
Molecular Targeted Therapies in Glioblastoma Multiforme: A Systematic Overview of Global Trends and Findings

 ,
,  ,
, 
Abstract
1. Introduction
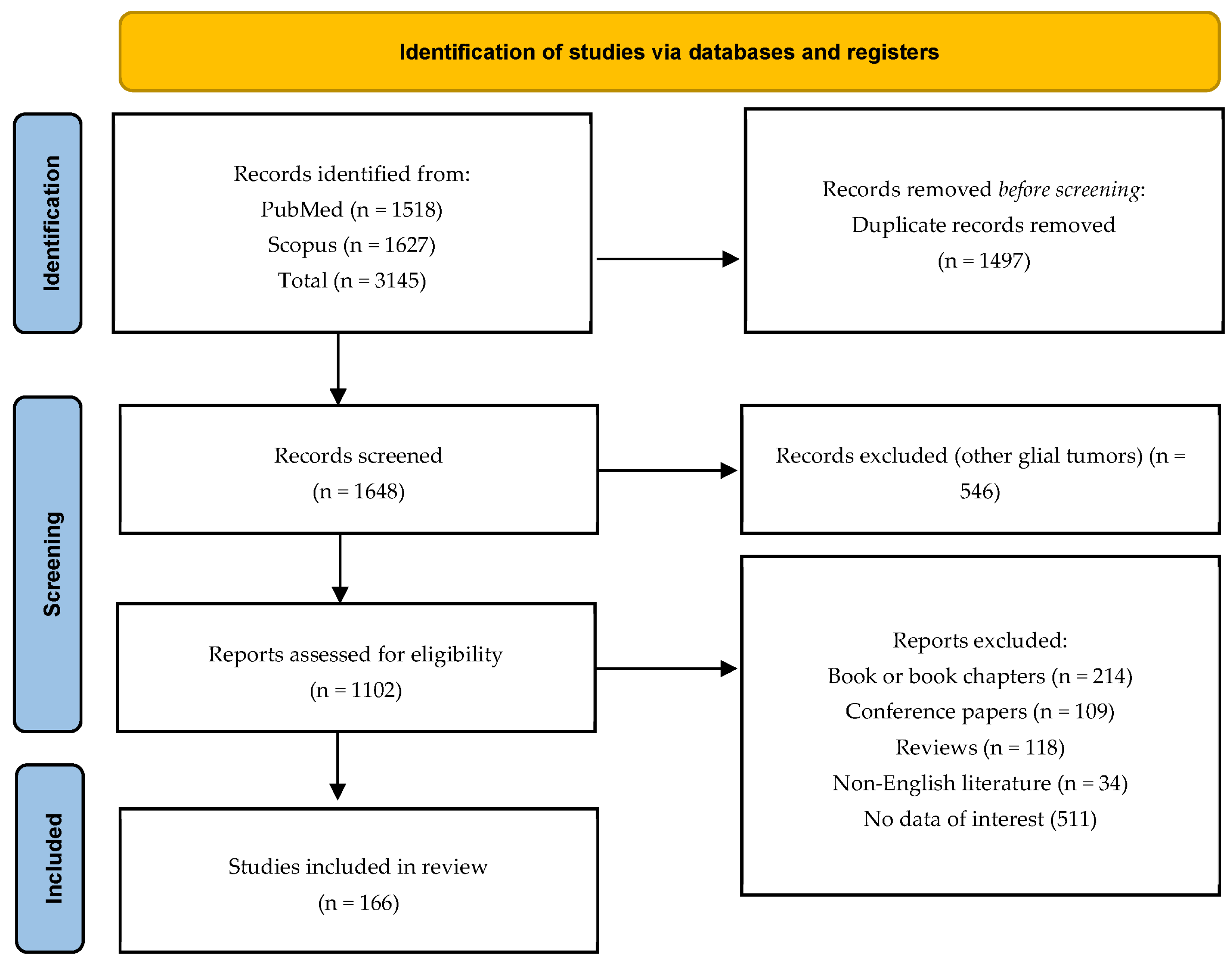
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion and Exclusion Criteria
2.3. Data Extraction and Processing
2.4. Statistical Analysis and Graphical Elements
3. Results
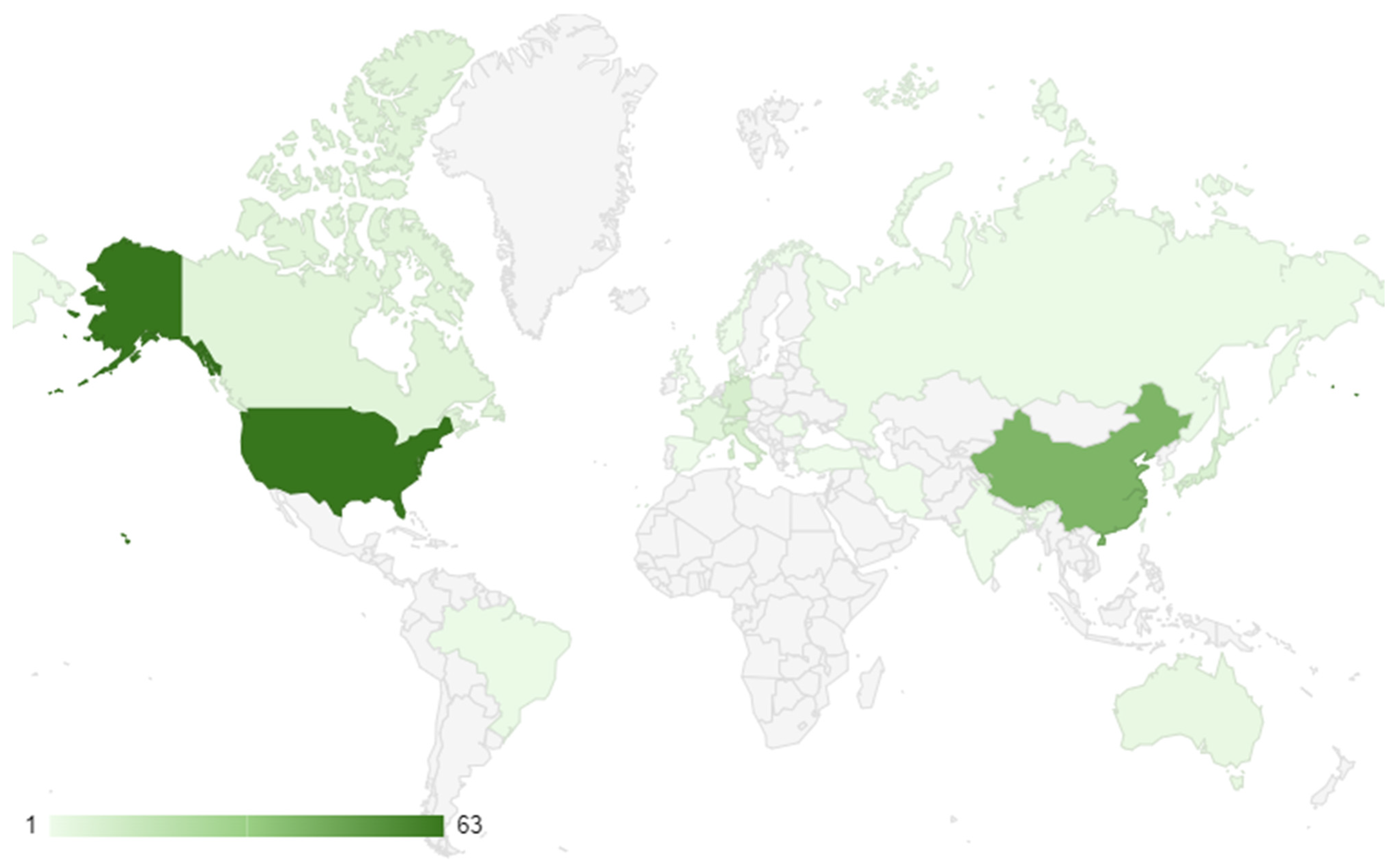
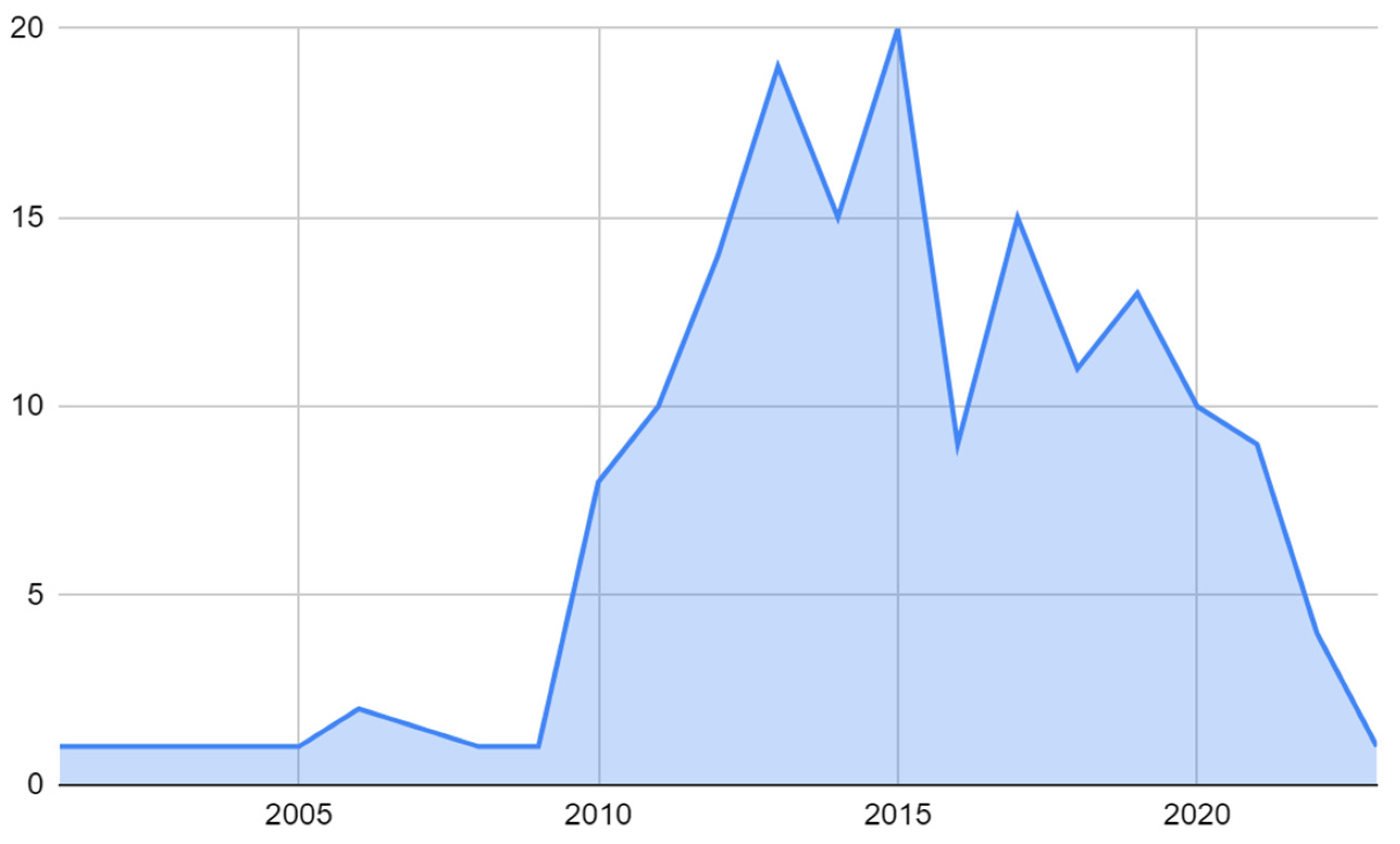
3.1. Global Research Trends
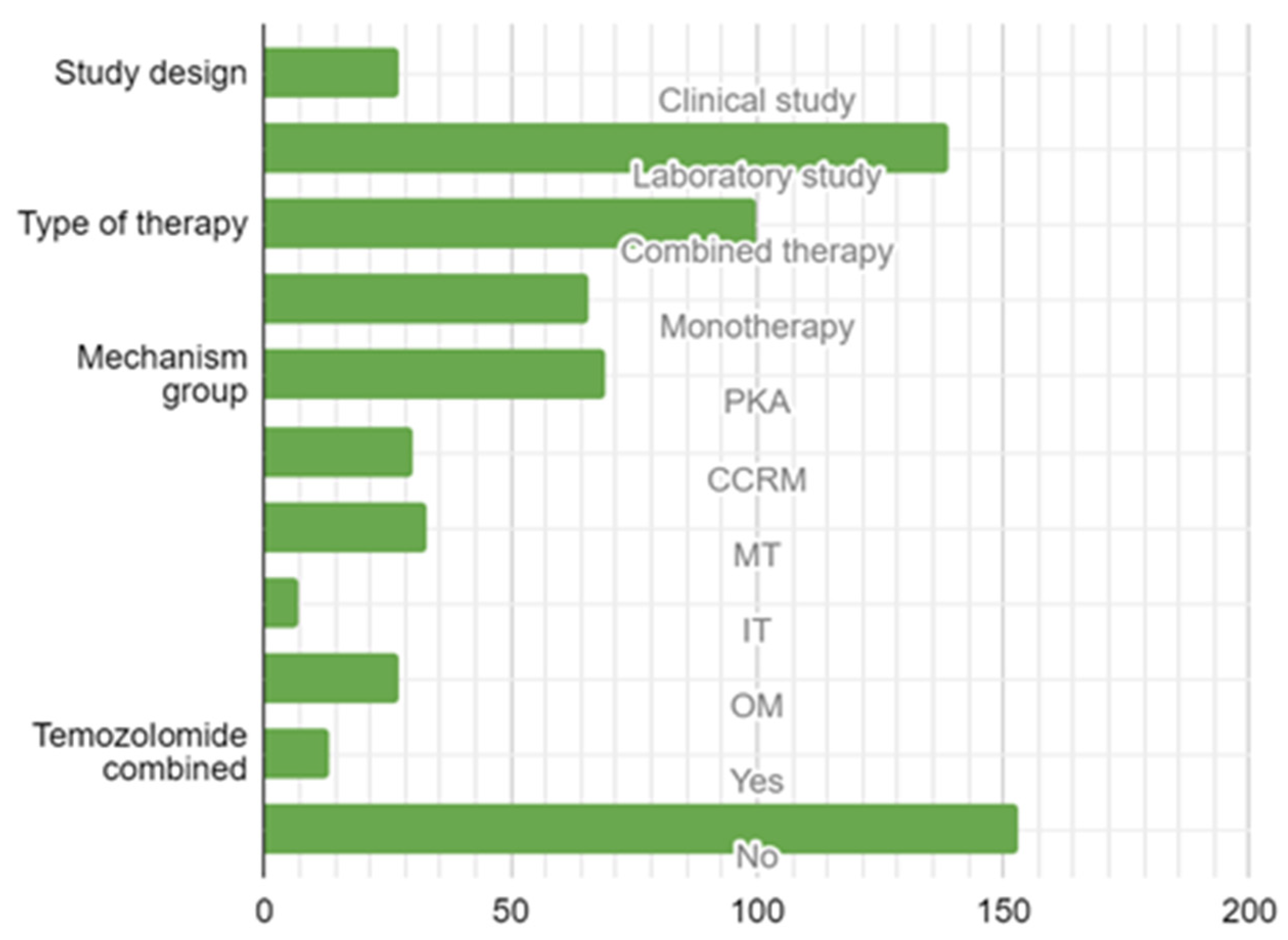
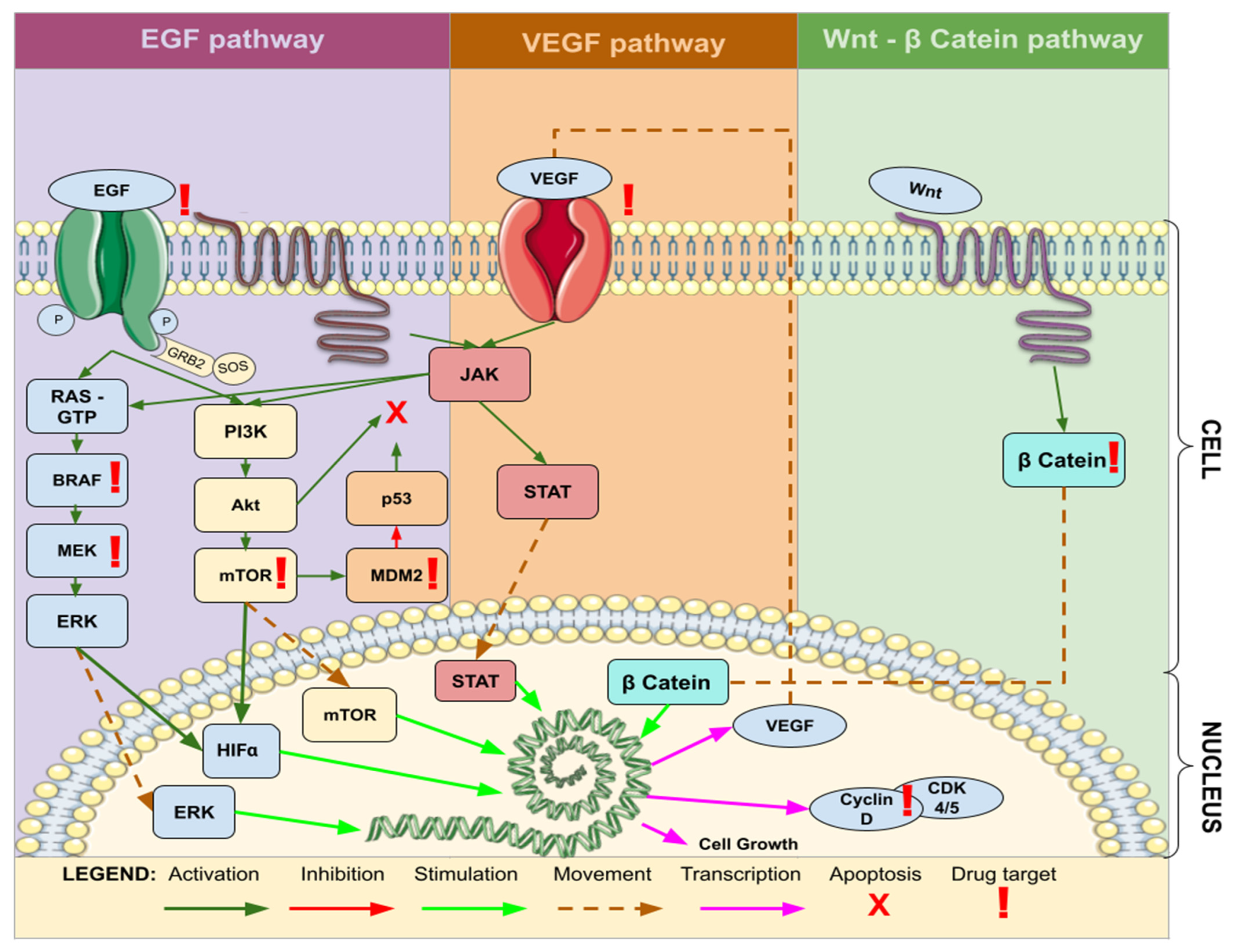
3.2. Study Design, Type of Target Therapy, and Molecular Mechanisms
3.3. Findings from Clinical Studies
3.4. Findings from Laboratory Studies
3.4.1. Overview of In Vitro Laboratory Studies
3.4.2. Overview of In Vivo Laboratory Studies
3.4.3. Overview of Combined Laboratory Studies
4. Discussion
4.1. Global and Research Trends of GBMs
4.2. Current State of Targeted Molecular Therapy in GBM Treatment
4.3. Effectiveness of Targeted Therapy in GBM Treatment
4.4. Promising Targeted Therapies for GBM Treatment
4.5. Advantages and Disadvantages in Molecular Targeted Therapy of GBM
4.6. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
| Search Strategy | |
| Search | (Glioblastoma multiforme OR GBM) AND (Molecular targeted therapy) |
| Filter | from 2000 to 2022 |
| Search details | ((“glioblastoma”[MeSH Terms] OR “glioblastoma”[All Fields] OR (“glioblastoma”[All Fields] AND “multiforme”[All Fields]) OR “glioblastoma multiforme”[All Fields] OR “GBM”[All Fields]) AND (“molecular targeted therapy”[MeSH Terms] OR (“molecular”[All Fields] AND “targeted”[All Fields] AND “therapy”[All Fields]) OR “molecular targeted therapy”[All Fields] OR (“protein kinase inhibitors”[Pharmacological Action] OR “protein kinase inhibitors”[MeSH Terms] OR (“protein”[All Fields] AND “kinase”[All Fields] AND “inhibitors”[All Fields]) OR “protein kinase inhibitors”[All Fields]) OR (“immunotherapy”[MeSH Terms] OR “immunotherapy”[All Fields] OR “immunotherapies”[All Fields] OR “immunotherapy s”[All Fields]) OR (“apoptosis”[MeSH Terms] OR “apoptosis”[All Fields]))) AND (2000:2022[pdat]) |
References
- Kanderi, T.; Gupta, V. Glioblastoma Multiforme. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac. J. Cancer Prev. 2017, 18, 3–9. [Google Scholar] [CrossRef]
- Stoyanov, G.S.; Dzhenkov, D.L. On the Concepts and History of Glioblastoma Multiforme—Morphology, Genetics and Epigenetics. Folia Med. 2018, 60, 48–66. [Google Scholar] [CrossRef]
- McCutcheon, I.E.; Preul, M.C. Historical Perspective on Surgery and Survival with Glioblastoma: How Far Have We Come? World Neurosurg. 2021, 149, 148–168. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, A.; Proietti, G.; Sica, G.; Scicchitano, B.M. Pathological and Molecular Features of Glioblastoma and Its Peritumoral Tissue. Cancers 2019, 11, 469. [Google Scholar] [CrossRef] [PubMed]
- Inda, M.-d.-M.; Bonavia, R.; Seoane, J. Glioblastoma Multiforme: A Look Inside Its Heterogeneous Nature. Cancers 2014, 6, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Scheithauer, B.W. Development of the WHO classification of tumors of the central nervous system: A historical perspective. Brain Pathol. 2009, 19, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, G.S.; Lyutfi, E.; Georgieva, R.; Georgiev, R.; Dzhenkov, D.L.; Petkova, L.; Ivanov, B.D.; Kaprelyan, A.; Ghenev, P. Reclassification of Glioblastoma Multiforme According to the 2021 World Health Organization Classification of Central Nervous System Tumors: A Single Institution Report and Practical Significance. Cureus 2022, 14, e21822. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Costa, A.; Osório, L.; Lago, R.C.; Linhares, P.; Carvalho, B.; Caeiro, C. Current Standards of Care in Glioblastoma Therapy. In Glioblastoma; De Vleeschouwer, S., Ed.; Codon Publications: Brisbane, Australia, 2017. [Google Scholar]
- Jain, K.K. A Critical Overview of Targeted Therapies for Glioblastoma. Front. Oncol. 2018, 8, 419. [Google Scholar] [CrossRef] [PubMed]
- McBain, C.; Lawrie, T.A.; Rogozińska, E.; Kernohan, A.; Robinson, T.; Jefferies, S. Treatment options for progression or recurrence of glioblastoma: A network meta-analysis. Cochrane Database Syst. Rev. 2021, 5, Cd013579. [Google Scholar] [CrossRef]
- Mofatteh, M.; Mashayekhi, M.S.; Arfaie, S.; Chen, Y.; Malhotra, A.K.; Alvi, M.A.; Sader, N.; Antonick, V.; Fatehi Hassanabad, M.; Mansouri, A.; et al. Suicidal Ideation and Attempts in Brain Tumor Patients and Survivors: A Systematic Review. Neuro-Oncol. Adv. 2023, 5, vdad058. [Google Scholar] [CrossRef]
- Zhang, P.; Xia, Q.; Liu, L.; Li, S.; Dong, L. Current Opinion on Molecular Characterization for GBM Classification in Guiding Clinical Diagnosis, Prognosis, and Therapy. Front. Mol. Biosci. 2020, 7, 562798. [Google Scholar] [CrossRef]
- Clark, K.; Voronovich, Z.; Horbinski, C. How molecular testing can help (and hurt) in the workup of gliomas. Am. J. Clin. Pathol. 2013, 139, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Trembath, D.G. Chapter 26—Molecular Testing for Glioblastoma. In Diagnostic Molecular Pathology; Coleman, W.B., Tsongalis, G.J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 339–347. [Google Scholar]
- Hersh, A.M.; Gaitsch, H.; Alomari, S.; Lubelski, D.; Tyler, B.M. Molecular Pathways and Genomic Landscape of Glioblastoma Stem Cells: Opportunities for Targeted Therapy. Cancers 2022, 14, 3743. [Google Scholar] [CrossRef] [PubMed]
- Di Cintio, F.; Dal Bo, M.; Baboci, L.; De Mattia, E.; Polano, M.; Toffoli, G. The Molecular and Microenvironmental Landscape of Glioblastomas: Implications for the Novel Treatment Choices. Front. Neurosci. 2020, 14, 603647. [Google Scholar] [CrossRef] [PubMed]
- Wagle, N.; Nguyen, M.; Carrillo, J.; Truong, J.; Dobrawa, L.; Kesari, S. Characterization of molecular pathways for targeting therapy in glioblastoma. Chin. Clin. Oncol. 2020, 9, 77. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.W.; Verhaak, R.G.W.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The Somatic Genomic Landscape of Glioblastoma. Cell 2013, 155, 462–477. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Abdul Rahim, S.A.; Dirkse, A.; Oudin, A.; Schuster, A.; Bohler, J.; Barthelemy, V.; Muller, A.; Vallar, L.; Janji, B.; Golebiewska, A. Regulation of hypoxia-induced autophagy in glioblastoma involves ATG9A. Br. J. Cancer 2017, 117, 813–825. [Google Scholar] [CrossRef]
- Aldea, M.D.; Petrushev, B.; Soritau, O.; Tomuleasa, C.I.; Berindan-Neagoe, I.; Filip, A.G.; Chereches, G.; Cenariu, M.; Craciun, L.; Tatomir, C.; et al. Metformin plus sorafenib highly impacts temozolomide resistant glioblastoma stem-like cells. J. Buon 2014, 19, 502–511. [Google Scholar]
- Angara, K.; Rashid, M.H.; Shankar, A.; Ara, R.; Iskander, A.; Borin, T.F.; Jain, M.; Achyut, B.R.; Arbab, A.S. Vascular mimicry in glioblastoma following anti-angiogenic and anti-20-HETE therapies. Histol. Histopathol. 2017, 32, 917. [Google Scholar]
- Anghileri, E.; Di Ianni, N.; Paterra, R.; Langella, T.; Zhao, J.; Eoli, M.; Patanè, M.; Pollo, B.; Cuccarini, V.; Iavarone, A. High tumor mutational burden and T-cell activation are associated with long-term response to anti-PD1 therapy in Lynch syndrome recurrent glioblastoma patient. Cancer Immunol. Immunother. 2021, 70, 831–842. [Google Scholar] [CrossRef]
- Arcella, A.; Biagioni, F.; Antonietta Oliva, M.; Bucci, D.; Frati, A.; Esposito, V.; Cantore, G.; Giangaspero, F.; Fornai, F. Rapamycin inhibits the growth of glioblastoma. Brain Res. 2013, 1495, 37–51. [Google Scholar] [CrossRef]
- Ariey-Bonnet, J.; Carrasco, K.; Le Grand, M.; Hoffer, L.; Betzi, S.; Feracci, M.; Tsvetkov, P.; Devred, F.; Collette, Y.; Morelli, X. In silico molecular target prediction unveils mebendazole as a potent MAPK14 inhibitor. Mol. Oncol. 2020, 14, 3083–3099. [Google Scholar] [CrossRef]
- Badruddoja, M.A.; Pazzi, M.; Sanan, A.; Schroeder, K.; Kuzma, K.; Norton, T.; Scully, T.; Mahadevan, D.; Ahmadi, M.M. Phase II study of bi-weekly temozolomide plus bevacizumab for adult patients with recurrent glioblastoma. Cancer Chemother. Pharmacol. 2017, 80, 715–721. [Google Scholar] [CrossRef]
- Bagca, B.G.; Ozates, N.P.; Asik, A.; Caglar, H.O.; Gunduz, C.; Avci, C.B. Temozolomide treatment combined with AZD3463 shows synergistic effect in glioblastoma cells. Biochem. Biophys. Res. Commun. 2020, 533, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Bähr, O.; Gross, S.; Harter, P.N.; Kirches, E.; Mawrin, C.; Steinbach, J.P.; Mittelbronn, M. ASA404, a vascular disrupting agent, as an experimental treatment approach for brain tumors. Oncol. Lett. 2017, 14, 5443–5451. [Google Scholar]
- Barbarisi, M.; Iaffaioli, R.V.; Armenia, E.; Schiavo, L.; De Sena, G.; Tafuto, S.; Barbarisi, A.; Quagliariello, V. Novel nanohydrogel of hyaluronic acid loaded with quercetin alone and in combination with temozolomide as new therapeutic tool, CD44 targeted based, of glioblastoma multiforme. J. Cell. Physiol. 2018, 233, 6550–6564. [Google Scholar] [CrossRef] [PubMed]
- Barone, A.; Sengupta, R.; Warrington, N.M.; Smith, E.; Wen, P.Y.; Brekken, R.A.; Romagnoli, B.; Douglas, G.; Chevalier, E.; Bauer, M.P. Combined VEGF and CXCR4 antagonism targets the GBM stem cell population and synergistically improves survival in an intracranial mouse model of glioblastoma. Oncotarget 2014, 5, 9811. [Google Scholar] [CrossRef] [PubMed]
- Benezra, M.; Hambardzumyan, D.; Penate-Medina, O.; Veach, D.R.; Pillarsetty, N.; Smith-Jones, P.; Phillips, E.; Ozawa, T.; Zanzonico, P.B.; Longo, V. Fluorine-labeled dasatinib nanoformulations as targeted molecular imaging probes in a PDGFB-driven murine glioblastoma model. Neoplasia 2012, 14, 1132–1143. [Google Scholar] [CrossRef] [PubMed]
- Blanco, V.M.; Chu, Z.; Vallabhapurapu, S.D.; Sulaiman, M.K.; Kendler, A.; Rixe, O.; Warnick, R.E.; Franco, R.S.; Qi, X. Phosphatidylserine-selective targeting and anticancer effects of SapC-DOPS nanovesicles on brain tumors. Oncotarget 2014, 5, 7105. [Google Scholar] [CrossRef]
- Blank, M.; Weinschenk, T.; Priemer, M.; Schluesener, H. Systematic Evolution of a DNA Aptamer Binding to Rat Brain Tumor Microvessels: SELECTIVE TARGETING OF ENDOTHELIAL REGULATORY PROTEIN PIGPEN. J. Biol. Chem. 2001, 276, 16464–16468. [Google Scholar] [CrossRef]
- Brown, N.; McBain, C.; Nash, S.; Hopkins, K.; Sanghera, P.; Saran, F.; Phillips, M.; Dungey, F.; Clifton-Hadley, L.; Wanek, K. Multi-center randomized phase II study comparing cediranib plus gefitinib with cediranib plus placebo in subjects with recurrent/progressive glioblastoma. PLoS ONE 2016, 11, e0156369. [Google Scholar] [CrossRef] [PubMed]
- Butowski, N.; Chang, S.M.; Lamborn, K.R.; Polley, M.Y.; Parvataneni, R.; Hristova-Kazmierski, M.; Musib, L.; Nicol, S.J.; Thornton, D.E.; Prados, M.D. Enzastaurin plus temozolomide with radiation therapy in glioblastoma multiforme: A phase I study. Neuro-Oncology 2010, 12, 608–613. [Google Scholar] [CrossRef]
- Bychkov, M.; Shulepko, M.; Osmakov, D.; Andreev, Y.; Sudarikova, A.; Vasileva, V.; Pavlyukov, M.S.; Latyshev, Y.A.; Potapov, A.A.; Kirpichnikov, M. Mambalgin-2 induces cell cycle arrest and apoptosis in glioma cells via interaction with ASIC1a. Cancers 2020, 12, 1837. [Google Scholar] [CrossRef]
- Camorani, S.; Crescenzi, E.; Colecchia, D.; Carpentieri, A.; Amoresano, A.; Fedele, M.; Chiariello, M.; Cerchia, L. Aptamer targeting EGFRvIII mutant hampers its constitutive autophosphorylation and affects migration, invasion and proliferation of glioblastoma cells. Oncotarget 2015, 6, 37570. [Google Scholar] [CrossRef]
- Caruana, B.T.; Skoric, A.; Brown, A.J.; Lutze-Mann, L.H. Site-1 protease, a novel metabolic target for glioblastoma. Biochem. Biophys. Res. Commun. 2017, 490, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Chen, P.-Y.; Lin, Y.-Y.; Feng, L.-Y.; Chen, S.-H.; Chen, C.-Y.; Huang, Y.-C.; Huang, C.-Y.; Jung, S.-M.; Chen, L.Y. Suppression of tumor growth via IGFBP3 depletion as a potential treatment in glioma. J. Neurosurg. 2019, 132, 168–179. [Google Scholar] [CrossRef]
- Chen, L.; Miao, W.; Tang, X.; Zhang, H.; Wang, S.; Luo, F.; Yan, J. Inhibitory effect of neuropilin-1 monoclonal antibody (NRP-1 MAb) on glioma tumor in mice. J. Biomed. Nanotechnol. 2013, 9, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Chen, J.; Zhu, Y.; Li, Y.; Wang, Y.; Chen, H.; Wang, J.; Li, X.; Liu, Y.; Li, B. CD163, a novel therapeutic target, regulates the proliferation and stemness of glioma cells via casein kinase 2. Oncogene 2019, 38, 1183–1199. [Google Scholar] [CrossRef]
- Chen, W.; Wu, M.; Cui, S.-T.; Zheng, Y.; Liu, Z.; Luo, L.-S. CircRNA Circ-ITCH inhibits the proliferation and invasion of glioma cells through targeting the miR-106a-5p/SASH1 Axis. Cell Transplant. 2021, 30, 0963689720983785. [Google Scholar] [CrossRef]
- Chen, Z.; Pan, X.; Georgakilas, A.G.; Chen, P.; Hu, H.; Yang, Y.; Tian, S.; Xia, L.; Zhang, J.; Cai, X.; et al. Tetramethylpyrazine (TMP) protects cerebral neurocytes and inhibits glioma by down regulating chemokine receptor CXCR4 expression. Cancer Lett. 2013, 336, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Ren, Z.; Liu, Z.; Sun, X.; Qian, R.; Cao, C.; Liu, B.; Wang, J.; Wang, H.; Guo, Y. Cysteine cathepsin C: A novel potential biomarker for the diagnosis and prognosis of glioma. Cancer Cell Int. 2022, 22, 53. [Google Scholar] [CrossRef] [PubMed]
- Chinnaiyan, P.; Won, M.; Wen, P.Y.; Rojiani, A.M.; Wendland, M.; Dipetrillo, T.A.; Corn, B.W.; Mehta, M.P. RTOG 0913: A phase 1 study of daily everolimus (RAD001) in combination with radiation therapy and temozolomide in patients with newly diagnosed glioblastoma. Int. J. Radiat. Oncol. Biol. Phys. 2013, 86, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Ciesielski, M.J.; Bu, Y.; Munich, S.A.; Teegarden, P.; Smolinski, M.P.; Clements, J.L.; Lau, J.Y.; Hangauer, D.G.; Fenstermaker, R.A. KX2-361: A novel orally bioavailable small molecule dual Src/tubulin inhibitor that provides long term survival in a murine model of glioblastoma. J. Neuro-Oncol. 2018, 140, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.L.; Molinaro, A.M.; Phillips, J.J.; Butowski, N.A.; Chang, S.M.; Perry, A.; Costello, J.F.; DeSilva, A.A.; Rabbitt, J.E.; Prados, M.D. A single-institution phase II trial of radiation, temozolomide, erlotinib, and bevacizumab for initial treatment of glioblastoma. Neuro-Oncology 2014, 16, 984–990. [Google Scholar] [CrossRef]
- Cloninger, C.; Bernath, A.; Bashir, T.; Holmes, B.; Artinian, N.; Ruegg, T.; Anderson, L.; Masri, J.; Lichtenstein, A.; Gera, J. Inhibition of SAPK2/p38 enhances sensitivity to mTORC1 inhibition by blocking IRES-mediated translation initiation in glioblastoma. Mol. Cancer Ther. 2011, 10, 2244–2256. [Google Scholar] [CrossRef]
- Colen, C.B.; Shen, Y.; Ghoddoussi, F.; Yu, P.; Francis, T.B.; Koch, B.J.; Monterey, M.D.; Galloway, M.P.; Sloan, A.E.; Mathupala, S.P. Metabolic targeting of lactate efflux by malignant glioma inhibits invasiveness and induces necrosis: An in vivo study. Neoplasia 2011, 13, 620–632. [Google Scholar] [CrossRef]
- D’Alessandris, Q.G.; Montano, N.; Cenci, T.; Martini, M.; Lauretti, L.; Bianchi, F.; Larocca, L.M.; Maira, G.; Fernandez, E.; Pallini, R. Targeted therapy with bevacizumab and erlotinib tailored to the molecular profile of patients with recurrent glioblastoma. Preliminary experience. Acta Neurochir. 2013, 155, 33–40. [Google Scholar] [CrossRef]
- Desjardins, A.; Reardon, D.A.; Coan, A.; Marcello, J.; Herndon Ii, J.E.; Bailey, L.; Peters, K.B.; Friedman, H.S.; Vredenburgh, J.J. Bevacizumab and daily temozolomide for recurrent glioblastoma. Cancer 2012, 118, 1302–1312. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.; Reardon, D.A.; Peters, K.B.; Threatt, S.; Coan, A.D.; Herndon, J.E., 2nd; Friedman, A.H.; Friedman, H.S.; Vredenburgh, J.J. A phase I trial of the farnesyl transferase inhibitor, SCH 66336, with temozolomide for patients with malignant glioma. J. Neurooncol. 2011, 105, 601–606. [Google Scholar] [CrossRef]
- Di Stefano, A.L.; Fucci, A.; Frattini, V.; Labussiere, M.; Mokhtari, K.; Zoppoli, P.; Marie, Y.; Bruno, A.; Boisselier, B.; Giry, M. Detection, characterization, and inhibition of FGFR–TACC fusions in IDH wild-type glioma. Clin. Cancer Res. 2015, 21, 3307–3317. [Google Scholar] [CrossRef]
- Dominguez, C.L.; Floyd, D.H.; Xiao, A.; Mullins, G.R.; Kefas, B.A.; Xin, W.; Yacur, M.N.; Abounader, R.; Lee, J.K.; Wilson, G.M.; et al. Diacylglycerol Kinase α Is a Critical Signaling Node and Novel Therapeutic Target in Glioblastoma and Other Cancers. Cancer Discov. 2013, 3, 782–797. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Zhou, J.-R.; Wang, D.-L.; Gong, K.; Zhang, Q.-J. Vitamin K1 enhances sorafenib-induced growth inhibition and apoptosis of human malignant glioma cells by blocking the Raf/MEK/ERK pathway. World J. Surg. Oncol. 2012, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Emlet, D.R.; Gupta, P.; Holgado-Madruga, M.; Del Vecchio, C.A.; Mitra, S.S.; Han, S.Y.; Li, G.; Jensen, K.C.; Vogel, H.; Xu, L.W.; et al. Targeting a glioblastoma cancer stem-cell population defined by EGF receptor variant III. Cancer Res. 2014, 74, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Farrell, P.J.; Matuszkiewicz, J.; Balakrishna, D.; Pandya, S.; Hixon, M.S.; Kamran, R.; Chu, S.; Lawson, J.D.; Okada, K.; Hori, A. MET tyrosine kinase inhibition enhances the antitumor efficacy of an HGF antibody. Mol. Cancer Ther. 2017, 16, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Huang, J.; Ding, Y.; Xie, F.; Shen, X. Tamoxifen-induced apoptosis of rat C6 glioma cells via PI3K/Akt, JNK and ERK activation. Oncol. Rep. 2010, 24, 1561–1567. [Google Scholar] [CrossRef]
- Fleurence, J.; Cochonneau, D.; Fougeray, S.; Oliver, L.; Geraldo, F.; Terme, M.; Dorvillius, M.; Loussouarn, D.; Vallette, F.; Paris, F. Targeting and killing glioblastoma with monoclonal antibody to O-acetyl GD2 ganglioside. Oncotarget 2016, 7, 41172. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.G.; Moretti, I.F.; Marie, S.K. Mitochondria transcription factor a: A putative target for the effect of melatonin on U87MG malignant glioma cell line. Molecules 2018, 23, 1129. [Google Scholar] [CrossRef]
- Ge, Y.-F.; Sun, J.; Jin, C.-J.; Cao, B.-Q.; Jiang, Z.-F.; Shao, J.-F. AntagomiR-27a targets FOXO3a in glioblastoma and suppresses U87 cell growth in vitro and in vivo. Asian Pac. J. Cancer Prev. 2013, 14, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Geletneky, K.; Hajda, J.; Angelova, A.L.; Leuchs, B.; Capper, D.; Bartsch, A.J.; Neumann, J.-O.; Schöning, T.; Hüsing, J.; Beelte, B. Oncolytic H-1 parvovirus shows safety and signs of immunogenic activity in a first phase I/IIa glioblastoma trial. Mol. Ther. 2017, 25, 2620–2634. [Google Scholar] [CrossRef]
- Genoud, V.; Espinoza, F.I.; Marinari, E.; Rochemont, V.; Dietrich, P.-Y.; McSheehy, P.; Bachmann, F.; Lane, H.A.; Walker, P.R. Treating ICB-resistant glioma with anti-CD40 and mitotic spindle checkpoint controller BAL101553 (lisavanbulin). JCI Insight 2021, 6, e142980. [Google Scholar] [CrossRef]
- Goswami, S.; Walle, T.; Cornish, A.E.; Basu, S.; Anandhan, S.; Fernandez, I.; Vence, L.; Blando, J.; Zhao, H.; Yadav, S.S. Immune profiling of human tumors identifies CD73 as a combinatorial target in glioblastoma. Nat. Med. 2020, 26, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Grinshtein, N.; Rioseco, C.C.; Marcellus, R.; Uehling, D.; Aman, A.; Lun, X.; Muto, O.; Podmore, L.; Lever, J.; Shen, Y. Small molecule epigenetic screen identifies novel EZH2 and HDAC inhibitors that target glioblastoma brain tumor-initiating cells. Oncotarget 2016, 7, 59360. [Google Scholar] [CrossRef]
- Grossman, R.; Tyler, B.; Rudek, M.A.; Kim, E.; Zadnik, P.; Khan, U.; Blakeley, J.O.; Pathak, A.P.; Brem, H. Microdialysis measurement of intratumoral temozolomide concentration after cediranib, a pan-VEGF receptor tyrosine kinase inhibitor, in a U87 glioma model. Cancer Chemother. Pharmacol. 2013, 72, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Wang, C.; Wang, X.; Ma, G.; Li, Y.; Cui, L.; Chen, Y.; Zhao, B.; Li, K. Efficient inhibition of human glioma development by RNA interference-mediated silencing of PAK5. Int. J. Biol. Sci. 2015, 11, 230. [Google Scholar] [CrossRef]
- Guo, L.; Fan, L.; Pang, Z.; Ren, J.; Ren, Y.; Li, J.; Chen, J.; Wen, Z.; Jiang, X. TRAIL and doxorubicin combination enhances anti-glioblastoma effect based on passive tumor targeting of liposomes. J. Control. Release 2011, 154, 93–102. [Google Scholar] [CrossRef]
- Harford-Wright, E.; Andre-Gregoire, G.; Jacobs, K.A.; Treps, L.; Le Gonidec, S.; Leclair, H.M.; Gonzalez-Diest, S.; Roux, Q.; Guillonneau, F.; Loussouarn, D. Pharmacological targeting of apelin impairs glioblastoma growth. Brain 2017, 140, 2939–2954. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, N.; Tsuboi, A.; Kagawa, N.; Chiba, Y.; Izumoto, S.; Kinoshita, M.; Kijima, N.; Oka, Y.; Morimoto, S.; Nakajima, H. Wilms tumor 1 peptide vaccination combined with temozolomide against newly diagnosed glioblastoma: Safety and impact on immunological response. Cancer Immunol. Immunother. 2015, 64, 707–716. [Google Scholar] [CrossRef]
- Hasselbalch, B.; Eriksen, J.G.; Broholm, H.; Christensen, I.J.; Grunnet, K.; Horsman, M.R.; Poulsen, H.S.; Stockhausen, M.T.; Lassen, U. Prospective evaluation of angiogenic, hypoxic and EGFR-related biomarkers in recurrent glioblastoma multiforme treated with cetuximab, bevacizumab and irinotecan. APMIS 2010, 118, 585–594. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Jabouille, A.; Steri, V.; Johansson-Percival, A.; Michael, I.P.; Kotamraju, V.R.; Junckerstorff, R.; Nowak, A.K.; Hamzah, J.; Lee, G. Vascular targeting of LIGHT normalizes blood vessels in primary brain cancer and induces intratumoural high endothelial venules. J. Pathol. 2018, 245, 209–221. [Google Scholar] [CrossRef]
- He, H.; Yao, M.; Zhang, W.; Tao, B.; Liu, F.; Li, S.; Dong, Y.; Zhang, C.; Meng, Y.; Li, Y. MEK2 is a prognostic marker and potential chemo-sensitizing target for glioma patients undergoing temozolomide treatment. Cell. Mol. Immunol. 2016, 13, 658–668. [Google Scholar] [CrossRef]
- Hong, H.; Stastny, M.; Brown, C.; Chang, W.-C.; Ostberg, J.R.; Forman, S.J.; Jensen, M.C. Diverse solid tumors expressing a restricted epitope of L1-CAM can be targeted by chimeric antigen receptor redirected T lymphocytes. J. Immunother. 2014, 37, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Xu, T.; Wang, Y.; Zhou, Y.; Yu, D.; Wang, Z.; He, L.; Chen, Z.; Zhang, Y.; Davidson, D. Cannabidiol inhibits human glioma by induction of lethal mitophagy through activating TRPV4. Autophagy 2021, 17, 3592–3606. [Google Scholar] [CrossRef]
- Huveldt, D.; Lewis-Tuffin, L.J.; Carlson, B.L.; Schroeder, M.A.; Rodriguez, F.; Giannini, C.; Galanis, E.; Sarkaria, J.N.; Anastasiadis, P.Z. Targeting Src family kinases inhibits bevacizumab-induced glioma cell invasion. PLoS ONE 2013, 8, e56505. [Google Scholar] [CrossRef] [PubMed]
- Ishiwata, T.; Teduka, K.; Yamamoto, T.; Kawahara, K.; Matsuda, Y.; Naito, Z. Neuroepithelial stem cell marker nestin regulates the migration, invasion and growth of human gliomas. Oncol. Rep. 2011, 26, 91–99. [Google Scholar] [CrossRef]
- Jaszberenyi, M.; Schally, A.V.; Block, N.L.; Zarandi, M.; Cai, R.-Z.; Vidaurre, I.; Szalontay, L.; Jayakumar, A.R.; Rick, F.G. Suppression of the proliferation of human U-87 MG glioblastoma cells by new antagonists of growth hormone-releasing hormone in vivo and in vitro. Target. Oncol. 2013, 8, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Wang, H.; Zhu, J.; Tang, Y.; Zhou, Y.; Zhu, L.; Gao, C.; Li, W.; You, W.; Yu, B.; et al. Correlation of Nrf2 and HIF-1α in glioblastoma and their relationships to clinicopathologic features and survival. Neurol. Res. 2013, 35, 1044–1050. [Google Scholar] [CrossRef]
- Jin, R.; Nakada, M.; Teng, L.; Furuta, T.; Sabit, H.; Hayashi, Y.; Demuth, T.; Hirao, A.; Sato, H.; Zhao, G.; et al. Combination therapy using Notch and Akt inhibitors is effective for suppressing invasion but not proliferation in glioma cells. Neurosci. Lett. 2013, 534, 316–321. [Google Scholar] [CrossRef]
- Joel, M.; Mughal, A.A.; Grieg, Z.; Murrell, W.; Palmero, S.; Mikkelsen, B.; Fjerdingstad, H.B.; Sandberg, C.J.; Behnan, J.; Glover, J.C. Targeting PBK/TOPK decreases growth and survival of glioma initiating cells in vitro and attenuates tumor growth in vivo. Mol. Cancer 2015, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Johanns, T.M.; Ferguson, C.J.; Grierson, P.M.; Dahiya, S.; Ansstas, G. Rapid clinical and radiographic response with combined dabrafenib and trametinib in adults with BRAF-mutated high-grade glioma. J. Natl. Compr. Cancer Netw. 2018, 16, 4–10. [Google Scholar] [CrossRef]
- Joshi, A.D.; Botham, R.C.; Schlein, L.J.; Roth, H.S.; Mangraviti, A.; Borodovsky, A.; Tyler, B.; Joslyn, S.; Looper, J.S.; Podell, M. Synergistic and targeted therapy with a procaspase-3 activator and temozolomide extends survival in glioma rodent models and is feasible for the treatment of canine malignant glioma patients. Oncotarget 2017, 8, 80124–80138. [Google Scholar] [CrossRef]
- Ju, R.-J.; Zeng, F.; Liu, L.; Mu, L.-M.; Xie, H.-J.; Zhao, Y.; Yan, Y.; Wu, J.-S.; Hu, Y.-J.; Lu, W.-L. Destruction of vasculogenic mimicry channels by targeting epirubicin plus celecoxib liposomes in treatment of brain glioma. Int. J. Nanomed. 2016, 11, 1131–1146. [Google Scholar]
- Junca, A.; Villalva, C.; Tachon, G.; Rivet, P.; Cortes, U.; Guilloteau, K.; Balbous, A.; Godet, J.; Wager, M.; Karayan-Tapon, L. Crizotinib targets in glioblastoma stem cells. Cancer Med. 2017, 6, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Park, H.; Zhao, H.-Y.; Jeon, R.; Ryu, J.-H.; Kim, W.-Y. Systemic approaches identify a garlic-derived chemical, Z-ajoene, as a glioblastoma multiforme cancer stem cell-specific targeting agent. Mol. Cells 2014, 37, 547. [Google Scholar] [CrossRef] [PubMed]
- Kanemaru, Y.; Natsumeda, M.; Okada, M.; Saito, R.; Kobayashi, D.; Eda, T.; Watanabe, J.; Saito, S.; Tsukamoto, Y.; Oishi, M. Dramatic response of BRAF V600E-mutant epithelioid glioblastoma to combination therapy with BRAF and MEK inhibitor: Establishment and xenograft of a cell line to predict clinical efficacy. Acta Neuropathol. Commun. 2019, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Kaneta, Y.; Ullrich, A. NEK9 depletion induces catastrophic mitosis by impairment of mitotic checkpoint control and spindle dynamics. Biochem. Biophys. Res. Commun. 2013, 442, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Kawauchi, D.; Takahashi, M.; Satomi, K.; Yamamuro, S.; Kobayashi, T.; Uchida, E.; Honda-Kitahara, M.; Narita, Y.; Iwadate, Y.; Ichimura, K. The ALK inhibitors, alectinib and ceritinib, induce ALK-independent and STAT3-dependent glioblastoma cell death. Cancer Sci. 2021, 112, 2442–2453. [Google Scholar] [CrossRef]
- Kim, S.-S.; Harford, J.B.; Moghe, M.; Rait, A.; Pirollo, K.F.; Chang, E.H. Targeted nanocomplex carrying siRNA against MALAT1 sensitizes glioblastoma to temozolomide. Nucleic Acids Res. 2018, 46, 1424–1440. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Ai, C.; Dong, F.; Xia, X.; Zhao, X.; Yang, C.; Kang, C.; Zhou, Y.; Zhao, Q.; Sun, X. Targeting of BMI-1 with PTC-209 inhibits glioblastoma development. Cell Cycle 2018, 17, 1199–1211. [Google Scholar] [CrossRef]
- Koul, D.; Shen, R.; Bergh, S.; Lu, Y.; de Groot, J.F.; Liu, T.J.; Mills, G.B.; Yung, W.A. Targeting integrin-linked kinase inhibits Akt signaling pathways and decreases tumor progression of human glioblastoma. Mol. Cancer Ther. 2005, 4, 1681–1688. [Google Scholar] [CrossRef] [PubMed]
- Koul, D.; Shen, R.; Kim, Y.-W.; Kondo, Y.; Lu, Y.; Bankson, J.; Ronen, S.M.; Kirkpatrick, D.L.; Powis, G.; Yung, W.A. Cellular and in vivo activity of a novel PI3K inhibitor, PX-866, against human glioblastoma. Neuro-Oncology 2010, 12, 559–569. [Google Scholar] [CrossRef]
- Kuan, C.-T.; Wakiya, K.; Herndon, J.E.; Lipp, E.S.; Pegram, C.N.; Riggins, G.J.; Rasheed, A.; Szafranski, S.E.; McLendon, R.E.; Wikstrand, C.J. MRP3: A molecular target for human glioblastoma multiforme immunotherapy. BMC Cancer 2010, 10, 468. [Google Scholar] [CrossRef] [PubMed]
- Lamour, V.; Henry, A.; Kroonen, J.; Nokin, M.J.; von Marschall, Z.; Fisher, L.W.; Chau, T.L.; Chariot, A.; Sanson, M.; Delattre, J.Y. Targeting osteopontin suppresses glioblastoma stem-like cell character and tumorigenicity in vivo. Int. J. Cancer 2015, 137, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Lassen, U.; Chinot, O.L.; McBain, C.; Mau-Sørensen, M.; Larsen, V.A.; Barrie, M.; Roth, P.; Krieter, O.; Wang, K.; Habben, K. Phase 1 dose-escalation study of the antiplacental growth factor monoclonal antibody RO5323441 combined with bevacizumab in patients with recurrent glioblastoma. Neuro-Oncology 2015, 17, 1007–1015. [Google Scholar] [CrossRef][Green Version]
- Lee, C.; Fotovati, A.; Triscott, J.; Chen, J.; Venugopal, C.; Singhal, A.; Dunham, C.; Kerr, J.M.; Verreault, M.; Yip, S.; et al. Polo-like kinase 1 inhibition kills glioblastoma multiforme brain tumor cells in part through loss of SOX2 and delays tumor progression in mice. Stem Cells 2012, 30, 1064–1075. [Google Scholar] [CrossRef]
- Lee, C.H.; Alpert, B.O.; Sankaranarayanan, P.; Alter, O. GSVD comparison of patient-matched normal and tumor aCGH profiles reveals global copy-number alterations predicting glioblastoma multiforme survival. PLoS ONE 2012, 7, e30098. [Google Scholar] [CrossRef]
- Lescarbeau, R.S.; Lei, L.; Bakken, K.K.; Sims, P.A.; Sarkaria, J.N.; Canoll, P.; White, F.M. Quantitative phosphoproteomics reveals Wee1 kinase as a therapeutic target in a model of proneural glioblastoma. Mol. Cancer Ther. 2016, 15, 1332–1343. [Google Scholar] [CrossRef]
- Li, C.; Shen, J.; Wei, X.; Xie, C.; Lu, W. Targeted delivery of a novel palmitylated D-peptide for antiglioblastoma molecular therapy. J. Drug Target. 2012, 20, 264–271. [Google Scholar] [CrossRef]
- Lian, S.; Shi, R.; Bai, T.; Liu, Y.; Miao, W.; Wang, H.; Liu, X.; Fan, Y. Anti-miRNA-23a oligonucleotide suppresses glioma cells growth by targeting apoptotic protease activating factor-1. Curr. Pharm. Des. 2013, 19, 6382–6389. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhou, L.; Shi, S.; Wang, Y.; Ni, X.; Xiao, F.; Wang, S.; Li, P.; Ding, K. Oligosaccharide G19 inhibits U-87 MG human glioma cells growth in vitro and in vivo by targeting epidermal growth factor (EGF) and activating p53/p21 signaling. Glycobiology 2014, 24, 748–765. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, X.; Feng, X.; Zhang, B.; Wang, J. Adenovirus-mediated delivery of bFGF small interfering RNA reduces STAT3 phosphorylation and induces the depolarization of mitochondria and apoptosis in glioma cells U251. J. Exp. Clin. Cancer Res. 2011, 30, 80. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Shi, L.; Shan, Q.; Cao, Q.; Yue, C.; Li, H.; Li, S.; Wang, J.; Gao, S. The third-generation EGFR inhibitor AZD9291 overcomes primary resistance by continuously blocking ERK signaling in glioblastoma. J. Exp. Clin. Cancer Res. 2019, 38, 219. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chhipa, R.R.; Nakano, I.; Dasgupta, B. The AMPK inhibitor compound C is a potent AMPK-independent antiglioma agent. Mol. Cancer Ther. 2014, 13, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, G.; De Salvo, G.L.; Brandes, A.A.; Eoli, M.; Rudà, R.; Faedi, M.; Lolli, I.; Pace, A.; Daniele, B.; Pasqualetti, F. Regorafenib compared with lomustine in patients with relapsed glioblastoma (REGOMA): A multicentre, open-label, randomised, controlled, phase 2 trial. Lancet Oncol. 2019, 20, 110–119. [Google Scholar] [CrossRef]
- Loskutov, Y.V.; Griffin, C.L.; Marinak, K.M.; Bobko, A.; Margaryan, N.V.; Geldenhuys, W.J.; Sarkaria, J.N.; Pugacheva, E.N. LPA signaling is regulated through the primary cilium: A novel target in glioblastoma. Oncogene 2018, 37, 1457–1471. [Google Scholar] [CrossRef]
- Luchman, H.A.; Stechishin, O.D.; Dang, N.H.; Blough, M.D.; Chesnelong, C.; Kelly, J.J.; Nguyen, S.A.; Chan, J.A.; Weljie, A.M.; Cairncross, J.G. An in vivo patient-derived model of endogenous IDH1-mutant glioma. Neuro-Oncology 2011, 14, 184–191. [Google Scholar] [CrossRef]
- Luwor, R.; Morokoff, A.P.; Amiridis, S.; D’Abaco, G.; Paradiso, L.; Stylli, S.S.; Nguyen, H.P.; Tarleton, M.; Young, K.A.; O’Brien, T.J. Targeting glioma stem cells by functional inhibition of dynamin 2: A novel treatment strategy for glioblastoma. Cancer Investig. 2019, 37, 144–155. [Google Scholar] [CrossRef]
- Balkhi, H.M.; Gul, T.; Haq, E. Anti-neoplastic and calcium modulatory action of caffeic acid phenethyl ester and dasatinib in C6 glial cells: A therapeutic perspective. CNS Neurol. Disord.-Drug Targets Former. Curr. Drug Targets-CNS Neurol. Disord. 2016, 15, 54–63. [Google Scholar] [CrossRef]
- Ma, J.-W.; Zhang, Y.; Li, R.; Ye, J.-C.; Li, H.-Y.; Zhang, Y.-K.; Ma, Z.-L.; Li, J.-Y.; Zhong, X.-Y.; Yang, X. Tetrandrine suppresses human glioma growth by inhibiting cell survival, proliferation and tumour angiogenesis through attenuating STAT3 phosphorylation. Eur. J. Pharmacol. 2015, 764, 228–239. [Google Scholar] [CrossRef]
- Mao, P.; Hever-Jardine, M.P.; Rahme, G.J.; Yang, E.; Tam, J.; Kodali, A.; Biswal, B.; Fadul, C.E.; Gaur, A.; Israel, M.A. Serine/threonine kinase 17A is a novel candidate for therapeutic targeting in glioblastoma. PLoS ONE 2013, 8, e81803. [Google Scholar] [CrossRef]
- Mason, W.P.; MacNeil, M.; Kavan, P.; Easaw, J.; Macdonald, D.; Thiessen, B.; Urva, S.; Lwin, Z.; McIntosh, L.; Eisenhauer, E. A phase I study of temozolomide and everolimus (RAD001) in patients with newly diagnosed and progressive glioblastoma either receiving or not receiving enzyme-inducing anticonvulsants: An NCIC CTG study. Investig. New Drugs 2012, 30, 2344–2351. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.-I.; Sato, A.; Okada, M.; Shibuya, K.; Seino, S.; Suzuki, K.; Watanabe, E.; Narita, Y.; Shibui, S.; Kayama, T. Targeting JNK for therapeutic depletion of stem-like glioblastoma cells. Sci. Rep. 2012, 2, 516. [Google Scholar] [CrossRef]
- Maxwell, M.J.; Arnold, A.; Sweeney, H.; Chen, L.; Lih, T.-S.M.; Schnaubelt, M.; Eberhart, C.G.; Rubens, J.A.; Zhang, H.; Clark, D.J. Unbiased proteomic and phosphoproteomic analysis identifies response signatures and novel susceptibilities after combined MEK and mTOR inhibition in BRAFV600E mutant glioma. Mol. Cell. Proteom. 2021, 20, 100123. [Google Scholar] [CrossRef]
- Merlino, F.; Daniele, S.; La Pietra, V.; Di Maro, S.; Di Leva, F.S.; Brancaccio, D.; Tomassi, S.; Giuntini, S.; Cerofolini, L.; Fragai, M. Simultaneous targeting of RGD-integrins and dual murine double minute proteins in glioblastoma multiforme. J. Med. Chem. 2018, 61, 4791–4809. [Google Scholar] [CrossRef]
- Michaud, K.; Solomon, D.A.; Oermann, E.; Kim, J.S.; Zhong, W.Z.; Prados, M.D.; Ozawa, T.; James, C.D.; Waldman, T. Pharmacologic inhibition of cyclin-dependent kinases 4 and 6 arrests the growth of glioblastoma multiforme intracranial xenografts. Cancer Res. 2010, 70, 3228–3238. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Pan, Y.; Joshi, K.; Purohit, D.; Hu, B.; Demir, H.; Mazumder, S.; Okabe, S.; Yamori, T.; Viapiano, M.; et al. Telomestatin impairs glioma stem cell survival and growth through the disruption of telomeric G-quadruplex and inhibition of the proto-oncogene, c-Myb. Clin Cancer Res 2012, 18, 1268–1280. [Google Scholar] [CrossRef]
- Mojarad-Jabali, S.; Farshbaf, M.; Hemmati, S.; Sarfraz, M.; Motasadizadeh, H.; Mojarrad, J.S.; Atyabi, F.; Zakeri-Milani, P.; Valizadeh, H. Comparison of three synthetic transferrin mimetic small peptides to promote the blood–brain barrier penetration of vincristine liposomes for improved glioma targeted therapy. Int. J. Pharm. 2022, 613, 121395. [Google Scholar] [CrossRef]
- Nandhu, M.S.; Behera, P.; Bhaskaran, V.; Longo, S.L.; Barrera-Arenas, L.M.; Sengupta, S.; Rodriguez-Gil, D.J.; Chiocca, E.A.; Viapiano, M.S. Development of a function-blocking antibody against fibulin-3 as a targeted reagent for glioblastoma. Clin. Cancer Res. 2018, 24, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Nayak, L.; Molinaro, A.M.; Peters, K.; Clarke, J.L.; Jordan, J.T.; de Groot, J.; Nghiemphu, L.; Kaley, T.; Colman, H.; McCluskey, C. Randomized phase II and biomarker study of pembrolizumab plus bevacizumab versus pembrolizumab alone for patients with recurrent glioblastoma. Clin. Cancer Res. 2021, 27, 1048–1057. [Google Scholar] [CrossRef]
- Niu, M.; Cai, W.; Liu, H.; Chong, Y.; Hu, W.; Gao, S.; Shi, Q.; Zhou, X.; Liu, X.; Yu, R. Plumbagin inhibits growth of gliomas in vivo via suppression of FOXM1 expression. J. Pharmacol. Sci. 2015, 128, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Nonnenmacher, L.; Westhoff, M.A.; Fulda, S.; Karpel-Massler, G.; Halatsch, M.E.; Engelke, J.; Simmet, T.; Corbacioglu, S.; Debatin, K.M. RIST: A potent new combination therapy for glioblastoma. Int. J. Cancer 2015, 136, E173–E187. [Google Scholar] [CrossRef] [PubMed]
- Pall, A.E.; Juratli, L.; Guntur, D.; Bandyopadhyay, K.; Kondapalli, K.C. A gain of function paradox: Targeted therapy for glioblastoma associated with abnormal NHE9 expression. J. Cell. Mol. Med. 2019, 23, 7859–7872. [Google Scholar] [CrossRef] [PubMed]
- Paternot, S.; Roger, P.P. Combined inhibition of MEK and mammalian target of rapamycin abolishes phosphorylation of cyclin-dependent kinase 4 in glioblastoma cell lines and prevents their proliferation. Cancer Res. 2009, 69, 4577–4581. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Yang, C.; Liu, Y.; Shen, C. miR-25-3p promotes glioma cell proliferation and migration by targeting FBXW7 and DKK3. Exp. Ther. Med. 2019, 18, 769–778. [Google Scholar] [CrossRef]
- Peng, R.; Jiang, B.; Ma, J.; Ma, Z.; Wan, X.; Liu, H.; Chen, Z.; Cheng, Q.; Chen, R. Forced downregulation of RACK1 inhibits glioma development by suppressing Src/Akt signaling activity. Oncol. Rep. 2013, 30, 2195–2202. [Google Scholar] [CrossRef]
- Pezuk, J.A.; Brassesco, M.S.; Morales, A.G.; de Oliveira, J.C.; de Paula Queiroz, R.G.; Machado, H.R.; Carlotti, C.G.; Neder, L.; Scrideli, C.A.; Tone, L.G. Polo-like kinase 1 inhibition causes decreased proliferation by cell cycle arrest, leading to cell death in glioblastoma. Cancer Gene Ther. 2013, 20, 499–506. [Google Scholar] [CrossRef]
- Premkumar, D.R.; Jane, E.P.; Pollack, I.F. Co-administration of NVP-AEW541 and dasatinib induces mitochondrial-mediated apoptosis through Bax activation in malignant human glioma cell lines. Int. J. Oncol. 2010, 37, 633–643. [Google Scholar]
- Preukschas, M.; Hagel, C.; Schulte, A.; Weber, K.; Lamszus, K.; Sievert, H.; Pällmann, N.; Bokemeyer, C.; Hauber, J.; Braig, M.; et al. Expression of eukaryotic initiation factor 5A and hypusine forming enzymes in glioblastoma patient samples: Implications for new targeted therapies. PLoS ONE 2012, 7, e43468. [Google Scholar] [CrossRef]
- Punganuru, S.R.; Arutla, V.; Zhao, W.; Rajaei, M.; Deokar, H.; Zhang, R.; Buolamwini, J.K.; Srivenugopal, K.S.; Wang, W. Targeted brain tumor therapy by inhibiting the MDM2 oncogene: In vitro and in vivo antitumor activity and mechanism of action. Cells 2020, 9, 1592. [Google Scholar] [CrossRef] [PubMed]
- Raub, T.J.; Wishart, G.N.; Kulanthaivel, P.; Staton, B.A.; Ajamie, R.T.; Sawada, G.A.; Gelbert, L.M.; Shannon, H.E.; Sanchez-Martinez, C.; De Dios, A. Brain Exposure of Two Selective Dual CDK4 and CDK6 Inhibitors and the Antitumor Activity of CDK4 and CDK6 Inhibition in Combination with Temozolomide in an Intracranial Glioblastoma Xenograft. Drug Metab. Dispos. 2015, 43, 1360–1371. [Google Scholar] [CrossRef] [PubMed]
- Reardon, D.A.; Brandes, A.A.; Omuro, A.; Mulholland, P.; Lim, M.; Wick, A.; Baehring, J.; Ahluwalia, M.S.; Roth, P.; Bähr, O.; et al. Effect of Nivolumab vs Bevacizumab in Patients With Recurrent Glioblastoma: The CheckMate 143 Phase 3 Randomized Clinical Trial. JAMA Oncol. 2020, 6, 1003–1010. [Google Scholar] [CrossRef]
- Renfrow, J.J.; Soike, M.H.; West, J.L.; Ramkissoon, S.H.; Metheny-Barlow, L.; Mott, R.T.; Kittel, C.A.; D’Agostino, R.B., Jr.; Tatter, S.B.; Laxton, A.W. Attenuating hypoxia driven malignant behavior in glioblastoma with a novel hypoxia-inducible factor 2 alpha inhibitor. Sci. Rep. 2020, 10, 15195. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Iizuka, Y.; Ohta, S.; Takahashi, S.; Nakamura, K.; Saya, H.; Yoshida, K.; Kawakami, Y.; Toda, M. Functional analysis of a novel glioma antigen, EFTUD1. Neuro-Oncology 2014, 16, 1618–1629. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Bringas, J.R.; Panner, A.; Tamas, M.; Pieper, R.O.; Berger, M.S.; Bankiewicz, K.S. Convection-enhanced delivery of tumor necrosis factor-related apoptosis-inducing ligand with systemic administration of temozolomide prolongs survival in an intracranial glioblastoma xenograft model. Cancer Res. 2004, 64, 6858–6862. [Google Scholar] [CrossRef]
- Salphati, L.; Heffron, T.P.; Alicke, B.; Nishimura, M.; Barck, K.; Carano, R.A.; Cheong, J.; Edgar, K.A.; Greve, J.; Kharbanda, S. Targeting the PI3K pathway in the brain—Efficacy of a PI3K inhibitor optimized to cross the blood–brain barrier. Clin. Cancer Res. 2012, 18, 6239–6248. [Google Scholar] [CrossRef]
- Sanai, N.; Li, J.; Boerner, J.; Stark, K.; Wu, J.; Kim, S.; Derogatis, A.; Mehta, S.; Dhruv, H.D.; Heilbrun, L.K. Phase 0 trial of AZD1775 in first-recurrence glioblastoma patients. Clin. Cancer Res. 2018, 24, 3820–3828. [Google Scholar] [CrossRef]
- Sanzey, M.; Abdul Rahim, S.A.; Oudin, A.; Dirkse, A.; Kaoma, T.; Vallar, L.; Herold-Mende, C.; Bjerkvig, R.; Golebiewska, A.; Niclou, S.P. Comprehensive analysis of glycolytic enzymes as therapeutic targets in the treatment of glioblastoma. PLoS ONE 2015, 10, e0123544. [Google Scholar] [CrossRef]
- Sathornsumetee, S.; Hjelmeland, A.B.; Keir, S.T.; McLendon, R.E.; Batt, D.; Ramsey, T.; Yusuff, N.; Rasheed, B.A.; Kieran, M.W.; Laforme, A. AAL881, a novel small molecule inhibitor of RAF and vascular endothelial growth factor receptor activities, blocks the growth of malignant glioma. Cancer Res. 2006, 66, 8722–8730. [Google Scholar] [CrossRef]
- Saunders, J.T.; Holmes, B.; Benavides-Serrato, A.; Kumar, S.; Nishimura, R.N.; Gera, J. Targeting the YAP-TEAD interaction interface for therapeutic intervention in glioblastoma. J. Neuro-Oncol. 2021, 152, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Sautter, L.; Hofheinz, R.; Tuettenberg, J.; Grimm, M.; Vajkoczy, P.; Groden, C.; Schmieder, K.; Hochhaus, A.; Wenz, F.; Giordano, F.A. Open-label phase II evaluation of imatinib in primary inoperable or incompletely resected and recurrent glioblastoma. Oncology 2020, 98, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Schleicher, S.M.; Thotala, D.K.; Linkous, A.G.; Hu, R.; Leahy, K.M.; Yazlovitskaya, E.M.; Hallahan, D.E. Autotaxin and LPA receptors represent potential molecular targets for the radiosensitization of murine glioma through effects on tumor vasculature. PLoS ONE 2011, 6, e22182. [Google Scholar] [CrossRef] [PubMed]
- See, W.L.; Tan, I.-L.; Mukherjee, J.; Nicolaides, T.; Pieper, R.O. Sensitivity of Glioblastomas to Clinically Available MEK Inhibitors Is Defined by Neurofibromin 1 Deficiency. Cancer Res. 2012, 72, 3350–3359. [Google Scholar] [CrossRef] [PubMed]
- Selvasaravanan, K.D.; Wiederspohn, N.; Hadzalic, A.; Strobel, H.; Payer, C.; Schuster, A.; Karpel-Massler, G.; Siegelin, M.D.; Halatsch, M.-E.; Debatin, K.-M. The limitations of targeting MEK signalling in Glioblastoma therapy. Sci. Rep. 2020, 10, 7401. [Google Scholar] [CrossRef] [PubMed]
- Shingu, T.; Holmes, L.; Henry, V.; Wang, Q.; Latha, K.; Gururaj, A.E.; Gibson, L.A.; Doucette, T.; Lang, F.F.; Rao, G. Suppression of RAF/MEK or PI3K synergizes cytotoxicity of receptor tyrosine kinase inhibitors in glioma tumor-initiating cells. J. Transl. Med. 2016, 14, 46. [Google Scholar] [CrossRef]
- Shulepko, M.; Bychkov, M.; Lyukmanova, E.; Kirpichnikov, M. Recombinant analogue of the human protein SLURP-1 inhibits the growth of U251 MG and A172 glioma cells. Dokl. Biochem. Biophys. 2020, 493, 211–214. [Google Scholar] [CrossRef]
- Signore, M.; Pelacchi, F.; Di Martino, S.; Runci, D.; Biffoni, M.; Giannetti, S.; Morgante, L.; De Majo, M.; Petricoin, E.; Stancato, L.; et al. Combined PDK1 and CHK1 inhibition is required to kill glioblastoma stem-like cells in vitro and in vivo. Cell Death Dis. 2014, 5, e1223. [Google Scholar] [CrossRef]
- Takano, S.; Tsuboi, K.; Matsumura, A.; Nose, T. Anti-vascular endothelial growth factor antibody and nimustine as combined therapy: Effects on tumor growth and angiogenesis in human glioblastoma xenografts. Neuro-Oncology 2003, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tchoghandjian, A.; Soubéran, A.; Tabouret, E.; Colin, C.; Denicolaï, E.; Jiguet-Jiglaire, C.; El-Battari, A.; Villard, C.; Baeza-Kallee, N.; Figarella-Branger, D. Inhibitor of apoptosis protein expression in glioblastomas and their in vitro and in vivo targeting by SMAC mimetic GDC-0152. Cell Death Dis. 2016, 7, e2325. [Google Scholar] [CrossRef]
- Thanasupawat, T.; Glogowska, A.; Burg, M.; Krcek, J.; Beiko, J.; Pitz, M.; Zhang, G.J.; Hombach-Klonisch, S.; Klonisch, T. C1q/TNF-related peptide 8 (CTRP 8) promotes temozolomide resistance in human glioblastoma. Mol. Oncol. 2018, 12, 1464–1479. [Google Scholar] [CrossRef] [PubMed]
- Tsigelny, I.F.; Mukthavaram, R.; Kouznetsova, V.L.; Chao, Y.; Babic, I.; Nurmemmedov, E.; Pastorino, S.; Jiang, P.; Calligaris, D.; Agar, N. Multiple spatially related pharmacophores define small molecule inhibitors of OLIG2 in glioblastoma. Oncotarget 2017, 8, 22370. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Niu, M.; Xie, P.; Yue, C.; Liu, N.; Qi, Z.; Gao, S.; Liu, H.; Shi, Q.; Yu, R. Smoothened is a poor prognosis factor and a potential therapeutic target in glioma. Sci. Rep. 2017, 7, 42630. [Google Scholar] [CrossRef]
- Venere, M.; Horbinski, C.; Crish, J.F.; Jin, X.; Vasanji, A.; Major, J.; Burrows, A.C.; Chang, C.; Prokop, J.; Wu, Q. The mitotic kinesin KIF11 is a driver of invasion, proliferation, and self-renewal in glioblastoma. Sci. Transl. Med. 2015, 7, 304ra143. [Google Scholar] [CrossRef]
- Vengoji, R.; Macha, M.A.; Nimmakayala, R.K.; Rachagani, S.; Siddiqui, J.A.; Mallya, K.; Gorantla, S.; Jain, M.; Ponnusamy, M.P.; Batra, S.K.; et al. Afatinib and Temozolomide combination inhibits tumorigenesis by targeting EGFRvIII-cMet signaling in glioblastoma cells. J. Exp. Clin. Cancer Res. 2019, 38, 266. [Google Scholar] [CrossRef]
- von Spreckelsen, N.; Fadzen, C.M.; Hartrampf, N.; Ghotmi, Y.; Wolfe, J.M.; Dubey, S.; Yang, B.Y.; Kijewski, M.F.; Wang, S.; Farquhar, C. Targeting glioblastoma using a novel peptide specific to a deglycosylated isoform of brevican. Adv. Ther. 2021, 4, 2000244. [Google Scholar] [CrossRef] [PubMed]
- Vredenburgh, J.J.; Desjardins, A.; Kirkpatrick, J.P.; Reardon, D.A.; Peters, K.B.; Herndon, J.E., II; Marcello, J.; Bailey, L.; Threatt, S.; Sampson, J.; et al. Addition of Bevacizumab to Standard Radiation Therapy and Daily Temozolomide Is Associated With Minimal Toxicity in Newly Diagnosed Glioblastoma Multiforme. Int. J. Radiat. Oncol. Biol. Phys. 2012, 82, 58–66. [Google Scholar] [CrossRef]
- Wang, F.; Bai, H.; Wang, J.; Bai, Y.; Dou, C. Glioma growth inhibition in vitro and in vivo by single chain variable fragments of the transferrin receptor conjugated to survivin small interfering RNA. J. Int. Med. Res. 2011, 39, 1701–1712. [Google Scholar] [CrossRef]
- Wang, F.; Huang, Q.; Zhou, L.-Y. Analysis of the treatment of gliomas with SEC therapy combined with radiochemotherapy. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 2400–2405. [Google Scholar]
- Wang, X.; Hua, Y.; Xu, G.; Deng, S.; Yang, D.; Gao, X. Targeting eZh2 for glioma therapy with a novel nanoparticle–sirNa complex. Int. J. Nanomed. 2019, 14, 2637–2653. [Google Scholar] [CrossRef]
- Wang, X.; Yang, K.; Wu, Q.; Kim, L.J.; Morton, A.R.; Gimple, R.C.; Prager, B.C.; Shi, Y.; Zhou, W.; Bhargava, S. Targeting pyrimidine synthesis accentuates molecular therapy response in glioblastoma stem cells. Sci. Transl. Med. 2019, 11, eaau4972. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pan, L.; Sheng, X.F.; Chen, S.; Dai, J.Z. Nimotuzumab, a humanized monoclonal antibody specific for the EGFR, in combination with temozolomide and radiation therapy for newly diagnosed glioblastoma multiforme: First results in Chinese patients. Asia Pac. J. Clin. Oncol. 2016, 12, e23–e29. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Nishijima, N.; Hirai, K.; Shibata, K.; Hase, A.; Yamanaka, T.; Inazu, M. Anticancer activity of Amb4269951, a choline transporter-like protein 1 inhibitor, in human glioma cells. Pharmaceuticals 2020, 13, 104. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; van den Bent, M.; Preusser, M.; Le Rhun, E.; Tonn, J.C.; Minniti, G.; Bendszus, M.; Balana, C.; Chinot, O.; Dirven, L. EANO guidelines on the diagnosis and treatment of diffuse gliomas of adulthood. Nat. Rev. Clin. Oncol. 2021, 18, 170–186. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, H.; Güttler, A.; Bache, M.; Taubert, H.; Rot, S.; Kessler, J.; Eckert, A.W.; Kappler, M.; Vordermark, D. Targeting of EGFR and HER2 with therapeutic antibodies and siRNA. Strahlenther. Onkol. 2015, 191, 180. [Google Scholar] [CrossRef] [PubMed]
- Wick, W.; Dettmer, S.; Berberich, A.; Kessler, T.; Karapanagiotou-Schenkel, I.; Wick, A.; Winkler, F.; Pfaff, E.; Brors, B.; Debus, J. N2M2 (NOA-20) phase I/II trial of molecularly matched targeted therapies plus radiotherapy in patients with newly diagnosed non-MGMT hypermethylated glioblastoma. Neuro-Oncology 2019, 21, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Gong, M.; Zou, Y.; Wang, Z.; Wu, B.; Zhang, S.; Li, L.; Jin, K.; Sun, C. Apatinib induces ferroptosis of glioma cells through modulation of the VEGFR2/Nrf2 pathway. Oxidative Med. Cell. Longev. 2022, 2022, 9925919. [Google Scholar] [CrossRef]
- Xiong, D.D.; Xu, W.Q.; He, R.Q.; Dang, Y.W.; Chen, G.; Luo, D.Z. In silico analysis identified miRNA-based therapeutic agents against glioblastoma multiforme. Oncol. Rep. 2019, 41, 2194–2208. [Google Scholar] [CrossRef]
- Xu, B.; Jiang, C.; Han, H.; Liu, H.; Tang, M.; Liu, L.; Ji, W.; Lu, X.; Yang, X.; Zhang, Y. Icaritin inhibits the invasion and epithelial-to-mesenchymal transition of glioblastoma cells by targeting EMMPRIN via PTEN/AKt/HIF-1α signalling. Clin. Exp. Pharmacol. Physiol. 2015, 42, 1296–1307. [Google Scholar] [CrossRef]
- Xu, T.-J.; Qiu, P.; Zhang, Y.-B.; Yu, S.-Y.; Xu, G.-M.; Yang, W. MiR-148a inhibits the proliferation and migration of glioblastoma by targeting ITGA9. Hum. Cell 2019, 32, 548–556. [Google Scholar] [CrossRef]
- Yan, D.; Kowal, J.; Akkari, L.; Schuhmacher, A.; Huse, J.; West, B.; Joyce, J. Inhibition of colony stimulating factor-1 receptor abrogates microenvironment-mediated therapeutic resistance in gliomas. Oncogene 2017, 36, 6049–6058. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Barth, R.F.; Wu, G.; Kawabata, S.; Sferra, T.J.; Bandyopadhyaya, A.K.; Tjarks, W.; Ferketich, A.K.; Moeschberger, M.L.; Binns, P.J. Molecular targeting and treatment of EGFRvIII-positive gliomas using boronated monoclonal antibody L8A4. Clin. Cancer Res. 2006, 12, 3792–3802. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wu, G.; Barth, R.F.; Swindall, M.R.; Bandyopadhyaya, A.K.; Tjarks, W.; Tordoff, K.; Moeschberger, M.; Sferra, T.J.; Binns, P.J.; et al. Molecular targeting and treatment of composite EGFR and EGFRvIII-positive gliomas using boronated monoclonal antibodies. Clin. Cancer Res. 2008, 14, 883–891. [Google Scholar] [CrossRef]
- Yao, T.-W.; Zhang, J.; Prados, M.; Weiss, W.A.; James, C.D.; Nicolaides, T. EGFR blockade prevents glioma escape from BRAFV600E targeted therapy. Oncotarget 2015, 6, 21993. [Google Scholar] [CrossRef]
- Zhang, C.; Burger, M.C.; Jennewein, L.; Genßler, S.; Schönfeld, K.; Zeiner, P.; Hattingen, E.; Harter, P.N.; Mittelbronn, M.; Tonn, T. ErbB2/HER2-specific NK cells for targeted therapy of glioblastoma. J. Natl. Cancer Inst. 2016, 108, djv375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yuan, X.-R.; Li, H.-Y.; Zhao, Z.-J.; Liao, Y.-W.; Wang, X.-Y.; Su, J.; Sang, S.-S.; Liu, Q. Anti-cancer effect of metabotropic glutamate receptor 1 inhibition in human glioma U87 cells: Involvement of PI3K/Akt/mTOR pathway. Cell. Physiol. Biochem. 2015, 35, 419–432. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Y.; Liu, X.Y.; Qin, Z.H.; Yang, J.M. Expression of elongation factor-2 kinase contributes to anoikis resistance and invasion of human glioma cells. Acta Pharmacol. Sin. 2011, 32, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; He, H.; Wang, C.; Tao, B.; Zhou, H.; Dong, Y.; Xiang, J.; Wang, L.; Luo, C.; Lu, Y. Downregulation of Id2 increases chemosensitivity of glioma. Tumor Biol. 2015, 36, 4189–4196. [Google Scholar] [CrossRef]
- Zustovich, F.; Landi, L.; Lombardi, G.; Porta, C.; Galli, L.; Fontana, A.; Amoroso, D.; Galli, C.; Andreuccetti, M.; Falcone, A. Sorafenib plus daily low-dose temozolomide for relapsed glioblastoma: A phase II study. Anticancer. Res. 2013, 33, 3487–3494. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zhang, X.; Gao, C.; Jiang, S.; Wu, H.; Liu, Z.; Dou, T. Burden and trends of brain and central nervous system cancer from 1990 to 2019 at the global, regional, and country levels. Arch. Public Health 2022, 80, 209. [Google Scholar] [CrossRef]
- Raja, N.; Hayes, L.; Basta, N.; McNally, R.J.Q. International trends in the incidence of brain tumours in children and young-adults and their association with indicators of economic development. Cancer Epidemiol. 2021, 74, 102006. [Google Scholar] [CrossRef] [PubMed]
- Grech, N.; Dalli, T.; Mizzi, S.; Meilak, L.; Calleja, N.; Zrinzo, A. Rising Incidence of Glioblastoma Multiforme in a Well-Defined Population. Cureus 2020, 12, e8195. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.R. Rising incidence of glioblastoma and meningioma in the United States: Projections through 2050. J. Clin. Oncol. 2012, 30, 2065. [Google Scholar] [CrossRef]
- Tebha, S.S.; Ali Memon, S.; Mehmood, Q.; Mukherjee, D.; Abdi, H.; Negida, A. Glioblastoma management in low and middle-income countries; existing challenges and policy recommendations. Brain Spine 2023, 3, 101775. [Google Scholar] [CrossRef]
- Cagney, D.N.; Alexander, B.M. The cost and value of glioblastoma therapy. Expert Rev. Anticancer. Ther. 2017, 17, 657–659. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.S.; Koffie, R.M.; Rattani, A.; Dewan, M.C.; Baticulon, R.E.; Qureshi, M.M.; Wahjoepramono, E.J.; Rosseau, G.; Park, K.; Nahed, B.V. Global incidence of brain and spinal tumors by geographic region and income level based on cancer registry data. J. Clin. Neurosci. 2019, 66, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Anjum, K.; Shagufta, B.I.; Abbas, S.Q.; Patel, S.; Khan, I.; Shah, S.A.A.; Akhter, N.; ul Hassan, S.S. Current status and future therapeutic perspectives of glioblastoma multiforme (GBM) therapy: A review. Biomed. Pharmacother. 2017, 92, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.; Hashem, S.A.; Al-Rashdan, A.; Ezam, N.; Nour, A.; Alsharbaji, A.; Sughayer, M.; Mohamad, I.; Elyan, M.; Addas, A.; et al. The challenges of managing glioblastoma multiforme in developing countries: A trade-off between cost and quality of care. Hematol. Oncol. Stem Cell Ther. 2011, 4, 116–120. [Google Scholar] [CrossRef]
- Wang, T.; Pham, A.; Yoo, S.; Attenello, F.J.; Jennelle, R.; Wagle, N.; Chang, E.L.; Zada, G. Identifying Disparities in Care in Treating Glioblastoma: A Retrospective Cohort Study of Patients Treated at a Safety-net Versus Private Hospital Setting. World Neurosurg. 2020, 137, e213–e220. [Google Scholar] [CrossRef]
- Ooi, S.Z.Y.; de Koning, R.; Egiz, A.; Dalle, D.U.; Denou, M.; Tsopmene, M.R.D.; Khan, M.; Takoukam, R.; Kotecha, J.; Sichimba, D.; et al. Management and outcomes of low-grade gliomas in Africa: A scoping review. Ann. Med. Surg. 2022, 74, 103246. [Google Scholar] [CrossRef]
- Balogun, J.A.; Adekanmbi, A.A. Management of glioblastoma: A perspective from Nigeria. Chin. Clin. Oncol. 2020, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Kanmounye, U.S.; Karekezi, C.; Nyalundja, A.D.; Awad, A.K.; Laeke, T.; Balogun, J.A. Adult brain tumors in Sub-Saharan Africa: A scoping review. Neuro-Oncology 2022, 24, 1799–1806. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; Steinbach, J.P.; Wick, W. Temozolomide: A milestone in the pharmacotherapy of brain tumors. Future Oncol. 2005, 1, 747–754. [Google Scholar] [CrossRef]
- Trinh, V.A.; Patel, S.P.; Hwu, W.J. The safety of temozolomide in the treatment of malignancies. Expert Opin. Drug Saf. 2009, 8, 493–499. [Google Scholar] [CrossRef]
- Singh, N.; Miner, A.; Hennis, L.; Mittal, S. Mechanisms of temozolomide resistance in glioblastoma—A comprehensive review. Cancer Drug Resist. 2021, 4, 17–43. [Google Scholar] [CrossRef]
- Ghosh, D.; Nandi, S.; Bhattacharjee, S. Combination therapy to checkmate Glioblastoma: Clinical challenges and advances. Clin. Transl. Med. 2018, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Qiu, W.; Sun, T.; Wang, L.; Du, C.; Hu, Y.; Liu, W.; Feng, F.; Chen, Y.; Sun, H. Therapeutic strategies of glioblastoma (GBM): The current advances in the molecular targets and bioactive small molecule compounds. Acta Pharm. Sin. B 2022, 12, 1781–1804. [Google Scholar] [CrossRef]
- Fisher, J.P.; Adamson, D.C. Current FDA-Approved Therapies for High-Grade Malignant Gliomas. Biomedicines 2021, 9, 324. [Google Scholar] [CrossRef]
- Zhu, P.; Du, X.L.; Lu, G.; Zhu, J.J. Survival benefit of glioblastoma patients after FDA approval of temozolomide concomitant with radiation and bevacizumab: A population-based study. Oncotarget 2017, 8, 44015–44031. [Google Scholar] [CrossRef]
- Yan, Y.; Xu, Z.; Dai, S.; Qian, L.; Sun, L.; Gong, Z. Targeting autophagy to sensitive glioma to temozolomide treatment. J. Exp. Clin. Cancer Res. 2016, 35, 23. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Huang, L.; Li, Y.; Fang, B.; Li, G.; Chen, L.; Xu, L. Mesenchymal Stem Cells and Cancer: Clinical Challenges and Opportunities. BioMed Res. Int. 2019, 2019, 2820853. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y. Temozolomide resistance in glioblastoma multiforme. Genes Dis. 2016, 3, 198–210. [Google Scholar] [CrossRef]
- Teraiya, M.; Perreault, H.; Chen, V.C. An overview of glioblastoma multiforme and temozolomide resistance: Can LC-MS-based proteomics reveal the fundamental mechanism of temozolomide resistance? Front. Oncol. 2023, 13, 1166207. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.-H.; Hsueh, W.-T.; Chuang, J.-Y.; Chang, K.-Y. Dissecting the mechanism of temozolomide resistance and its association with the regulatory roles of intracellular reactive oxygen species in glioblastoma. J. Biomed. Sci. 2021, 28, 18. [Google Scholar] [CrossRef] [PubMed]
- Eom, K.Y.; Cho, B.J.; Choi, E.J.; Kim, J.H.; Chie, E.K.; Wu, H.G.; Kim, I.H.; Paek, S.H.; Kim, J.S.; Kim, I.A. The Effect of Chemoradiotherapy with SRC Tyrosine Kinase Inhibitor, PP2 and Temozolomide on Malignant Glioma Cells In vitro and In vivo. Cancer Res. Treat. 2016, 48, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Gainer, J.L.; Sheehan, J.P.; Larner, J.M.; Jones, D.R. Trans sodium crocetinate with temozolomide and radiation therapy for glioblastoma multiforme. J. Neurosurg. 2017, 126, 460–466. [Google Scholar] [CrossRef]
- Lam, F.C.; Morton, S.W.; Wyckoff, J.; Vu Han, T.-L.; Hwang, M.K.; Maffa, A.; Balkanska-Sinclair, E.; Yaffe, M.B.; Floyd, S.R.; Hammond, P.T. Enhanced efficacy of combined temozolomide and bromodomain inhibitor therapy for gliomas using targeted nanoparticles. Nat. Commun. 2018, 9, 1991. [Google Scholar] [CrossRef]
- Ling, S.; Song, L.; Fan, N.; Feng, T.; Liu, L.; Yang, X.; Wang, M.; Li, Y.; Tian, Y.; Zhao, F.; et al. Combination of metformin and sorafenib suppresses proliferation and induces autophagy of hepatocellular carcinoma via targeting the mTOR pathway. Int. J. Oncol. 2017, 50, 297–309. [Google Scholar] [CrossRef]
- Hatanpaa, K.J.; Burma, S.; Zhao, D.; Habib, A.A. Epidermal growth factor receptor in glioma: Signal transduction, neuropathology, imaging, and radioresistance. Neoplasia 2010, 12, 675–684. [Google Scholar] [CrossRef]
- Minniti, G.; Muni, R.; Lanzetta, G.; Marchetti, P.; Enrici, R.M. Chemotherapy for Glioblastoma: Current Treatment and Future Perspectives for Cytotoxic and Targeted Agents. Anticancer. Res. 2009, 29, 5171. [Google Scholar]
- Veliz, I.; Loo, Y.; Castillo, O.; Karachaliou, N.; Nigro, O.; Rosell, R. Advances and challenges in the molecular biology and treatment of glioblastoma-is there any hope for the future? Ann. Transl. Med. 2015, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, S.K.; Brothers, S.P.; Wahlestedt, C. Emerging treatment strategies for glioblastoma multiforme. EMBO Mol. Med. 2014, 6, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Shergalis, A.; Bankhead, A., 3rd; Luesakul, U.; Muangsin, N.; Neamati, N. Current Challenges and Opportunities in Treating Glioblastoma. Pharmacol. Rev. 2018, 70, 412–445. [Google Scholar] [CrossRef]
- Tangsrivimol, J.A.; Schonfeld, E.; Zhang, M.; Veeravagu, A.; Smith, T.R.; Härtl, R.; Lawton, M.T.; El-Sherbini, A.H.; Prevedello, D.M.; Glicksberg, B.S.; et al. Artificial Intelligence in Neurosurgery: A State-of-the-Art Review from Past to Future. Diagnostics 2023, 13, 2429. [Google Scholar] [CrossRef] [PubMed]
- Ou, A.; Yung, W.K.A.; Majd, N. Molecular Mechanisms of Treatment Resistance in Glioblastoma. Int. J. Mol. Sci. 2021, 22, 351. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Abbreviation | Explanation |
|---|---|---|
| Protein kinase pathway group | PKP | Mechanisms related to protein kinases. |
| Cell cycle-related mechanisms | CCRM | Mechanisms associated with cell cycle, apoptosis, and transcription pathways. |
| Microenvironmental mechanisms | MT | Mechanisms in the tumor’s surrounding environment, including angiogenesis, cell–cell adhesion, and iron/cation regulation. |
| Immunomodulatory targets | IT | Targets that modulate the immune response. |
| Other targets | OT | Targets not falling into the previous categories. |
| Reference | Year | N | Male/Female Ratio | Years (Median and IQR) | Study Design | Molecular Mechanism | Molecular Target | Therapy | Success Rate/Outcome | Main Findings |
|---|---|---|---|---|---|---|---|---|---|---|
| Sanai et al. [141] | 2018 | 20 | 12/8 | 59 (28–81) | NRCT | PKP | Wee1K | AZD1775 | N/A | AZD1775 reaches therapeutic concentrations in GBM, well tolerated. |
| Wick et al. [169] | 2019 | 450 | N/A | N/A | NRCT (II) | PKP | ALK CDK4/6 mTOR MDM2 SHH | Alectinib Palbociclib Temsirolimus Idasanutlin Vismodegib | N/A | NCT Neuro Master Match (N2M2) trial uses GBM molecular signatures for treatment. |
| Sauter et al. [145] | 2022 | 51 | 36/15 | Primary: 63 (35–78); Recurrent: 52.5 (29–70) | NRCT (II) | OM | CSF1R, ABL, cKIT, PDGFR | Imatinib | mPFS 2.8 m in arm A and 2.1 m in arm B. mOS was 5.0 (0.8–30) m in arm A and 6.5 m in arm B. | Imatinib shows no significant effect on GBM. |
| Wang et al. [165] | 2014 | 92 | 21/5 | 50 (18–76) | Pilot clinical study | MT | EGFR | Nimotuzumab + TMZ + RT | mOS 15.9 m; mPFS 10 m. | Nimotuzumab, TMZ, and RT offer similar survival times. |
| Hasselbalch et al. [74] | 2010 | 37 | 21/16 | 57.9 (23.8–70.3) | Prospective study | MT | EGFR, VEGF, topoisomerase I | Cetuximab + bevacizumab + irinotecan | mPFS in CBI (n = 37) corresponded to 17 w. | No biomarkers identify bevacizumab benefits. |
| Mason et al. [116] | 2012 | 32 | 22/10 | 53 (43–71) | RCT (I) | PKP | mTOR1 | Everolimus + TMZ | N/A | Everolimus (5 days) + TMZ is an appropriate phase II dose. |
| Chinnaiyan et al. [48] | 2013 | 25 | 14/11 | 57 (31–73) | RCT (I) | PKP | mTOR | Everolimus + TMZ + RT | N/A | Everolimus (10 mg) + RT/TMZ: tolerable, acceptable toxicity. |
| Lassen et al. [99] | 2015 | 22 | 32/12 | 60 (37–72) | RCT (I) | MT | Placental growth factor (PlGF) + VEGF | RO5323441 + Bevacizumab | mPFS 3.5 m, mOS 8.5 m. | RO5323441 plus bevacizumab toxicity is manageable. |
| Desjardins et al. [54] | 2012 | 32 | 19/13 | 56 (25–80) | RCT (II) | MT | VEGF | Bevacizumab | MPFS was 15.8 w. | Combined temozolomide and bevacizumab show activity and tolerance. |
| Vredenburgh et al. [160] | 2012 | 125 | 74/51 | 56.2 (19–80) | RCT (II) | MT | VEGF | Bevacizumab + RT + TMZ | Medaian PFS was 13.8 m. | Bevacizumab addition to TMZ and RT has minimal toxicity. |
| Zustovich et al. [182] | 2013 | 43 | 18/25 | 60 (36.1–77.0) | RCT (II) | PKP | Multitarget kinase | Sorafenib | Median time-to-progression was 3.2 m. | Sorafenib + TMZ is safe with activity in relapsed GBM. |
| Clarke et al. [50] | 2014 | 59 | N/A | 90 (60–100) | RCT (II) | MT | VEGF + tyrosine kinase | Bevacizumab + Erlotinib | OS: 19.8 m, PFS: 13.5 m. | Bevacizumab/erlotinib/TMZ/radiotherapy improves progression-free survival. |
| Brown et al. [37] | 2016 | 38 | 27/11 | 57.0 (30–71) | RCT (II) | MT | VEGFR + EGFR | Cediranib + Gefitinib/placebo | PFS (cediranib + gefitinib): 3.6 m, PFS (cediranib + placebo): 2.8 m. | Cediranib and gefitinib combination improves PFS. |
| Badruddoja et al. [29] | 2017 | 30 | 19/11 | 55 (18–82) | RCT (II) | MT | VEGF MGMT | Bevacizumab + TMZ | Overall response rate from diagnosis was 51 w, the PFS-6 was 52%, and median time to tumor progression was 5.5 m. | Bevacizumab + temozolomide is a salvage regimen for recurrent GBM. |
| Lombardi et al. [109] | 2019 | 119 | 84/35 | 54·8 (46·8–61·3) | RCT (II) | PKP | Multitarget kinase + mTOR | regorafenib | Survival 24.8 m with regorafenib vs. 6.2 m for patients with progressive disease. | REGOMA showed survival benefits with regorafenib in recurrent GBM. |
| Nayak et al. [124] | 2021 | 80 | 54/26 | 53 (42–60) | RCT (II) | IT | PD1 + VEGF | Pembrolizumab + Bevacizumab | PFS-6: 26.0% and OS 8.8 m with bevacizumab. PFS-6 was 6.7%, mOS was 10.3 m w/o bevacizumab. | Pembrolizumab +/− bevacizumab is not effective in therapy. |
| Weller et al. [167] | 2017 | 745 | N/A | N/A | RCT (III) | MT | EGFRvIII | TMZ +/− Rindopepimut | N/A | Rindopepimut monotherapy does not reduce GBM mortality. |
| Reardon et al. [136] | 2020 | 439 | 235/204 | 55.5 (22–77) | RCT (III) | IT | PD1 | Nivolumab | mOS (nivolumab): 9.8 m; bevacizumab, 10.0 m; mOS-12 42% in both groups. | Nivolumab monotherapy is as effective as bevacizumab. |
| D’Alessandris et al. [53] | 2013 | 10 | 19/7 | 52.5 (29–77) | RTC | MT | VEGF + EGFRvIII | Bevacizumab + Erlotinib | mPFS 8.0 m; mOS 9.5 m. | Molecular analysis improves RR and PFS at 6 months. |
| Butowski et al. [38] | 2010 | 12 | 8/4 | 11 patients > 18 and <65 y 1 patient > 65 y | RTC | PKP | Protein kinase C-beta + PI3K/Akt | Enzastaurin + TMZ | Median survival: 14.6 m, and 1/4 patients > 2 y. | Enzastaurin + RT/TMZ: Well tolerated. |
| Hashimoto et al. [73] | 2015 | 7 | 4/3 | 49 (41–60) | RTC (I) | OM | WT1 (Wilms Tumor 1) | WT1 peptide vaccination + TMZ | 4 patients remained in an NR status after GTR, two showed complete response. | Combined peptide vaccination with temozolomide is safe. |
| Desjardins et al. [55] | 2011 | 36 | 29/7 | 52 (26–74) | RTC (I) | OM | Farnesyl transferase | SCH 66336 | mOS (14.3 m); mPFS (4.5 m); PFS-6 (41.7%). | SCH 66336 dose specified for strata. |
| Geletneky et al. [65] | 2017 | 18 | 14/4 | 57.8 ± 10.6 | RTC (I/II) | OM | Protein NS1 | Rat H-1 parvovirus (H-1PV) | PFS-6: 27%; mPFS: 111 d. | H-1PV is safe with favorable PFS. |
| Kanemaru et al. [90] | 2019 | 1 | 1/0 | N/A | Case report | PKP | BRAF + MEK | Dabrafenib + Trametinib | N/A | Dabrafenib and trametinib + radiation showed strong response in epithelioid GBM. |
| Anghileri et al. [26] | 2021 | 1 | 0/1 | N/A | Case report | IT | PD1 | Nivolumab | N/A | Nivolumab is useful for GBM patients. |
| Johanns et al. [85] | 2018 | 2 | 1/1 | N/A | Case series | PKP | BRAF + MEK | Dabrafenib + Trametinib | N/A | PT1: 11mo therapy improved function, then progressed. PT2: 3 mo therapy allowed ambulation, but ended fatally. |
| Reference | Year | Species/Culture Type | Molecular Mechanism | Molecular Target | Therapy | Success Rate/Outcome | Main Study Findings |
|---|---|---|---|---|---|---|---|
| Blank et al. [36] | 2001 | HCC (endothelial YPEN-1 (CRL-222), microglial cell line N9, rat GBM cell line C6) | MT | O-acetyl GD2 ganglioside | Anti-GD2 Antibody | N/A | O-acetyl GD2 ganglioside prevents glioma proliferation. |
| Koul et al. [95] | 2005 | ACC (U87, U251, LN229, SNB-19, U373, and D54 human GBM cell lines) | PKP | Integrin-linked kinase | QLT0276 In DMSO | ILK inhibition decreased the in vitro invasive capability of glioma cells, concomitant with a decrease in MMP-2 secretion. | ILK inhibition down-regulates proliferation and invasion. |
| Paternot et al. [128] | 2009 | HCC (T98G, U-87 MG, and U-138 MG) | PKP | mTOR1 + MEK1/2 | Rapamycin + PD184352 | Complete inhibition of DNA synthesis and pRb phosphorylation requires the combined inhibition of MEK1/2 and mTOR-raptor pathways. | Combined mTOR1 and MEK1/2 inhibition in CDK4-dysregulated tumors. |
| Premkumar et al. [151] | 2010 | HCC (U87, T98G, U373, LN229 and A172) | PKP | IGF1R + Src | NVP-AEW541 + Dasatinib | The effect on the induced formation of Bax homodimers (42 kDa), homotrimers (63 kDa), and homotetramers (84 kDa) was significantly reduced by transfection with Bcl-2 and Myr-Akt. | Dual IGF1R and Src inhibition increases apoptosis in glioma cells. |
| Siegelin et al. [132] | 2010 | HCC | PKP | BRAF | Sorafenib | N/A | Sorafenib has potent in vivo and in vitro anti-glioma activity. |
| Cloninger et al. [51] | 2011 | HCC (U87 and LN229 parental lines) | PKP | SAPK2/p38 + mTORC1 | SB203580 + Rapamycin | Significant inhibition of tumor growth rate 76% (at end of dosing period) and tumor growth delay, 16.5 days. | SAPK2/p38 + mTORC1 inhibitors for synergistic response. |
| Liu et al. [106] | 2011 | HCC (U251) | PKP | bFGF | Anti-bFGF siRNA | Cytochrome C, Caspase3, and Bax were markedly higher in the Ad-bFGF-siRNA group than in the control group. | bFGF siRNA is a potential glioma treatment. |
| Zhang et al. [180] | 2011 | HCC (T98G and LN-229) | CCRM | ID2 | Anti ID2 siRNA | The viability of cultured glioma cells was reduced in eEF-2 kinase knock-down when compared with control cells. | ID2 upregulation decreases glioma apoptosis; targeting increases sensitivity. |
| Ishiwata et al. [80] | 2011 | HCC and ACC/ A172 | OM | hnRNP A1/B2 | Β-Asarone | The growth rate and motility of Nes cells were higher than those of the mock cells | β-Asarone inhibits EMT and invasion. |
| Du et al. [58] | 2012 | HCC (BT325 and U251) | PKP | Raf/MEK/ERK signaling pathway | Sorafenib + Vitamin K (VK1) | The combination of low-concentration sorafenib (2.5 μM) and VK1 (50 μM) exhibited strong synergistic action by inhibiting protein expression of Bcl-2 and Mcl-1, leading to induction of cell apoptosis. | Sorafenib + VK1 induces apoptosis through protein regulation. |
| Lee et al. [101] | 2012 | HCC | CCRM | Wee1K | Mk-1775 | The median survival time of the patients under 50 years old is 34 months, almost three times longer than the 12-month median survival time of the patients > 50. | Wee1K phosphorylation is an effective anti-tumor target. |
| Golubovskaya et al. [30] | 2013 | HCC | PKP | FAK | Y15 | N/A | FAK autophosphorylation blockade with Y15 is a potential GBM therapy. |
| Jin et al. [83] | 2013 | HCC (U251, U87) | PKP | Akt + NOTCH | MRK003 + MK-2206 | Combination treatment was superior to monotherapy in both U251 and U87 cells. | Akt and NOTCH inhibition decreases glioma proliferation. |
| Pezuk et al. [131] | 2013 | HCC (U251, U138, U87, T98G, U343, MO59K, LN319, SF188) | PKP | PLK1 | Bi2536 + Tmz | PLK1 possible therapeutic target; BI 2536 inhibited tumor growth in vivo. | PLK1 inhibition + TMZ are effective in vitro. |
| Kaneta et al. [91] | 2013 | HCC (U1242) | CCRM | BMI-1 | Ptc-209 | The inhibition of NEK9 suggested as a novel anticancer therapeutic strategy. | Tumor growth is attenuated by PTC-2009; potential BMI-1 inhibitor. |
| Lian et al. [104] | 2013 | HCC | CCRM | EGFR | AZD9291 | miR-23a might be employed as a novel prognostic marker and a therapeutic target for glioma. | AZD9291 is efficient in GBM preclinical models. |
| Mao et al. [115] | 2013 | HCC (U87, SF268, A172 and U118) | CCRM | MDM2/4 + α5β1/αvβ3 | Compound 9 | Targeting STK17A may lead to the development of new therapies for GBM and sensitize cancers to existing therapies. | Compound 9 inhibits p53, shows anti-glioma potential. |
| Ji et al. [82] | 2013 | HCC | MT | VEGFR | Axitinib | High expression levels of Nrf2 and HIF-1alpha correlated with low 1-year survival rate. median OS 13 mo. | Axitinib exhibits antiangiogenic activity and prolongs survival. |
| Aldea et al. [24] | 2014 | HCC | PKP | mTOR + RAF | Metformin + Sorafenib | Metformin + sorafenib could be combined into an efficient in vitro treatment strategy and this association is superior to either drug used alone or when compared with the use of TMZ. | Metformin + sorafenib is effective for TMZ-resistant GBM cells. |
| Emlet et al. [59] | 2014 | HCC (U87) | PKP | EGFRvIII + CD133 | Egfrviii + CD133 AB | The specific lysis of the EGFRvIII+/CD133+ population significantly reduces the implantation of primary GBM tumors in mice and prolongs survival. | EGFRvIII + CD133 BsAb target cancer stem cells. |
| Hong et al. [77] | 2014 | HCC | PKP | Aurora-A kinase | Alisertib | CE7 epitope was highly detected in GBM and it represents a potential therapeutic target. | Inhibiting Aurora-A kinase enhances radiation effects. |
| Jung et al. [89] | 2014 | HCC | PKP | FOXO3A | Z-Ajoene | Z-ajoene is a potential candidate for the treatment of GBM. | Z-ajoene targets glioma CSCs via FOXO3A pathway. |
| Liu et al. [105] | 2014 | HCC (CE7R+ T cells) | PKP | EGFR and PI3K/Akt | G19 | G19 inhibited cell proliferation of U-87 MG human glioma cells in vitro and in vivo | G19 targets EGFR and PI3K/Akt, inducing redox stress. |
| Liu et al. [108] | 2014 | HCC (T98G, A172, and U87) | PKP | AMPK | Compound C | Compound C at 10 μM inhibited proliferation and glioma formation of human U87MG glioma cells in vivo. | Compound C is a potent anti-glioma agent. |
| Camorani et al. [40] | 2015 | HCC (U87MG) | PKP | EGFRvIII | CL4 Aptamer + EGFR Tkis | Combined treatment with CL4 and Gint4.T aptamers led to a consistently higher inhibition of cell growth. | CL4 and gefitinib cooperate with anti-PDGFRβ aptamer. |
| Ma et al. [114] | 2015 | HCC (U251 and U87) | PKP | STAT3 | Tetrandrine | Higher expressions of STAT3 in patients with glioma received lower survival rates. | Tetrandrine inhibits glioma growth without affecting embryos. |
| Wichmann et al. [168] | 2015 | HCC (U251 and LN-229) | PKP | EGFR and HER2 | siRNA + Cetuximab + Trastuzumab | Knock-down of HER2 reduces clonogenic survival in both GBM cell lines. | EGFR and HER2 siRNA reduce GBM growth rate. |
| Zhao et al. [181] | 2015 | HCC (U87) | CCRM | CDK + Aurora (dual inhibitor) | Jnj-7706621 | Id2 is a good molecular target for GBM gene therapy. | JNJ-7706621 shows potential for GBM treatment. |
| Xu et al. [172] | 2015 | HCC (U87MG) | IT | CXCR4 | POL5551 + MCR89 | Icaritin is a promising anti-cancer agent in the treatment of GBM. | Higher POL5551 concentrations improve survival, especially with VEGF antagonism. |
| Junca et al. [88] | 2017 | HCC | PKP | ALK, ROS1, MET | Crizotinib | Overexpression was associated with poor prognosis with a survival of 11.7 months against 14.3 months for patients whose tumors did not express or had low expression of MET. | MET and ALK overexpression in glioma; crizotinib potential. |
| Thanasupawat et al. [154] | 2017 | HCC (U87MG) | PKP | FGFR | Dovitinib | N/A | Alternation of dovitinib and TMZ reduces GBM viability. |
| Caruana et al. [41] | 2017 | HCC (T98G) | OM | APLNR | MM54 Or MM193 (APLNR Antagonists) | N/A | APLNR inhibition significantly reduces tumor growth. |
| Barbarisi et al. [32] | 2018 | HCC | PKP | CD44 | Quercetin + TMZ | N/A | CD44-targeted nanocarriers deliver quercetin to GBM. |
| Merlino et al. [119] | 2018 | HCC (U87MG) | CCRM | CDK 4/6 | PD-0332991 | N/A | PD-0332991 inhibits glioma growth, increases survival. |
| Franco et al. [63] | 2018 | HCC (U87MG) | MT | LTβR | Light-VTP | N/A | LIGHT-VTP prevents angiogenesis and promotes immune infiltration. |
| Pall et al. [127] | 2019 | HCC (hBMVECs, U251n and U87, RAW264.7) | MT | HIF2α | PT2385 | N/A | HIF2α is a reasonable therapeutic target; PT2385 is effective. |
| Xiong et al. [171] | 2019 | HCC (MCF7, HL60, MCF7) | MT | STING | ASA404 | N/A | ASA404 efficacy varies by administration method. |
| Peng et al. [129] | 2019 | HCC (U-373MG Uppsala, U-87MG Uppsala, U251 and T98G) | OM | EFTUD1 | EFTUD1 shRNA | N/A | EFTUD1 overexpression is associated with glioma. |
| Ariey-Bonnet et al. [28] | 2020 | HCC (U87, U87vIII, T98G, and U251) | PKP | MAPK14 | BMZ | N/A | BMZ inhibits MAPK14, with anticancer properties. |
| Bagca et al. [30] | 2020 | HCC (T98G) | PKP | ALK | AZD3463 + TMZ | N/A | Combo with AZD3463 may enhance TMZ in GBM. |
| Bychkov et al. [39] | 2020 | HCC (U251 MG and A172) | CCRM | S100A9 (one of the heterodimers for calprotectin) | shRNA | N/A | S100A9 knockdown demonstrates anticancer potential. |
| Cheng et al. [47] | 2022 | HCC (LN-229, T98, A172, and human astrocyte) | PKP | CTSC | Piperlongumine + Scopoletin | N/A | CTSC is a MAPK biomarker; piperlongumine and scopoletin inhibit growth. |
| Reference | Year | Species/Culture Type | Molecular Mechanism | Molecular Target | Therapy | Success Rate/Outcome | Main Study Findings |
|---|---|---|---|---|---|---|---|
| Takano et al. [152] | 2003 | A (mice) | MT | CTL1 (choline transporter-like protein 1) | AMB4269951 | MST: 52.8 ± 5.5 days (ACNU + VEGF therapy significantly mouse survival. | Amb4269951 has significant antitumor effects in glioma. |
| Saito et al. [139] | 2004 | A (rats) | OM | α7 nAChR | Rslurp-1 | Rats who received combination therapy survived more than 80 days and revealed fibrous scar tissue at necropsy. | rSLURP-1 demonstrates antitumor activity. |
| SathornSumetee et al. [143] | 2006 | A (mice) | PKP | BRAF, CRAF, VEGFR | AA1881 | Median life spans of 12 days for control mice and 44 days for AAL881-treated animals. | AAL881 inhibits glioma growth, well tolerated. |
| Yang et al. [175] | 2006 | A (rats) | CCRM | EF2-kinase | EF2-siRNA | MST (bioconjugate in combination with BPA) = 85.5 days compared with 70.4 days for those that received it alone, 40.1 days for BPA alone, and 30.3 days for irradiated controls. | EF2 regulates cell migration; knockdown inhibits these properties. |
| Yang et al. [176] | 2008 | A (rats) | PKP | EGFR | Boronated EGFR MAB + Cetuximab | The MST of animals that received both boronated mAbs was 55 days compared with MSTs of 36 and 38 days for animals that received either one or the other boronated mAb. | Both EGFR and EGFRvIII tumors must be targeted for glioma. |
| Feng et al. [61] | 2010 | A (Rats) | PKP | PI3K/Akt; JNK; ERK | Tamoxifen | Treatment with TAM at 20 μM caused about half of the C6 glioma cells to die after 24 h. | TAM-induced apoptosis reveals potential targets. |
| Koul et al. [96] | 2010 | A (mice) | PKP | PI3K/Akt | Px-866 | The median survival time for controls was 32 days; treated with PX-866 was significantly longer at 39 days. | PX-866 inhibits growth, induces G1 arrest in mice. |
| Colen et al. [52] | 2011 | HCC (U87-MG and U251-MG) | OM | MALAT1 | Nanocomplex Targeting MALAT1 + TMZ | A 50% survival rate was observed with the nude rat model with no tumor recurrence after treatment. | Combined TMZ with MALAT1 silencing offers a survival benefit. |
| Joshi et al. [86] | 2012 | HCC (serum-cultured and oncosphere lines) | PKP | Multitarget kinases | Gefitinib + Erlotinib + Sunitinib | Use of PAC-1 with TMZ substantially improved the median survival to 205 days compared to vehicle, compared to TMZ alone. | Sunitinib combinations are effective in vitro, not in vivo. |
| Li et al. [103] | 2012 | A (mice) | CCRM | miR-23a (APAF1) | Anti-mir-23a | The MST of intracranial U87 glioblastoma-bearing nude mice treated with RGD-liposomal pDP (29 days) was significantly longer than that of mice treated with blank RGD-liposome (23 days) (p < 0.001). | miR-23a upregulated in gliomas; knockdown reduces survivability. |
| Arcella et al. [27] | 2013 | A (mice) | PKP | mTOR | Rapamycin | Rapamycin-treated mice survive almost double that observed in vehicle-treated mice. | Rapamycin is a potent mTOR inhibitor for GBM. |
| Grossman et al. [69] | 2013 | A (rats) | MT | TRPV4 | Cannabidiol (CBD) | TMZ intratumoral concentrations do not decline in the setting of the oral tyrosine kinase inhibitor cediranib. | CBD induces lethal mitophagy; TRPV4 is a target. |
| Chen et al. [46] | 2013 | A (rats) | OM | Nestin | Anti-Nestin IGG | TMP shown to be a potential therapeutic candidate for the treatment of resistant malignant gliomas. | Nestin downregulation is associated with reduced glioma proliferation and migration. |
| Wang et al. [162] | 2014 | H | PKP | RAS | Mir-143 | ER 51.6% ER 63.6% ER 32.0% | miR-143 downregulated in glioma, inactivates RAS. |
| Shingu et al. [149] | 2015 | A (mice) | PKP | MEK, EGFR, PI3K | Various Small Molecule Inhibitors | Combination of erlotinib and sorafenib tended to improve survival of nude mice bearing GSC11 brain tumors. | Most synergistic drug combinations affect RTKs and MEK/ERK or PI3K. |
| Yao et al. [177] | 2015 | A (mice) | PKP | EGFR and BRAF | BRAF(V600E) Inhibitor PLX4720 | BRAFV600E + EGFR inhibitors showed dramatic reduction in tumor growth and extended survival compared to vehicle or single-drug-treated counterparts. | Inhibiting EGFR and BRAF(V600E) reduces proliferation. |
| Venere et al. [157] | 2015 | A (mice) | OM | IDH1R132H | Wm17 | median survival of 36 days versus 24 days for the DMSO vehicle cohort. | WM17 is a mutant IDH1 inhibitor. |
| Balkhi et al. [113] | 2016 | A (rat) | PKP | Multitarget kinases | Caffeic Acid Phenethyl Ester (CAPE) + Dasatinib | N/A | Combo therapy inhibits migration and invasiveness, and reduces survival. |
| He et al. [76] | 2016 | A (mice) | PKP | MEK2 | MEK2 Antibody | Si-MEK2-infected U87 cell glioma burden mice had longer survival times (48 d) compared with Si-ctl-infected glioma burden mice (30.4). | MEK2 antagonists sensitize TMZ treatment in GBM. |
| Ju et al. [87] | 2016 | A (mice) | PKP | COX-2 | Celecoxib | Physiological saline survival (days): range 13–25, Targeting epirubicin + celecoxib liposomes survival (days): 15–36. | Targeting epirubicin plus celecoxib liposomes effective in glioma. |
| Zhang et al. [178] | 2016 | A (mice) | PKP | HER2 | HER2-specific NK cells | Median survival of 200.5 days upon treatment with NK-92/5.28.z vs 73 days upon treatment with parental NK-92 cells. | Modified HER2-specific NK cells effective against GBM. |
| Grinshtein et al. [68] | 2016 | A (mice) | CCRM | BAG3 | BAG3 siRNA | N/A | BAG3 is highly expressed in gliomas; a therapeutic target. |
| Lescarbeau et al. [102] | 2016 | A (mice) | CCRM | p53/MDM2 | D-PMNIbeta | Mean survival time after treatment initiation was 22 days with MK-1775 treatment and only 13 days with control. | D-PMIBeta an effective p53 inhibitor. |
| Tchoghandjian et al. [153] | 2016 | A (mice) | CCRM | EGFR | Afatinib + TMZ | Treatment significantly increased mouse survival in a dose-dependent manner. | Afatinib + TMZ significantly delays progression. |
| Fleurence et al. [62] | 2016 | A (mice) | MT | Pan-VEGF | Cediranib + TMZ | N/A | Intratumoral TMZ concentrations are slightly increased. |
| Farrell et al. [60] | 2017 | A (mice) | PKP | MET | WO2010/019899A1 + PF04217903 + Crizotinib | N/A | Dual targeting of HGF and MET could be effective. |
| Yan et al. [174] | 2017 | A (mice) | PKP | CSF-1R + cKIT + RTKs | PLX3397 + Vatalanib + Dovitinib | N/A | PLX3397 is effective, improves TKI efficacy. |
| Joshi et al. [86] | 2017 | A (mice) | CCRM | Phospholipase C | D609 | Rats treated with PAC-1 showed significantly improved survival (59 d) was 350% longer than for the untreated control rats (17 d), and 40% of the animals survived until the end of the experiment (day 200). | Chronic D609 treatment leads to decreased Olig2 biomarker levels. |
| Abdul Rahim et al. [23] | 2017 | A (mice) | MT | Phosphatidylserine | SAPC-DOPS | ATG9A knockdown led to a significant increase in mouse survival (+12–18%, Chloroquine treatment (20 mg kg−1) significantly prolonged survival of P3 mice (+18.4%). | SAPc-DOPS targets GBM effectively. |
| Angara et al. [25] | 2017 | A (rat) | MT | Endothelial pigpen protein | Aptamer III.1 | N/A | Aptamer III.1 is a potential GBM treatment. |
| Bäehr et al. [31] | 2017 | A (mice) | IT | ATX + LPA receptors | siRNA | Median survival was 36 days for PBS and 40 days for ASA404. | ATX and LPA receptor downregulation enhances radio-sensitivity. |
| Harford-Wright et al. [72] | 2017 | A (mice) | OM | IDH1R132H | AGI-5198 (In Combo with HDACi) | N/A | AGI-5198 attenuates HDACi resistance. |
| Tu et al. [156] | 2017 | A (mice) | OM | 14-3-2003 | siRNA | N/A | 14-3-3 downregulation decreases glioma survival. |
| Ciesielski et al. [49] | 2018 | A (mice) | PKP | Src-kinase + tubulin polymerization inhibitory activity | Kx2-361 | N/A | Active against GL261 gliomas in mice. |
| Kong et al. [94] | 2018 | A (mice) | CCRM | OPN | shRNA | N/A | U87-MG sphere cells’ tumorigenic potential abrogated upon OPN silencing. |
| He et al. [75] | 2018 | A (mice) | MT | VEGF + Src Family kinases | Bevacizumab + Dasatinib | N/A | Dasatinib may block bevacizumab-induced invasion. |
| Nandhu et al. [123] | 2018 | A (mice) | MT | NHE9 | Gold NEPTT | Intravenously injected mAb428.2 reduced tumor volume and significantly improved survival in all the fibulin-3-expressing models, extending median survival by 28% (GBM09) to 64% (GBM34) in the GBM xenografts. However, mAb428.2 did not prolong the survival of mice carrying fibulin-3-negative COLO201 tumor cells. | Gold nanoparticle-enabled photothermal therapy (NEPTT) kills tumor cells. |
| Kim et al. [93] | 2018 | A (mice) | OM | LPAR1/3 | KI16425 | Survival was extended by scL-siMAL + TMZ (50% of the mice were surviving at day 38). | LPA signaling knockdown reduces tumor growth. |
| Loskutov et al. [110] | 2018 | A (mice) | OM | PRC2 + BET bromodomain proteins | JQ1 + I-BET | N/A | H3K27M mutation effects are reduced by inhibiting PRC2 and BET proteins. |
| Chen et al. [44] | 2019 | A (mice) | PKP | CD163 pathway (CK2, kinase) | TBB | N/A | TBB inhibits CK2, crucial for tumor growth. |
| Chen et al. [42] | 2019 | A (mice) | CCRM | HDAC/EZH2 | Compound 26/UNC1999 | IGFBP3 siRNA-treated mice showed better OS compared with siCtrl-treated mice, and the median survival of siCtrl-, siIBP3-1-, and siIBP3-2-treated mice was 25, 32, and 35 d. | HDAC and EZH2 inhibition shows synergistic effects. |
| Liu et al. [107] | 2019 | A (mice) | CCRM | STK17A | Anti-STK17A shRNA | N/A | STK17A indicates worse prognosis; knockdown reduces survivability. |
| Vengoji et al. [158] | 2019 | A (mice) | CCRM | Survivin | Survivin-siRNA/Transferrin Receptor Conjugate | N/A | Conjugate decreases survivin expression, increases survival. |
| Wang et al. [163] | 2019 | A (mice) | CCRM | Carbamoyl-phosphate synthetase (CAD) | Teriflunomide | N/A | Targeting pyrimidine synthesis may improve outcomes. |
| Xu et al. [173] | 2019 | A (mice) | MT | CD73 | Anti-CD73 | N/A | Combination therapy targets CD73; anti-CD73 testing suggested. |
| Luwor et al. [112] | 2019 | A (mice) | OM | eIF-5A, DHS, DOHH (both eIF-5A activators) | Gc7 | N/A | eIF5-A is a potential therapeutic target. |
| Selvasaravanan et al. [148] | 2020 | A (mice) | PKP | MEK or PI3K | Trametinib + Pictilisib | N/A | MEK inhibition is not superior to PI3K inhibition. |
| Punganuru et al. [134] | 2020 | A (mice) | CCRM | HSP90 | BIIB021 + 17-AAG (HSP90 Inhibitor) + BRAFi +/Or MEKi | N/A | HSP90 inhibitor overcomes limitations of BRAFV600E therapy. |
| Renfrow et al. [137] | 2020 | A (mice) | MT | VEGF | Anti-VEGF AB + Nimustine | PT2385 single-agent treatment did improve mOS compared to placebo; no difference in animal survival was seen in combination treatment with radiation (RTtemozolomide TMZPT2385). | Combining antiangiogenic therapy with chemotherapy is promising. |
| Watanabe et al. [166] | 2020 | A (mice) | MT | Calmodulin, EGFR, aromatase | W-13 + Gefitinib + Exemestane | N/A | Identified miRNA-based chemicals for therapy. |
| Goswami et al. [67] | 2020 | A (mice) | IT | EMMPRIN | Icaritin | Improvement in survival was noted in WT and CD73−/− mice treated with anti-PD-1 + anti-CTLA-4 compared to untreated controls. Following treatment of anti-PD-1 + anti-CTLA-4, CD73−/− mice had improved survival as compared to WT GBM tumor-bearing mice. | Icaritin targets EMMPRIN, inhibiting GBM cell invasion and EMT. |
| Shulepko et al. [150] | 2020 | H | OM | KIF11 | Ipinesib | N/A | KIF11 inhibition halts tumor growth. |
| Kawauchi et al. [92] | 2021 | A (mice) | PKP | ALK | Alectinib + Ceritinib | Ceritinib or alectinib significantly prolonged the survival of mice harboring intracerebral U87MG or GSC23 xenografts. | Second-gen ALK inhibitors are potent against GBM. |
| Maxwell et al. [118] | 2021 | A (mice) | PKP | mTOR1/2 + MEK | TAK228 + Trametinib | N/A | mTOR1/2 and MEK inhibitors induce proteomic changes. |
| Genoud et al. [66] | 2021 | A (mice) | CCRM | PAK5 | PAK5 shRNA | Significant increase in survival with BAL101553 + aCD40 (49 d), compared with BAL101553 monotherapy (42). | PAK5 overexpressed in glioma; its inhibition is promising. |
| Huang et al. [78] | 2021 | A (mice) | MT | Growth-Hormone Releasing Hormone | MIA-604 + MIA-690 | N/A | GHRH antagonists augment standard treatments. |
| Chen et al. [45] | 2021 | A (mice) | OM | EEF1A1 + RPL11 | Puromycin + Doxorubicin + Daunorubicin + Mitoxantrone | N/A | Database analysis identifies target genes and potential drugs for glioma treatment. |
| Saunders et al. [144] | 2021 | A (mice) | OM | Smoothened | Gdc-0449 | NSC682769 treated GFAP-EGFRvIII × GFAP-Cre+/Rictor mice had a marked increase in OS with more than 75% of mice surviving at 20 weeks at 20 mg/kg and 60% of mice surviving to 20 weeks receiving the lower 5 mg/kg regimen. | Smoothened is a prognostic biomarker. |
| von Spreckelsen et al. [159] | 2021 | A (mice) | OM | FTO | SPI1 Inhibitor DB2313 | N/A | FTO is a novel prognostic indicator. |
| Xia et al. [170] | 2022 | A (mice) | MT | ITGA9 | miR-148a | U251 + U87 cell survival overexpressing Nrf2 remarkably increased 24, 48, and 72 h after treatment with apatinib, in comparison with cells transfected with the empty vector. | miR-148a suppresses GBM malignancy via ITGA9 targeting. |
| Wang et al. [164] | 2022 | A (mice) | CCRM | BCL6 | RI-BPi | The combination of lapatinib and teriflunomide yielded the greatest efficacy in tumor control and OS. | BCL6 overexpression in glioma worsens prognosis; RI-BPI reduces tumor growth. |
| Joel et al. [84] | 2015 | N/A | PKP | PBK/TOPK | Hi-Topk-032 | PBK may serve as a potential therapeutic target in GBM tumors. | HITOPK-032 diminishes tumor growth. |
| Reference | Year | Species/Culture Type | Molecular Mechanism | Molecular Target | Therapy | Success Rate/Outcome | Main Study Findings |
|---|---|---|---|---|---|---|---|
| Kuan et al. [97] | 2010 | HCC (D54 MG, D247 MG, D392 MG, and D245 MG, T98G and U251 MG | MT | TfR (transferrin receptor) | T12 + B6 + T7 (Tfr-Targeting Peptides) | High levels of RNA remain a significant predictor of survival. | T7-modified liposomes penetrate the BBB effectively. |
| Guo et al. [71] | 2011 | HCC (U87MG) | CCRM | CDK 4/6 + PDGFRα | Lenvatinib + Crenolanib + Abemaciclib + Palbociclib | Compared to physiological saline, DOX-LP+ TRAIL-LP, DOX-LP, and DOX significantly prolonged the survival time (48,49,36 days). | PDGFRα and CDK4/6 signaling blockade for a splice variant. |
| Wang et al. [161] | 2011 | HCC (U87) | CCRM | EZH2 | EZH2si-DMC | Decrease in proliferation of cancer cells, reduction in cancer cell survival in vitro, and a reduction in tumor volume in nude mice. | DMC nanoparticle-mediated EZH2-siRNA decreases tumor size. |
| Schleicher et al. [146] | 2011 | HCC (Endothelial cells, HUVEC) and ACC (mice) (bEND.3) | IT | FPR | F2 Procyanidins | Mice treated with BrP-LPA and irradiation showed a tumor growth delay of 6.8 days compared to mice treated with irradiation alone. | F2 procyanidins downregulate FPR, exerting a cytotoxic effect. |
| Benezra et al. [34] | 2012 | HCC | PKP | Multitarget kinases | Dasatinib | Mice gavaged with saline vehicle survived 14 to 17 days posttreatment, whereas dasatinib-gavaged mice survived 18 to 30 days. | Dasatinib boosts survival in mouse GBM. |
| Matsuda et al. [117] | 2012 | HCC (TGS01, GS-Y01) | PKP | JNK | Sp600125 | All mice survived beyond 12 months after treatment, with no significant differences found in general health status as assessed by body weight and survival, and in cognitive function. | JNK is a target for stem-like potential in GBM. |
| Salphati et al. [140] | 2012 | HCC (MDR1-MDCKI, Bcrp1-MDCKII, Bcrp-MDCKII, Mdr1a-LLC-PK | PKP | PI3K | Gne-317 | In the GBM10 model, mice that were treated with GNE-317 experienced a marked survival benefit. | GNE-317 is a PI3K inhibitor for GBM. |
| See et al. [147] | 2012 | HCC (TCC; LN229, A172, T98G, MO59J, LN18, U87, U138) or the UCSF BTRC Tissue Core (U251, U373, SF188, U343, SF126, SF210, SF268, SF295, SF539) | PKP | MEK + PI3K/mTOR | Vemurafenib + PI103 | PD0325901 suppressed the growth of LN229 tumors and increased the survival of LN229-bearing animals, but had no significant effect on the intracranial growth of U251 cells or the survival of U251 tumor-bearing mice. | MEK inhibitor-resistant GBM lines respond to dual therapy. |
| Miyazaki et al. [121] | 2012 | HCC (GBM146, 157, 205, 206, 218, 1600, 2313, and 13, f16w and 1105A) | OM | TRAILR | Recombinant TRAIL + TMZ | N/A | TMZ + TRAIL synergistically improve survival in tumor-bearing rats. |
| Preukschas et al. [133] | 2012 | H/HCC (G55T2, U87-MG) | OM | YAP1 | Nsc682769 | Glioma patients with a high expression of eIF-5A have a lower probability of survival, compared to patients with an intermediate expression. | NSC682769 is a YAP1 inhibitor, decreasing glioma growth. |
| Dominguez et al. [57] | 2013 | HCC (U87, U251) | PKP | DGK-α | R59022 + R59949 + siRNA | N/A | DGK-α is a potential glioma target linked to pathways. |
| Peng et al. [130] | 2013 | HCC (U87 and CHG-5) | PKP | RACK1-PKC | siRNA | N/A | RACK1 is a glioma development target via SRC/Akt activity. |
| Chen et al. [43] | 2013 | HCC (U87) | MT | TFAM | Melatonin + TMZ | N/A | Melatonin enhances TMZ effects via TFAM inhibition. |
| Huveldt et al. [79] | 2013 | HCC | MT | Nrf2 | siRNA | Dasatinib effectively blocked the increased invasion induced by bevacizumab, thus its combination is recommended to be used in clinical settings. | Nrf2 promotes glioma proliferation; siRNA is a potential drug. |
| Jaszberenyi et al. [81] | 2013 | HCC (U-87) | MT | MRP3 | Anti-MRP Antibody | GHRH antagonists can increase the direct inhibitory effect of traditional chemotherapeutic drugs. | MRP3 is overexpressed in gliomas; specific antibodies decrease growth. |
| Luchman et al. [111] | 2011 | HCC (BT142) | PKP | mTOR1/2 | AZD8055 | N/A | Dual mTOR1/2 inhibition + TMZ for resistant GBM. |
| Qin et al. [135] | 2014 | A (mice and rats) + HCC | PKP | EMP2 | Anti-EMP2 antibodies/Anti-EMP2 Igg1 | Abemaciclib + TMZ increased survival by 31–37,5 days. | EMP2 promotes migration/invasion via protein kinases. |
| Signore et al. [151] | 2014 | HCC (U87MG) | PKP | PDK1 + CHK1 | UCN-01 | Combined inhibition of PDK1 and CHK1 represents a potentially effective therapeutic approach to growth reduction of human GBM. | UCN-01 downregulates PDK1 and CHK1, killing tumor cells. |
| Blanco et al. [35] | 2014 | HCC (U87ΔEGFR-Luc, U87-MG) | MT | NRP-1 | NRP-1 Mab | SapC-DOPS nanovesicles target tumor cells and exert antitumor actions both in vitro and in vivo | NRP-1Mab inhibits glioma growth and invasion. |
| Barone et al. [33] | 2014 | HCC (U87) | OM | Lactate (monocarboxylate) transporters | ACCA | CXCR4 antagonist + POL5551 + mcr84 can increase median OS in GBM xenografts compared to treatment with either drug as monotherapy. | ACCA inhibits lactate transport, a potential brain tumor target. |
| Saito et al. [138] | 2014 | HCC (U87MG and T98G) | OM | A1CF + FAM224A | shRNA | Combination therapy consisting of EFTUD1 downregulation with an autophagy blocker enhanced the antitumor effect. | A1CF/FAM224A/miR-590-3p/ZNF143 loop regulates tumor progression. |
| Di Stefano et al. [56] | 2015 | HCC | PKP | FGFR kinase | JNJ-42756493 | Targeted inhibition of FGFR-TK with JNJ-42756493 may provide clinical benefits for patients with recurrent glioma. | JNJ-42756493 inhibits growth and regression in GBM. |
| Zhang et al. [179] | 2015 | HCC (U87) | PKP | mGluR1 | siRNA, Selective Antagonists Riluzole + BAY36-7620 | mGluR1 is a potential therapeutic target for the treatment of human gliomas. | mGluR1 inhibition demonstrated antitumor activity. |
| Ge et al. [64] | 2015 | HCC (U87) | CCRM | Tumor checkpoint controller targeting microtubules | BAL101553 | anti-miR-27a could inhibit the growth of GBM and has potential for clinical application. | BAL101553 is a promising GBM agent. |
| Gu et al. [70] | 2015 | HCC (U87, SHG-44, CHG-5, and U251) | CCRM | DR4/5 | TRAIL + Doxorubicin | Inhibition of PAK5 by lentivirus-mediated RNAi suppressed glioma development. | TRAIL-LP and DOX-LP are stronger against GBM in vivo. |
| Lamour et al. [98] | 2015 | HCC (U87-MG and U251-MG) | CCRM | PLK1 | Bi2536 | N/A | PLK1 is critical to glioma cell survival. |
| Niu et al. [125] | 2015 | HCC (U87, A172, SHG44, and U251) | CCRM | XIAP + BCL-2 | RIST + ARIST | Plumbagin inhibited glioma cell proliferation and promoted apoptosis in a nude mouse model. | RIST and aRIST prolong survival, reduce tumor burden. |
| Nonnenmacher et al. [126] | 2015 | HCC (A172, D54, U118, U138, T98G, U87-MG) | CCRM | MGMT | PRIMA-1MET | RIST therapy can be considered a promising treatment strategy for GBM. | PRIMA-1MET targets p53, an effective therapy. |
| Sanzey et al. [142] | 2015 | HCC (NCH421k, NCH660h, NCH465, NCH601 and NCH644 | OM | DLL3 | Rova-T | Glycolysis is a promising target for GBM therapy. | DLL3 is selectively expressed in glioma; targetable with Rova-T. |
| Tsigelny et al. [155] | 2017 | HCC and ACC (mice GBM4, GBM8, U87, and NHA) | PKP | OLIG2 | SKOG102 | N/A | SKOG102 inhibits glioma growth via OLIG2 downregulation. |
| Mojarad-Jabali et al. [122] | 2022 | HCC (T7, B6, and T12 peptides) | MT | Fibulin-3 | Mab428.2 | N/A | mAb428.2 inhibits fibulin-3, reduces tumor growth, and extends survival. |
| Michaud et al. [120] | 2010 | HCC (U87MG, U138MG, M059J, Hs683, H4, A172, LN18, LN229, CCF-STTG1, T98G, DBTRG-05MG, 8MGBA, 42MGBA, DKMG, GAMG, GMS10, LN405, SNB19, AM38, NMC-G1, and KG-1-C) | CCRM | FOXM1 | Plumbagin | The antitumor activity of PD0332991, when used with radiation either concurrently or sequentially, is superior to monotherapy. | Plumbagin inhibits glioma proliferation, induces apoptosis. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Begagić, E.; Pugonja, R.; Bečulić, H.; Čeliković, A.; Tandir Lihić, L.; Kadić Vukas, S.; Čejvan, L.; Skomorac, R.; Selimović, E.; Jaganjac, B.; et al. Molecular Targeted Therapies in Glioblastoma Multiforme: A Systematic Overview of Global Trends and Findings. Brain Sci. 2023, 13, 1602. https://doi.org/10.3390/brainsci13111602
Begagić E, Pugonja R, Bečulić H, Čeliković A, Tandir Lihić L, Kadić Vukas S, Čejvan L, Skomorac R, Selimović E, Jaganjac B, et al. Molecular Targeted Therapies in Glioblastoma Multiforme: A Systematic Overview of Global Trends and Findings. Brain Sciences. 2023; 13(11):1602. https://doi.org/10.3390/brainsci13111602
Chicago/Turabian StyleBegagić, Emir, Ragib Pugonja, Hakija Bečulić, Amila Čeliković, Lejla Tandir Lihić, Samra Kadić Vukas, Lejla Čejvan, Rasim Skomorac, Edin Selimović, Belma Jaganjac, and et al. 2023. "Molecular Targeted Therapies in Glioblastoma Multiforme: A Systematic Overview of Global Trends and Findings" Brain Sciences 13, no. 11: 1602. https://doi.org/10.3390/brainsci13111602
APA StyleBegagić, E., Pugonja, R., Bečulić, H., Čeliković, A., Tandir Lihić, L., Kadić Vukas, S., Čejvan, L., Skomorac, R., Selimović, E., Jaganjac, B., Juković-Bihorac, F., Jusić, A., & Pojskić, M. (2023). Molecular Targeted Therapies in Glioblastoma Multiforme: A Systematic Overview of Global Trends and Findings. Brain Sciences, 13(11), 1602. https://doi.org/10.3390/brainsci13111602