
Spatial Olfactory Memory and Spatial Olfactory Navigation, Assessed with a Variant of Corsi Test, Is Modulated by Gender and Sporty Activity

 ,
, 


Abstract
:1. Introduction
1.1. Human Spatial Representation and Navigation
1.2. Spatial Navigation and Olfaction in Humans
1.3. Spatial Navigation in Athletes and Non-Athletes
1.4. Visual–Spatial Memory and Olfaction
2. Materials and Methods
2.1. Participants
2.2. Olfactory Stimuli
2.3. Assessment
2.4. Statistical Analysis
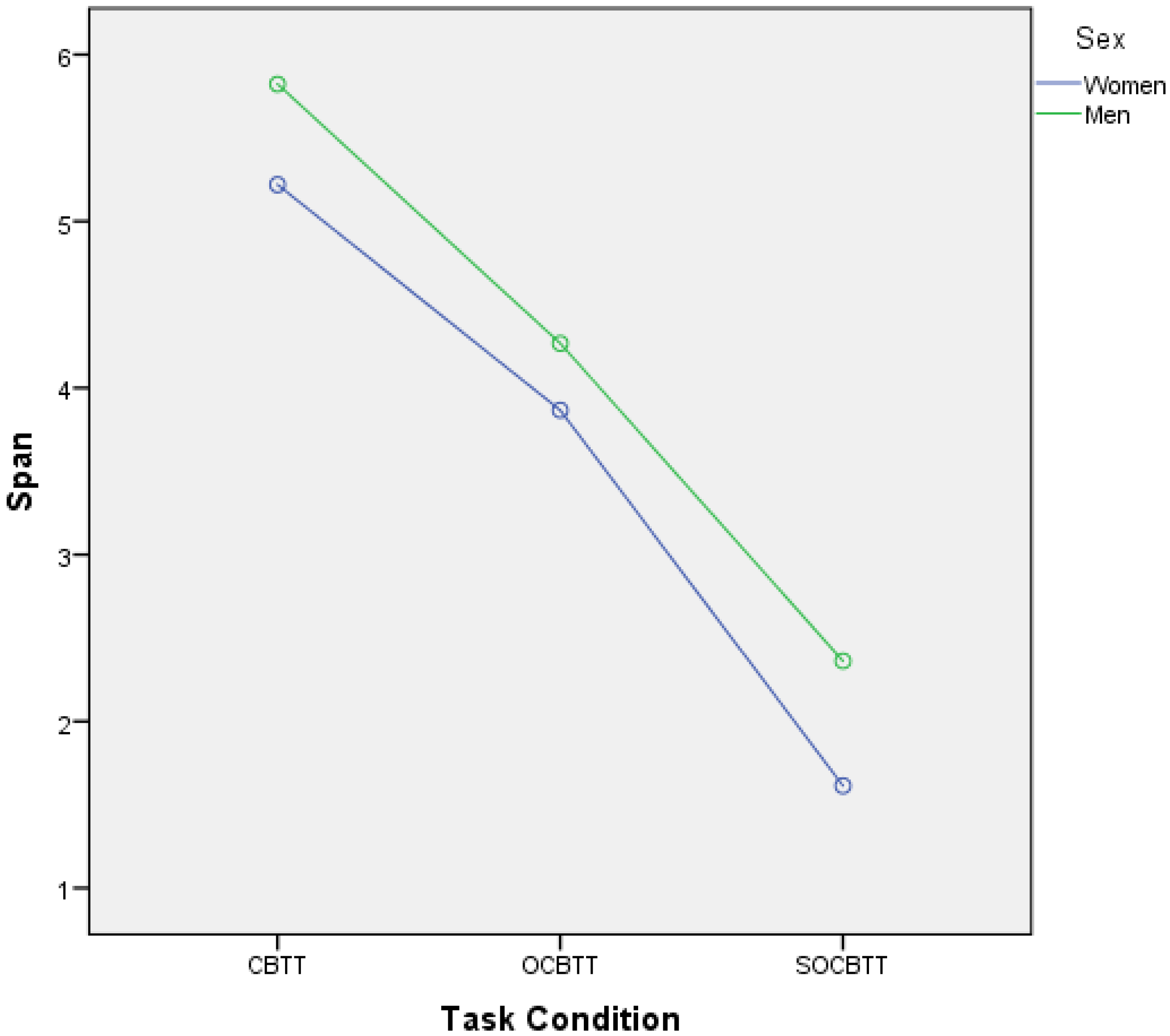
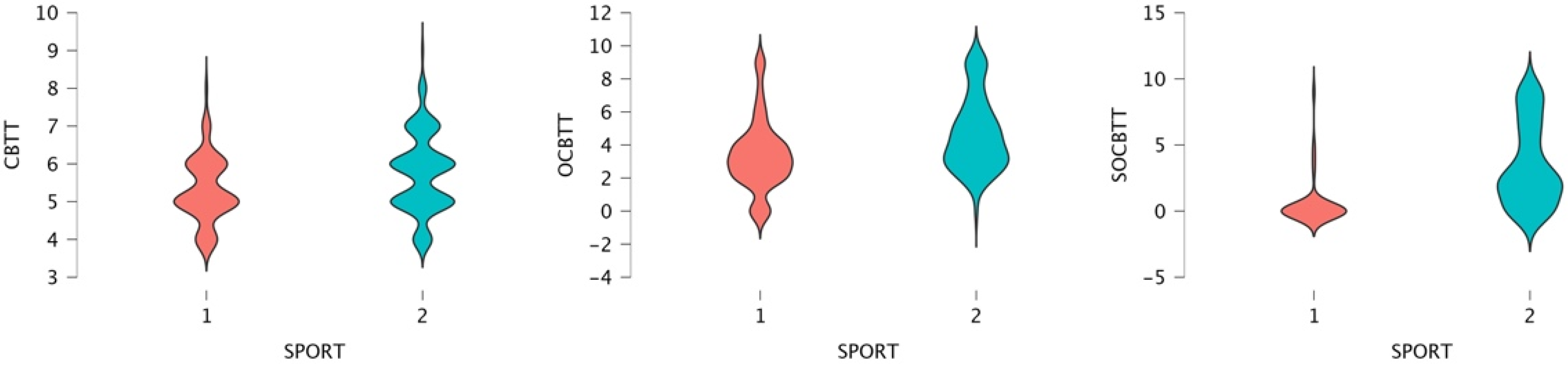
3. Results
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Werner, S.; Krieg-Brueckner, B.; Mallot, H.; Schweizer, K.; Freksa, C. Spatial Cognition: The Role of Landmark, Route, and Survey Knowledge in Human and Robot Navigation. In Informatik ’97 Informatik als Innovationsmotor; Springer: Berlin, Heidelberg, 1997; Available online: http://www.cosy.informatik.uni-bremen.de/spp/SPP_onlines/ProjektD/gi97.pdf (accessed on 1 August 2022). [CrossRef]
- Chan, E.; Baumann, O.; Bellgrove, M.; Mattingley, J. From Objects to Landmarks: The Function of Visual Location Information in Spatial Navigation. Front. Psychol. 2012, 3, 304. [Google Scholar] [CrossRef]
- Chrastil, E.R. Neural Evidence Supports a Novel Framework for Spatial Navigation. Psychon. Bull. Rev. 2013, 20, 208–227. [Google Scholar] [CrossRef] [PubMed]
- Chrastil, E.R.; Warren, W.H. Active and Passive Spatial Learning in Human Navigation: Acquisition of Survey Knowledge. J. Exp. Psychol. Learn. Mem. Cogn. 2013, 39, 1520–1537. [Google Scholar] [CrossRef]
- Ekstrom, A.D.; Isham, E.A. Human Spatial Navigation: Representations across Dimensions and Scales. Curr. Opin. Behav. Sci. 2017, 17, 84–89. [Google Scholar] [CrossRef]
- Ekstrom, A.D.; Kahana, M.J.; Caplan, J.B.; Fields, T.A.; Isham, E.A.; Newman, E.L.; Fried, I. Cellular Networks Underlying Human Spatial Navigation. Nature 2003, 425, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Eichenbaum, H. The Role of the Hippocampus in Navigation Is Memory. J. Neurophysiol. 2017, 117, 1785–1796. [Google Scholar] [CrossRef]
- Epstein, R.A.; Patai, E.Z.; Julian, J.B.; Spiers, H.J. The Cognitive Map in Humans: Spatial Navigation and Beyond. Nat. Neurosci. 2017, 20, 1504–1513. [Google Scholar] [CrossRef]
- Epstein, R.A. Parahippocampal and Retrosplenial Contributions to Human Spatial Navigation. Trends Cogn. Sci. 2008, 12, 388–396. [Google Scholar] [CrossRef]
- Jacobs, J.; Weidemann, C.T.; Miller, J.F.; Solway, A.; Burke, J.F.; Wei, X.-X.; Suthana, N.; Sperling, M.R.; Sharan, A.D.; Fried, I.; et al. Direct Recordings of Grid-like Neuronal Activity in Human Spatial Navigation. Nat. Neurosci. 2013, 16, 1188–1190. [Google Scholar] [CrossRef]
- Jacobs, J.; Korolev, I.O.; Caplan, J.B.; Ekstrom, A.D.; Litt, B.; Baltuch, G.; Fried, I.; Schulze-Bonhage, A.; Madsen, J.R.; Kahana, M.J. Right-Lateralized Brain Oscillations in Human Spatial Navigation. J. Cogn. Neurosci. 2010, 22, 824–836. [Google Scholar] [CrossRef]
- Moffat, S.D.; Elkins, W.; Resnick, S.M. Age Differences in the Neural Systems Supporting Human Allocentric Spatial Navigation. Neurobiol. Aging 2006, 27, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Sneider, J.T.; Hamilton, D.A.; Cohen-Gilbert, J.E.; Crowley, D.J.; Rosso, I.M.; Silveri, M.M. Sex Differences in Spatial Navigation and Perception in Human Adolescents and Emerging Adults. Behav. Process. 2015, 111, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Baizan, C.; Arias, J.L.; Mendez, M. Spatial Memory Assessment Reveals Age-Related Differences in Egocentric and Allocentric Memory Performance. Behav. Brain Res. 2020, 388, 112646. [Google Scholar] [CrossRef]
- Jacobs, L.F. From Chemotaxis to the Cognitive Map: The Function of Olfaction. Proc. Natl. Acad. Sci. USA 2012, 109, 10693–10700. [Google Scholar] [CrossRef] [PubMed]
- Aboitiz, F.; Montiel, J.F. Olfaction, Navigation, and the Origin of Isocortex. Front. Neurosci. 2015, 9, 402. [Google Scholar] [CrossRef]
- Rossier, J.; Schenk, F. Olfactory and/or Visual Cues for Spatial Navigation through Ontogeny: Olfactory Cues Enable the Use of Visual Cues. Behav. Neurosci. 2003, 117, 412–425. [Google Scholar] [CrossRef]
- Gagliardo, A.; Bried, J.; Lambardi, P.; Luschi, P.; Wikelski, M.; Bonadonna, F. Oceanic Navigation in Cory’s Shearwaters: Evidence for a Crucial Role of Olfactory Cues for Homing after Displacement. J. Exp. Biol. 2013, 216, 2798–2805. [Google Scholar] [CrossRef]
- Bao, X.; Gjorgieva, E.; Shanahan, L.K.; Howard, J.D.; Kahnt, T.; Gottfried, J.A. Grid-like Neural Representations Support Olfactory Navigation of a Two-Dimensional Odor Space. Neuron 2019, 102, 1066–1075.e5. [Google Scholar] [CrossRef]
- Dahmani, L.; Courcot, B.; Near, J.; Patel, R.; Amaral, R.S.C.; Chakravarty, M.M.; Bohbot, V.D. Fimbria-Fornix Volume Is Associated with Spatial Memory and Olfactory Identification in Humans. Front. Syst. Neurosci. 2020, 13, 87. [Google Scholar] [CrossRef]
- Dahmani, L.; Patel, R.M.; Yang, Y.; Chakravarty, M.M.; Fellows, L.K.; Bohbot, V.D. An Intrinsic Association between Olfactory Identification and Spatial Memory in Humans. Nat. Commun. 2018, 9, 4162. [Google Scholar] [CrossRef]
- Hamburger, K.; Knauff, M. Odors Can Serve as Landmarks in Human Wayfinding. Cogn. Sci. 2019, 43, e12798. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, L.F.; Arter, J.; Cook, A.; Sulloway, F.J. Olfactory Orientation and Navigation in Humans. PLoS ONE 2015, 10, e0129387. [Google Scholar] [CrossRef] [PubMed]
- Goodrich-Hunsaker, N.J.; Gilbert, P.E.; Hopkins, R.O. The Role of the Human Hippocampus in Odor-Place Associative Memory. Chem. Senses 2009, 34, 513–521. [Google Scholar] [CrossRef] [PubMed]
- van Uffelen, J.G.Z.; Chin, A.; Paw, M.J.M.; Hopman-Rock, M.; van Mechelen, W. The Effects of Exercise on Cognition in Older Adults with and without Cognitive Decline: A Systematic Review. Clin. J. Sport Med. 2008, 18, 486–500. [Google Scholar] [CrossRef]
- Sabia, S.; Dugravot, A.; Dartigues, J.-F.; Abell, J.; Elbaz, A.; Kivimäki, M.; Singh-Manoux, A. Physical Activity, Cognitive Decline, and Risk of Dementia: 28 Year Follow-up of Whitehall II Cohort Study. BMJ 2017, 357, j2709. [Google Scholar] [CrossRef]
- Bherer, L.; Erickson, K.I.; Liu-Ambrose, T. A Review of the Effects of Physical Activity and Exercise on Cognitive and Brain Functions in Older Adults. J. Aging Res. 2013, 2013, 657508. [Google Scholar] [CrossRef]
- Brasure, M.; Desai, P.; Davila, H.; Nelson, V.A.; Calvert, C.; Jutkowitz, E.; Butler, M.; Fink, H.A.; Ratner, E.; Hemmy, L.S.; et al. Physical Activity Interventions in Preventing Cognitive Decline and Alzheimer-Type Dementia: A Systematic Review. Ann. Intern. Med. 2018, 168, 30–38. [Google Scholar] [CrossRef]
- Erickson, K.I.; Hillman, C.H.; Kramer, A.F. Physical Activity, Brain, and Cognition. Curr. Opin. Behav. Sci. 2015, 4, 27–32. [Google Scholar] [CrossRef]
- Lista, I.; Sorrentino, G. Biological Mechanisms of Physical Activity in Preventing Cognitive Decline. Cell. Mol. Neurobiol. 2010, 30, 493–503. [Google Scholar] [CrossRef]
- Park, H.; Park, J.H.; Na, H.R.; Hiroyuki, S.; Kim, G.M.; Jung, M.K.; Kim, W.K.; Park, K.W. Combined Intervention of Physical Activity, Aerobic Exercise, and Cognitive Exercise Intervention to Prevent Cognitive Decline for Patients with Mild Cognitive Impairment: A Randomized Controlled Clinical Study. J. Clin. Med. 2019, 8, E940. [Google Scholar] [CrossRef]
- Colcombe, S.J.; Erickson, K.I.; Scalf, P.E.; Kim, J.S.; Prakash, R.; McAuley, E.; Elavsky, S.; Marquez, D.X.; Hu, L.; Kramer, A.F. Aerobic Exercise Training Increases Brain Volume in Aging Humans. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2006, 61, 1166–1170. [Google Scholar] [CrossRef]
- Erickson, K.I.; Prakash, R.S.; Voss, M.W.; Chaddock, L.; Hu, L.; Morris, K.S.; White, S.M.; Wójcicki, T.R.; McAuley, E.; Kramer, A.F. Aerobic Fitness Is Associated with Hippocampal Volume in Elderly Humans. Hippocampus 2009, 19, 1030–1039. [Google Scholar] [CrossRef]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise Training Increases Size of Hippocampus and Improves Memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar] [CrossRef]
- Brini, S.; Sohrabi, H.R.; Peiffer, J.J.; Karrasch, M.; Hämäläinen, H.; Martins, R.N.; Fairchild, T.J. Physical Activity in Preventing Alzheimer’s Disease and Cognitive Decline: A Narrative Review. Sports Med. 2018, 48, 29–44. [Google Scholar] [CrossRef]
- Guure, C.B.; Ibrahim, N.A.; Adam, M.B.; Said, S.M. Impact of Physical Activity on Cognitive Decline, Dementia, and Its Subtypes: Meta-Analysis of Prospective Studies. BioMed Res. Int. 2017, 2017, 9016924. [Google Scholar] [CrossRef]
- Cynthia, J.; Lubis, L. Vitriana Spatial Ability Differences in Athletes and Non-Athletes. AMJ 2016, 3, 533–537. [Google Scholar] [CrossRef]
- Jansen, P.; Lehmann, J. Mental Rotation Performance in Soccer Players and Gymnasts in an Object-Based Mental Rotation Task. Adv. Cogn. Psychol. 2013, 9, 92–98. [Google Scholar] [CrossRef]
- Simpon, W.F.; Coady, R.C.; Osowski, E.E.; Bode, D.S. The Effect of Aromatherapy on Exercise Performance. Kinesiol. On-Line. 2001, 9. [Google Scholar]
- Marchand, S.; Arsenault, P. Odors Modulate Pain PerceptionA Gender-Specific Effect. Physiol. Behav. 2002, 76, 251–256. [Google Scholar] [CrossRef]
- Barker, S.; Grayhem, P.; Koon, J.; Perkins, J.; Whalen, A.; Raudenbush, B. Improved Performance on Clerical Tasks Associated with Administration of Peppermint Odor. Percept. Mot. Ski. 2003, 97, 1007–1010. [Google Scholar] [CrossRef]
- Gamble, C. The Cambridge Encyclopedia of Human Evolution. Int. J. Osteoarchaeol. 1994, 4, 265–266. [Google Scholar] [CrossRef]
- Raudenbush, B.; Corley, N.; Eppich, W. Enhancing Athletic Performance through the Administration of Peppermint Odor. J. Sport Exerc. Psychol. 2001, 23, 156–160. [Google Scholar] [CrossRef]
- Basevitch, I.; Thompson, B.; Braun, R.; Razon, S.; Arsal, G.; Tokac, U.; Filho, E.M.; Nascimento, T.; Tenenbaum, G. Olfactory Effects on Attention Allocation and Perception of Exertion. Sport Psychol. 2011, 25, 144–158. [Google Scholar] [CrossRef]
- Corsi, P.M. Human Memory and the Medial Temporal Region of the Brain. Available online: https://escholarship.mcgill.ca/concern/theses/05741s554 (accessed on 1 August 2022).
- Orsini, A.; Grossi, D.; Capitani, E.; Laiacona, M.; Papagno, C.; Vallar, G. Verbal and Spatial Immediate Memory Span: Normative Data from 1355 Adults and 1112 Children. Ital. J. Neurol. Sci. 1987, 8, 537–548. [Google Scholar] [CrossRef]
- Milner, B. Interhemispheric Differences in the Localization of Psychological Processes in Man. Br. Med. Bull. 1971, 27, 272–277. [Google Scholar] [CrossRef]
- Bianchi, A.; Dai Prà, M. Twenty Years after Spinnler and Tognoni: New Instruments in the Italian Neuropsychologist’s Toolbox. Neurol. Sci. 2008, 29, 209–217. [Google Scholar] [CrossRef]
- Bannerman, D.; Yee, B.; Lemaire, M.; Wilbrecht, L.; Jarrard, L.; Iversen, S.; Rawlins, J.; Good, M. The Role of the Entorhinal Cortex in Two Forms of Spatial Learning and Memory. Exp. Brain Res. 2001, 141, 281–303. [Google Scholar] [CrossRef]
- Invitto, S.; Grasso, A. Chemosensory Perception: A Review on Electrophysiological Methods in “Cognitive Neuro-Olfactometry”. Chemosensors 2019, 7, 45. [Google Scholar] [CrossRef]
- Bowman, J.K.; Boone, R.T.; Goldman, S.; Auerbach, A. The Athletic Intelligence Quotient and Performance Outcomes in Professional Baseball. Front. Psychol. 2021, 12, 629827. [Google Scholar] [CrossRef]
- Wilson, R.S.; David, G.K.; Murphy, S.C.; Angilletta, M.J.; Niehaus, A.C.; Hunter, A.H.; Smith, M.D. Skill Not Athleticism Predicts Individual Variation in Match Performance of Soccer Players. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170953. [Google Scholar] [CrossRef]
- Invitto, S.; Piraino, G.; Ciccarese, V.; Carmillo, L.; Caggiula, M.; Trianni, G.; Nicolardi, G.; Di Nuovo, S.; Balconi, M. Potential Role of OERP as Early Marker of Mild Cognitive Impairment. Front. Aging Neurosci. 2018, 10, 272. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Odorant | Common Parfum Name | Corsi Block Number |
|---|---|---|
| Acetophenone | Solvent | 8 |
| Carvone | Mint | 2 |
| Cinnamaldehyde | Cinnamon | 9 |
| Eucalyptol | Eucalyptus | 1 |
| Eugenol | Cloves | 3 |
| Geraniol | Geranium | 5 |
| Hexanal | Grass | 7 |
| Isoamyl Acetate | Banana | 4 |
| Phenethyl Alcohol | Rose | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Invitto, S.; Accogli, G.; Leucci, M.; Salonna, M.; Serio, T.; Fancello, F.; Ciccarese, V.; Lankford, D. Spatial Olfactory Memory and Spatial Olfactory Navigation, Assessed with a Variant of Corsi Test, Is Modulated by Gender and Sporty Activity. Brain Sci. 2022, 12, 1108. https://doi.org/10.3390/brainsci12081108
Invitto S, Accogli G, Leucci M, Salonna M, Serio T, Fancello F, Ciccarese V, Lankford D. Spatial Olfactory Memory and Spatial Olfactory Navigation, Assessed with a Variant of Corsi Test, Is Modulated by Gender and Sporty Activity. Brain Sciences. 2022; 12(8):1108. https://doi.org/10.3390/brainsci12081108
Chicago/Turabian StyleInvitto, Sara, Giuseppe Accogli, Mariangela Leucci, Marika Salonna, Tonia Serio, Francesca Fancello, Vincenzo Ciccarese, and Dion Lankford. 2022. "Spatial Olfactory Memory and Spatial Olfactory Navigation, Assessed with a Variant of Corsi Test, Is Modulated by Gender and Sporty Activity" Brain Sciences 12, no. 8: 1108. https://doi.org/10.3390/brainsci12081108
APA StyleInvitto, S., Accogli, G., Leucci, M., Salonna, M., Serio, T., Fancello, F., Ciccarese, V., & Lankford, D. (2022). Spatial Olfactory Memory and Spatial Olfactory Navigation, Assessed with a Variant of Corsi Test, Is Modulated by Gender and Sporty Activity. Brain Sciences, 12(8), 1108. https://doi.org/10.3390/brainsci12081108