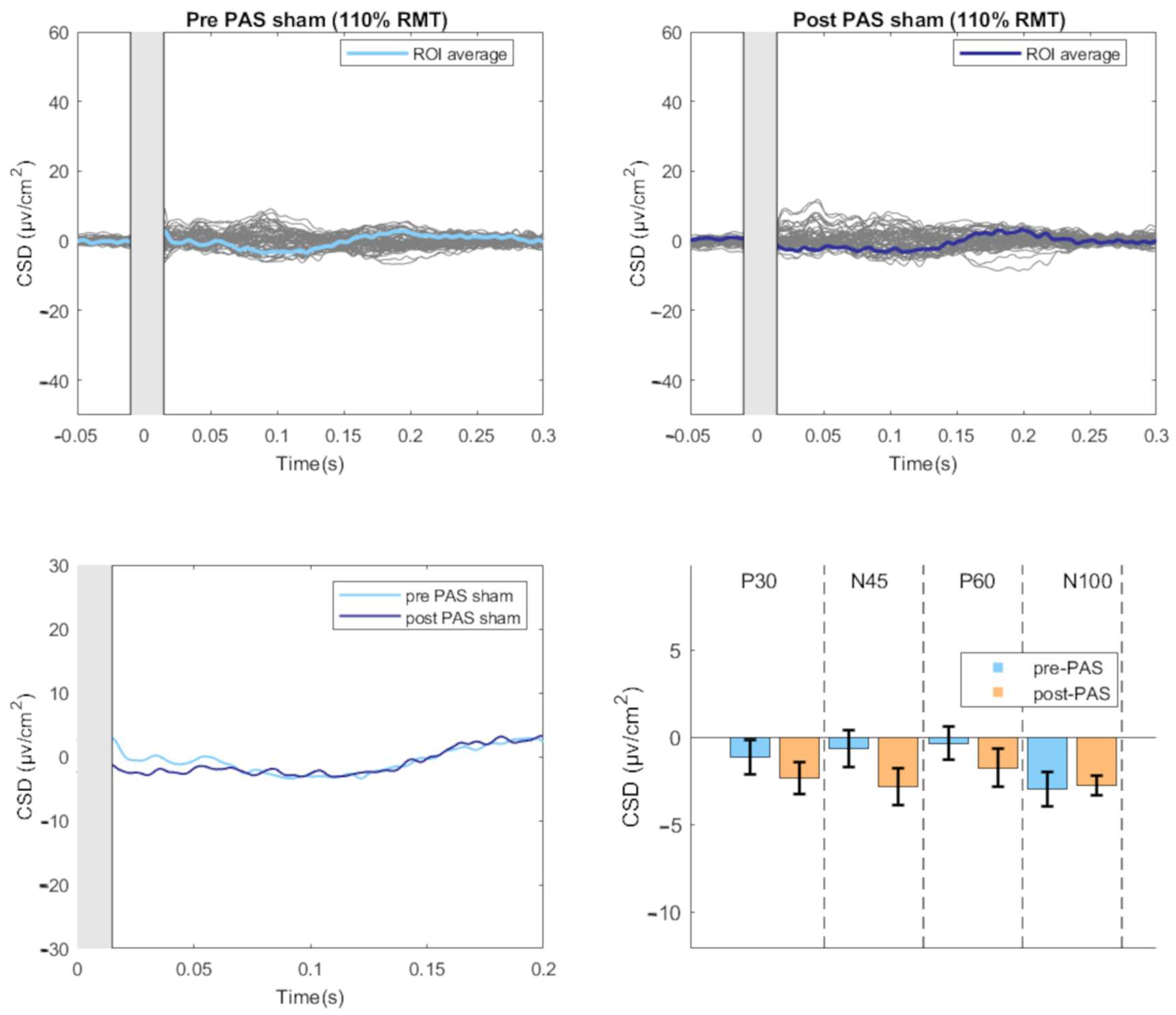
3.2. Effect of PAS on TEPs
3.3. Modulation of TEPs during the PAS Paradigm
3.4. Relationship between PAS-Induced Effects on MEPs and TEPs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stefan, K. Induction of Plasticity in the Human Motor Cortex by Paired Associative Stimulation. Brain 2000, 123, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Stefan, K.; Kunesch, E.; Benecke, R.; Cohen, L.G.; Classen, J. Mechanisms of Enhancement of Human Motor Cortex Excitability Induced by Interventional Paired Associative Stimulation. J. Physiol. 2002, 543, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Wolters, A.; Sandbrink, F.; Schlottmann, A.; Kunesch, E.; Stefan, K.; Cohen, L.G.; Benecke, R.; Classen, J. A Temporally Asymmetric Hebbian Rule Governing Plasticity in the Human Motor Cortex. J. Neurophysiol. 2003, 89, 2339–2345. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, U.; Iliać, T.V.; Pauli, C.; Meintzschel, F.; Ruge, D. Learning Modifies Subsequent Induction of Long-Term Potentiation-Like and Long-Term Depression-Like Plasticity in Human Motor Cortex. J. Neurosci. 2004, 24, 1666–1672. [Google Scholar] [CrossRef] [PubMed]
- Meunier, S.; Russmann, H.; Simonetta-Moreau, M.; Hallett, M. Changes in Spinal Excitability after PAS. J. Neurophysiol. 2007, 97, 3131–3135. [Google Scholar] [CrossRef]
- Suppa, A.; Quartarone, A.; Siebner, H.; Chen, R.; Di Lazzaro, V.; Del Giudice, P.; Paulus, W.; Rothwell, J.C.; Ziemann, U.; Classen, J. The Associative Brain at Work: Evidence from Paired Associative Stimulation Studies in Humans. Clin. Neurophysiol. 2017, 128, 2140–2164. [Google Scholar] [CrossRef]
- Carson, R.G.; Kennedy, N.C. Modulation of Human Corticospinal Excitability by Paired Associative Stimulation. Front. Hum. Neurosci. 2013, 7, 00823. [Google Scholar] [CrossRef]
- Paus, T.; Sipila, P.K.; Strafella, A.P. Synchronization of Neuronal Activity in the Human Primary Motor Cortex by Transcranial Magnetic Stimulation: An EEG Study. J. Neurophysiol. 2001, 86, 1983–1990. [Google Scholar] [CrossRef]
- Leodori, G.; Thirugnanasambandam, N.; Conn, H.; Popa, T.; Berardelli, A.; Hallett, M. Intracortical Inhibition and Surround Inhibition in the Motor Cortex: A TMS-EEG Study. Front. Neurosci. 2019, 13, 612. [Google Scholar] [CrossRef]
- Tremblay, S.; Rogasch, N.C.; Premoli, I.; Blumberger, D.M.; Casarotto, S.; Chen, R.; Di Lazzaro, V.; Farzan, F.; Ferrarelli, F.; Fitzgerald, P.B.; et al. Clinical Utility and Prospective of TMS–EEG. Clin. Neurophysiol. 2019, 130, 802–844. [Google Scholar] [CrossRef]
- Julkunen, P.; Kimiskidis, V.K.; Belardinelli, P. Bridging the Gap: TMS-EEG from Lab to Clinic. J. Neurosci. Methods 2022, 369, 109482. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Pavon, J.C.; Veniero, D.; Bergmann, T.O.; Belardinelli, P.; Bortoletto, M.; Casarotto, S.; Casula, E.P.; Farzan, F.; Fecchio, M.; Julkunen, P.; et al. TMS Combined with EEG: Recommendations and Open Issues for Data Collection and Analysis. Brain Stimul. 2023, 16, 567–593. [Google Scholar] [CrossRef]
- Leodori, G.; Fabbrini, A.; De Bartolo, M.I.; Costanzo, M.; Asci, F.; Palma, V.; Belvisi, D.; Conte, A.; Berardelli, A. Cortical Mechanisms Underlying Variability in Intermittent Theta-Burst Stimulation-Induced Plasticity: A TMS-EEG Study. Clin. Neurophysiol. 2021, 132, 2519–2531. [Google Scholar] [CrossRef] [PubMed]
- Farzan, F.; Barr, M.S.; Hoppenbrouwers, S.S.; Fitzgerald, P.B.; Chen, R.; Pascual-Leone, A.; Daskalakis, Z.J. The EEG Correlates of the TMS-Induced EMG Silent Period in Humans. NeuroImage 2013, 83, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Premoli, I.; Király, J.; Müller-Dahlhaus, F.; Zipser, C.M.; Rossini, P.; Zrenner, C.; Ziemann, U.; Belardinelli, P. Short-Interval and Long-Interval Intracortical Inhibition of TMS-Evoked EEG Potentials. Brain Stimul. 2018, 11, 818–827. [Google Scholar] [CrossRef]
- Rogasch, N.C.; Daskalakis, Z.J.; Fitzgerald, P.B. Mechanisms Underlying Long-Interval Cortical Inhibition in the Human Motor Cortex: A TMS-EEG Study. J. Neurophysiol. 2013, 109, 89–98. [Google Scholar] [CrossRef]
- Fernandez, L.; Biabani, M.; Do, M.; Opie, G.M.; Hill, A.T.; Barham, M.P.; Teo, W.-P.; Byrne, L.K.; Rogasch, N.C.; Enticott, P.G. Assessing Cerebellar-Cortical Connectivity Using Concurrent TMS-EEG: A Feasibility Study. J. Neurophysiol. 2021, 125, 1768–1787. [Google Scholar] [CrossRef]
- Ferreri, F.; Pasqualetti, P.; Määttä, S.; Ponzo, D.; Ferrarelli, F.; Tononi, G.; Mervaala, E.; Miniussi, C.; Rossini, P.M. Human Brain Connectivity during Single and Paired Pulse Transcranial Magnetic Stimulation. NeuroImage 2011, 54, 90–102. [Google Scholar] [CrossRef]
- Hallett, M.; Di Iorio, R.; Rossini, P.M.; Park, J.E.; Chen, R.; Celnik, P.; Strafella, A.P.; Matsumoto, H.; Ugawa, Y. Contribution of Transcranial Magnetic Stimulation to Assessment of Brain Connectivity and Networks. Clin. Neurophysiol. 2017, 128, 2125–2139. [Google Scholar] [CrossRef]
- Kimiskidis, V.K. Transcranial Magnetic Stimulation (TMS) Coupled with Electroencephalography (EEG): Biomarker of the Future. Rev. Neurol. 2016, 172, 123–126. [Google Scholar] [CrossRef]
- Kallioniemi, E.; Daskalakis, Z.J. Identifying Novel Biomarkers with TMS-EEG–Methodological Possibilities and Challenges. J. Neurosci. Methods 2022, 377, 109631. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Pajevic, S.; Chen, L.; Leodori, G.; Vial, F.; Avram, A.V.; Zhang, Y.; McGurrin, P.; Cohen, L.G.; Basser, P.J.; et al. Identifying Transcranial Magnetic Stimulation Induced EEG Signatures of Different Neuronal Elements in Primary Motor Cortex. Clin. Neurophysiol. 2022, 141, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Belardinelli, P.; Ziemann, U. Bihemispheric Sensorimotor Oscillatory Network States Determine Cortical Responses to Transcranial Magnetic Stimulation. Brain Stimul. 2022, 15, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Bikmullina, R.; Kičić, D.; Carlson, S.; Nikulin, V.V. Electrophysiological Correlates of Short-Latency Afferent Inhibition: A Combined EEG and TMS Study. Exp. Brain Res. 2009, 194, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Ponzo, D.; Hukkanen, T.; Mervaala, E.; Könönen, M.; Pasqualetti, P.; Vecchio, F.; Rossini, P.M.; Määttä, S. Human Brain Cortical Correlates of Short-Latency Afferent Inhibition: A Combined EEG–TMS Study. J. Neurophysiol. 2012, 108, 314–323. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Profice, P.; Ranieri, F.; Capone, F.; Dileone, M.; Oliviero, A.; Pilato, F. I-Wave Origin and Modulation. Brain Stimul. 2012, 5, 512–525. [Google Scholar] [CrossRef]
- Rossi, S.; Antal, A.; Bestmann, S.; Bikson, M.; Brewer, C.; Brockmöller, J.; Carpenter, L.L.; Cincotta, M.; Chen, R.; Daskalakis, J.D.; et al. Safety and Recommendations for TMS Use in Healthy Subjects and Patient Populations, with Updates on Training, Ethical and Regulatory Issues: Expert Guidelines. Clin. Neurophysiol. 2021, 132, 269–306. [Google Scholar] [CrossRef]
- Russo, S.; Sarasso, S.; Puglisi, G.E.; Dal Palù, D.; Pigorini, A.; Casarotto, S.; D’Ambrosio, S.; Astolfi, A.; Massimini, M.; Rosanova, M.; et al. TAAC-TMS Adaptable Auditory Control: A Universal Tool to Mask TMS Clicks. J. Neurosci. Methods 2022, 370, 109491. [Google Scholar] [CrossRef]
- Leodori, G.; Rocchi, L.; Mancuso, M.; De Bartolo, M.I.; Baione, V.; Costanzo, M.; Belvisi, D.; Conte, A.; Defazio, G.; Berardelli, A. The Effect of Stimulation Frequency on Transcranial Evoked Potentials. Transl. Neurosci. 2022, 13, 211–217. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An Open Source Toolbox for Analysis of Single-Trial EEG Dynamics Including Independent Component Analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Rogasch, N.C.; Sullivan, C.; Thomson, R.H.; Rose, N.S.; Bailey, N.W.; Fitzgerald, P.B.; Farzan, F.; Hernandez-Pavon, J.C. Analysing Concurrent Transcranial Magnetic Stimulation and Electroencephalographic Data: A Review and Introduction to the Open-Source TESA Software. NeuroImage 2017, 147, 934–951. [Google Scholar] [CrossRef] [PubMed]
- Leodori, G.; Mancuso, M.; Maccarrone, D.; Tartaglia, M.; Ianniello, A.; Certo, F.; Baione, V.; Ferrazzano, G.; Malimpensa, L.; Belvisi, D.; et al. Neural Bases of Motor Fatigue in Multiple Sclerosis: A Multimodal Approach Using Neuromuscular Assessment and TMS-EEG. Neurobiol. Dis. 2023, 180, 106073. [Google Scholar] [CrossRef] [PubMed]
- Leodori, G.; De Bartolo, M.I.; Guerra, A.; Fabbrini, A.; Rocchi, L.; Latorre, A.; Paparella, G.; Belvisi, D.; Conte, A.; Bhatia, K.P.; et al. Motor Cortical Network Excitability in Parkinson’s Disease. Mov. Disord. 2022, 37, 734–744. [Google Scholar] [CrossRef]
- Mazzi, C.; Mazzeo, G.; Savazzi, S. Markers of TMS-Evoked Visual Conscious Experience in a Patient with Altitudinal Hemianopia. Conscious. Cogn. 2017, 54, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Bertazzoli, G.; Esposito, R.; Mutanen, T.P.; Ferrari, C.; Ilmoniemi, R.J.; Miniussi, C.; Bortoletto, M. The Impact of Artifact Removal Approaches on TMS–EEG Signal. NeuroImage 2021, 239, 118272. [Google Scholar] [CrossRef]
- Mutanen, T.P.; Kukkonen, M.; Nieminen, J.O.; Stenroos, M.; Sarvas, J.; Ilmoniemi, R.J. Recovering TMS-Evoked EEG Responses Masked by Muscle Artifacts. NeuroImage 2016, 139, 157–166. [Google Scholar] [CrossRef]
- Mutanen, T.P.; Metsomaa, J.; Makkonen, M.; Varone, G.; Marzetti, L.; Ilmoniemi, R.J. Source-Based Artifact-Rejection Techniques for TMS–EEG. J. Neurosci. Methods 2022, 382, 109693. [Google Scholar] [CrossRef]
- Kayser, J.; Tenke, C.E. Issues and Considerations for Using the Scalp Surface Laplacian in EEG/ERP Research: A Tutorial Review. Int. J. Psychophysiol. 2015, 97, 189–209. [Google Scholar] [CrossRef]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open Source Software for Advanced Analysis of MEG, EEG, and Invasive Electrophysiological Data. Comput. Intell. Neurosci. 2011, 2011, 1–9. [Google Scholar] [CrossRef]
- Wolters, A.; Schmidt, A.; Schramm, A.; Zeller, D.; Naumann, M.; Kunesch, E.; Benecke, R.; Reiners, K.; Classen, J. Timing-Dependent Plasticity in Human Primary Somatosensory Cortex: Timing-Dependent Plasticity in Human Somatosensory Cortex. J. Physiol. 2005, 565, 1039–1052. [Google Scholar] [CrossRef]
- Yamashita, A.; Murakami, T.; Hattori, N.; Miyai, I.; Ugawa, Y. Intensity Dependency of Peripheral Nerve Stimulation in Spinal LTP Induced by Paired Associative Corticospinal-Motoneuronal Stimulation (PCMS). PLoS ONE 2021, 16, e0259931. [Google Scholar] [CrossRef] [PubMed]
- Bonato, C.; Miniussi, C.; Rossini, P.M. Transcranial Magnetic Stimulation and Cortical Evoked Potentials: A TMS/EEG Co-Registration Study. Clin. Neurophysiol. 2006, 117, 1699–1707. [Google Scholar] [CrossRef] [PubMed]
- Mäki, H.; Ilmoniemi, R.J. The Relationship between Peripheral and Early Cortical Activation Induced by Transcranial Magnetic Stimulation. Neurosci. Lett. 2010, 478, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Cash, R.F.H.; Noda, Y.; Zomorrodi, R.; Radhu, N.; Farzan, F.; Rajji, T.K.; Fitzgerald, P.B.; Chen, R.; Daskalakis, Z.J.; Blumberger, D.M. Characterization of Glutamatergic and GABAA-Mediated Neurotransmission in Motor and Dorsolateral Prefrontal Cortex Using Paired-Pulse TMS–EEG. Neuropsychopharmacology 2017, 42, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Fröhlich, F. Pinging the Brain with Transcranial Magnetic Stimulation Reveals Cortical Reactivity in Time and Space. Brain Stimul. 2021, 14, 304–315. [Google Scholar] [CrossRef]
- Komssi, S.; Kähkönen, S.; Ilmoniemi, R.J. The Effect of Stimulus Intensity on Brain Responses Evoked by Transcranial Magnetic Stimulation: TMS-Evoked Brain Responses. Hum. Brain Mapp. 2004, 21, 154–164. [Google Scholar] [CrossRef]
- Fecchio, M.; Pigorini, A.; Comanducci, A.; Sarasso, S.; Casarotto, S.; Premoli, I.; Derchi, C.-C.; Mazza, A.; Russo, S.; Resta, F.; et al. The Spectral Features of EEG Responses to Transcranial Magnetic Stimulation of the Primary Motor Cortex Depend on the Amplitude of the Motor Evoked Potentials. PLoS ONE 2017, 12, e0184910. [Google Scholar] [CrossRef]
- Leodori, G.; Belvisi, D.; De Bartolo, M.I.; Fabbrini, A.; Costanzo, M.; Vial, F.; Conte, A.; Hallett, M.; Berardelli, A. Re-emergent Tremor in Parkinson’s Disease: The Role of the Motor Cortex. Mov. Disord. 2020, 35, 1002–1011. [Google Scholar] [CrossRef]
- Biabani, M.; Fornito, A.; Coxon, J.P.; Fulcher, B.D.; Rogasch, N.C. The Correspondence between EMG and EEG Measures of Changes in Cortical Excitability Following Transcranial Magnetic Stimulation. J. Physiol. 2021, 599, 2907–2932. [Google Scholar] [CrossRef]
- Kallioniemi, E. Cortical Excitability Measures from TMS-EEG and TMS-EMG–Two Sides of the Same Story? J. Physiol. 2021, 599, 2779–2780. [Google Scholar] [CrossRef]
- Petrichella, S.; Johnson, N.; He, B. The Influence of Corticospinal Activity on TMS-Evoked Activity and Connectivity in Healthy Subjects: A TMS-EEG Study. PLoS ONE 2017, 12, e0174879. [Google Scholar] [CrossRef] [PubMed]
- Siebner, H.R.; Funke, K.; Aberra, A.S.; Antal, A.; Bestmann, S.; Chen, R.; Classen, J.; Davare, M.; Di Lazzaro, V.; Fox, P.T.; et al. Transcranial Magnetic Stimulation of the Brain: What Is Stimulated?—A Consensus and Critical Position Paper. Clin. Neurophysiol. 2022, 140, 59–97. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Time Course | FDI Muscle | APB Muscle | ||
|---|---|---|---|---|
| Mean Amplitude (mV) | Std. Deviation | Mean Amplitude (mV) | Std. Deviation | |
| Baseline | 1.229 | 0.393 | 0.881 | 0.432 |
| T1 Post-Pas | 1.844 | 0.862 | 0.917 | 0.532 |
| T2 Post-PAS | 2.094 | 0.724 | 0.909 | 0.476 |
| T3 Post-PAS | 1.885 | 0.707 | 0.882 | 0.450 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costanzo, M.; Leodori, G.; Cutrona, C.; Marchet, F.; De Bartolo, M.I.; Mancuso, M.; Belvisi, D.; Conte, A.; Berardelli, A.; Fabbrini, G. Motor Cortical Correlates of Paired Associative Stimulation Induced Plasticity: A TMS-EEG Study. Brain Sci. 2023, 13, 921. https://doi.org/10.3390/brainsci13060921
Costanzo M, Leodori G, Cutrona C, Marchet F, De Bartolo MI, Mancuso M, Belvisi D, Conte A, Berardelli A, Fabbrini G. Motor Cortical Correlates of Paired Associative Stimulation Induced Plasticity: A TMS-EEG Study. Brain Sciences. 2023; 13(6):921. https://doi.org/10.3390/brainsci13060921
Chicago/Turabian StyleCostanzo, Matteo, Giorgio Leodori, Carolina Cutrona, Francesco Marchet, Maria Ilenia De Bartolo, Marco Mancuso, Daniele Belvisi, Antonella Conte, Alfredo Berardelli, and Giovanni Fabbrini. 2023. "Motor Cortical Correlates of Paired Associative Stimulation Induced Plasticity: A TMS-EEG Study" Brain Sciences 13, no. 6: 921. https://doi.org/10.3390/brainsci13060921
APA StyleCostanzo, M., Leodori, G., Cutrona, C., Marchet, F., De Bartolo, M. I., Mancuso, M., Belvisi, D., Conte, A., Berardelli, A., & Fabbrini, G. (2023). Motor Cortical Correlates of Paired Associative Stimulation Induced Plasticity: A TMS-EEG Study. Brain Sciences, 13(6), 921. https://doi.org/10.3390/brainsci13060921