
Cross-Modal Plasticity during Self-Motion Perception


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Cross-Modal Calibration and Recalibration
3. Self-Motion Perception Offers a Prime Substrate to Study Cross-Modal Recalibration
3.1. Long-Term Visual–Vestibular Recalibration
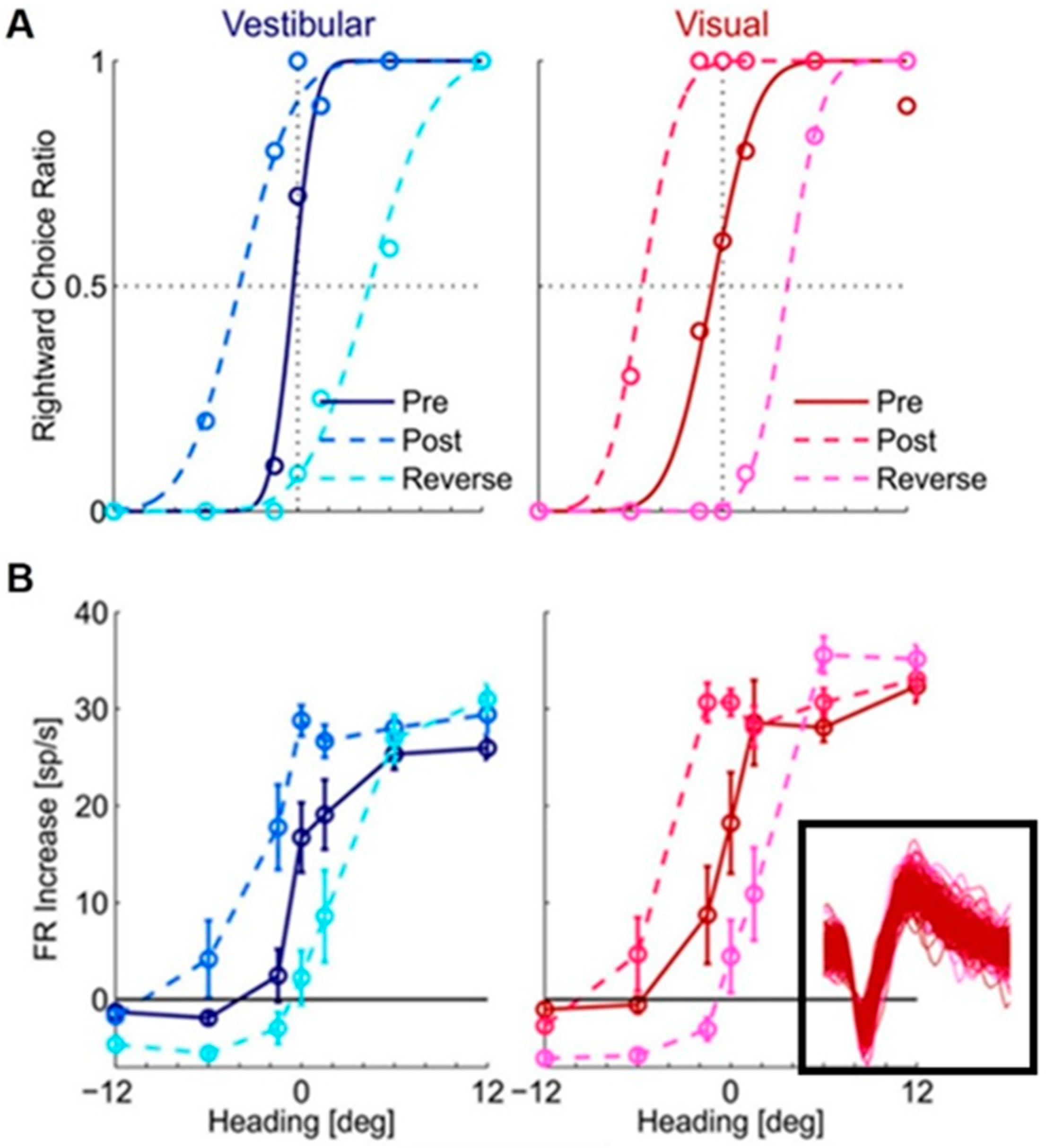
3.2. Short-Term Visual–Vestibular Recalibration
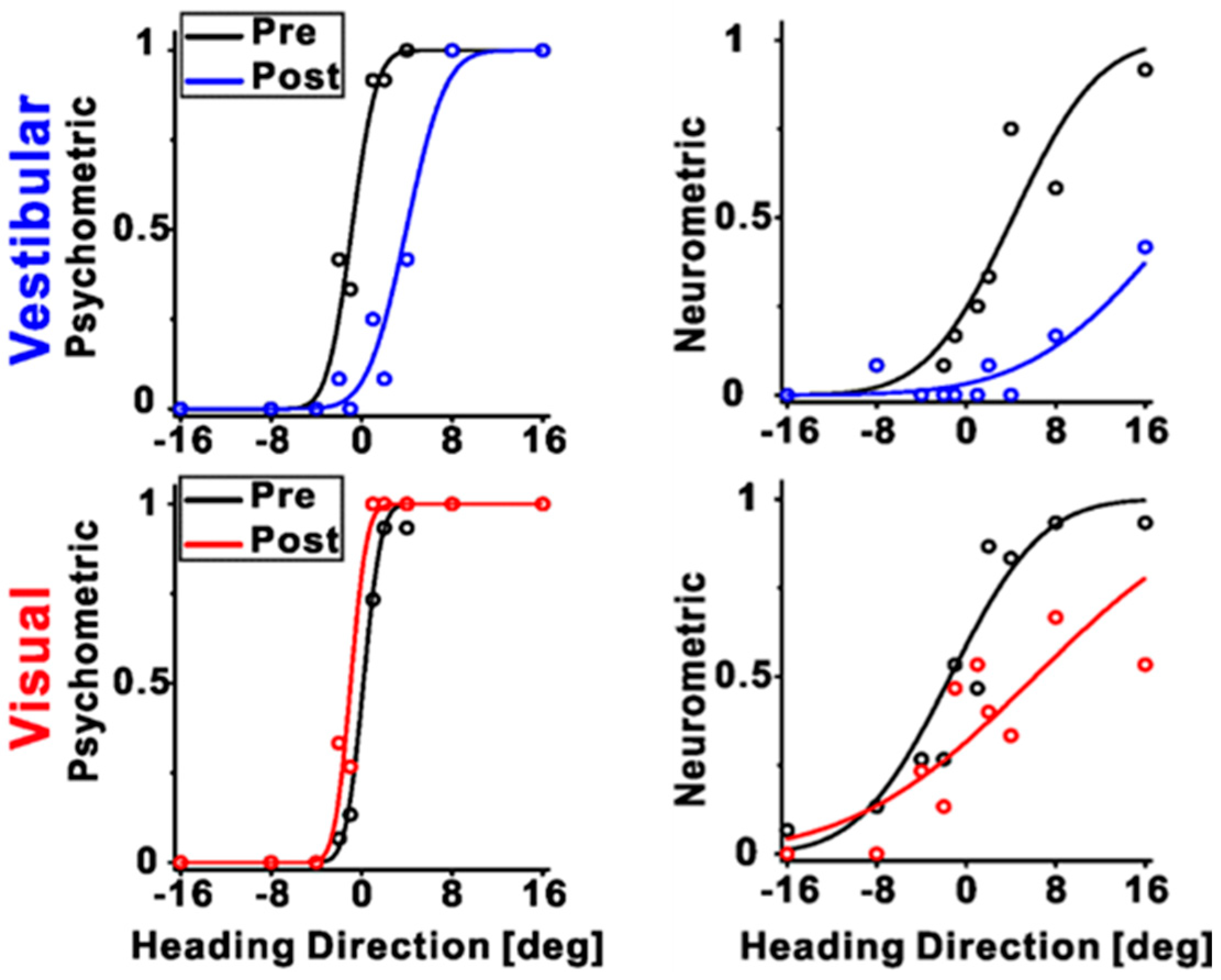
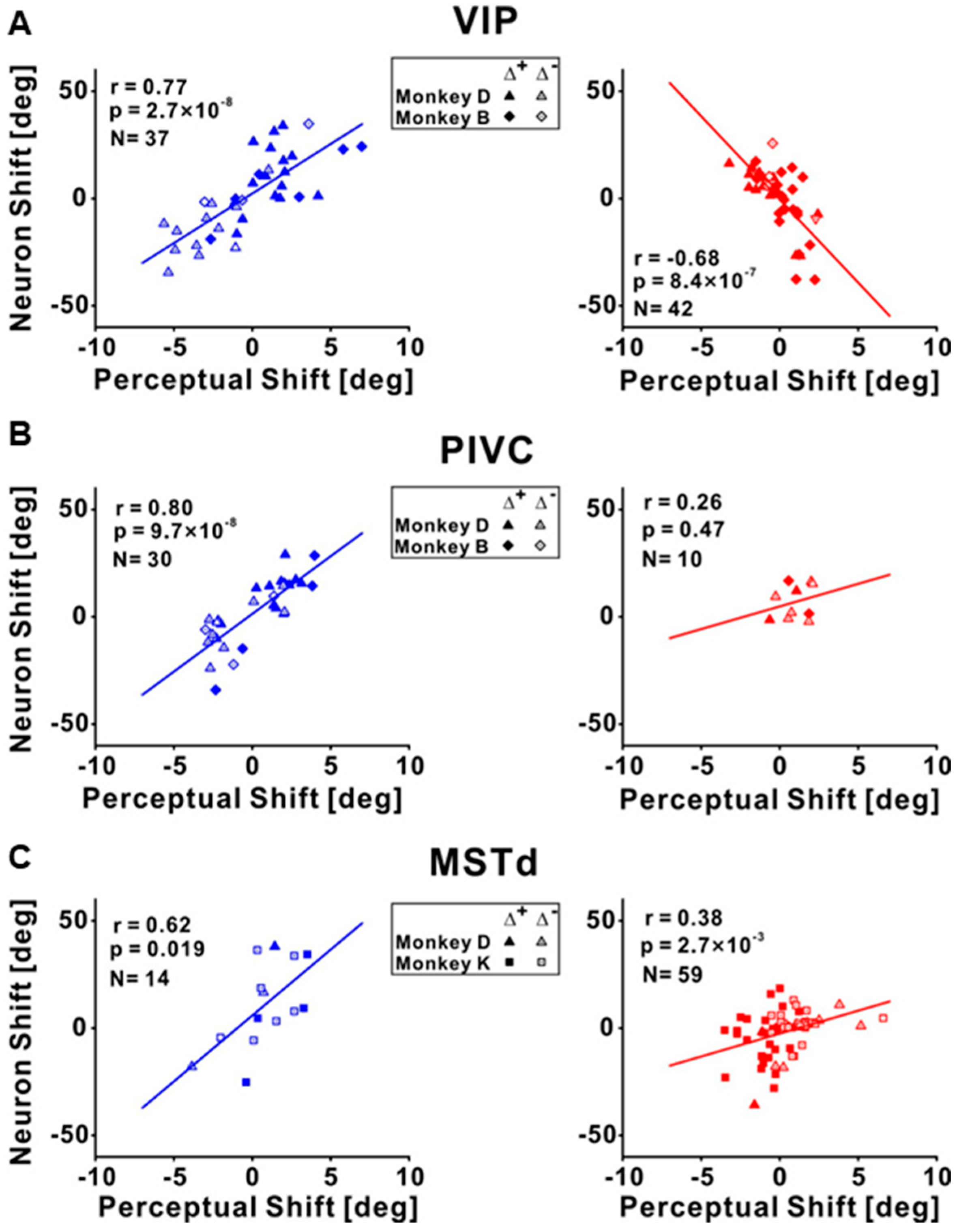
4. Neural Correlates of Visual–Vestibular Recalibration
5. Summary and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stein, B.E.; Stanford, T.R.; Rowland, B.A. Development of multisensory integration from the perspective of the individual neuron. Nat. Rev. Neurosci. 2014, 15, 520–535. [Google Scholar] [CrossRef]
- Von Kriegstein, K. Frontiers in Neuroscience a Multisensory Perspective on Human Auditory Communication. In The Neural Bases of Multisensory Processes; Murray, M.M., Wallace, M.T., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2012. [Google Scholar]
- Kayser, C.; Shams, L. Multisensory causal inference in the brain. PLoS Biol. 2015, 13, e1002075. [Google Scholar] [CrossRef] [PubMed]
- Camarata, S.; Miller, L.J.; Wallace, M.T. Evaluating Sensory Integration/Sensory Processing Treatment: Issues and Analysis. Front. Integr. Neurosci. 2020, 14, 556660. [Google Scholar] [CrossRef] [PubMed]
- Bronstein, A.M. Multisensory integration in balance control. Handb. Clin. Neurol. 2016, 137, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Marco, E.J.; Hinkley, L.B.; Hill, S.S.; Nagarajan, S.S. Sensory processing in autism: A review of neurophysiologic findings. Pediatr. Res. 2011, 69, 48r–54r. [Google Scholar] [CrossRef]
- Meilleur, A.; Foster, N.E.V.; Coll, S.M.; Brambati, S.M.; Hyde, K.L. Unisensory and multisensory temporal processing in autism and dyslexia: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2020, 116, 44–63. [Google Scholar] [CrossRef]
- Zhou, H.Y.; Wang, Y.M.; Zhang, R.T.; Cheung, E.F.C.; Pantelis, C.; Chan, R.C.K. Neural Correlates of Audiovisual Temporal Binding Window in Individuals With Schizotypal and Autistic Traits: Evidence from Resting-State Functional Connectivity. Autism Res. 2021, 14, 668–680. [Google Scholar] [CrossRef]
- Stein, B.E.; Stanford, T.R.; Rowland, B.A. Multisensory Integration and the Society for Neuroscience: Then and Now. J. Neurosci. 2020, 40, 3–11. [Google Scholar] [CrossRef]
- Gu, Y.; Watkins, P.V.; Angelaki, D.E.; DeAngelis, G.C. Visual and nonvisual contributions to three-dimensional heading selectivity in the medial superior temporal area. J. Neurosci. 2006, 26, 73–85. [Google Scholar] [CrossRef]
- Chen, A.; Deangelis, G.C.; Angelaki, D.E. Functional specializations of the ventral intraparietal area for multisensory heading discrimination. J. Neurosci. 2013, 33, 3567–3581. [Google Scholar] [CrossRef]
- Gori, M. Multisensory Integration and Calibration in Children and Adults with and without Sensory and Motor Disabilities. Multisens. Res. 2015, 28, 71–99. [Google Scholar] [CrossRef] [PubMed]
- Van Mier, H.I. Combining visual and haptic practice significantly reduced deviations in haptic parallelity matching. Acta Psychol. 2020, 203, 103008. [Google Scholar] [CrossRef] [PubMed]
- Gori, M.; Sandini, G.; Burr, D. Development of visuo-auditory integration in space and time. Front. Integr. Neurosci. 2012, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Valori, I.; McKenna-Plumley, P.E.; Bayramova, R.; Callegher, C.Z.; Altoè, G.; Farroni, T. Proprioceptive accuracy in Immersive Virtual Reality: A developmental perspective. PLoS ONE 2020, 15, e0222253. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.D.; Sigman, M.; Crist, R.E. The neural basis of perceptual learning. Neuron 2001, 31, 681–697. [Google Scholar] [CrossRef]
- Bavelier, D.; Neville, H.J. Cross-modal plasticity: Where and how? Nat. Rev. Neurosci. 2002, 3, 443–452. [Google Scholar] [CrossRef]
- Berger, C.C.; Ehrsson, H.H. Mental Imagery Induces Cross-Modal Sensory Plasticity and Changes Future Auditory Perception. Psychol. Sci. 2018, 29, 926–935. [Google Scholar] [CrossRef]
- Shimojo, S.; Shams, L. Sensory modalities are not separate modalities: Plasticity and interactions. Curr. Opin. Neurobiol. 2001, 11, 505–509. [Google Scholar] [CrossRef]
- Canon, L.K. Intermodality inconsistency of input and directed attention as determinants of the nature of adaptation. J. Exp. Psychol. 1970, 84, 141–147. [Google Scholar] [CrossRef]
- Radeau, M.; Bertelson, P. The after-effects of ventriloquism. Q. J. Exp. Psychol. 1974, 26, 63–71. [Google Scholar] [CrossRef]
- Recanzone, G.H. Rapidly induced auditory plasticity: The ventriloquism aftereffect. Proc. Natl. Acad. Sci. USA 1998, 95, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Vroomen, J.; Keetels, M.; de Gelder, B.; Bertelson, P. Recalibration of temporal order perception by exposure to audio-visual asynchrony. Brain Res. Cogn. Brain Res. 2004, 22, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Woods, T.M.; Recanzone, G.H. Visually induced plasticity of auditory spatial perception in macaques. Curr. Biol. 2004, 14, 1559–1564. [Google Scholar] [CrossRef]
- Kramer, A.; Röder, B.; Bruns, P. Feedback Modulates Audio-Visual Spatial Recalibration. Front. Integr. Neurosci. 2019, 13, 74. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.M.; Akeroyd, M.A.; Roach, N.W.; Webb, B.S. Multiple spatial reference frames underpin perceptual recalibration to audio-visual discrepancies. PLoS ONE 2021, 16, e0251827. [Google Scholar] [CrossRef] [PubMed]
- Botvinick, M.; Cohen, J. Rubber hands ‘feel’ touch that eyes see. Nature 1998, 391, 756. [Google Scholar] [CrossRef]
- Tsakiris, M.; Haggard, P. Experimenting with the acting self. Cogn. Neuropsychol. 2005, 22, 387–407. [Google Scholar] [CrossRef]
- Abdulkarim, Z.; Hayatou, Z.; Ehrsson, H.H. Sustained rubber hand illusion after the end of visuotactile stimulation with a similar time course for the reduction of subjective ownership and proprioceptive drift. Exp. Brain Res. 2021, 239, 3471–3486. [Google Scholar] [CrossRef]
- Thériault, R.; Landry, M.; Raz, A. The Rubber Hand Illusion: Top-down attention modulates embodiment. Q. J. Exp. Psychol. 2022, 75, 2129–2148. [Google Scholar] [CrossRef]
- Bruns, P.; Röder, B. Sensory recalibration integrates information from the immediate and the cumulative past. Sci. Rep. 2015, 5, 12739. [Google Scholar] [CrossRef]
- Lewald, J. Rapid adaptation to auditory-visual spatial disparity. Learn. Mem. 2002, 9, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Burge, J.; Ernst, M.O.; Banks, M.S. The statistical determinants of adaptation rate in human reaching. J. Vis. 2008, 8, 20. [Google Scholar] [CrossRef]
- Burge, J.; Girshick, A.R.; Banks, M.S. Visual-haptic adaptation is determined by relative reliability. J. Neurosci. 2010, 30, 7714–7721. [Google Scholar] [CrossRef] [PubMed]
- Zierul, B.; Röder, B.; Tempelmann, C.; Bruns, P.; Noesselt, T. The role of auditory cortex in the spatial ventriloquism aftereffect. Neuroimage 2017, 162, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Kayser, C. The Neurophysiological Basis of the Trial-Wise and Cumulative Ventriloquism Aftereffects. J. Neurosci. 2021, 41, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Scheliga, S.; Kellermann, T.; Lampert, A.; Rolke, R.; Spehr, M.; Habel, U. Neural correlates of multisensory integration in the human brain: An ALE meta-analysis. Rev. Neurosci. 2023, 34, 223–245. [Google Scholar] [CrossRef]
- Fernández, L.M.; Torralba, M.; Soto-Faraco, S. Theta oscillations reflect conflict processing in the perception of the McGurk illusion. Eur. J. Neurosci. 2018, 48, 2630–2641. [Google Scholar] [CrossRef]
- Knudsen, E.I. Instructed learning in the auditory localization pathway of the barn owl. Nature 2002, 417, 322–328. [Google Scholar] [CrossRef]
- Linkenhoker, B.A.; Knudsen, E.I. Incremental training increases the plasticity of the auditory space map in adult barn owls. Nature 2002, 419, 293–296. [Google Scholar] [CrossRef]
- Kaliuzhna, M.; Ferrè, E.R.; Herbelin, B.; Blanke, O.; Haggard, P. Multisensory effects on somatosensation: A trimodal visuo-vestibular-tactile interaction. Sci. Rep. 2016, 6, 26301. [Google Scholar] [CrossRef]
- Hummel, N.; Cuturi, L.F.; MacNeilage, P.R.; Flanagin, V.L. The effect of supine body position on human heading perception. J. Vis. 2016, 16, 19. [Google Scholar] [CrossRef] [PubMed]
- Bayramova, R.; Valori, I.; McKenna-Plumley, P.E.; Callegher, C.Z.; Farroni, T. The role of vision and proprioception in self-motion encoding: An immersive virtual reality study. Atten. Percept. Psychophys. 2021, 83, 2865–2878. [Google Scholar] [CrossRef] [PubMed]
- Dokka, K.; DeAngelis, G.C.; Angelaki, D.E. Multisensory Integration of Visual and Vestibular Signals Improves Heading Discrimination in the Presence of a Moving Object. J. Neurosci. 2015, 35, 13599–13607. [Google Scholar] [CrossRef] [PubMed]
- Deveze, A.; Bernard-Demanze, L.; Xavier, F.; Lavieille, J.P.; Elziere, M. Vestibular compensation and vestibular rehabilitation. Current concepts and new trends. Neurophysiol. Clin. 2014, 44, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Tighilet, B.; Chabbert, C. Adult neurogenesis promotes balance recovery after vestibular loss. Prog. Neurobiol. 2019, 174, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Black, F.O.; Paloski, W.H.; Doxey-Gasway, D.D.; Reschke, M.F. Vestibular plasticity following orbital spaceflight: Recovery from postflight postural instability. Acta Otolaryngol. Suppl. 1995, 520 Pt 2, 450–454. [Google Scholar] [CrossRef]
- Shupak, A.; Gordon, C.R. Motion sickness: Advances in pathogenesis, prediction, prevention, and treatment. Aviat. Space Environ. Med. 2006, 77, 1213–1223. [Google Scholar]
- Crane, B.T. Direction specific biases in human visual and vestibular heading perception. PLoS ONE 2012, 7, e51383. [Google Scholar] [CrossRef]
- Gordon, C.R.; Fletcher, W.A.; Melvill Jones, G.; Block, E.W. Adaptive plasticity in the control of locomotor trajectory. Exp. Brain Res. 1995, 102, 540–545. [Google Scholar] [CrossRef]
- Nooij, S.A.; Groen, E.L. Rolling into spatial disorientation: Simulator demonstration of the post-roll (Gillingham) illusion. Aviat. Space Environ. Med. 2011, 82, 505–512. [Google Scholar] [CrossRef]
- Fetsch, C.R.; Turner, A.H.; DeAngelis, G.C.; Angelaki, D.E. Dynamic reweighting of visual and vestibular cues during self-motion perception. J. Neurosci. 2009, 29, 15601–15612. [Google Scholar] [CrossRef] [PubMed]
- Zaidel, A.; Turner, A.H.; Angelaki, D.E. Multisensory calibration is independent of cue reliability. J. Neurosci. 2011, 31, 13949–13962. [Google Scholar] [CrossRef] [PubMed]
- Zaidel, A.; Ma, W.J.; Angelaki, D.E. Supervised calibration relies on the multisensory percept. Neuron 2013, 80, 1544–1557. [Google Scholar] [CrossRef] [PubMed]
- Shalom-Sperber, S.; Chen, A.; Zaidel, A. Rapid cross-sensory adaptation of self-motion perception. Cortex 2022, 148, 14–30. [Google Scholar] [CrossRef]
- Alais, D.; Burr, D. The ventriloquist effect results from near-optimal bimodal integration. Curr. Biol. 2004, 14, 257–262. [Google Scholar] [CrossRef]
- Crane, B.T. Limited interaction between translation and visual motion aftereffects in humans. Exp. Brain Res. 2013, 224, 165–178. [Google Scholar] [CrossRef][Green Version]
- Cuturi, L.F.; MacNeilage, P.R. Optic flow induces nonvisual self-motion aftereffects. Curr. Biol. 2014, 24, 2817–2821. [Google Scholar] [CrossRef]
- Chen, A.; DeAngelis, G.C.; Angelaki, D.E. Representation of vestibular and visual cues to self-motion in ventral intraparietal cortex. J. Neurosci. 2011, 31, 12036–12052. [Google Scholar] [CrossRef]
- Chen, A.; DeAngelis, G.C.; Angelaki, D.E. Convergence of vestibular and visual self-motion signals in an area of the posterior sylvian fissure. J. Neurosci. 2011, 31, 11617–11627. [Google Scholar] [CrossRef]
- Gu, Y.; Cheng, Z.; Yang, L.; DeAngelis, G.C.; Angelaki, D.E. Multisensory Convergence of Visual and Vestibular Heading Cues in the Pursuit Area of the Frontal Eye Field. Cereb. Cortex 2016, 26, 3785–3801. [Google Scholar] [CrossRef]
- Zhao, B.; Zhang, Y.; Chen, A. Encoding of vestibular and optic flow cues to self-motion in the posterior superior temporal polysensory area. J. Physiol. 2021, 599, 3937–3954. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Angelaki, D.E.; Deangelis, G.C. Neural correlates of multisensory cue integration in macaque MSTd. Nat. Neurosci. 2008, 11, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Deangelis, G.C.; Angelaki, D.E. Causal links between dorsal medial superior temporal area neurons and multisensory heading perception. J. Neurosci. 2012, 32, 2299–2313. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Gu, Y.; Liu, S.; DeAngelis, G.C.; Angelaki, D.E. Evidence for a Causal Contribution of Macaque Vestibular, but Not Intraparietal, Cortex to Heading Perception. J. Neurosci. 2016, 36, 3789–3798. [Google Scholar] [CrossRef]
- Zaidel, A.; DeAngelis, G.C.; Angelaki, D.E. Decoupled choice-driven and stimulus-related activity in parietal neurons may be misrepresented by choice probabilities. Nat. Commun. 2017, 8, 715. [Google Scholar] [CrossRef]
- Gu, Y. Vestibular signals in primate cortex for self-motion perception. Curr. Opin. Neurobiol. 2018, 52, 10–17. [Google Scholar] [CrossRef]
- Morgan, M.L.; Deangelis, G.C.; Angelaki, D.E. Multisensory integration in macaque visual cortex depends on cue reliability. Neuron 2008, 59, 662–673. [Google Scholar] [CrossRef]
- Fetsch, C.R.; Pouget, A.; DeAngelis, G.C.; Angelaki, D.E. Neural correlates of reliability-based cue weighting during multisensory integration. Nat. Neurosci. 2011, 15, 146–154. [Google Scholar] [CrossRef]
- Zaidel, A.; Laurens, J.; DeAngelis, G.C.; Angelaki, D.E. Supervised Multisensory Calibration Signals Are Evident in VIP but Not MSTd. J. Neurosci. 2021, 41, 10108–10119. [Google Scholar] [CrossRef]
- Zeng, F.; Zaidel, A.; Chen, A. Contrary neuronal recalibration in different multisensory cortical areas. eLife 2023, 12, e82895. [Google Scholar] [CrossRef]
- Zaidel, A.; Salomon, R. Multisensory decisions from self to world. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2023, 378, 20220335. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, J.C.; Rowland, B.A.; Stanford, T.R.; Stein, B.E. A neural network model of multisensory integration also accounts for unisensory integration in superior colliculus. Brain Res. 2008, 1242, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.D.; Chen, Z.; Xiang, X.; Hu, M.; Xie, H.; Jia, X.; Cai, F.; Cui, Y.; Chen, Z.; Qian, L.; et al. Zona incerta GABAergic neurons integrate prey-related sensory signals and induce an appetitive drive to promote hunting. Nat. Neurosci. 2019, 22, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.D.; Lindsay, N.M.; Deschênes, M.; Kleinfeld, D. Vibrissa Self-Motion and Touch Are Reliably Encoded along the Same Somatosensory Pathway from Brainstem through Thalamus. PLoS Biol. 2015, 13, e1002253. [Google Scholar] [CrossRef] [PubMed]
- Bina, L.; Romano, V.; Hoogland, T.M.; Bosman, L.W.J.; De Zeeuw, C.I. Purkinje cells translate subjective salience into readiness to act and choice performance. Cell Rep. 2021, 37, 110116. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, R.; Zeng, F.; Wang, Q.; Chen, A. Cross-Modal Plasticity during Self-Motion Perception. Brain Sci. 2023, 13, 1504. https://doi.org/10.3390/brainsci13111504
Lin R, Zeng F, Wang Q, Chen A. Cross-Modal Plasticity during Self-Motion Perception. Brain Sciences. 2023; 13(11):1504. https://doi.org/10.3390/brainsci13111504
Chicago/Turabian StyleLin, Rushi, Fu Zeng, Qingjun Wang, and Aihua Chen. 2023. "Cross-Modal Plasticity during Self-Motion Perception" Brain Sciences 13, no. 11: 1504. https://doi.org/10.3390/brainsci13111504
APA StyleLin, R., Zeng, F., Wang, Q., & Chen, A. (2023). Cross-Modal Plasticity during Self-Motion Perception. Brain Sciences, 13(11), 1504. https://doi.org/10.3390/brainsci13111504