
Cognitive Avoidance Is Associated with Decreased Brain Responsiveness to Threat Distractors under High Perceptual Load

 ,
,
Abstract
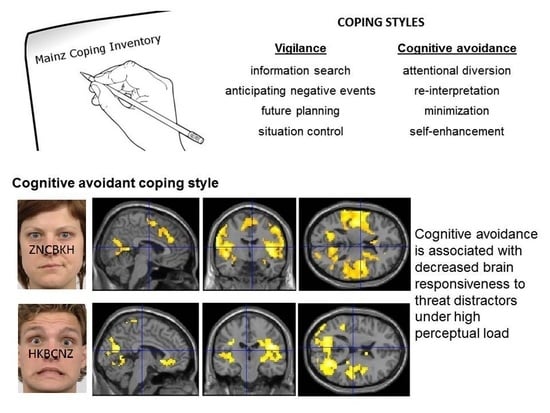
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Psychometric Measures
2.3. fMRI Experiment: Perceptual Load Task
2.4. MRI Acquisition and Preprocessing
2.5. Data Analyses
3. Results
3.1. Neural Main Effects
3.1.1. Main Effects of Threatening Faces under Low Perceptual Load
3.1.2. Main Effects of Threatening Faces under High Perceptual Load
3.2. Relationship among Brain Activation to Threat Faces and Coping Styles
3.2.1. Vigilance
3.2.2. Cognitive Avoidance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davis, M.; Walker, D.L.; Miles, L.; Grillon, C. Phasic vs sustained fear in rats and humans: Role of the extended amygdala in fear vs anxiety. Neuropsychopharmacology 2010, 35, 105–135. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.J.; Davis, M.; Öhman, A. Fear and anxiety: Animal models and human cognitive psychophysiology. J. Affect. Disord. 2000, 61, 137–159. [Google Scholar] [CrossRef]
- Sylvers, P.; Lilienfeld, S.O.; LaPrairie, J.L. Differences between trait fear and trait anxiety: Implications for psychopathology. Clin. Psychol. Rev. 2011, 31, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Suls, J. Toxic affect: Are anger, anxiety, and depression independent risk factors for cardiovascular disease? Emotion Rev. 2018, 10, 6–17. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 0-89042-555-8. [Google Scholar]
- Taylor, S.E.; Stanton, A.L. Coping resources, coping processes, and mental health. Annu. Rev. Clin. Psychol. 2007, 3, 377–401. [Google Scholar] [CrossRef] [PubMed]
- Mund, M.; Mitte, K. The costs of repression: A meta-analysis on the relation between repressive coping and somatic diseases. Health Psychol. 2012, 31, 640–649. [Google Scholar] [CrossRef]
- Myers, L.B. The importance of the repressive coping style: Findings from 30 years of research. Anxiety Stress Coping 2010, 23, 3–17. [Google Scholar] [CrossRef]
- Collins, S.; Woolfson, L.M.; Durkin, K. Effects on coping skills and anxiety of a universal school-based mental health intervention delivered in Scottish primary schools. Sch. Psychol. Int. 2014, 35, 85–100. [Google Scholar] [CrossRef]
- Krohne, H.W. Vigilance and cognitive avoidance as concepts in coping research. In Attention and Avoidance; Krohne, H.W., Ed.; Hogrefe & Huber: Seattle, WA, USA, 1993; pp. 19–50. [Google Scholar]
- Krohne, H.W.; Egloff, B.; Varner, L.J.; Burns, L.R.; Weidner, G.; Ellis, H.C. The assessment of dispositional vigilance and cognitive avoidance: Factorial structure, psychometric properties, and validity of the Mainz Coping Inventory. Cogn. Ther. Res. 2000, 24, 297–311. [Google Scholar] [CrossRef]
- Krohne, H.W.; Schmukle, S.C.; Burns, L.R.; Egloff, B.; Spielberger, C.D. The measurement of coping in achievement situations: An international comparison. Pers. Individ. Dif. 2001, 30, 1225–1243. [Google Scholar] [CrossRef]
- Weinberger, D.A.; Schwartz, G.E.; Davidson, R.J. Low-anxious, high-anxious, and repressive coping styles: Psychometric patterns and behavioral and physiological responses to stress. J. Abnorm. Psychol. 1979, 88, 369–380. [Google Scholar] [CrossRef]
- Egloff, B.; Hock, M. A comparison of two approaches to the assessment of coping styles. Pers. Individ. Dif. 1997, 23, 913–916. [Google Scholar] [CrossRef]
- Kohlmann, C.-W. Rigid and flexible modes of coping: Related to coping style? Anxiety Stress Coping 1993, 6, 107–123. [Google Scholar] [CrossRef]
- Gebhardt, C.; Rose, N.; Mitte, K. Fact or artefact: An item response theory analysis of median split based repressor classification. Br. J. Health Psychol. 2014, 19, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Rauch, A.V.; Ohrmann, P.; Bauer, J.; Kugel, H.; Engelien, A.; Arolt, V.; Heindel, W.; Suslow, T. Cognitive coping style modulates neural responses to emotional faces in healthy humans: A 3-T fMRI study. Cereb. Cortex 2007, 17, 2526–2535. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.G.; Rauch, A.V.; Kugel, H.; Ter Horst, L.; Bauer, J.; Dannlowski, U.; Ohrmann, P.; Lindner, C.; Donges, U.-S.; Kersting, A.; et al. High responsivity to threat during the initial stage of perception in repression: A 3 T fMRI study. Soc. Cogn. Affect. Neurosci. 2012, 7, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Rauch, A.V.; Ter Horst, L.; Paul, V.G.; Bauer, J.; Dannlowski, U.; Konrad, C.; Ohrmann, P.; Kugel, H.; Egloff, B.; Arolt, V.; et al. Influence of repressive coping style on cortical activation during encoding of angry faces. PLoS ONE 2014, 9, e112398. [Google Scholar] [CrossRef]
- Klucken, T.; Kruse, O.; Schweckendiek, J.; Stark, R. Increased skin conductance responses and neural activity during fear conditioning are associated with a repressive coping style. Front. Behav. Neurosci. 2015, 9, 132. [Google Scholar] [CrossRef]
- Stevens, F.L.; Hurley, R.A.; Taber, K.H. Anterior cingulate cortex: Unique role in cognition and emotion. J. Neuropsychiatry Clin. Neurosci. 2011, 23, 121–125. [Google Scholar] [CrossRef]
- Goldin, P.R.; McRae, K.; Ramel, W.; Gross, J.J. The neural bases of emotion regulation: Reappraisal and suppression of negative emotion. Biol. Psychiatry 2008, 63, 577–586. [Google Scholar] [CrossRef]
- Egloff, B.; Krohne, H.W. The measurement of vigilance and cognitive avoidance: Investigations with the “Angstbewältigungsinventar” (ABI). Diagnostica 1998, 44, 189–200. [Google Scholar]
- Günther, V.; Jahn, S.; Webelhorst, C.; Bodenschatz, C.M.; Bujanow, A.; Mucha, S.; Kersting, A.; Hoffmann, K.-T.; Egloff, B.; Lobsien, D.; et al. Coping with anxiety: Brain structural correlates of vigilance and cognitive avoidance. Front. Psychiatry 2022, 13, 869367. [Google Scholar] [CrossRef]
- Leehr, E.J.; Redlich, R.; Zaremba, D.; Dohm, K.; Böhnlein, J.; Grotegerd, D.; Kähler, C.; Repple, J.; Förster, K.; Opel, N.; et al. Structural and functional neural correlates of vigilant and avoidant regulation style. J. Affect. Disord. 2019, 258, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Liddell, B.J.; Brown, K.J.; Kemp, A.H.; Barton, M.J.; Das, P.; Peduto, A.; Gordon, E.; Williams, L.M. A direct brainstem-amygdala-cortical ‘alarm’ system for subliminal signals of fear. NeuroImage 2005, 24, 235–243. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, J.E. The Emotional Brain: The Mysterious Underspinnings of Emotional Life; Simon & Schuster Paperbacks: New York, NY, USA, 1996; ISBN 9780684836591. [Google Scholar]
- Vuilleumier, P.; Armony, J.L.; Driver, J.; Dolan, R.J. Distinct spatial frequency sensitivities for processing faces and emotional expressions. Nat. Neurosci. 2003, 6, 624–631. [Google Scholar] [CrossRef]
- Herrmann, M.J.; Ehlis, A.-C.; Muehlberger, A.; Fallgatter, A.J. Source localization of early stages of face processing. Brain Topogr. 2005, 18, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Adolphs, R. Neural systems for recognizing emotion. Curr. Opin. Neurobiol. 2002, 12, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, S.; Cardellicchio, P.; Di Fazio, C.; Nazzi, C.; Fracasso, A.; Borgomaneri, S. Stopping in (e)motion: Reactive action inhibition when facing valence-independent emotional stimuli. Front. Behav. Neurosci. 2022, 16, 998714. [Google Scholar] [CrossRef] [PubMed]
- Kirby, L.A.J.; Robinson, J.L. Affective mapping: An activation likelihood estimation (ALE) meta-analysis. Brain Cogn. 2017, 118, 137–148. [Google Scholar] [CrossRef]
- Sawada, R.; Sato, W. Emotional attention capture by facial expressions. Sci. Rep. 2015, 5, 14042. [Google Scholar] [CrossRef]
- Dolan, R.J.; Vuilleumier, P. Amygdala automaticity in emotional processing. Ann. NY Acad. Sci. 2003, 985, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.T.; Zajonc, R.B. Affect, cognition, and awareness: Affective priming with optimal and suboptimal stimulus exposures. J. Pers. Soc. Psychol. 1993, 64, 723–739. [Google Scholar] [CrossRef] [PubMed]
- Okon-Singer, H.; Tzelgov, J.; Henik, A. Distinguishing between automaticity and attention in the processing of emotionally significant stimuli. Emotion 2007, 7, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.J.; Jenkins, R.; Lawrence, A.D. Neural processing of fearful faces: Effects of anxiety are gated by perceptual capacity limitations. Cereb. Cortex 2007, 17, 1595–1603. [Google Scholar] [CrossRef]
- Lavie, N. Perceptual load as a necessary condition for selective attention. J. Exp. Psychol. Hum. Percept. Perform. 1995, 21, 451–468. [Google Scholar] [CrossRef]
- Lavie, N. Attention, distraction, and cognitive control under load. Curr. Dir. Psychol. Sci. 2010, 19, 143–148. [Google Scholar] [CrossRef]
- Brockhoff, L.; Schindler, S.; Bruchmann, M.; Straube, T. Effects of perceptual and working memory load on brain responses to task-irrelevant stimuli: Review and implications for future research. Neurosci. Biobehav. Rev. 2022, 135, 104580. [Google Scholar] [CrossRef]
- Wheaton, M.G.; Fitzgerald, D.A.; Phan, K.L.; Klumpp, H. Perceptual load modulates anterior cingulate cortex response to threat distractors in generalized social anxiety disorder. Biol. Psychol. 2014, 101, 13–17. [Google Scholar] [CrossRef]
- Wieser, M.J.; Keil, A. Fearful faces heighten the cortical representation of contextual threat. NeuroImage 2014, 86, 317–325. [Google Scholar] [CrossRef]
- Izard, C.E. The Psychology of Emotions; Plenum Press: New York, NY, USA, 1991. [Google Scholar]
- Aleman, A.; Swart, M. Sex differences in neural activation to facial expressions denoting contempt and disgust. PLoS ONE 2008, 3, e3622. [Google Scholar] [CrossRef]
- Wittchen, H.; Wunderlich, U.; Gruschwitz, S.; Zaudig, M. SKID-I. Strukturiertes Klinisches Interview für DSM-IV; Hogrefe: Göttingen, Germany, 1997. [Google Scholar]
- Laux, L.; Glanzmann, P.; Schaffner, P.; Spielberger, C.D. State-Trait-Angstinventar (STAI). Manual; Beltz: Weinheim, Germany, 1981. [Google Scholar]
- Langner, O.; Dotsch, R.; Bijlstra, G.; Wigboldus, D.H.J.; Hawk, S.T.; van Knippenberg, A. Presentation and validation of the Radboud Faces Database. Cogn. Emot. 2010, 24, 1377–1388. [Google Scholar] [CrossRef]
- Edwards, B.G.; Calhoun, V.D.; Kiehl, K.A. Joint ICA of ERP and fMRI during error-monitoring. NeuroImage 2012, 59, 1896–1903. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mugler, J.P.; Brookeman, J.R. Three-dimensional magnetization-prepared rapid gradient-echo imaging (3D MP RAGE). Magn. Reason. Med. 1990, 15, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Maldjian, J.A.; Laurienti, P.J.; Kraft, R.A.; Burdette, J.H. An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. NeuroImage 2003, 19, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Moors, A. Automaticity: Componential, causal, and mechanistic explanations. Annu. Rev. Psychol. 2016, 67, 263–287. [Google Scholar] [CrossRef]
- Okon-Singer, H.; Mehnert, J.; Hoyer, J.; Hellrung, L.; Schaare, H.L.; Dukart, J.; Villringer, A. Neural control of vascular reactions: Impact of emotion and attention. J. Neurosci. 2014, 34, 4251–4259. [Google Scholar] [CrossRef]
- Tamietto, M.; de Gelder, B. Neural bases of the non-conscious perception of emotional signals. Nat. Rev. Neurosci. 2010, 11, 697–709. [Google Scholar] [CrossRef]
- Saalmann, Y.B.; Kastner, S. Cognitive and perceptual functions of the visual thalamus. Neuron 2011, 71, 209–223. [Google Scholar] [CrossRef]
- Saalmann, Y.B. Intralaminar and medial thalamic influence on cortical synchrony, information transmission and cognition. Front. Syst. Neurosci. 2014, 8, 83. [Google Scholar] [CrossRef]
- Abivardi, A.; Bach, D.R. Deconstructing white matter connectivity of human amygdala nuclei with thalamus and cortex subdivisions in vivo. Hum. Brain Mapp. 2017, 38, 3927–3940. [Google Scholar] [CrossRef]
- Davis, M.; Whalen, P.J. The amygdala: Vigilance and emotion. Mol. Psychiatry 2001, 6, 13–34. [Google Scholar] [CrossRef]
- Phelps, E.A.; LeDoux, J.E. Contributions of the amygdala to emotion processing: From animal models to human behavior. Neuron 2005, 48, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Pessoa, L.; Adolphs, R. Emotion processing and the amygdala: From a ‘low road’ to ‘many roads’ of evaluating biological significance. Nat. Rev. Neurosci. 2010, 11, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R. Cuneus. In Encyclopedia of Clinical Neuropsychology; Kreutzer, J., DeLuca, J., Caplan, B., Eds.; Springer International Publishing: Cham, Switzerland, 2011; ISBN 978-3-319-56782-2. [Google Scholar]
- Engell, A.D.; Haxby, J.V. Facial expression and gaze-direction in human superior temporal sulcus. Neuropsychologia 2007, 45, 3234–3241. [Google Scholar] [CrossRef]
- Nomi, J.S.; Scherfeld, D.; Friederichs, S.; Schäfer, R.; Franz, M.; Wittsack, H.-J.; Azari, N.P.; Missimer, J.; Seitz, R.J. On the neural networks of empathy: A principal component analysis of an fMRI study. Behav. Brain Funct. 2008, 4, 41. [Google Scholar] [CrossRef]
- Ochsner, K.N.; Silvers, J.A.; Buhle, J.T. Functional imaging studies of emotion regulation: A synthetic review and evolving model of the cognitive control of emotion. Ann. N. Y. Acad. Sci. 2012, 1251, E1–E24. [Google Scholar] [CrossRef]
- Vanderhasselt, M.-A.; Kühn, S.; de Raedt, R. ‘Put on your poker face’: Neural systems supporting the anticipation for expressive suppression and cognitive reappraisal. Soc. Cogn. Affect. Neurosci. 2013, 8, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Cavanna, A.E.; Trimble, M.R. The precuneus: A review of its functional anatomy and behavioural correlates. Brain 2006, 129, 564–583. [Google Scholar] [CrossRef] [PubMed]
- Numssen, O.; Bzdok, D.; Hartwigsen, G. Functional specialization within the inferior parietal lobes across cognitive domains. Elife 2021, 10, e63591. [Google Scholar] [CrossRef]
- Shomstein, S. Cognitive functions of the posterior parietal cortex: Top-down and bottom-up attentional control. Front. Integr. Neurosci. 2012, 6, 38. [Google Scholar] [CrossRef]
- Fan, J.; Gu, X.; Liu, X.; Guise, K.G.; Park, Y.; Martin, L.; Marchena, A.; de Tang, C.Y.; Minzenberg, M.J.; Hof, P.R. Involvement of the anterior cingulate and frontoinsular cortices in rapid processing of salient facial emotional information. NeuroImage 2011, 54, 2539–2546. [Google Scholar] [CrossRef] [PubMed]
- Adolphs, R.; Spezio, M. Role of the amygdala in processing visual social stimuli. Prog. Brain Res. 2006, 156, 363–378. [Google Scholar] [PubMed]
- Said, C.P.; Haxby, J.V.; Todorov, A. Brain systems for assessing the affective value of faces. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 1660–1670. [Google Scholar] [CrossRef]
- Leech, R.; Sharp, D.J. The role of the posterior cingulate cortex in cognition and disease. Brain 2014, 137, 12–32. [Google Scholar] [CrossRef] [PubMed]
- Maddock, R.J.; Garrett, A.S.; Buonocore, M.H. Posterior cingulate cortex activation by emotional words: FMRI evidence from a valence decision task. Hum. Brain Mapp. 2003, 18, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Brewer, J.A.; Garrison, K.A.; Whitfield-Gabrieli, S. What about the "self" is processed in the posterior cingulate cortex? Front. Hum. Neurosci. 2013, 7, 647. [Google Scholar] [CrossRef]
- Cauda, F.; Costa, T.; Torta, D.M.E.; Sacco, K.; D’Agata, F.; Duca, S.; Geminiani, G.; Fox, P.T.; Vercelli, A. Meta-analytic clustering of the insular cortex: Characterizing the meta-analytic connectivity of the insula when involved in active tasks. NeuroImage 2012, 62, 343–355. [Google Scholar] [CrossRef]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Craig, A.D.B. How do you feel—Now? The anterior insula and human awareness. Nat. Rev. Neurosci. 2009, 10, 59–70. [Google Scholar] [CrossRef]
- Uddin, L.Q.; Nomi, J.S.; Hébert-Seropian, B.; Ghaziri, J.; Boucher, O. Structure and function of the human insula. J. Clin. Neurophysiol. 2017, 34, 300–306. [Google Scholar] [CrossRef]
- Gehrlach, D.A.; Dolensek, N.; Klein, A.S.; Roy Chowdhury, R.; Matthys, A.; Junghänel, M.; Gaitanos, T.N.; Podgornik, A.; Black, T.D.; Reddy Vaka, N.; et al. Aversive state processing in the posterior insular cortex. Nat. Neurosci. 2019, 22, 1424–1437. [Google Scholar] [CrossRef] [PubMed]
- Kurth, F.; Zilles, K.; Fox, P.T.; Laird, A.R.; Eickhoff, S.B. A link between the systems: Functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct. Funct. 2010, 214, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Straube, T.; Miltner, W.H.R. Attention to aversive emotion and specific activation of the right insula and right somatosensory cortex. NeuroImage 2011, 54, 2534–2538. [Google Scholar] [CrossRef]
- Terasawa, Y.; Fukushima, H.; Umeda, S. How does interoceptive awareness interact with the subjective experience of emotion? An fMRI study. Hum. Brain Mapp. 2013, 34, 598–612. [Google Scholar] [CrossRef]
- Phillips, M.L.; Ladouceur, C.D.; Drevets, W.C. A neural model of voluntary and automatic emotion regulation: Implications for understanding the pathophysiology and neurodevelopment of bipolar disorder. Mol. Psychiatry 2008, 13, 829, 833–857. [Google Scholar] [CrossRef]
- Heilbronner, S.R.; Hayden, B.Y. Dorsal anterior cingulate cortex: A bottom-up view. Annu. Rev. Neurosci. 2016, 39, 149–170. [Google Scholar] [CrossRef] [PubMed]
- Etkin, A.; Egner, T.; Kalisch, R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cogn. Sci. 2011, 15, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Gerber, A.J.; Posner, J.; Gorman, D.; Colibazzi, T.; Yu, S.; Wang, Z.; Kangarlu, A.; Zhu, H.; Russell, J.; Peterson, B.S. An affective circumplex model of neural systems subserving valence, arousal, and cognitive overlay during the appraisal of emotional faces. Neuropsychologia 2008, 46, 2129–2139. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, R.P.; Kirlic, N.; Misaki, M.; Bodurka, J.; Rhudy, J.L.; Paulus, M.P.; Drevets, W.C. Increased anterior insula activity in anxious individuals is linked to diminished perceived control. Transl. Psychiatry 2015, 5, e591. [Google Scholar] [CrossRef]
- Brooks, S.J.; Savov, V.; Allzén, E.; Benedict, C.; Fredriksson, R.; Schiöth, H.B. Exposure to subliminal arousing stimuli induces robust activation in the amygdala, hippocampus, anterior cingulate, insular cortex and primary visual cortex: A systematic meta-analysis of fMRI studies. NeuroImage 2012, 59, 2962–2973. [Google Scholar] [CrossRef]
- Critchley, H.D.; Rotshtein, P.; Nagai, Y.; O’Doherty, J.; Mathias, C.J.; Dolan, R.J. Activity in the human brain predicting differential heart rate responses to emotional facial expressions. NeuroImage 2005, 24, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Gianaros, P.J.; Jennings, J.R.; Sheu, L.K.; Derbyshire, S.W.G.; Matthews, K.A. Heightened functional neural activation to psychological stress covaries with exaggerated blood pressure reactivity. Hypertension 2007, 49, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Burklund, L.J.; Eisenberger, N.I.; Lieberman, M.D. The face of rejection: Rejection sensitivity moderates dorsal anterior cingulate activity to disapproving facial expressions. Soc. Neuro. Sci. 2007, 2, 238–253. [Google Scholar] [CrossRef] [PubMed]
- Eisenberger, N.I.; Lieberman, M.D.; Williams, K.D. Does rejection hurt? An FMRI study of social exclusion. Science 2003, 302, 290–292. [Google Scholar] [CrossRef]
- Schwerdtfeger, A.R.; Rathner, E.-M. The ecological validity of the autonomic-subjective response dissociation in repressive coping. Anxiety Stress Coping 2016, 29, 241–258. [Google Scholar] [CrossRef]
- Grant, D.M.; Wingate, L.R.; Rasmussen, K.A.; Davidson, C.L.; Slish, M.L.; Rhoades-Kerswill, S.; Mills, A.C.; Judah, M.R. An examination of the reciprocal relationship between avoidance coping and symptoms of anxiety and depression. J. Soc. Clin. Psychol. 2013, 32, 878–896. [Google Scholar] [CrossRef]
- Coifman, K.G.; Bonanno, G.A.; Ray, R.D.; Gross, J.J. Does repressive coping promote resilience? Affective-autonomic response discrepancy during bereavement. J. Pers. Soc. Psychol. 2007, 92, 745–758. [Google Scholar] [CrossRef]
- Bonanno, G.A.; Keltner, D.; Holen, A.; Horowitz, M.J. When avoiding unpleasant emotions might not be such a bad thing: Verbal-autonomic response dissociation and midlife conjugal bereavement. J. Pers. Soc. Psychol. 1995, 69, 975–989. [Google Scholar] [CrossRef]
- Krohne, H.W. The concept of coping modes: Relating cognitive person variables to actual coping behavior. Adv. Behav. Res. Ther. 1989, 11, 235–248. [Google Scholar] [CrossRef]
- Kohlmann, C.-W.; Weidner, G.; Messina, C.R. Avoidant coping style and verbal-cardiovascular response dissociation. Psychol. Health 1996, 11, 371–384. [Google Scholar] [CrossRef]
- Howard, S.; Myers, L.B.; Hughes, B.M. Repressive coping and cardiovascular reactivity to novel and recurrent stress. Anxiety Stress Coping 2017, 30, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Brosschot, J.F.; Janssen, E. Continuous monitoring of affective-autonomic response dissociation in repressers during negative emotional stimulation. Pers. Individ. Dif. 1998, 25, 69–84. [Google Scholar] [CrossRef]
- Jørgensen, M.M.; Zachariae, R. Repressive coping style and autonomic reactions to two experimental stressors in healthy men and women. Scand. J. Psychol. 2006, 47, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Moos, R.H.; Holahan, C.J. Dispositional and contextual perspectives on coping: Toward an integrative framework. J. Clin. Psychol. 2003, 59, 1387–1403. [Google Scholar] [CrossRef] [PubMed]
- Rosser, B.A. Intolerance of uncertainty as a transdiagnostic mechanism of psychological difficulties: A systematic review of evidence pertaining to causality and temporal precedence. Cogn. Ther. Res. 2019, 43, 438–463. [Google Scholar] [CrossRef]
- Brühl, A.B.; Delsignore, A.; Komossa, K.; Weidt, S. Neuroimaging in social anxiety disorder—A meta-analytic review resulting in a new neurofunctional model. Neurosci. Biobehav. Rev. 2014, 47, 260–280. [Google Scholar] [CrossRef]
- Etkin, A.; Klemenhagen, K.C.; Dudman, J.T.; Rogan, M.T.; Hen, R.; Kandel, E.R.; Hirsch, J. Individual differences in trait anxiety predict the response of the basolateral amygdala to unconsciously processed fearful faces. Neuron 2004, 44, 1043–1055. [Google Scholar] [CrossRef]
- Fu, X.; Taber-Thomas, B.C.; Pérez-Edgar, K. Frontolimbic functioning during threat-related attention: Relations to early behavioral inhibition and anxiety in children. Biol. Psychol. 2017, 122, 98–109. [Google Scholar] [CrossRef]
- Heitmann, C.Y.; Feldker, K.; Neumeister, P.; Brinkmann, L.; Schrammen, E.; Zwitserlood, P.; Straube, T. Brain activation to task-irrelevant disorder-related threat in social anxiety disorder: The impact of symptom severity. Neuroimage Clin. 2017, 14, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Blair, K.S.; Geraci, M.; Korelitz, K.; Otero, M.; Towbin, K.; Ernst, M.; Leibenluft, E.; Blair, R.J.R.; Pine, D.S. The pathology of social phobia is independent of developmental changes in face processing. Am. J. Psychiatry 2011, 168, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Feldker, K.; Heitmann, C.Y.; Neumeister, P.; Brinkmann, L.; Bruchmann, M.; Zwitserlood, P.; Straube, T. Cardiorespiratory concerns shape brain responses during automatic panic-related scene processing in patients with panic disorder. J. Psychiatry Neurosci. 2018, 43, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Lipka, J.; Hoffmann, M.; Miltner, W.H.R.; Straube, T. Effects of cognitive-behavioral therapy on brain responses to subliminal and supraliminal threat and their functional significance in specific phobia. Biol. Psychiatry 2014, 76, 869–877. [Google Scholar] [CrossRef]
- Wright, C.I.; Fischer, H.; Whalen, P.J.; McInerney, S.C.; Shin, L.M.; Rauch, S.L. Differential prefrontal cortex and amygdala habituation to repeatedly presented emotional stimuli. Neuroreport 2001, 12, 379–383. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
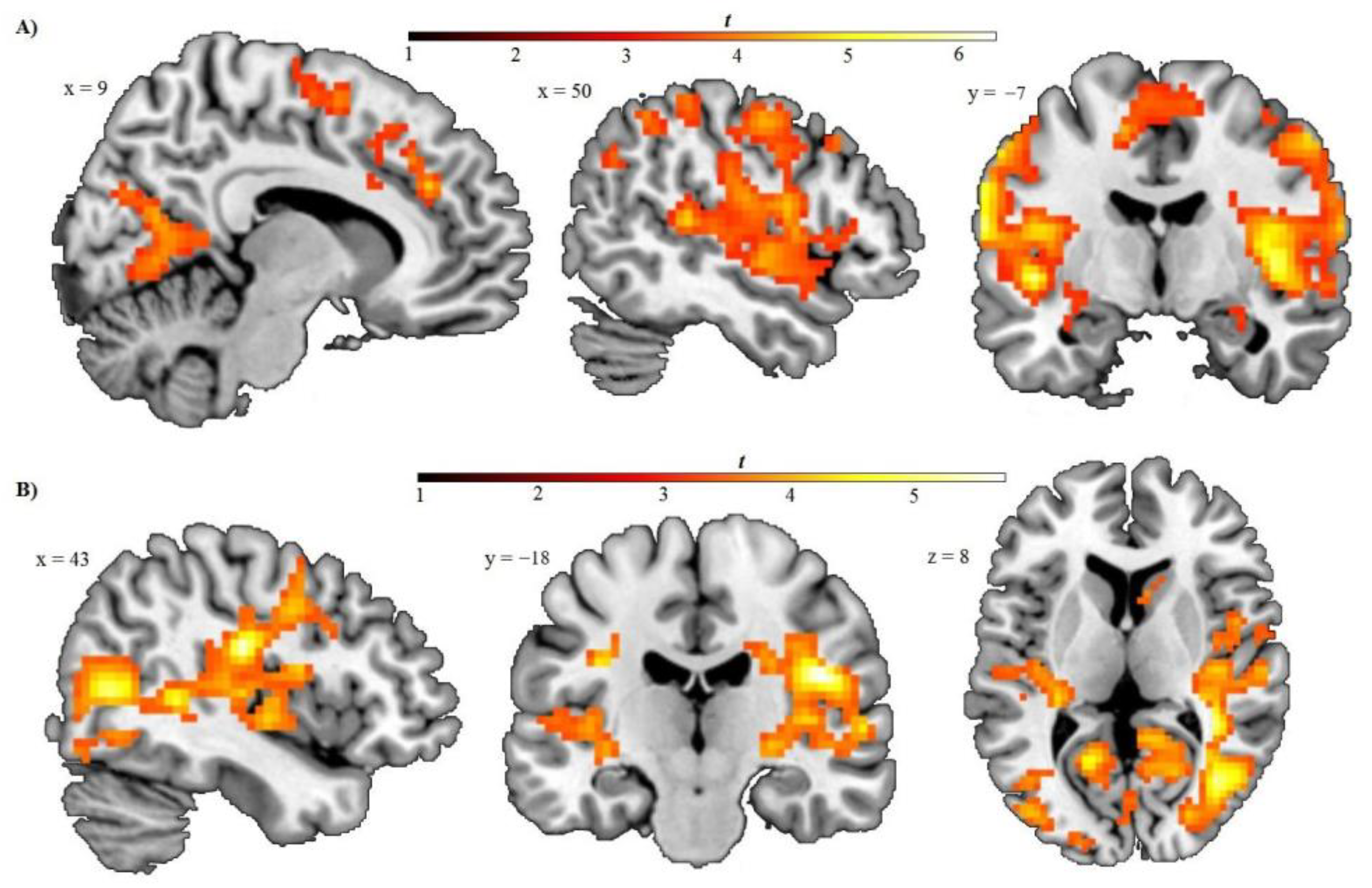{kind=link}
| Coping | Trait Anxiety | ||
|---|---|---|---|
| M (SD) | MCI-CAV | STAI | |
| MCI-VIG | 22.00 (6.80) | −0.41 ** | 0.35 * |
| MCI-CAV | 23.75 (5.11) | - | −0.36 * |
| STAI-trait | 35.05 (6.82) | - | - |
| Hemisphere | Peak T-Value | Peak-Level | Cluster | Cluster-Level | Peak MNI | |||
|---|---|---|---|---|---|---|---|---|
| puncorrected | Size (Voxels) | pFWE | x | y | z | |||
| Low perceptual load | - | - | - | - | - | - | - | - |
| Contempt > neutral | - | - | - | - | - | - | - | - |
| Superior extending to inferior parietal lobule | R | 5.61 | <0.001 | 183 | 0.004 | 42 | −55 | 55 |
| Middle temporal extending to fusiform gyrus | R | 5.23 | <0.001 | 536 | <0.001 | 57 | −46 | −5 |
| Inferior frontal gyrus | R | 5.33 | <0.001 | 278 | 0.003 | 45 | 26 | 22 |
| Inferior parietal lobule (BA40) | L | 5.10 | <0.001 | 183 | 0.004 | −42 | −52 | 55 |
| Inferior temporal gyrus | L | 4.57 | <0.001 | 171 | 0.005 | −42 | −49 | −17 |
| Thalamus (ROI) | L | 3.69 | <0.001 | 1 | 0.04 | 9 | −28 | 1 |
| Fear > neutral | - | - | - | - | - | - | - | - |
| - | - | - | - | - | - | - | - | - |
| High perceptual load | - | - | - | - | - | - | - | - |
| Contempt > neutral | - | - | - | - | - | - | - | |
| - | - | - | - | - | - | - | - | - |
| Fear > neutral | - | - | - | - | - | - | - | |
| Inferior frontal extending to precentral gyrus | L | 4.73 | <0.001 | 230 | 0.003 | −51 | 17 | 19 |
| Superior temporal gyrus | R | 4.89 | <0.001 | 267 | 0.001 | −57 | −43 | 13 |
| Superior temporal gyrus | L | 5.33 | <0.001 | 217 | 0.004 | −57 | −46 | 13 |
| Cerebellum | R | 4.62 | <0.001 | 419 | <0.001 | 42 | −67 | −29 |
| Hemisphere | Peak T-Value | Peak-Level | Cluster | Cluster-Level | Peak MNI | |||
|---|---|---|---|---|---|---|---|---|
| puncorrected | Size (Voxels) | pFWE | x | y | z | |||
| Contempt (vs. neutral) | - | - | - | - | - | - | - | - |
| Left postcentral gyrus, extending to the following regions: | L/R | 6.29 | <0.001 | 5770 | <0.001 | −63 | −10 | 16 |
| Bilateral precentral (BA6), superior, and middle frontal gyrus (BA8) | L/R | 5.18 | <0.001 | - | - | −51 | −1 | 16 |
| Bilateral posterior and anterior insula | L/R | 6.14 | <0.001 | - | - | 42 | −4 | 4 |
| Bilateral lingual and parahippocampal gyrus, calcarine sulcus and precuneus | L/R | 5.16 | <0.001 | - | - | −18 | −55 | 1 |
| Bilateral superior temporal gyrus | L/R | 5.38 | <0.001 | - | - | −48 | −37 | 19 |
| Bilateral supramarginal and inferior parietal gyrus | L/R | 4.67 | <0.001 | - | - | −63 | −19 | 37 |
| Amygdala | L/R | 3.46 | <0.001 | - | - | −27 | −7 | −11 |
| Thalamus | L | 3.79 | <0.001 | - | - | −6 | −10 | 16 |
| Dorsal anterior and middle cingulate gyrus (BA32), extending to medial frontal cortex (BA6,8,9) | R | 4.59 | <0.001 | 214 | 0.002 | 9 | 35 | 28 |
| Bilateral supplementary motor area (BA6), extending to the left medial frontal and middle cingulate gyrus (BA31) | L/R | 4.51 | <0.001 | 221 | 0.001 | −12 | −7 | 52 |
| Right postcentral gyrus extending to inferior parietal gyrus | R | 4.13 | <0.001 | 158 | 0.008 | 33 | −40 | 58 |
| Fear (vs. neutral) | ||||||||
| Subcentral gyrus, extending to the following regions: | L/R | 5.67 | <0.001 | 3634 | <0.001 | 45 | −19 | 22 |
| Bilateral superior temporal gyrus | L/R | 4.69 | <0.001 | - | - | 45 | −10 | −2 |
| Posterior insula | R | 4.81 | <0.001 | - | - | 45 | −10 | 1 |
| Precentral gyrus | R | 4.68 | <0.001 | - | - | |||
| Bilateral posterior cingulate | L/R | 3.77 | <0.001 | - | - | 12 | −40 | 28 |
| Bilateral lingual gyrus, cuneus and precuneus | L/R | 5.05 | <0.001 | - | - | 15 | −43 | −5 |
| Amygdala | R | 3.49 | <0.001 | - | - | 27 | −4 | −11 |
| Thalamus | L/R | 4.25 | <0.001 | - | - | −3 | −13 | −2 |
| Bilateral anterior cingulate gyrus (BA25, 32) | L/R | 4.75 | <0.001 | 353 | 0.001 | −3 | 17 | −5 |
| Fusiform gyrus | L | 4.22 | <0.001 | 307 | <0.001 | −36 | −46 | −23 |
| Postcentral gyrus | R | 4.49 | <0.001 | 161 | 0.008 | 12 | −43 | 73 |
| Postcentral extending to superior temporal gyrus | L | 4.41 | <0.001 | 110 | 0.03 | −36 | −19 | 28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Günther, V.; Strukova, M.; Pecher, J.; Webelhorst, C.; Engelmann, S.; Kersting, A.; Hoffmann, K.-T.; Egloff, B.; Okon-Singer, H.; Lobsien, D.; et al. Cognitive Avoidance Is Associated with Decreased Brain Responsiveness to Threat Distractors under High Perceptual Load. Brain Sci. 2023, 13, 618. https://doi.org/10.3390/brainsci13040618
Günther V, Strukova M, Pecher J, Webelhorst C, Engelmann S, Kersting A, Hoffmann K-T, Egloff B, Okon-Singer H, Lobsien D, et al. Cognitive Avoidance Is Associated with Decreased Brain Responsiveness to Threat Distractors under High Perceptual Load. Brain Sciences. 2023; 13(4):618. https://doi.org/10.3390/brainsci13040618
Chicago/Turabian StyleGünther, Vivien, Mariia Strukova, Jonas Pecher, Carolin Webelhorst, Simone Engelmann, Anette Kersting, Karl-Titus Hoffmann, Boris Egloff, Hadas Okon-Singer, Donald Lobsien, and et al. 2023. "Cognitive Avoidance Is Associated with Decreased Brain Responsiveness to Threat Distractors under High Perceptual Load" Brain Sciences 13, no. 4: 618. https://doi.org/10.3390/brainsci13040618
APA StyleGünther, V., Strukova, M., Pecher, J., Webelhorst, C., Engelmann, S., Kersting, A., Hoffmann, K.-T., Egloff, B., Okon-Singer, H., Lobsien, D., & Suslow, T. (2023). Cognitive Avoidance Is Associated with Decreased Brain Responsiveness to Threat Distractors under High Perceptual Load. Brain Sciences, 13(4), 618. https://doi.org/10.3390/brainsci13040618