
Shared Mechanisms of GABAergic and Opioidergic Transmission Regulate Corticolimbic Reward Systems and Cognitive Aspects of Motivational Behaviors



Abstract
1. Introduction
2. Neurobiology of Corticolimbic Endogenous Opioidergic Neurons and GABAergic Interneurons
3. Neuroanatomical Studies of Corticolimbic GABAergic Interneurons and Opioidergic System in Reward-Related Mechanisms
3.1. Prefrontal Cortex
3.2. Hippocampus
3.3. The Amygdala

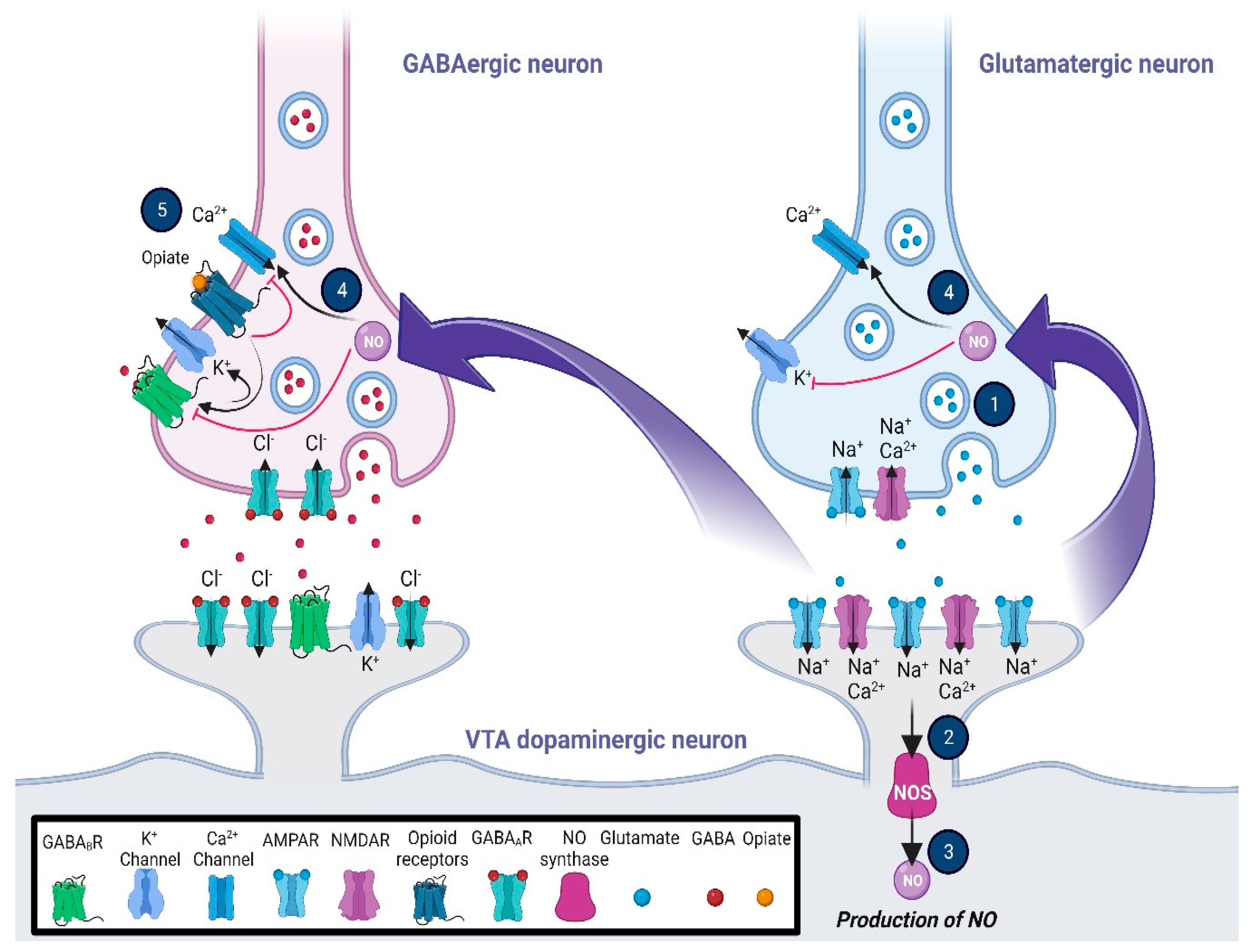{kind=link}
{kind=link}
| Opioid Receptor | GABAergic Marker | Brain Region |
|---|---|---|
| κ-opioid receptors | Glutamate decarboxylase [91] | Hippocampus [92] |
| κ-opioid receptors | Calretinin [93] | Dorsal striatum [93] |
| κ-opioid receptors | Parvalbumin [94] | Basolateral amygdala [94] |
| δ-opioid receptors | Somatostatin [95] | Dentate gyrus [95] |
| δ-opioid receptors | Parvalbumin [96] | Hippocampal CA1 [96] |
| δ-opioid receptors | Parvalbumin [97] | Hippocampal CA2 [97] |
| δ-opioid receptors | Somatostatin [98] | Prelimbic cortex [98] |
| δ-opioid receptors | Parvalbumin [98] | Prelimbic cortex [98] |
| δ-opioid receptors | Calretinin [93] | Nucleus accumbens core [93] |
| μ-opioid receptors | Glutamate decarboxylase [23] | Ventral tegmental area [23] |
| μ-opioid receptors | Glutamate decarboxylase [73] | Dentate gyrus [73] |
| μ-opioid receptors | Parvalbumin [73] | Dentate gyrus [73] |
| μ-opioid receptors | Somatostatin [73] | Hippocampus (OLM) [23] |
| μ-opioid receptors | Somatostatin [99] | Dentate gyrus [99] |
| μ-opioid receptors | Parvalbumin [100] | Hippocampal CA1 [100] |
| μ-opioid receptors | Parvalbumin [101] | Orbitofrontal cortex [101] |
| μ-opioid receptors | Calbindin-D28K [102] | Dorsal Striatum (DA neurons) [102] |
| μ-opioid receptors | Calbindin-D28K [96] | Nucleus accumbens core (MSNs) [96] |
| μ-opioid receptors | Vasoactive intestinal peptide [103] | Neocortex [103] |
| μ-opioid receptors | Vasoactive intestinal peptide [104] | Anterior cingulate cortex (L1) [104] |
| μ-opioid receptors | Somatostatin [104] | Anterior cingulate cortex (L5 and 6b) [104] |
| μ-opioid receptors | Calretinin [105] | Central amygdala [105] |
| μ-opioid receptors | Glutamate decarboxylase [56] | Nucleus accumbens [56] |
| μ-opioid receptors | Parvalbumin [106] | Hippocampal CA3 [106] |
4. Opioid and GABA Receptor Dynamics in Motivational Behaviors
5. Neurochemical Markers for the Colocalization of Opioidergic Neurons and GABAergic Interneurons
5.1. Opioids and Glutamate Decarboxylase 65/67
5.2. Opioids and Somatostatin
5.3. Opioids and Parvalbumin
5.4. Opioids and Calbindin-D28K
5.5. Opioids and Vasoactive Intestinal Peptide
5.6. Opioids and Calretinin
5.7. Opioids and Neuropeptide Y
6. Opioids Regulate GABAergic Transmission-Induced Changes in Plasticity
7. Intracellular Mechanism Underlying Opioid–GABA Crosstalk
7.1. Intracellular G Protein Involvement in Opioid/GABA Reinforcement
7.2. Opioidergic/GABAergic Intracellular Signaling Pathways
8. Inhibitory Mechanisms of Reward–Aversion in Addiction
9. GABAergic/Opioidergic Systems Modulating Feedback and Feedforward Activity
10. Opioids and GABA in Network Oscillations
11. Modulation of GABAergic/Opioidergic Transmission in Pathology and Therapeutics
12. Discussion
13. Limitations
14. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Q.; Ding, S.-L.; Li, Y.; Royall, J.; Feng, D.; Lesnar, P.; Graddis, N.; Naeemi, M.; Facer, B.; Ho, A. The Allen mouse brain common coordinate framework: A 3D reference atlas. Cell 2020, 181, 936–953.e920. [Google Scholar] [CrossRef] [PubMed]
- Joffe, M.E.; Grueter, C.A.; Grueter, B.A. Biological substrates of addiction. Wiley Interdiscip. Rev. Cogn. Sci. 2014, 5, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Creed, M.C.; Ntamati, N.R.; Tan, K.R. VTA GABA neurons modulate specific learning behaviors through the control of dopamine and cholinergic systems. Front. Behav. Neurosci. 2014, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.-H. Stress and the dopaminergic reward system. Exp. Mol. Med. 2020, 52, 1879–1890. [Google Scholar] [CrossRef]
- Bariselli, S.; Glangetas, C.; Tzanoulinou, S.; Bellone, C. Ventral tegmental area subcircuits process rewarding and aversive experiences. J. Neurochem. 2016, 139, 1071–1080. [Google Scholar] [CrossRef]
- Reeves, K.C.; Shah, N.; Muñoz, B.; Atwood, B.K. Opioid receptor-mediated regulation of neurotransmission in the brain. Front. Mol. Neurosci. 2022, 15, 919773. [Google Scholar] [CrossRef]
- Qu, C.-L.; Huo, F.-Q.; Huang, F.-S.; Tang, J.-S. Activation of mu-opioid receptors in the ventrolateral orbital cortex inhibits the GABAergic miniature inhibitory postsynaptic currents in rats. Neurosci. Lett. 2015, 592, 64–69. [Google Scholar] [CrossRef]
- Bull, F.A.; Baptista-Hon, D.T.; Lambert, J.J.; Walwyn, W.; Hales, T.G. Morphine activation of mu opioid receptors causes disinhibition of neurons in the ventral tegmental area mediated by β-arrestin2 and c-Src. Sci. Rep. 2017, 7, 9969. [Google Scholar] [CrossRef]
- Svoboda, K.R.; Adams, C.E.; Lupica, C.R. Opioid receptor subtype expression defines morphologically distinct classes of hippocampal interneurons. J. Neurosci. 1999, 19, 85–95. [Google Scholar] [CrossRef]
- Johnson, S.; North, R. Opioids excite dopamine neurons by hyperpolarization of local interneurons. J. Neurosci. 1992, 12, 483–488. [Google Scholar] [CrossRef]
- Asgharpour-Masouleh, N.; Rezayof, A.; Alijanpour, S.; Delphi, L. Pharmacological activation of mediodorsal thalamic GABA-A receptors modulates morphine/cetirizine-induced changes in the prefrontal cortical GFAP expression in a rat model of neuropathic pain. Behav. Brain Res. 2023, 438, 114213. [Google Scholar] [CrossRef] [PubMed]
- Zarrindast, M.-R.; Ahmadi, S.; Haeri-Rohani, A.; Rezayof, A.; Jafari, M.-R.; Jafari-Sabet, M. GABAA receptors in the basolateral amygdala are involved in mediating morphine reward. Brain Res. 2004, 1006, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Pathan, H.; Williams, J. Basic opioid pharmacology: An update. Br. J. Pain 2012, 6, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Darcq, E.; Kieffer, B.L. Opioid receptors: Drivers to addiction? Nat. Rev. Neurosci. 2018, 19, 499–514. [Google Scholar] [CrossRef]
- Nummenmaa, L.; Saanijoki, T.; Tuominen, L.; Hirvonen, J.; Tuulari, J.J.; Nuutila, P.; Kalliokoski, K. μ-opioid receptor system mediates reward processing in humans. Nat. Commun. 2018, 9, 1500. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.A.; Basbaum, A.I.; Naidu, R.K. Opioid agonists & antagonists. In Basic & Clinical Pharmacology & Toxicology, 13th ed.; Katzung, B., Trevor, A., Eds.; Wiley: Hoboken, NJ, USA, 2015; p. 531. [Google Scholar]
- Ádám, D.; Arany, J.; Tóth, K.F.; Tóth, B.I.; Szöllősi, A.G.; Oláh, A. Opioidergic signaling—A neglected, yet potentially important player in atopic dermatitis. Int. J. Mol. Sci. 2022, 23, 4140. [Google Scholar] [CrossRef]
- Lam, M.P.; Gianoulakis, C. Effects of acute ethanol on corticotropin-releasing hormone and β-endorphin systems at the level of the rat central amygdala. Psychopharmacology 2011, 218, 229–239. [Google Scholar] [CrossRef]
- Netto, C.A. Role of brain Β-endorphin in memory modulation revisited. Neuroscience 2022, 497, 30–38. [Google Scholar] [CrossRef]
- Pilozzi, A.; Carro, C.; Huang, X. Roles of β-endorphin in stress, behavior, neuroinflammation, and brain energy metabolism. Int. J. Mol. Sci. 2020, 22, 338. [Google Scholar] [CrossRef]
- Schoenfeld, T.J.; Swanson, C. A Runner’s High for New Neurons? Potential Role for Endorphins in Exercise Effects on Adult Neurogenesis. Biomolecules 2021, 11, 1077. [Google Scholar] [CrossRef]
- Rysztak, L.G.; Jutkiewicz, E.M. The role of enkephalinergic systems in substance use disorders. Front. Syst. Neurosci. 2022, 16, 932546. [Google Scholar] [CrossRef] [PubMed]
- Kudo, T.; Konno, K.; Uchigashima, M.; Yanagawa, Y.; Sora, I.; Minami, M.; Watanabe, M. GABA ergic neurons in the ventral tegmental area receive dual GABA/enkephalin-mediated inhibitory inputs from the bed nucleus of the stria terminalis. Eur. J. Neurosci. 2014, 39, 1796–1809. [Google Scholar] [CrossRef]
- Hang, A.; Wang, Y.-J.; He, L.; Liu, J.-G. The role of the dynorphin/κ opioid receptor system in anxiety. Acta Pharmacol. Sin. 2015, 36, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Sardella, T.C.; Polgár, E.; Garzillo, F.; Furuta, T.; Kaneko, T.; Watanabe, M.; Todd, A.J. Dynorphin is expressed primarily by GABAergic neurons that contain galanin in the rat dorsal horn. Mol. Pain 2011, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Gilpin, N.W.; Roberto, M.; Koob, G.F.; Schweitzer, P. Kappa opioid receptor activation decreases inhibitory transmission and antagonizes alcohol effects in rat central amygdala. Neuropharmacology 2014, 77, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Bagosi, Z.; Jászberényi, M.; Bujdosó, E.; Szabó, G.; Telegdy, G. The effects of endomorphins and diprotin A on striatal dopamine release induced by electrical stimulation—An in vitro superfusion study in rats. Neurochem. Int. 2006, 49, 665–668. [Google Scholar] [CrossRef]
- Bagosi, Z.; Jászberényi, M.; Telegdy, G. The Effects of Endomorphins on Striatal [3 H] Gaba Release Induced by Electrical Stimulation: An In vitro Superfusion Study in Rats. Neurochem. Res. 2009, 34, 905–908. [Google Scholar] [CrossRef]
- Isaacson, J.S.; Scanziani, M. How inhibition shapes cortical activity. Neuron 2011, 72, 231–243. [Google Scholar] [CrossRef]
- Tremblay, R.; Lee, S.; Rudy, B. GABAergic interneurons in the neocortex: From cellular properties to circuits. Neuron 2016, 91, 260–292. [Google Scholar] [CrossRef]
- Lombardi, A.; Luhmann, H.J.; Kilb, W. Modelling the spatial and temporal constrains of the GABAergic influence on neuronal excitability. PLoS Comput. Biol. 2021, 17, e1009199. [Google Scholar] [CrossRef]
- Toniolo, S.; Sen, A.; Husain, M. Modulation of brain hyperexcitability: Potential new therapeutic approaches in Alzheimer’s disease. Int. J. Mol. Sci. 2020, 21, 9318. [Google Scholar] [CrossRef]
- Fogaça, M.V.; Duman, R.S. Cortical GABAergic dysfunction in stress and depression: New insights for therapeutic interventions. Front. Cell. Neurosci. 2019, 13, 87. [Google Scholar] [CrossRef]
- Lee, S.-E.; Lee, Y.; Lee, G.H. The regulation of glutamic acid decarboxylases in GABA neurotransmission in the brain. Arch. Pharmacal Res. 2019, 42, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Clausen, K.M.H.W.R.; Larsson, B.F.O.; Schousboe, P.K.L.A. 18 Functional and Pharmacological Aspects of GABA Transporters; Springer: New York, NY, USA, 2006. [Google Scholar]
- Ghit, A.; Assal, D.; Al-Shami, A.S.; Hussein, D.E.E. GABAA receptors: Structure, function, pharmacology, and related disorders. J. Genet. Eng. Biotechnol. 2021, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.W.; Liang, J. Role of GABAA receptors in alcohol use disorders suggested by chronic intermittent ethanol (CIE) rodent model. Mol. Brain 2017, 10, 1–20. [Google Scholar] [CrossRef]
- Agrawal, A.; Pergadia, M.L.; Saccone, S.F.; Hinrichs, A.L.; Lessov-Schlaggar, C.N.; Saccone, N.L.; Neuman, R.J.; Breslau, N.; Johnson, E.; Hatsukami, D. Gamma-aminobutyric acid receptor genes and nicotine dependence: Evidence for association from a case–control study. Addiction 2008, 103, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Morley, K.I.; Hansell, N.K.; Pergadia, M.L.; Montgomery, G.W.; Statham, D.J.; Todd, R.D.; Madden, P.A.; Heath, A.C.; Whitfield, J. Autosomal linkage analysis for cannabis use behaviors in Australian adults. Drug Alcohol Depend. 2008, 98, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Yang, M.; Mat, W.-K.; Tsang, S.-Y.; Su, Z.; Jiang, X.; Ng, S.-K.; Liu, S.; Hu, T.; Pun, F. GABRB2 haplotype association with heroin dependence in Chinese population. PLoS ONE 2015, 10, e0142049. [Google Scholar] [CrossRef] [PubMed]
- Cancedda, L.; Fiumelli, H.; Chen, K.; Poo, M.-M. Excitatory GABA action is essential for morphological maturation of cortical neurons in vivo. J. Neurosci. 2007, 27, 5224–5235. [Google Scholar] [CrossRef]
- Patrizi, A.; Scelfo, B.; Viltono, L.; Briatore, F.; Fukaya, M.; Watanabe, M.; Strata, P.; Varoqueaux, F.; Brose, N.; Fritschy, J.-M. Synapse formation and clustering of neuroligin-2 in the absence of GABAA receptors. Proc. Natl. Acad. Sci. USA 2008, 105, 13151–13156. [Google Scholar] [CrossRef]
- Hanics, J.; Barna, J.; Xiao, J.; Millán, J.L.; Fonta, C.; Négyessy, L. Ablation of TNAP function compromises myelination and synaptogenesis in the mouse brain. Cell Tissue Res. 2012, 349, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Palpagama, T.H.; Sagniez, M.; Kim, S.; Waldvogel, H.J.; Faull, R.L.; Kwakowsky, A. GABAA receptors are well preserved in the hippocampus of aged mice. eNeuro 2019, 6, ENEURO.0496-18.2019. [Google Scholar] [CrossRef] [PubMed]
- Sugita, S.; North, R. Opioid actions on neurons of rat lateral amygdala in vitro. Brain Res. 1993, 612, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Cohen, G.A.; Doze, V.A.; Madison, D.V. Opioid inhibition of GABA release from presynaptic terminals of rat hippocampal interneurons. Neuron 1992, 9, 325–335. [Google Scholar] [CrossRef]
- Li, C.; Sugam, J.A.; Lowery-Gionta, E.G.; McElligott, Z.A.; McCall, N.M.; Lopez, A.J.; McKlveen, J.M.; Pleil, K.E.; Kash, T.L. Mu opioid receptor modulation of dopamine neurons in the periaqueductal gray/dorsal raphe: A role in regulation of pain. Neuropsychopharmacology 2016, 41, 2122–2132. [Google Scholar] [CrossRef]
- Khastkhodaei, Z.; Muthuraman, M.; Yang, J.-W.; Groppa, S.; Luhmann, H.J. Functional and directed connectivity of the cortico-limbic network in mice in vivo. Brain Struct. Funct. 2021, 226, 685–700. [Google Scholar] [CrossRef]
- Preston, A.R.; Eichenbaum, H. Interplay of hippocampus and prefrontal cortex in memory. Curr. Biol. 2013, 23, R764–R773. [Google Scholar] [CrossRef]
- Meyer, H.S.; Schwarz, D.; Wimmer, V.C.; Schmitt, A.C.; Kerr, J.N.; Sakmann, B.; Helmstaedter, M. Inhibitory interneurons in a cortical column form hot zones of inhibition in layers 2 and 5A. Proc. Natl. Acad. Sci. USA 2011, 108, 16807–16812. [Google Scholar] [CrossRef]
- Goldberg, E.M.; Coulter, D.A. Mechanisms of epileptogenesis: A convergence on neural circuit dysfunction. Nat. Rev. Neurosci. 2013, 14, 337–349. [Google Scholar] [CrossRef]
- Marín, O. Interneuron dysfunction in psychiatric disorders. Nat. Rev. Neurosci. 2012, 13, 107–120. [Google Scholar] [CrossRef]
- Le Merrer, J.; Becker, J.A.; Befort, K.; Kieffer, B.L. Reward processing by the opioid system in the brain. Physiol. Rev. 2009, 89, 1379–1412. [Google Scholar] [CrossRef] [PubMed]
- Zieglgänsberger, W.; French, E.D.; Siggins, G.R.; Bloom, F.E. Opioid peptides may excite hippocampal pyramidal neurons by inhibiting adjacent inhibitory interneurons. Science 1979, 205, 415–417. [Google Scholar] [CrossRef]
- Listos, J.; Łupina, M.; Talarek, S.; Mazur, A.; Orzelska-Górka, J.; Kotlińska, J. The mechanisms involved in morphine addiction: An overview. Int. J. Mol. Sci. 2019, 20, 4302. [Google Scholar] [CrossRef] [PubMed]
- Hinkle, C.; Dedkov, E.; Buono, R.; Ferraro, T. Colocalization of MOR1 and GAD67 in mouse nucleus accumbens. bioRxiv 2019, 574434. [Google Scholar] [CrossRef]
- Carlén, M. What constitutes the prefrontal cortex? Science 2017, 358, 478–482. [Google Scholar] [CrossRef]
- Yizhar, O.; Fenno, L.E.; Prigge, M.; Schneider, F.; Davidson, T.J.; O’shea, D.J.; Sohal, V.S.; Goshen, I.; Finkelstein, J.; Paz, J.T. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature 2011, 477, 171–178. [Google Scholar] [CrossRef]
- Liu, L.; Xu, H.; Wang, J.; Li, J.; Tian, Y.; Zheng, J.; He, M.; Xu, T.-L.; Wu, Z.-Y.; Li, X.-M. Cell type–differential modulation of prefrontal cortical GABAergic interneurons on low gamma rhythm and social interaction. Sci. Adv. 2020, 6, eaay4073. [Google Scholar] [CrossRef]
- Royer, S.; Zemelman, B.V.; Losonczy, A.; Kim, J.; Chance, F.; Magee, J.C.; Buzsáki, G. Control of timing, rate and bursts of hippocampal place cells by dendritic and somatic inhibition. Nat. Neurosci. 2012, 15, 769–775. [Google Scholar] [CrossRef]
- Taki, K.; Kaneko, T.; Mizuno, N. A group of cortical interneurons expressing μ-opioid receptor-like immunoreactivity: A double immunofluorescence study in the rat cerebral cortex. Neuroscience 2000, 98, 221–231. [Google Scholar] [CrossRef]
- Witkowski, G.; Szulczyk, P. Opioid μ receptor activation inhibits sodium currents in prefrontal cortical neurons via a protein kinase A-and C-dependent mechanism. Brain Res. 2006, 1094, 92–106. [Google Scholar] [CrossRef]
- Colasanti, A.; Searle, G.E.; Long, C.J.; Hill, S.P.; Reiley, R.R.; Quelch, D.; Erritzoe, D.; Tziortzi, A.C.; Reed, L.J.; Lingford-Hughes, A.R.; et al. Endogenous Opioid Release in the Human Brain Reward System Induced by Acute Amphetamine Administration. Biol. Psychiatry 2012, 72, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.M.; O’Neil, J.P.; Janabi, M.; Marks, S.M.; Jagust, W.J.; Fields, H.L. Alcohol consumption induces endogenous opioid release in the human orbitofrontal cortex and nucleus accumbens. Sci. Transl. Med. 2012, 4, ra116. [Google Scholar] [CrossRef] [PubMed]
- Blasio, A.; Steardo, L.; Sabino, V.; Cottone, P. Opioid system in the medial prefrontal cortex mediates binge-like eating. Addict. Biol. 2014, 19, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Kong, E.; Lee, K.-H.; Do, J.; Kim, P.; Lee, D. Dynamic and stable hippocampal representations of social identity and reward expectation support associative social memory in male mice. Nat. Commun. 2023, 14, 2597. [Google Scholar] [CrossRef]
- Frank, L.E.; Preston, A.R.; Zeithamova, D. Functional connectivity between memory and reward centers across task and rest track memory sensitivity to reward. Cogn. Affect. Behav. Neurosci. 2019, 19, 503–522. [Google Scholar] [CrossRef]
- Ito, R.; Robbins, T.W.; Pennartz, C.M.; Everitt, B.J. Functional interaction between the hippocampus and nucleus accumbens shell is necessary for the acquisition of appetitive spatial context conditioning. J. Neurosci. 2008, 28, 6950–6959. [Google Scholar] [CrossRef]
- Guo, M.; Cao, D.; Zhu, S.; Fu, G.; Wu, Q.; Liang, J.; Cao, M. Chronic exposure to morphine decreases the expression of EAAT3 via opioid receptors in hippocampal neurons. Brain Res. 2015, 1628, 40–49. [Google Scholar] [CrossRef]
- Bezaire, M.J.; Soltesz, I. Quantitative assessment of CA1 local circuits: Knowledge base for interneuron-pyramidal cell connectivity. Hippocampus 2013, 23, 751–785. [Google Scholar] [CrossRef]
- McKenzie, S. Inhibition shapes the organization of hippocampal representations. Hippocampus 2018, 28, 659–671. [Google Scholar] [CrossRef]
- Drake, C.T.; Milner, T.A. Mu opioid receptors are in somatodendritic and axonal compartments of GABAergic neurons in rat hippocampal formation. Brain Res. 1999, 849, 203–215. [Google Scholar] [CrossRef]
- Drake, C.T.; Milner, T.A. Mu opioid receptors are in discrete hippocampal interneuron subpopulations. Hippocampus 2002, 12, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.H.; Han, K.S.; Lee, J.; Bae, J.Y.; An, H.; Park, S.; Oh, S.J.; Kim, E.; Hwang, E.; Bae, Y.C.; et al. Expression of µ-Opioid Receptor in CA1 Hippocampal Astrocytes. Exp. Neurobiol. 2018, 27, 120–128. [Google Scholar] [CrossRef]
- Matthies, H.; Schroeder, H.; Becker, A.; Loh, H.; Höllt, V.; Krug, M. Lack of expression of long-term potentiation in the dentate gyrus but not in the CA1 region of the hippocampus of mu-opioid receptor-deficient mice. Neuropharmacology 2000, 39, 952–960. [Google Scholar] [CrossRef]
- Sagratella, S.; Longo, R.; Domenici, M.R. Selective opposite modulation of dentate granule cells excitability by mu and kappa opioids in rat hippocampal slices. Neurosci. Lett. 1996, 205, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Andreu, V.; Langreck, C.; Pekarskaya, E.A.; Grinnell, S.G.; Allain, F.; Magalong, V.; Pintar, J.; Kieffer, B.L.; Harris, A.Z. Mu opioid receptors on hippocampal GABAergic interneurons are critical for the antidepressant effects of tianeptine. Neuropsychopharmacology 2022, 47, 1387–1397. [Google Scholar] [CrossRef]
- Zhang, T.A.; Tang, J.; Pidoplichko, V.I.; Dani, J.A. Addictive nicotine alters local circuit inhibition during the induction of in vivo hippocampal synaptic potentiation. J. Neurosci. 2010, 30, 6443–6453. [Google Scholar] [CrossRef] [PubMed]
- Aery Jones, E.A.; Rao, A.; Zilberter, M.; Djukic, B.; Bant, J.S.; Gillespie, A.K.; Koutsodendris, N.; Nelson, M.; Yoon, S.Y.; Huang, K.; et al. Dentate gyrus and CA3 GABAergic interneurons bidirectionally modulate signatures of internal and external drive to CA1. Cell Rep. 2021, 37, 110159. [Google Scholar] [CrossRef]
- Janak, P.H.; Tye, K.M. From circuits to behaviour in the amygdala. Nature 2015, 517, 284–292. [Google Scholar] [CrossRef]
- Sah, P.; Faber, E.L.; Lopez de Armentia, M.; Power, J. The amygdaloid complex: Anatomy and physiology. Physiol. Rev. 2003, 83, 803–834. [Google Scholar] [CrossRef]
- McDONALD, A.J. Is there an amygdala and how far does it extend? An anatomical perspective. Ann. N. Y. Acad. Sci. 2003, 985, 1–21. [Google Scholar] [CrossRef]
- Wilson, M.A.; Junor, L. The role of amygdalar mu-opioid receptors in anxiety-related responses in two rat models. Neuropsychopharmacology 2008, 33, 2957–2968. [Google Scholar] [CrossRef]
- Lebow, M.A.; Chen, A. Overshadowed by the amygdala: The bed nucleus of the stria terminalis emerges as key to psychiatric disorders. Mol. Psychiatry 2016, 21, 450–463. [Google Scholar] [CrossRef] [PubMed]
- Lichtenberg, N.T.; Wassum, K.M. Amygdala mu-opioid receptors mediate the motivating influence of cue-triggered reward expectations. Eur. J. Neurosci. 2017, 45, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.L.; McKay, P.F.; Seyoum, R.; Milbourne, D.; Yin, W.; Sarma, P.; Cook, J.M.; June, H.L. GABAA and opioid receptors of the central nucleus of the amygdala selectively regulate ethanol-maintained behaviors. Neuropsychopharmacology 2004, 29, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Kang-Park, M.; Kieffer, B.L.; Roberts, A.J.; Siggins, G.R.; Moore, S.D. κ-Opioid receptors in the central amygdala regulate ethanol actions at presynaptic GABAergic sites. J. Pharm. Exp. 2013, 346, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Roberto, M.; Madamba, S.G.; Moore, S.D.; Tallent, M.K.; Siggins, G.R. Ethanol increases GABAergic transmission at both pre-and postsynaptic sites in rat central amygdala neurons. Proc. Natl. Acad. Sci. USA 2003, 100, 2053–2058. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Muller, J.F.; McDonald, A.J. Mu opioid receptor localization in the basolateral amygdala: An ultrastructural analysis. Neuroscience 2015, 303, 352–363. [Google Scholar] [CrossRef]
- Muller, J.F.; Mascagni, F.; McDonald, A.J. Postsynaptic targets of somatostatin-containing interneurons in the rat basolateral amygdala. J. Comp. Neurol. 2007, 500, 513–529. [Google Scholar] [CrossRef]
- West, A.M.; Holleran, K.M.; Jones, S.R. Kappa Opioid Receptors Reduce Serotonin Uptake and Escitalopram Efficacy in the Mouse Substantia Nigra Pars Reticulata. Int. J. Mol. Sci. 2023, 24, 2080. [Google Scholar] [CrossRef]
- Commons, K.G.; Milner, T.A. Localization of delta opioid receptor immunoreactivity in interneurons and pyramidal cells in the rat hippocampus. J. Comp. Neurol. 1997, 381, 373–387. [Google Scholar] [CrossRef]
- Bayassi-Jakowicka, M.; Lietzau, G.; Czuba, E.; Steliga, A.; Waśkow, M.; Kowiański, P. Neuroplasticity and multilevel system of connections determine the integrative role of nucleus accumbens in the brain reward system. Int. J. Mol. Sci. 2021, 22, 9806. [Google Scholar] [CrossRef] [PubMed]
- Przybysz, K.R.; Gamble, M.E.; Diaz, M.R. Moderate adolescent chronic intermittent ethanol exposure sex-dependently disrupts synaptic transmission and kappa opioid receptor function in the basolateral amygdala of adult rats. Neuropharmacology 2021, 188, 108512. [Google Scholar] [CrossRef] [PubMed]
- Drake, C.T.; Chavkin, C.; Milner, T.A. Opioid systems in the dentate gyrus. Prog. Brain Res. 2007, 163, 245–814. [Google Scholar] [PubMed]
- He, J.; Kleyman, M.; Chen, J.; Alikaya, A.; Rothenhoefer, K.M.; Ozturk, B.E.; Wirthlin, M.; Bostan, A.C.; Fish, K.; Byrne, L.C. Transcriptional and anatomical diversity of medium spiny neurons in the primate striatum. Curr. Biol. 2021, 31, 5473–5486.e6. [Google Scholar] [CrossRef]
- Piskorowski, R.A.; Chevaleyre, V. Delta-opioid receptors mediate unique plasticity onto parvalbumin-expressing interneurons in area CA2 of the hippocampus. J. Neurosci. 2013, 33, 14567–14578. [Google Scholar] [CrossRef]
- Jiang, C.; Wang, X.; Le, Q.; Liu, P.; Liu, C.; Wang, Z.; He, G.; Zheng, P.; Wang, F.; Ma, L. Morphine coordinates SST and PV interneurons in the prelimbic cortex to disinhibit pyramidal neurons and enhance reward. Mol. Psychiatry 2021, 26, 1178–1193. [Google Scholar] [CrossRef] [PubMed]
- Drake, C.T.; Milner, T.A. Mu opioid receptors are extensively co-localized with parvalbumin, but not somatostatin, in the dentate gyrus. Neurosci. Lett. 2006, 403, 176–180. [Google Scholar] [CrossRef]
- Shao, C.; Chen, P.; Chen, Q.; Zhao, M.; Zhang, W.-N.; Yang, K. Mu opioid receptors inhibit GABA release from parvalbumin interneuron terminals onto CA1 pyramidal cells. Biochem. Biophys. Res. Commun. 2020, 522, 1059–1062. [Google Scholar] [CrossRef]
- Lau, B.K.; Ambrose, B.P.; Thomas, C.S.; Qiao, M.; Borgland, S.L. Mu-opioids suppress GABAergic synaptic transmission onto orbitofrontal cortex pyramidal neurons with subregional selectivity. J. Neurosci. 2020, 40, 5894–5907. [Google Scholar] [CrossRef]
- Trifu, S.; Drăgan-Serban, F.; Jianu, E. Neurotransmitters, neuromodellers, anatomical structures involved in psychiatric pathologies. Recent. Pharm. 2021, 147, 144–174. [Google Scholar]
- Dutkiewicz, A.P.; Morielli, A.D. μ Opioid Receptors Modulate Action Potential Kinetics and Firing Frequency in Neocortical Interneurons. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zamfir, M.; Sharif, B.; Locke, S.; Ehrlich, A.T.; Ochandarena, N.E.; Scherrer, G.; Ribeiro-da-Silva, A.; Kieffer, B.L.; Séguéla, P. Distinct and sex-specific expression of mu opioid receptors in anterior cingulate and somatosensory S1 cortical areas. Pain 2023, 164, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.M.; Cross, J.; Eraslan, I.M.; Attawar, A.; Ch’ng, S.; Dhar, P.; Samarasinghe, R.; Gray, L.; Lawrence, A.J. Phenotyping neurons activated in the mouse brain during restoration of salt debt. J. Chem. Neuroanat. 2019, 101, 101665. [Google Scholar] [CrossRef] [PubMed]
- Capogna, M.; Gahwiler, B.H.; Thompson, S.M. Presynaptic inhibition of calcium-dependent and-independent release elicited with ionomycin, gadolinium, and alpha-latrotoxin in the hippocampus. J. Neurophysiol. 1996, 75, 2017–2028. [Google Scholar] [CrossRef]
- Lutz, P.-E.; Kieffer, B.L. Opioid receptors: Distinct roles in mood disorders. Trends Neurosci. 2013, 36, 195–206. [Google Scholar] [CrossRef]
- Hörtnagl, H.; Tasan, R.O.; Wieselthaler, A.; Kirchmair, E.; Sieghart, W.; Sperk, G. Patterns of mRNA and protein expression for 12 GABAA receptor subunits in the mouse brain. Neuroscience 2013, 236, 345–372. [Google Scholar] [CrossRef]
- Lingford-Hughes, A.; Myers, J.; Watson, B.; Reid, A.G.; Kalk, N.; Feeney, A.; Hammers, A.; Riaño-Barros, D.A.; McGinnity, C.J.; Taylor, L.G. Using [11C] Ro15 4513 PET to characterise GABA-benzodiazepine receptors in opiate addiction: Similarities and differences with alcoholism. Neuroimage 2016, 132, 1–7. [Google Scholar] [CrossRef]
- Schoffelmeer, A.N.; Wardeh, G.; Vanderschuren, L.J. Morphine acutely and persistently attenuates nonvesicular GABA release in rat nucleus accumbens. Synapse 2001, 42, 87–94. [Google Scholar] [CrossRef]
- Sasaki, K.; Fan, L.-W.; Tien, L.-T.; Ma, T.; Loh, H.H.; Ho, K. The interaction of morphine and γ-aminobutyric acid (GABA) ergic systems in anxiolytic behavior: Using μ-opioid receptor knockout mice. Brain Res. Bull. 2002, 57, 689–694. [Google Scholar] [CrossRef]
- Kim, Y.R.; Shim, H.G.; Kim, C.-E.; Kim, S.J. The effect of µ-opioid receptor activation on GABAergic neurons in the spinal dorsal horn. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2018, 22, 419. [Google Scholar] [CrossRef]
- Xi, Z.X.; Stein, E.A. Increased mesolimbic GABA concentration blocks heroin self-administration in the rat. J. Pharm. Exp. 2000, 294, 613–619. [Google Scholar]
- He, X.J.; Patel, J.; Weiss, C.E.; Ma, X.; Bloodgood, B.L.; Banghart, M.R. Convergent, functionally independent signaling by mu and delta opioid receptors in hippocampal parvalbumin interneurons. eLife 2021, 10, e69746. [Google Scholar] [CrossRef] [PubMed]
- Der-Avakian, A.; Markou, A. The neurobiology of anhedonia and other reward-related deficits. Trends Neurosci. 2012, 35, 68–77. [Google Scholar] [CrossRef]
- Bali, A.; Randhawa, P.K.; Jaggi, A.S. Stress and opioids: Role of opioids in modulating stress-related behavior and effect of stress on morphine conditioned place preference. Neurosci. Biobehav. Rev. 2015, 51, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Theile, J.W.; Morikawa, H.; Gonzales, R.A.; Morrisett, R.A. Ethanol enhances GABAergic transmission onto dopamine neurons in the ventral tegmental area of the rat. Alcohol. Clin. Exp. Res. 2008, 32, 1040–1048. [Google Scholar] [CrossRef]
- Kelm, M.K.; Criswell, H.E.; Breese, G.R. Ethanol-enhanced GABA release: A focus on G protein-coupled receptors. Brain Res. Rev. 2011, 65, 113–123. [Google Scholar] [CrossRef]
- Matsui, A.; Jarvie, B.C.; Robinson, B.G.; Hentges, S.T.; Williams, J.T. Separate GABA afferents to dopamine neurons mediate acute action of opioids, development of tolerance, and expression of withdrawal. Neuron 2014, 82, 1346–1356. [Google Scholar] [CrossRef]
- Steffensen, S.C.; Stobbs, S.H.; Colago, E.E.; Lee, R.-S.; Koob, G.F.; Gallegos, R.A.; Henriksen, S.J. Contingent and non-contingent effects of heroin on mu-opioid receptor-containing ventral tegmental area GABA neurons. Exp. Neurol. 2006, 202, 139–151. [Google Scholar] [CrossRef]
- Speciale, S.; Manaye, K.; Sadeq, M.; German, D. Opioid receptors in midbrain dopaminergic regions of the rat II. Kappa and delta receptor autoradiography. J. Neural Transm./Gen. Sect. JNT 1993, 91, 53–66. [Google Scholar] [CrossRef]
- Pirino, B.E.; Spodnick, M.B.; Gargiulo, A.T.; Curtis, G.R.; Barson, J.R.; Karkhanis, A.N. Kappa-opioid receptor-dependent changes in dopamine and anxiety-like or approach-avoidance behavior occur differentially across the nucleus accumbens shell rostro-caudal axis. Neuropharmacology 2020, 181, 108341. [Google Scholar] [CrossRef]
- Edwards, N.J.; Tejeda, H.A.; Pignatelli, M.; Zhang, S.; McDevitt, R.A.; Wu, J.; Bass, C.E.; Bettler, B.; Morales, M.; Bonci, A. Circuit specificity in the inhibitory architecture of the VTA regulates cocaine-induced behavior. Nat. Neurosci. 2017, 20, 438–448. [Google Scholar] [CrossRef]
- Margolis, E.B.; Hjelmstad, G.O.; Bonci, A.; Fields, H.L. κ-Opioid agonists directly inhibit midbrain dopaminergic neurons. J. Neurosci. 2003, 23, 9981–9986. [Google Scholar] [CrossRef] [PubMed]
- Bergevin, A.; Girardot, D.; Bourque, M.-J.; Trudeau, L.-E. Presynaptic μ-opioid receptors regulate a late step of the secretory process in rat ventral tegmental area GABAergic neurons. Neuropharmacology 2002, 42, 1065–1078. [Google Scholar] [CrossRef] [PubMed]
- Hjelmstad, G.O.; Xia, Y.; Margolis, E.B.; Fields, H.L. Opioid modulation of ventral pallidal afferents to ventral tegmental area neurons. J. Neurosci. 2013, 33, 6454–6459. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.; Wessendorf, M.; Williams, J. A subset of ventral tegmental area neurons is inhibited by dopamine, 5-hydroxytryptamine and opioids. Neuroscience 1997, 77, 155–166. [Google Scholar] [CrossRef]
- Pina, M.M.; Pati, D.; Hwa, L.S.; Wu, S.Y.; Mahoney, A.A.; Omenyi, C.G.; Navarro, M.; Kash, T.L. The kappa opioid receptor modulates GABA neuron excitability and synaptic transmission in midbrain projections from the insular cortex. Neuropharmacology 2020, 165, 107831. [Google Scholar] [CrossRef]
- Yokota, E.; Koyanagi, Y.; Yamamoto, K.; Oi, Y.; Koshikawa, N.; Kobayashi, M. Opioid subtype-and cell-type-dependent regulation of inhibitory synaptic transmission in the rat insular cortex. Neuroscience 2016, 339, 478–490. [Google Scholar] [CrossRef]
- Huo, F.-Q.; Wang, J.; Li, Y.-Q.; Chen, T.; Han, F.; Tang, J.-S. GABAergic neurons express μ-opioid receptors in the ventrolateral orbital cortex of the rat. Neurosci. Lett. 2005, 382, 265–268. [Google Scholar]
- Lupica, C.R. Delta and mu enkephalins inhibit spontaneous GABA-mediated IPSCs via a cyclic AMP-independent mechanism in the rat hippocampus. J. Neurosci. 1995, 15, 737–749. [Google Scholar] [CrossRef]
- Krook-Magnuson, E.; Luu, L.; Lee, S.-H.; Varga, C.; Soltesz, I. Ivy and neurogliaform interneurons are a major target of μ-opioid receptor modulation. J. Neurosci. 2011, 31, 14861–14870. [Google Scholar] [CrossRef]
- Fan, K.-M.; Qiu, L.-J.; Ma, N.; Du, Y.-N.; Qian, Z.-Q.; Wei, C.-L.; Han, J.; Ren, W.; Shi, M.-M.; Liu, Z.-Q. Acute stress facilitates LTD induction at glutamatergic synapses in the hippocampal CA1 region by activating μ-opioid receptors on GABAergic neurons. Front. Neurosci. 2019, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Lafourcade, C.A.; Alger, B.E. Distinctions among GABA A and GABA B responses revealed by calcium channel antagonists, cannabinoids, opioids, and synaptic plasticity in rat hippocampus. Psychopharmacology 2008, 198, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Capogna, M.; Gähwiler, B.; Thompson, S. Mechanism of mu-opioid receptor-mediated presynaptic inhibition in the rat hippocampus in vitro. J. Physiol. 1993, 470, 539–558. [Google Scholar] [CrossRef] [PubMed]
- Terman, G.W.; Drake, C.T.; Simmons, M.L.; Milner, T.A.; Chavkin, C. Opioid modulation of recurrent excitation in the hippocampal dentate gyrus. J. Neurosci. 2000, 20, 4379–4388. [Google Scholar] [CrossRef]
- Li, C.; Pleil, K.E.; Stamatakis, A.M.; Busan, S.; Vong, L.; Lowell, B.B.; Stuber, G.D.; Kash, T.L. Presynaptic inhibition of gamma-aminobutyric acid release in the bed nucleus of the stria terminalis by kappa opioid receptor signaling. Biol. Psychiatry 2012, 71, 725–732. [Google Scholar] [CrossRef]
- Kupchik, Y.M.; Scofield, M.D.; Rice, K.C.; Cheng, K.; Roques, B.P.; Kalivas, P.W. Cocaine dysregulates opioid gating of GABA neurotransmission in the ventral pallidum. J. Neurosci. 2014, 34, 1057–1066. [Google Scholar] [CrossRef]
- Banasr, M.; Lepack, A.; Fee, C.; Duric, V.; Maldonado-Aviles, J.; DiLeone, R.; Sibille, E.; Duman, R.S.; Sanacora, G. Characterization of GABAergic marker expression in the chronic unpredictable stress model of depression. Chronic Stress 2017, 1, 2470547017720459. [Google Scholar] [CrossRef]
- Udakis, M.; Pedrosa, V.; Chamberlain, S.E.; Clopath, C.; Mellor, J.R. Interneuron-specific plasticity at parvalbumin and somatostatin inhibitory synapses onto CA1 pyramidal neurons shapes hippocampal output. Nat. Commun. 2020, 11, 4395. [Google Scholar] [CrossRef]
- Lazarus, M.S.; Krishnan, K.; Huang, Z.J. GAD67 deficiency in parvalbumin interneurons produces deficits in inhibitory transmission and network disinhibition in mouse prefrontal cortex. Cereb. Cortex 2015, 25, 1290–1296. [Google Scholar] [CrossRef]
- Margolis, E.B.; Toy, B.; Himmels, P.; Morales, M.; Fields, H.L. Identification of rat ventral tegmental area GABAergic neurons. PLoS ONE 2012, 7, e42365. [Google Scholar] [CrossRef]
- Ronström, J.W.; Johnson, N.L.; Jones, S.T.; Werner, S.J.; Wadsworth, H.A.; Brundage, J.N.; Stolp, V.; Graziane, N.M.; Silberman, Y.; Steffensen, S.C. Opioid-Induced Reductions in Amygdala Lateral Paracapsular GABA Neuron Circuit Activity. Int. J. Mol. Sci. 2023, 24, 1929. [Google Scholar] [CrossRef] [PubMed]
- Seno, F.; Sgobbi, R.; Nobre, M. Contributions of the GABAergic system of the prelimbic cortex and basolateral amygdala to morphine withdrawal-induced contextual fear. Physiol. Behav. 2022, 254, 113868. [Google Scholar] [CrossRef] [PubMed]
- Galaj, E.; Han, X.; Shen, H.; Jordan, C.J.; He, Y.; Humburg, B.; Bi, G.-H.; Xi, Z.-X. Dissecting the role of GABA neurons in the VTA versus SNr in opioid reward. J. Neurosci. 2020, 40, 8853–8869. [Google Scholar] [CrossRef] [PubMed]
- Ntamati, N.R.; Lüscher, C. VTA projection neurons releasing GABA and glutamate in the dentate gyrus. eNeuro 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Gomperts, S.N.; Kloosterman, F.; Wilson, M.A. VTA neurons coordinate with the hippocampal reactivation of spatial experience. eLife 2015, 4, e05360. [Google Scholar] [CrossRef]
- Nam, M.-H.; Won, W.; Han, K.-S.; Lee, C.J. Signaling mechanisms of μ-opioid receptor (MOR) in the hippocampus: Disinhibition versus astrocytic glutamate regulation. Cell. Mol. Life Sci. 2021, 78, 415–426. [Google Scholar] [CrossRef]
- Kecskés, A.; Pohóczky, K.; Kecskés, M.; Varga, Z.V.; Kormos, V.; Szőke, É.; Henn-Mike, N.; Fehér, M.; Kun, J.; Gyenesei, A. Characterization of neurons expressing the novel analgesic drug target somatostatin receptor 4 in mouse and human brains. Int. J. Mol. Sci. 2020, 21, 7788. [Google Scholar] [CrossRef]
- Holly, E.N.; Davatolhagh, M.F.; Choi, K.; Alabi, O.O.; Cifuentes, L.V.; Fuccillo, M.V. Striatal low-threshold spiking interneurons regulate goal-directed learning. Neuron 2019, 103, 92–101.e106. [Google Scholar] [CrossRef]
- Jeong, H.; Kim, D.; Song, M.; Paik, S.-B.; Jung, M.W. Distinct roles of parvalbumin-and somatostatin-expressing neurons in flexible representation of task variables in the prefrontal cortex. Prog. Neurobiol. 2020, 187, 101773. [Google Scholar] [CrossRef]
- Riedemann, T. Diversity and function of somatostatin-expressing interneurons in the cerebral cortex. Int. J. Mol. Sci. 2019, 20, 2952. [Google Scholar] [CrossRef]
- Cattaneo, S.; Zaghi, M.; Maddalena, R.; Bedogni, F.; Sessa, A.; Taverna, S. Somatostatin-expressing interneurons co-release GABA and glutamate onto different postsynaptic targets in the striatum. bioRxiv 2019, 566984. [Google Scholar] [CrossRef]
- Robinson, S.L.; Thiele, T.E. A role for the neuropeptide somatostatin in the neurobiology of behaviors associated with substances abuse and affective disorders. Neuropharmacology 2020, 167, 107983. [Google Scholar] [CrossRef] [PubMed]
- Grimm, C.; Balsters, J.H.; Zerbi, V. Shedding light on social reward circuitry:(un) common blueprints in humans and rodents. NeuroSci 2021, 27, 159–183. [Google Scholar] [CrossRef]
- Bouarab, C.; Thompson, B.; Polter, A.M. VTA GABA neurons at the interface of stress and reward. Front. Neural Circuits 2019, 13, 78. [Google Scholar] [CrossRef]
- Nagaeva, E.; Zubarev, I.; Bengtsson Gonzales, C.; Forss, M.; Nikouei, K.; de Miguel, E.; Elsilä, L.; Linden, A.-M.; Hjerling-Leffler, J.; Augustine, G.J. Heterogeneous somatostatin-expressing neuron population in mouse ventral tegmental area. eLife 2020, 9, e59328. [Google Scholar] [CrossRef] [PubMed]
- Kántás, B.; Szőke, É.; Börzsei, R.; Bánhegyi, P.; Asghar, J.; Hudhud, L.; Steib, A.; Hunyady, Á.; Horváth, Á.; Kecskés, A. In Silico, In Vitro and In Vivo Pharmacodynamic Characterization of Novel Analgesic Drug Candidate Somatostatin SST4 Receptor Agonists. Front. Pharmacol. 2021, 11, 601887. [Google Scholar] [CrossRef]
- Thompson, J.M.; Neugebauer, V. Cortico-limbic pain mechanisms. Neurosci. Lett. 2019, 702, 15–23. [Google Scholar] [CrossRef]
- Shao, C.; Dong, J.; Zhao, M.; Liu, S.; Wang, X.; Yu, Y.; Fang, L.; Zhu, Z.; Chen, Q.; Xiao, X. Presynaptic GABAB receptors differentially modulate GABA release from cholecystokinin and parvalbumin interneurons onto CA1 pyramidal neurons: A cell type-specific labeling and activating study. Neurosci. Lett. 2022, 772, 136448. [Google Scholar] [CrossRef]
- Nasrallah, K.; Therreau, L.; Robert, V.; Huang, A.J.; McHugh, T.J.; Piskorowski, R.A.; Chevaleyre, V. Routing hippocampal information flow through parvalbumin interneuron plasticity in area CA2. Cell Rep. 2019, 27, 86–98.e83. [Google Scholar] [CrossRef]
- Hitti, F.L.; Siegelbaum, S.A. The hippocampal CA2 region is essential for social memory. Nature 2014, 508, 88–92. [Google Scholar] [CrossRef]
- Graham, K.; Spruston, N.; Bloss, E.B. Hippocampal and thalamic afferents form distinct synaptic microcircuits in the mouse infralimbic frontal cortex. Cell Rep. 2021, 37, 109837. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ahmed, S.; Neagu, G.; Wang, Y.; Li, Z.; Wen, J.; Liu, C.; Vreugdenhil, M. μ-Opioid receptor activation modulates CA3-to-CA1 gamma oscillation phase-coupling. IBRO Rep. 2019, 6, 122–131. [Google Scholar] [CrossRef]
- Vollmer, K.M.; Green, L.M.; Grant, R.I.; Winston, K.T.; Doncheck, E.M.; Bowen, C.W.; Paniccia, J.E.; Clarke, R.E.; Tiller, A.; Siegler, P.N. An opioid-gated thalamoaccumbal circuit for the suppression of reward seeking in mice. Nat. Commun. 2022, 13, 6865. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, P.; Ma, L.; Wang, F. Opioid signal transduction regulates the dendritic morphology of somatostatin and parvalbumin interneurons in the medial prefrontal cortex. Neuroreport 2019, 30, 592–599. [Google Scholar] [CrossRef]
- McMahon, A.; Wong, B.S.; Iacopino, A.M.; Ng, M.C.; Chi, S.; German, D.C. Calbindin-D28k buffers intracellular calcium and promotes resistance to degeneration in PC12 cells. Mol. Brain Res. 1998, 54, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Pert, C.B.; Kuhar, M.J.; Snyder, S.H. Opiate receptor: Autoradiographic localization in rat brain. Proc. Natl. Acad. Sci. USA 1976, 73, 3729–3733. [Google Scholar] [CrossRef]
- Castro, D.C.; Bruchas, M.R. A motivational and neuropeptidergic hub: Anatomical and functional diversity within the nucleus accumbens shell. Neuron 2019, 102, 529–552. [Google Scholar] [CrossRef]
- Pan, J.; Yu, J.; Sun, L.; Xie, C.; Chang, L.; Wu, J.; Hawes, S.; Saez–Atienzar, S.; Zheng, W.; Kung, J. ALDH1A1 regulates postsynaptic μ–opioid receptor expression in dorsal striatal projection neurons and mitigates dyskinesia through transsynaptic retinoic acid signaling. Sci. Rep. 2019, 9, 1–14. [Google Scholar]
- Zhou, K.; Zhu, Y. The paraventricular thalamic nucleus: A key hub of neural circuits underlying drug addiction. Pharmacol. Res. 2019, 142, 70–76. [Google Scholar] [CrossRef]
- Hu, H.; Cui, Y.; Yang, Y. Circuits and functions of the lateral habenula in health and in disease. Nat. Rev. Neurosci. 2020, 21, 277–295. [Google Scholar] [CrossRef]
- Kingsbury, M.A. New perspectives on vasoactive intestinal polypeptide as a widespread modulator of social behavior. Curr. Opin. Behav. Sci. 2015, 6, 139–147. [Google Scholar] [CrossRef]
- Achilly, N.P. Properties of VIP+ synapses in the suprachiasmatic nucleus highlight their role in circadian rhythm. J. Neurophysiol. 2016, 115, 2701–2704. [Google Scholar] [CrossRef] [PubMed]
- Férézou, I.; Hill, E.L.; Cauli, B.; Gibelin, N.; Kaneko, T.; Rossier, J.; Lambolez, B. Extensive overlap of mu-opioid and nicotinic sensitivity in cortical interneurons. Cereb. Cortex 2007, 17, 1948–1957. [Google Scholar] [CrossRef]
- Dupret, D.; O’Neill, J.; Csicsvari, J. Dynamic reconfiguration of hippocampal interneuron circuits during spatial learning. Neuron 2013, 78, 166–180. [Google Scholar] [CrossRef]
- Poulin, J.-F.; Zou, J.; Drouin-Ouellet, J.; Kim, K.-Y.A.; Cicchetti, F.; Awatramani, R.B. Defining midbrain dopaminergic neuron diversity by single-cell gene expression profiling. Cell Rep. 2014, 9, 930–943. [Google Scholar] [CrossRef]
- Newmyer, B.A.; Whindleton, C.M.; Klein, P.M.; Beenhakker, M.P.; Jones, M.K.; Scott, M.M. VIPergic neurons of the infralimbic and prelimbic cortices control palatable food intake through separate cognitive pathways. JCI Insight 2019, 4, e126283. [Google Scholar] [CrossRef]
- Luo, X.; Muñoz-Pino, E.; Francavilla, R.; Vallée, M.; Droit, A.; Topolnik, L. Transcriptomic profile of the subiculum-projecting VIP GABAergic neurons in the mouse CA1 hippocampus. Brain Struct. Funct. 2019, 224, 2269–2280. [Google Scholar] [CrossRef] [PubMed]
- Berríos-Cárcamo, P.; Quezada, M.; Santapau, D.; Morales, P.; Olivares, B.; Ponce, C.; Ávila, A.; De Gregorio, C.; Ezquer, M.; Quintanilla, M.E. A novel morphine drinking model of opioid dependence in rats. Int. J. Mol. Sci. 2022, 23, 3874. [Google Scholar] [CrossRef]
- Viena, T.D.; Rasch, G.E.; Silva, D.; Allen, T.A. Calretinin and calbindin architecture of the midline thalamus associated with prefrontal–hippocampal circuitry. Hippocampus 2021, 31, 770–789. [Google Scholar] [CrossRef]
- Maal-Bared, G. Identifying the Anatomical Layout and Molecular Properties of the GABA A Motivational Switch in the Ventral Tegmental Area; University of Toronto (Canada): Toronto, ON, Canada, 2021. [Google Scholar]
- Porcino, C.; Briglia, M.; Aragona, M.; Mhalhel, K.; Laurà, R.; Levanti, M.; Abbate, F.; Montalbano, G.; Germanà, G.; Lauriano, E.R. Potential Neuroprotective Role of Calretinin-N18 and Calbindin-D28k in the Retina of Adult Zebrafish Exposed to Different Wavelength Lights. Int. J. Mol. Sci. 2023, 24, 1087. [Google Scholar] [CrossRef] [PubMed]
- Carney, A.E.; Clarke, C.; Pratt, W.E. Administration of Neuropeptide Y into the rat nucleus accumbens shell, but not core, attenuates the motivational impairment from systemic dopamine receptor antagonism by α-flupenthixol. Neurosci. Lett. 2023, 797, 137069. [Google Scholar] [CrossRef]
- You, Z.-B.; Gardner, E.L.; Galaj, E.; Moore, A.R.; Buck, T.; Jordan, C.J.; Humburg, B.A.; Bi, G.-H.; Xi, Z.-X.; Leggio, L. Involvement of the ghrelin system in the maintenance of oxycodone self-administration: Converging evidence from endocrine, pharmacologic and transgenic approaches. Mol. Psychiatry 2022, 27, 2171–2181. [Google Scholar] [CrossRef] [PubMed]
- Elvig, S.K.; McGinn, M.A.; Smith, C.; Arends, M.A.; Koob, G.F.; Vendruscolo, L.F. Tolerance to alcohol: A critical yet understudied factor in alcohol addiction. Pharmacol. Biochem. Behav. 2021, 204, 173155. [Google Scholar] [CrossRef]
- Rezitis, J.; Herzog, H.; Ip, C.K. Neuropeptide Y interaction with dopaminergic and serotonergic pathways: Interlinked neurocircuits modulating hedonic eating behaviours. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 113, 110449. [Google Scholar] [CrossRef]
- Maric, T.; Tobin, S.; Quinn, T.; Shalev, U. Food deprivation-like effects of neuropeptide Y on heroin self-administration and reinstatement of heroin seeking in rats. Behav. Brain Res. 2008, 194, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Boey, D.; Lee, N.; Schwarzer, C.; Sainsbury, A.; Herzog, H. Distribution of prodynorphin mRNA and its interaction with the NPY system in the mouse brain. Neuropeptides 2006, 40, 115–123. [Google Scholar] [CrossRef]
- Ghule, A.; Rácz, I.; Bilkei-Gorzo, A.; Leidmaa, E.; Sieburg, M.; Zimmer, A. Modulation of feeding behavior and metabolism by dynorphin. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Warthen, K.G.; Sanford, B.; Walker, K.; Jones, K.G.; Angstadt, M.; Sripada, C.; Goldman, D.; Zubieta, J.-K.; Welsh, R.C.; Burmeister, M. Neuropeptide Y and representation of salience in human nucleus accumbens. Neuropsychopharmacology 2019, 44, 495–502. [Google Scholar] [CrossRef]
- Maffei, A. The many forms and functions of long term plasticity at GABAergic synapses. Neural Plast. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nugent, F.S.; Penick, E.C.; Kauer, J.A. Opioids block long-term potentiation of inhibitory synapses. Nature 2007, 446, 1086–1090. [Google Scholar] [CrossRef]
- Kullmann, D.M.; Lamsa, K.P. LTP and LTD in cortical GABAergic interneurons: Emerging rules and roles. Neuropharmacology 2011, 60, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.-Z.; Ye, J.-H. Ethanol blocks long-term potentiation of GABAergic synapses in the ventral tegmental area involving μ-opioid receptors. Neuropsychopharmacology 2010, 35, 1841–1849. [Google Scholar] [CrossRef] [PubMed]
- Matsui, A.; Williams, J.T. Opioid-sensitive GABA inputs from rostromedial tegmental nucleus synapse onto midbrain dopamine neurons. J. Neurosci. 2011, 31, 17729–17735. [Google Scholar] [CrossRef]
- Laurent, R.S.; Damonte, V.M.; Tsuda, A.C.; Kauer, J.A. Periaqueductal gray and rostromedial tegmental inhibitory afferents to VTA have distinct synaptic plasticity and opiate sensitivity. Neuron 2020, 106, 624–636.e4. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, Y.; Jin, S.; Lin, L.; Duan, S.; Si, K.; Gong, W.; Julius Zhu, J. Organizational principles of amygdalar input-output neuronal circuits. Mol. Psychiatry 2021, 26, 7118–7129. [Google Scholar] [CrossRef] [PubMed]
- Blaesse, P.; Goedecke, L.; Bazelot, M.; Capogna, M.; Pape, H.-C.; Jüngling, K. μ-Opioid receptor-mediated inhibition of intercalated neurons and effect on synaptic transmission to the central amygdala. J. Neurosci. 2015, 35, 7317–7325. [Google Scholar] [CrossRef]
- Finnegan, T.F.; Chen, S.-R.; Pan, H.-L. μ Opioid receptor activation inhibits GABAergic inputs to basolateral amygdala neurons through Kv1. 1/1.2 channels. J. Neurophysiol. 2006, 95, 2032–2041. [Google Scholar] [CrossRef]
- Crowley, N.A.; Bloodgood, D.W.; Hardaway, J.A.; Kendra, A.M.; McCall, J.G.; Al-Hasani, R.; McCall, N.M.; Yu, W.; Schools, Z.L.; Krashes, M.J. Dynorphin controls the gain of an amygdalar anxiety circuit. Cell Rep. 2016, 14, 2774–2783. [Google Scholar] [CrossRef]
- Huge, V.; Rammes, G.; Beyer, A.; Zieglgänsberger, W.; Azad, S.C. Activation of kappa opioid receptors decreases synaptic transmission and inhibits long-term potentiation in the basolateral amygdala of the mouse. Eur. J. Pain 2009, 13, 124–129. [Google Scholar] [CrossRef]
- Klenowski, P.; Morgan, M.; Bartlett, S.E. The role of δ-opioid receptors in learning and memory underlying the development of addiction. Br. J. Pharmacol. 2015, 172, 297–310. [Google Scholar] [CrossRef]
- Pieretti, S.; Di Giannuario, A.; Domenici, M.; Sagratella, S.; Capasso, A.; Sorrentino, L.; Loizzo, A. Dexamethasone-induced selective inhibition of the central μ opioid receptor: Functional in vivo and in vitro evidence in rodents. Br. J. Pharmacol. 1994, 113, 1416–1422. [Google Scholar] [CrossRef] [PubMed]
- Banghart, M.R.; Neufeld, S.Q.; Wong, N.C.; Sabatini, B.L. Enkephalin disinhibits mu opioid receptor-rich striatal patches via delta opioid receptors. Neuron 2015, 88, 1227–1239. [Google Scholar] [CrossRef] [PubMed]
- McDermott, C.M.; Schrader, L.A. Activation of κ opioid receptors increases intrinsic excitability of dentate gyrus granule cells. J. Physiol. 2011, 589, 3517–3532. [Google Scholar] [CrossRef] [PubMed]
- Nadeau, S.E. Neural mechanisms of emotions, alexithymia, and depression. Handb. Clin. Neurol. 2021, 183, 299–313. [Google Scholar]
- Rolls, E.T. The cingulate cortex and limbic systems for emotion, action, and memory. Brain Struct. Funct. 2019, 224, 3001–3018. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, G.; Sterlini, B.; Tardito, S.; Inglese, M.; Corradi, A.; Filaci, G.; Amore, M.; Magioncalda, P.; Martino, M. Opioidergic system and functional architecture of intrinsic brain activity: Implications for psychiatric disorders. Neurosci. 2020, 26, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Birdsong, W.T.; Jongbloets, B.C.; Engeln, K.A.; Wang, D.; Scherrer, G.; Mao, T. Synapse-specific opioid modulation of thalamo-cortico-striatal circuits. eLife 2019, 8, e45146. [Google Scholar] [CrossRef]
- Casello, S.M.; Flores, R.J.; Yarur, H.E.; Wang, H.; Awanyai, M.; Arenivar, M.A.; Jaime-Lara, R.B.; Bravo-Rivera, H.; Tejeda, H.A. Neuropeptide system regulation of prefrontal cortex circuitry: Implications for neuropsychiatric disorders. Front. Neural Circuits 2022, 16, 796443. [Google Scholar] [CrossRef]
- Tejeda, H.A.; Counotte, D.S.; Oh, E.; Ramamoorthy, S.; Schultz-Kuszak, K.N.; Bäckman, C.M.; Chefer, V.; O’donnell, P.; Shippenberg, T.S. Prefrontal cortical kappa-opioid receptor modulation of local neurotransmission and conditioned place aversion. Neuropsychopharmacology 2013, 38, 1770–1779. [Google Scholar] [CrossRef]
- Yamada, D.; Takahashi, J.; Iio, K.; Nagase, H.; Saitoh, A. Modulation of glutamatergic synaptic transmission and neuronal excitability in the prelimbic medial prefrontal cortex via delta-opioid receptors in mice. Biochem. Biophys. Res. Commun. 2021, 560, 192–198. [Google Scholar] [CrossRef]
- Rola, R.; Jarkiewicz, M.; Szulczyk, P. Modulation of Ca 2+ channel current by µ opioid receptors in prefrontal cortex pyramidal neurons in rats. Acta Neurobiol. Exp. 2008, 68, 10–25. [Google Scholar]
- Bloodgood, D.W.; Hardaway, J.A.; Stanhope, C.M.; Pati, D.; Pina, M.M.; Neira, S.; Desai, S.; Boyt, K.M.; Palmiter, R.D.; Kash, T.L. Kappa opioid receptor and dynorphin signaling in the central amygdala regulates alcohol intake. Mol. Psychiatry 2021, 26, 2187–2199. [Google Scholar] [CrossRef] [PubMed]
- Przybysz, K.; Werner, D.; Diaz, M. Age-dependent regulation of GABA transmission by kappa opioid receptors in the basolateral amygdala of Sprague-Dawley rats. Neuropharmacology 2017, 117, 124–133. [Google Scholar] [CrossRef]
- Hein, M.; Ji, G.; Tidwell, D.; D’Souza, P.; Kiritoshi, T.; Yakhnitsa, V.; Navratilova, E.; Porreca, F.; Neugebauer, V. Kappa opioid receptor activation in the amygdala disinhibits CRF neurons to generate pain-like behaviors. Neuropharmacology 2021, 185, 108456. [Google Scholar] [CrossRef]
- Dumont, É.C.; Williams, J.T. Noradrenaline triggers GABAA inhibition of bed nucleus of the stria terminalis neurons projecting to the ventral tegmental area. J. Neurosci. 2004, 24, 8198–8204. [Google Scholar] [CrossRef] [PubMed]
- Beltran-Campos, V.; Silva-Vera, M.; García-Campos, M.; Diaz-Cintra, S. Effects of morphine on brain plasticity. Neurol. ÍA (Engl. Ed.) 2015, 30, 176–180. [Google Scholar] [CrossRef]
- Kahle, K.T.; Staley, K.J.; Nahed, B.V.; Gamba, G.; Hebert, S.C.; Lifton, R.P.; Mount, D.B. Roles of the cation–chloride cotransporters in neurological disease. Nat. Clin. Pract. Neurol. 2008, 4, 490–503. [Google Scholar] [CrossRef] [PubMed]
- Korczak, M.; Kurowski, P.; Leśniak, A.; Grönbladh, A.; Filipowska, A.; Bujalska-Zadrożny, M. GABAB receptor intracellular signaling: Novel pathways for depressive disorder treatment? Eur. J. Pharmacol. 2020, 885, 173531. [Google Scholar] [CrossRef]
- Meredith, G.E.; Baldo, B.A.; Andrezjewski, M.E.; Kelley, A.E. The structural basis for mapping behavior onto the ventral striatum and its subdivisions. Brain Struct. Funct. 2008, 213, 17–27. [Google Scholar] [CrossRef]
- Karami, M.; Zarrindast, M.R. Morphine sex-dependently induced place conditioning in adult Wistar rats. Eur. J. Pharmacol. 2008, 582, 78–87. [Google Scholar] [CrossRef]
- Joshi, A.; Schott, M.; La Fleur, S.E.; Barrot, M. Role of the striatal dopamine, GABA and opioid systems in mediating feeding and fat intake. Neurosci. Biobehav. Rev. 2022, 139, 104726. [Google Scholar] [CrossRef] [PubMed]
- Sustkova-Fiserova, M.; Charalambous, C.; Khryakova, A.; Certilina, A.; Lapka, M.; Šlamberová, R. The Role of Ghrelin/GHS-R1A Signaling in Nonalcohol Drug Addictions. Int. J. Mol. Sci. 2022, 23, 761. [Google Scholar] [CrossRef]
- Fields, H.L.; Margolis, E.B. Understanding opioid reward. Trends Neurosci. 2015, 38, 217–225. [Google Scholar] [CrossRef]
- Terunuma, M. Diversity of structure and function of GABAB receptors: A complexity of GABAB-mediated signaling. Proc. Jpn. Acad. Ser. B 2018, 94, 390–411. [Google Scholar] [CrossRef]
- Al-Hasani, R.; Bruchas, M.R. Molecular mechanisms of opioid receptor-dependent signaling and behavior. J. Am. Soc. Anesthesiol. 2011, 115, 1363–1381. [Google Scholar] [CrossRef] [PubMed]
- Mattheisen, G.B.; Tsintsadze, T.; Smith, S.M. Strong G-protein-mediated inhibition of sodium channels. Cell Rep. 2018, 23, 2770–2781. [Google Scholar] [CrossRef]
- Gaiarsa, J.-L.; Porcher, C. Emerging neurotrophic role of GABAB receptors in neuronal circuit development. Front. Cell. Neurosci. 2013, 7, 206. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.; Rondard, P.; Xu, C.; Bertaso, F.; Cao, F.; Zhang, X.; Pin, J.-P.; Liu, J. Dominant role of GABAB2 and Gβγ for GABAB receptor-mediated-ERK1/2/CREB pathway in cerebellar neurons. Cell. Signal. 2007, 19, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Miyatake, M.; Rubinstein, T.J.; McLennan, G.P.; Belcheva, M.M.; Coscia, C.J. Inhibition of EGF-induced ERK/MAP kinase-mediated astrocyte proliferation by μ opioids: Integration of G protein and β-arrestin 2-dependent pathways. J. Neurochem. 2009, 110, 662–674. [Google Scholar] [CrossRef]
- Wang, Y.; Gai, S.; Zhang, W.; Huang, X.; Ma, S.; Huo, Y.; Wu, Y.; Tu, H.; Pin, J.-P.; Rondard, P. The GABAB receptor mediates neuroprotection by coupling to G13. Sci. Signal. 2021, 14, eaaz4112. [Google Scholar] [CrossRef]
- Jamshidi, R.J.; Sullivan, L.C.; Jacobs, B.A.; Chavera, T.A.; Berg, K.A.; Clarke, W.P. Long-term reduction of kappa opioid receptor function by the biased ligand, norbinaltorphimine, requires c-Jun N-terminal kinase activity and new protein synthesis in peripheral sensory neurons. J. Pharmacol. Exp. Ther. 2016, 359, 319–328. [Google Scholar] [CrossRef]
- Wamsteeker Cusulin, J.I.; Füzesi, T.; Inoue, W.; Bains, J.S. Glucocorticoid feedback uncovers retrograde opioid signaling at hypothalamic synapses. Nat. Neurosci. 2013, 16, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Bloem, B.; Poorthuis, R.B.; Mansvelder, H.D. Cholinergic modulation of the medial prefrontal cortex: The role of nicotinic receptors in attention and regulation of neuronal activity. Front. Neural Circuits 2014, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Hjelmstad, G.O.; Fields, H.L. Kappa opioid receptor activation in the nucleus accumbens inhibits glutamate and GABA release through different mechanisms. J. Neurophysiol. 2003, 89, 2389–2395. [Google Scholar] [CrossRef] [PubMed]
- Kano, H.; Toyama, Y.; Imai, S.; Iwahashi, Y.; Mase, Y.; Yokogawa, M.; Osawa, M.; Shimada, I. Structural mechanism underlying G protein family-specific regulation of G protein-gated inwardly rectifying potassium channel. Nat. Commun. 2019, 10, 2008. [Google Scholar] [CrossRef] [PubMed]
- Wimpey, T.L.; Chavkin, C. Opioids activate both an inward rectifier and a novel voltage-gated potassium conductance in the hippocampal formation. Neuron 1991, 6, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Lüscher, C.; Slesinger, P.A. Emerging roles for G protein-gated inwardly rectifying potassium (GIRK) channels in health and disease. Nat. Rev. Neurosci. 2010, 11, 301–315. [Google Scholar] [CrossRef]
- McHugh, R.K.; Taghian, N.R.; Nguyen, M.D. Anxiety and opioid use disorder: Potential targets for treatment and prevention. Curr. Addict. Rep. 2021, 8, 136–143. [Google Scholar] [CrossRef]
- Langdon, K.J.; Dove, K.; Ramsey, S. Comorbidity of opioid-related and anxiety-related symptoms and disorders. Curr. Opin. Psychol. 2019, 30, 17–23. [Google Scholar] [CrossRef]
- Umberg, E.N.; Pothos, E.N. Neurobiology of aversive states. Physiol. Behav. 2011, 104, 69–75. [Google Scholar] [CrossRef]
- Blum, K.; Chen, T.J.; Bailey, J.; Bowirrat, A.; Femino, J.; Chen, A.L.; Simpatico, T.; Morse, S.; Giordano, J.; Damle, U. Can the chronic administration of the combination of buprenorphine and naloxone block dopaminergic activity causing anti-reward and relapse potential? Mol. Neurobiol. 2011, 44, 250–268. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.P. Anticraving medications for relapse prevention: A possible new class of psychoactive medications. Am. J. Psychiatry 2005, 162, 1423–1431. [Google Scholar] [CrossRef]
- Cahill, C.M.; Walwyn, W.; Taylor, A.M.; Pradhan, A.A.; Evans, C.J. Allostatic mechanisms of opioid tolerance beyond desensitization and downregulation. Trends Pharmacol. Sci. 2016, 37, 963–976. [Google Scholar] [CrossRef]
- Russell, S.E.; Puttick, D.J.; Sawyer, A.M.; Potter, D.N.; Mague, S.; Carlezon, W.A.; Chartoff, E.H. Nucleus accumbens AMPA receptors are necessary for morphine-withdrawal-induced negative-affective states in rats. J. Neurosci. 2016, 36, 5748–5762. [Google Scholar] [CrossRef]
- Margolis, E.B.; Karkhanis, A.N. Dopaminergic cellular and circuit contributions to kappa opioid receptor mediated aversion. Neurochem. Int. 2019, 129, 104504. [Google Scholar] [CrossRef] [PubMed]
- Allouche, S.; Noble, F.; Marie, N. Opioid receptor desensitization: Mechanisms and its link to tolerance. Front. Pharmacol. 2014, 5, 280. [Google Scholar] [CrossRef] [PubMed]
- Addolorato, G.; Leggio, L.; Hopf, F.W.; Diana, M.; Bonci, A. Novel therapeutic strategies for alcohol and drug addiction: Focus on GABA, ion channels and transcranial magnetic stimulation. Neuropsychopharmacology 2012, 37, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Bagley, E.E. Opioid and GABAB receptors differentially couple to an adenylyl cyclase/protein kinase A downstream effector after chronic morphine treatment. Front. Pharmacol. 2014, 5, 148. [Google Scholar] [CrossRef]
- Gurgenidze, S.; Bäuerle, P.; Schmitz, D.; Vida, I.; Gloveli, T.; Dugladze, T. Cell-type specific inhibition controls the high-frequency oscillations in the medial entorhinal cortex. Int. J. Mol. Sci. 2022, 23, 14087. [Google Scholar] [CrossRef]
- Bastos, A.M.; Usrey, W.M.; Adams, R.A.; Mangun, G.R.; Fries, P.; Friston, K.J. Canonical microcircuits for predictive coding. Neuron 2012, 76, 695–711. [Google Scholar] [CrossRef]
- Asede, D.; Doddapaneni, D.; Bolton, M.M. Amygdala Intercalated Cells: Gate Keepers and Conveyors of Internal State to the Circuits of Emotion. J. Neurosci. 2022, 42, 9098–9109. [Google Scholar] [CrossRef]
- Asede, D.; Doddapaneni, D.; Chavez, A.; Okoh, J.; Ali, S.; Von-Walter, C.; Bolton, M.M. Apical intercalated cell cluster: A distinct sensory regulator in the amygdala. Cell Rep. 2021, 35, 109151. [Google Scholar] [CrossRef] [PubMed]
- Gregoriou, G.C.; Patel, S.D.; Pyne, S.; Winters, B.L.; Bagley, E.E. Opioid withdrawal abruptly disrupts amygdala circuit function by reducing peptide actions. J. Neurosci. 2023, 43, 1668–1681. [Google Scholar] [CrossRef] [PubMed]
- Gregoriou, G.C.; Kissiwaa, S.A.; Patel, S.D.; Bagley, E.E. Dopamine and opioids inhibit synaptic outputs of the main island of the intercalated neurons of the amygdala. Eur. J. Neurosci. 2019, 50, 2065–2074. [Google Scholar] [CrossRef]
- Xie, L.; Wu, H.; Chen, Q.; Xu, F.; Li, H.; Xu, Q.; Jiao, C.; Sun, L.; Ullah, R.; Chen, X. Divergent modulation of pain and anxiety by GABAergic neurons in the ventrolateral periaqueductal gray and dorsal raphe. Neuropsychopharmacology 2022, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Yakel, J.L. Cholinergic Regulation of Hippocampal Theta Rhythm. Biomedicines 2022, 10, 745. [Google Scholar] [CrossRef] [PubMed]
- Mann, E.O.; Paulsen, O. Role of GABAergic inhibition in hippocampal network oscillations. Trends Neurosci. 2007, 30, 343–349. [Google Scholar] [CrossRef]
- Alger, B.E.; Nagode, D.A.; Tang, A.-H. Muscarinic cholinergic receptors modulate inhibitory synaptic rhythms in hippocampus and neocortex. Front. Synaptic Neurosci. 2014, 6, 18. [Google Scholar] [CrossRef]
- Ieong, H.F.-H.; Gao, F.; Yuan, Z. Machine learning: Assessing neurovascular signals in the prefrontal cortex with non-invasive bimodal electro-optical neuroimaging in opiate addiction. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Schall, T.A.; Wright, W.J.; Dong, Y. Nucleus accumbens fast-spiking interneurons in motivational and addictive behaviors. Mol. Psychiatry 2021, 26, 234–246. [Google Scholar] [CrossRef]
- Plumbly, W.; Brandon, N.; Deeb, T.Z.; Hall, J.; Harwood, A.J. L-type voltage-gated calcium channel regulation of in vitro human cortical neuronal networks. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Zych, S.M.; Ford, C.P. Divergent properties and independent regulation of striatal dopamine and GABA co-transmission. Cell Rep. 2022, 39, 110823. [Google Scholar] [CrossRef]
- Pfeiffer, M.; Koch, T.; Schröder, H.; Laugsch, M.; Höllt, V.; Schulz, S. Heterodimerization of somatostatin and opioid receptors cross-modulates phosphorylation, internalization, and desensitization. J. Biol. Chem. 2002, 277, 19762–19772. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, L.; Losi, G.; Lia, A.; Melone, M.; Chiavegato, A.; Gómez-Gonzalo, M.; Sessolo, M.; Bovetti, S.; Forli, A.; Zonta, M. Interneuron-specific signaling evokes distinctive somatostatin-mediated responses in adult cortical astrocytes. Nat. Commun. 2018, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Bienvenu, T.C.; Busti, D.; Micklem, B.R.; Mansouri, M.; Magill, P.J.; Ferraguti, F.; Capogna, M. Large intercalated neurons of amygdala relay noxious sensory information. J. Neurosci. 2015, 35, 2044–2057. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ari, Y.; Gaiarsa, J.-L.; Tyzio, R.; Khazipov, R. GABA: A pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiol. Rev. 2007, 87, 1215–1284. [Google Scholar] [CrossRef]
- Wang, D.D.; Kriegstein, A.R. Defining the role of GABA in cortical development. J. Physiol. 2009, 587, 1873–1879. [Google Scholar] [CrossRef]
- Wu, Y.K.; Miehl, C.; Gjorgjieva, J. Regulation of circuit organization and function through inhibitory synaptic plasticity. Trends Neurosci. 2022, 45, 884–898. [Google Scholar] [CrossRef]
- Xu, M.-Y.; Wong, A.H. GABAergic inhibitory neurons as therapeutic targets for cognitive impairment in schizophrenia. Acta Pharmacol. Sin. 2018, 39, 733–753. [Google Scholar] [CrossRef]
- Zorrilla de San Martin, J.; Donato, C.; Peixoto, J.; Aguirre, A.; Choudhary, V.; De Stasi, A.M.; Lourenço, J.; Potier, M.C.; Bacci, A. Alterations of specific cortical GABAergic circuits underlie abnormal network activity in a mouse model of Down syndrome. eLife 2020, 9, e58731. [Google Scholar] [CrossRef]
- Tabuchi, K.; Blundell, J.; Etherton, M.R.; Hammer, R.E.; Liu, X.; Powell, C.M.; Südhof, T.C. A neuroligin-3 mutation implicated in autism increases inhibitory synaptic transmission in mice. Science 2007, 318, 71–76. [Google Scholar] [CrossRef]
- Nomura, T. Interneuron Dysfunction and Inhibitory Deficits in Autism and Fragile X Syndrome. Cells 2021, 10, 2610. [Google Scholar] [CrossRef] [PubMed]
- Jackson, G.M.; Draper, A.; Dyke, K.; Pépés, S.E.; Jackson, S.R. Inhibition, disinhibition, and the control of action in Tourette syndrome. Trends Cogn. Sci. 2015, 19, 655–665. [Google Scholar] [CrossRef]
- Tian, Y.; Gunther, J.R.; Liao, I.H.; Liu, D.; Ander, B.P.; Stamova, B.S.; Lit, L.; Jickling, G.C.; Xu, H.; Zhan, X. GABA-and acetylcholine-related gene expression in blood correlate with tic severity and microarray evidence for alternative splicing in Tourette syndrome: A pilot study. Brain Res. 2011, 1381, 228–236. [Google Scholar] [CrossRef]
- Pérez-Cremades, D.; Hernández, S.; Blasco-Ibáñez, J.M.; Crespo, C.; Nacher, J.; Varea, E. Alteration of inhibitory circuits in the somatosensory cortex of Ts65Dn mice, a model for Down’s syndrome. J. Neural Transm. 2010, 117, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Cué, C.; Martínez, P.; Rueda, N.; Vidal, R.; García, S.; Vidal, V.; Corrales, A.; Montero, J.A.; Pazos, Á.; Flórez, J. Reducing GABAA α5 receptor-mediated inhibition rescues functional and neuromorphological deficits in a mouse model of down syndrome. J. Neurosci. 2013, 33, 3953–3966. [Google Scholar] [CrossRef]
- Han, S.; Tai, C.; Westenbroek, R.E.; Yu, F.H.; Cheah, C.S.; Potter, G.B.; Rubenstein, J.L.; Scheuer, T.; de La Iglesia, H.O.; Catterall, W.A. Autistic-like behaviour in Scn1a+/− mice and rescue by enhanced GABA-mediated neurotransmission. Nature 2012, 489, 385–390. [Google Scholar] [CrossRef]
- Olmos-Serrano, J.L.; Corbin, J.G.; Burns, M.P. The GABAA receptor agonist THIP ameliorates specific behavioral deficits in the mouse model of fragile X syndrome. Dev. Neurosci. 2011, 33, 395–403. [Google Scholar] [CrossRef]
- Fernandez, F.; Morishita, W.; Zuniga, E.; Nguyen, J.; Blank, M.; Malenka, R.C.; Garner, C.C. Pharmacotherapy for cognitive impairment in a mouse model of Down syndrome. Nat. Neurosci. 2007, 10, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Jazvinscak Jembrek, M.; Vlainic, J. GABA receptors: Pharmacological potential and pitfalls. Curr. Pharm. Des. 2015, 21, 4943–4959. [Google Scholar] [CrossRef]
- He, S.; Ma, J.; Liu, N.; Yu, X. Early enriched environment promotes neonatal GABAergic neurotransmission and accelerates synapse maturation. J. Neurosci. 2010, 30, 7910–7916. [Google Scholar] [CrossRef]
- Loisy, M.; Farah, A.; Fafouri, A.; Fanton, A.; Ahmadi, M.; Therreau, L.; Chevaleyre, V.; Piskorowski, R.A. Environmental enrichment and social isolation modulate inhibitory transmission and plasticity in hippocampal area CA2. Hippocampus 2023, 33, 197–207. [Google Scholar] [CrossRef]
- Sgroi, S.; Tonini, R. Opioidergic modulation of striatal circuits, implications in Parkinson’s disease and levodopa induced dyskinesia. Front. Neurol. 2018, 9, 524. [Google Scholar] [CrossRef]
- Ponterio, G.; Tassone, A.; Sciamanna, G.; Vanni, V.; Meringolo, M.; Santoro, M.; Mercuri, N.B.; Bonsi, P.; Pisani, A. Enhanced mu opioid receptor–dependent opioidergic modulation of striatal cholinergic transmission in DYT1 dystonia. Mov. Disord. 2018, 33, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.D.; Abi-Dargham, A. The role of dynorphin and the kappa opioid receptor in the symptomatology of schizophrenia: A review of the evidence. Biol. Psychiatry 2019, 86, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Pellissier, L.P.; Pujol, C.; Becker, J.; Le Merrer, J. Delta opioid receptors: Learning and motivation. Delta Opioid Recept. Pharmacol. Ther. Appl. 2018, 247, 227–260. [Google Scholar]
- Tanguturi, P.; Streicher, J.M. The role of opioid receptors in modulating Alzheimer’s Disease. Front. Pharmacol. 2023, 14, 1056402. [Google Scholar] [CrossRef] [PubMed]


| Protein/Cascade | GABAB Receptors | Opioidergic Receptors |
|---|---|---|
| The activity of adenylyl cyclase | Inhibition [227] | Inhibition [228] |
| Cyclic adenosine monophosphate levels | Reduction [227] | Reduction [228] |
| The activity of GIRK channels | Enhance [227] | Enhance [228] |
| The activity of voltage-gated Ca2+ channel | Inhibition [227] | Inhibition [228] |
| The activity of voltage-gated Na+ channel | Inhibition [229] | Inhibition [62] |
| The activity of phospholipase C | Enhance [230] | Enhance [55] |
| Secretion of brain-derived neurotrophic factor | Increase [230] | Increase [55] |
| P38/MAPK signaling | Enhance [230] | Enhance [55] |
| The activity of ERK | Enhance [231] | Enhance [232] |
| The activity of phosphorylated JNK | Enhance [233] | Enhance [234] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosseinzadeh Sahafi, O.; Sardari, M.; Alijanpour, S.; Rezayof, A. Shared Mechanisms of GABAergic and Opioidergic Transmission Regulate Corticolimbic Reward Systems and Cognitive Aspects of Motivational Behaviors. Brain Sci. 2023, 13, 815. https://doi.org/10.3390/brainsci13050815
Hosseinzadeh Sahafi O, Sardari M, Alijanpour S, Rezayof A. Shared Mechanisms of GABAergic and Opioidergic Transmission Regulate Corticolimbic Reward Systems and Cognitive Aspects of Motivational Behaviors. Brain Sciences. 2023; 13(5):815. https://doi.org/10.3390/brainsci13050815
Chicago/Turabian StyleHosseinzadeh Sahafi, Oveis, Maryam Sardari, Sakineh Alijanpour, and Ameneh Rezayof. 2023. "Shared Mechanisms of GABAergic and Opioidergic Transmission Regulate Corticolimbic Reward Systems and Cognitive Aspects of Motivational Behaviors" Brain Sciences 13, no. 5: 815. https://doi.org/10.3390/brainsci13050815
APA StyleHosseinzadeh Sahafi, O., Sardari, M., Alijanpour, S., & Rezayof, A. (2023). Shared Mechanisms of GABAergic and Opioidergic Transmission Regulate Corticolimbic Reward Systems and Cognitive Aspects of Motivational Behaviors. Brain Sciences, 13(5), 815. https://doi.org/10.3390/brainsci13050815