
Occlusal Disharmony—A Potential Factor Promoting Depression in a Rat Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
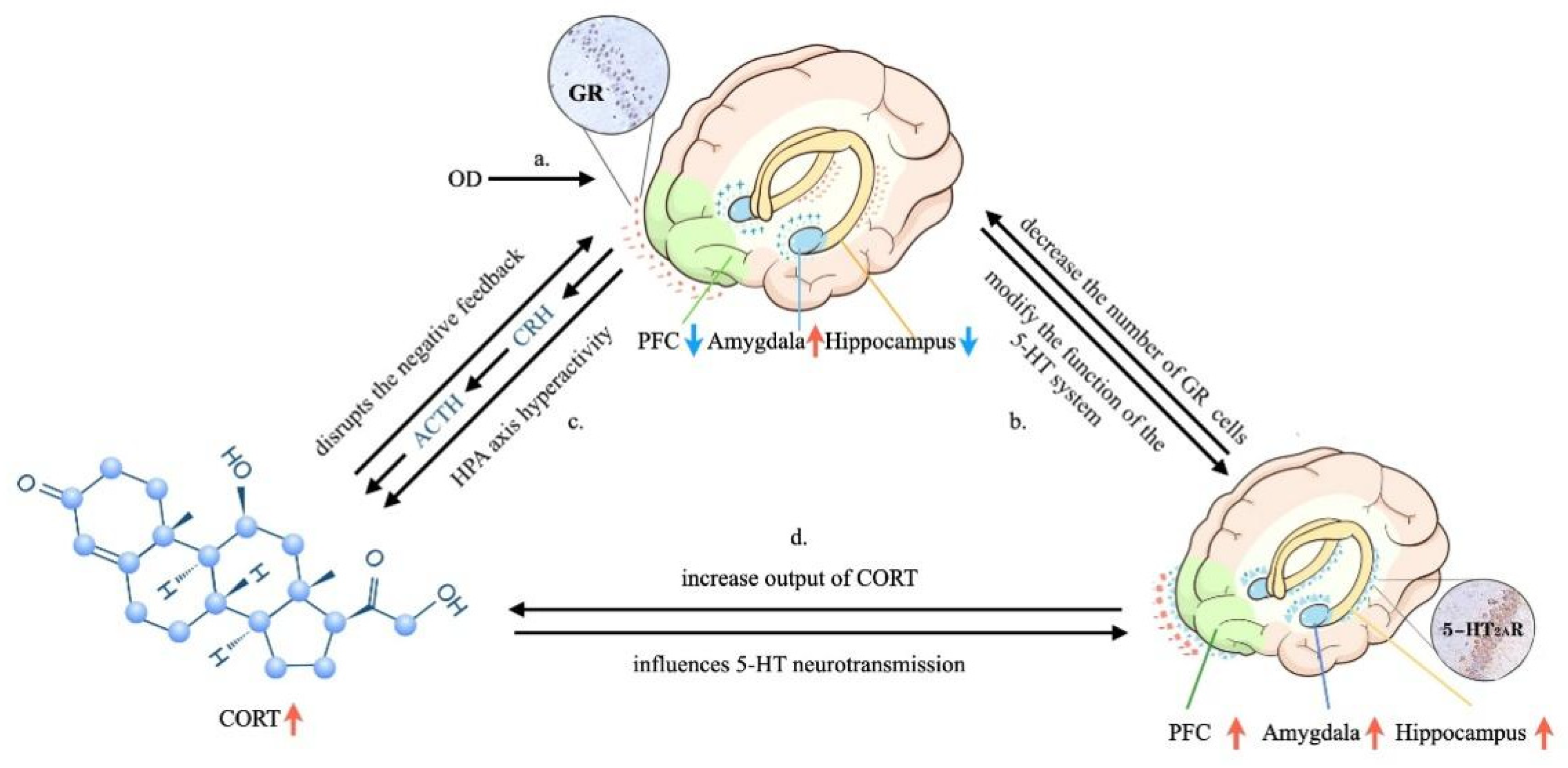
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Preparation
2.2. Anatomy of Molar Teeth in Sprague–Dawley Rats
2.3. Animal Model of Occlusal Disharmony (OD)
2.4. Animal Model of Chronic Unpredictable Stress (CUS)
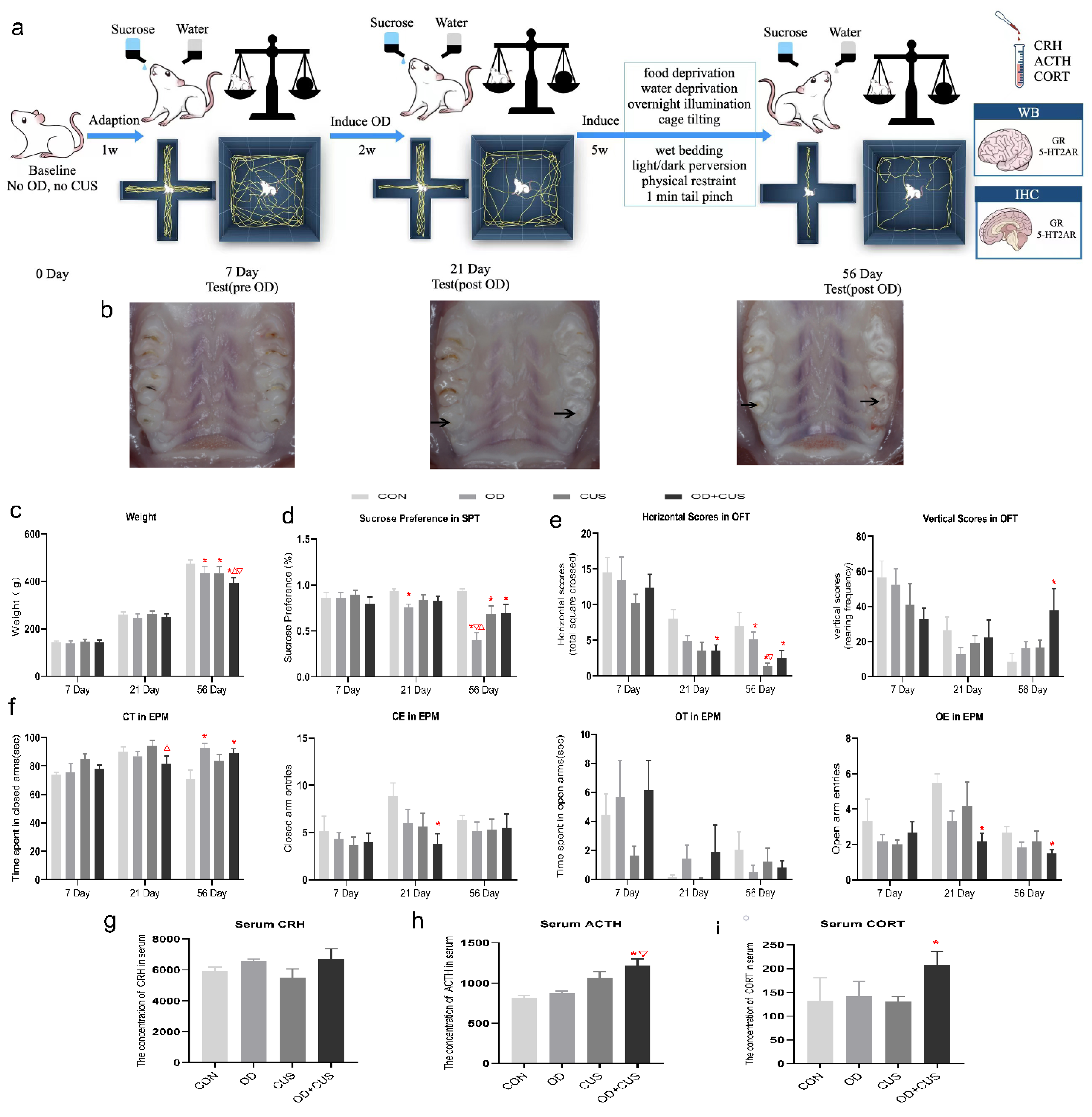
2.5. Experimental Design
2.6. Behavioral Tests
2.6.1. Weight Measurement
2.6.2. Elevated Plus Maze (EPM) Test
2.6.3. Open-Field Test (OFT)
2.6.4. Sucrose Preference Test
2.7. Enzyme-Linked Immunosorbent Assay (ELISA) Analysis
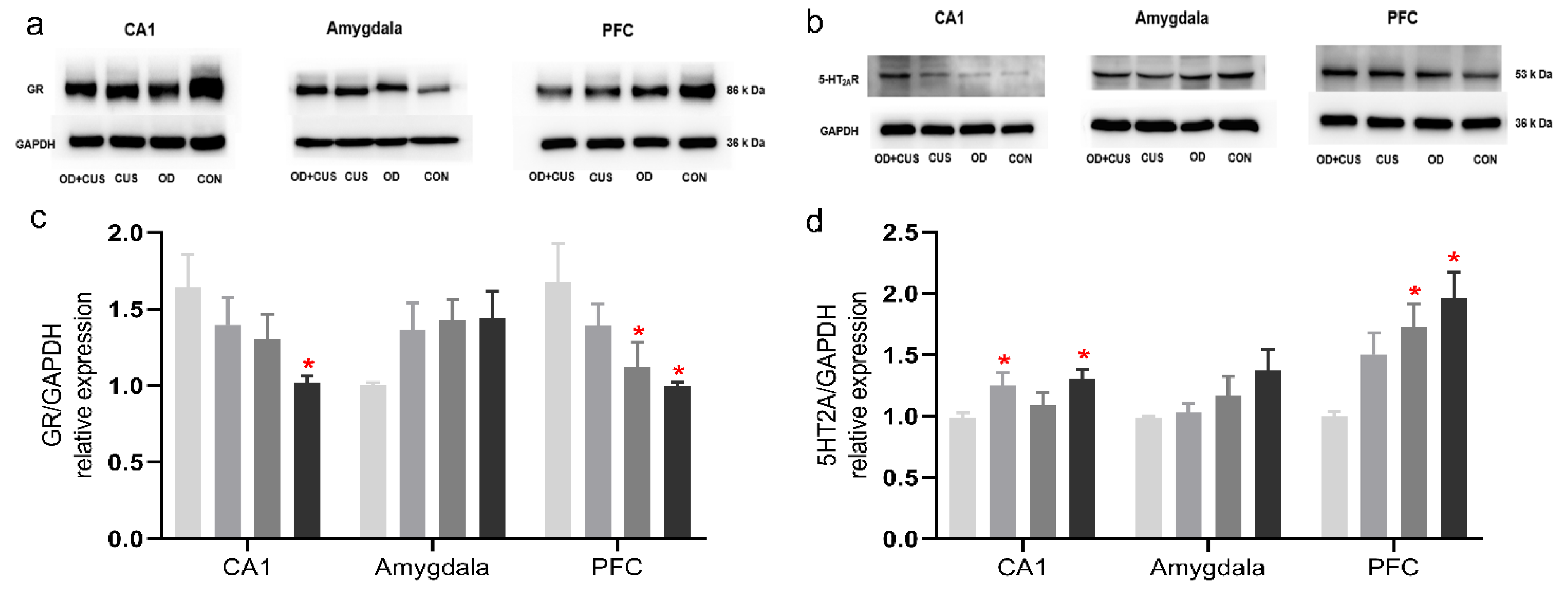
2.8. Western Blot of GR and 5-HT2AR Expression
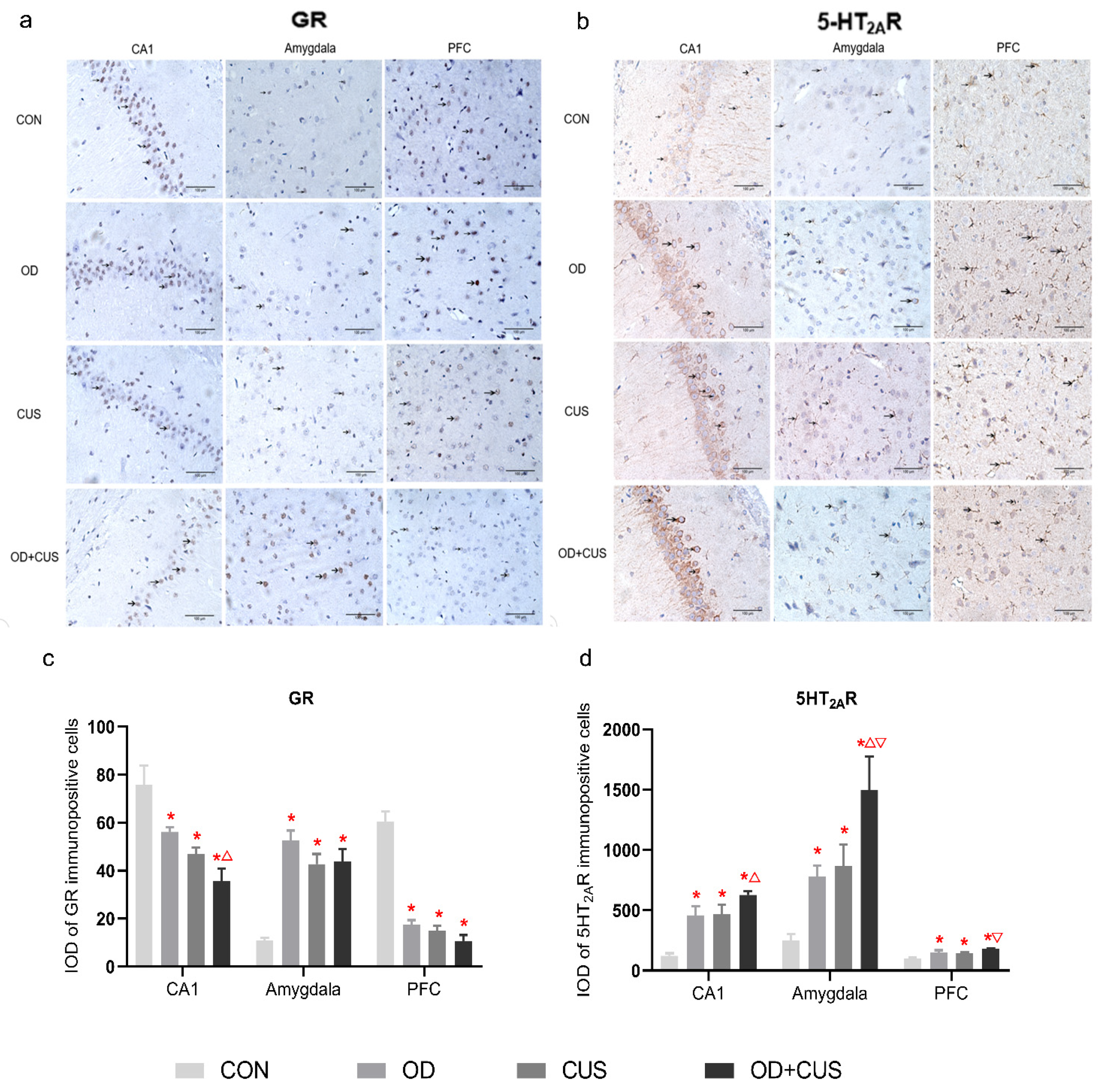
2.9. Immunohistochemistry of GR and 5-HT2AR Expression
2.10. Statistical Analysis
3. Results
3.1. Weight
3.2. Sucrose Preference Test
3.3. Open-Field Test
3.4. Elevated Plus Maze Test
3.5. Serum Concentrations of CRH, ACTH and CORT
3.6. Western Blot for GR and 5-HT2AR Expression
3.7. Immunohistochemistry of GR and 5-HT2AR Expression
4. Discussion
4.1. Design of the Experimental Procedure
4.2. Development of a New OD Animal Model
4.3. OD Is a Potential Factor Promoting Depression in a Rat Model
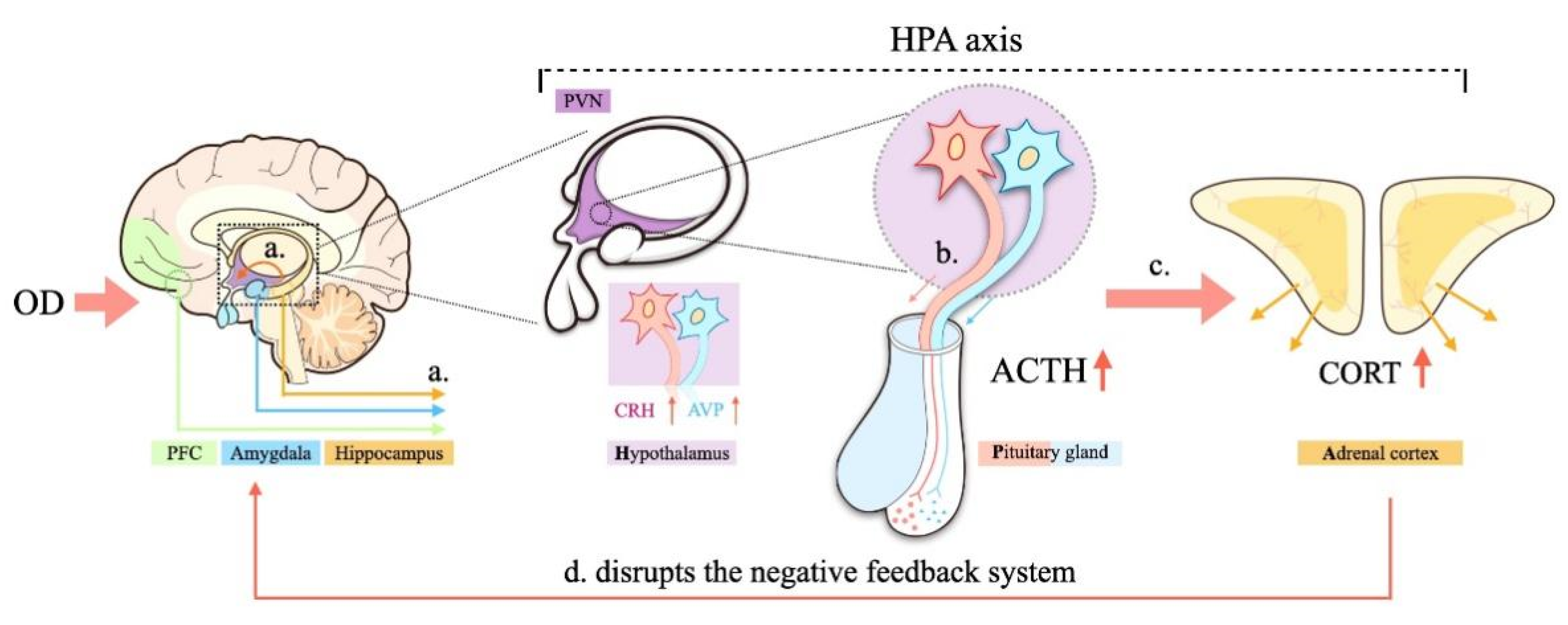
4.4. Activity of the Hypothalamus–Pituitary–Adrenal (HPA) Axis
4.5. Regulatory Mechanisms of the 5-HT System in Depression
4.6. Relationships between Depression-like Behavior and OD: Possible Mechanisms Underlying the Effects of OD
4.7. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Black, J.; Bond, M.A.; Hawkins, R.; Black, E. Test of a clinical model of poor physical health and suicide: The role of depression, psychosocial stress, interpersonal conflict, and panic. J. Affect. Disord. 2019, 257, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Braunsteiner, N.; Vickers, E.R.; Shparberg, R. Psychological Issues for Patients Undergoing Stem Cell Therapy and Regenerative Medicine. Open J. Regen. Med. 2018, 7, 1–17. [Google Scholar] [CrossRef][Green Version]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef]
- Klerman, G.L.; Weissman, M.M. The changing epidemiology of depression. Clin. Chem. 1988, 34, 807–812. [Google Scholar] [CrossRef]
- Che, Y.; Zhou, Z.; Shu, Y.; Zhai, C.; Zhu, Y.; Gong, S.; Cui, J.; Wang, J.-F. Chronic unpredictable stress impairs endogenous antioxidant defense in rat brain. Neurosci. Lett. 2015, 584, 208–213. [Google Scholar] [CrossRef]
- Katz, R.J. Animal model of depression: Pharmacological sensitivity of a hedonic deficit. Pharm. Biochem. Behav. 1982, 16, 965–968. [Google Scholar] [CrossRef]
- Belzung, C.; Willner, P.; Philippot, P. Depression: From psychopathology to pathophysiology. Curr. Opin. Neurobiol. 2015, 30, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. The chronic mild stress (CUS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. Validity, reliability and utility of the chronic mild stress model of depression: A 10-year review and evaluation. Psy-chopharmacology 1997, 134, 319–329. [Google Scholar] [CrossRef]
- Willner, P.; Mitchell, P.J. The validity of animal models of predisposition to depression. Behav. Pharmacol. 2002, 13, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. Reliability of the chronic mild stress model of depression: A user survey. Neurobiol. Stress 2017, 6, 68–77. [Google Scholar] [CrossRef]
- The Academy of prosthodontics. The Glossary of Prosthodontic Terms: Ninth Edition. J. Prosthet. Dent. 2017, 117, e1–e105. [Google Scholar] [CrossRef] [PubMed]
- Leon-Salazar, V.; Morrow, L.; Schiffman, E.L. Pain and persistent occlusal awareness: What should dentists do? J. Am. Dent. Assoc. 2012, 143, 989–991. [Google Scholar] [CrossRef]
- Narita, N.; Kamiya, K.; Makiyama, Y.; Iwaki, S.; Komiyama, O.; Ishii, T.; Wake, H. Prefrontal modulation during chewing performance in occlusal dysesthesia patients: A functional near-infrared spectroscopy study. Clin. Oral Investig. 2019, 23, 1181–1196. [Google Scholar] [CrossRef]
- Oguchi, H.; Yamauchi, Y.; Karube, Y.; Suzuki, N.; Tamaki, K. Occlusal Dysesthesia: A Clinical Report on the Psychosomatic Man-agement of a Japanese Patient Cohort. Int. J. Prosthodont. 2017, 30, 142–146. [Google Scholar] [CrossRef][Green Version]
- Hara, E.S.; Matsuka, Y.; Minakuchi, H.; Clark, G.T.; Kuboki, T. Occlusal dysesthesia: A qualitative systematic review of the epidemi-ology, aetiology and management. J. Oral Rehabil. 2012, 39, 630–638. [Google Scholar] [CrossRef]
- Tang, X.; Li, J.; Jiang, T.; Han, S.H.; Yao, D.Y. Experimental occlusal disharmony—A promoting factor for anxiety in rats under chronic psychological stress. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 75, 165–175. [Google Scholar] [CrossRef]
- Ekuni, D.; Tomofuji, T.; Irie, K.; Azuma, T.; Endo, Y.; Kasuyama, K.; Morita, M. Occlusal disharmony increases amyloid-beta in the rat hippocampus. NeuroMolecular Med. 2011, 13, 197–203. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, K.X.; Yin, N.N.; Zhang, C.K.; Shi, M.H.; Zhang, H.Y.; Wang, D.-M.; Xu, Z.-J.; Zhang, J.-D.; Li, J.-L.; et al. Malocclusion Generates Anxiety-Like Behavior Through a Putative Lateral Habenula-Mesencephalic Trigeminal Nucleus Pathway. Front. Mol. Neurosci. 2019, 12, 174. [Google Scholar] [CrossRef]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef]
- Rackers, H.S.; Thomas, S.; Williamson, K.; Posey, R.; Kimmel, M.C. Emerging literature in the Microbiota-Brain Axis and Perinatal Mood and Anxiety Disorders. Psychoneuroendocrinology 2018, 95, 86–96. [Google Scholar] [CrossRef]
- Zeni, A.L.B.; Camargo, A.; Dalmagro, A.P. Ferulic acid reverses depression-like behavior and oxidative stress induced by chronic corticosterone treatment in mice. Steroids 2017, 125, 131–136. [Google Scholar] [CrossRef]
- Artigas, F. Developments in the field of antidepressants, where do we go now? Eur. Neuropsychopharmacol. 2015, 25, 657–670. [Google Scholar] [CrossRef]
- Perkins, S.C.; Finegood, E.D.; Swain, J.E. Poverty and language development: Roles of parenting and stress. Innov. Clin. Neurosci. 2013, 10, 10–19. [Google Scholar]
- Azuma, K.; Zhou, Q.; Niwa, M.; Kubo, K.Y. Association between Mastication, the Hippocampus, and the HPA Axis: A Compre-hensive Review. Int. J. Mol. Sci. 2017, 18, 1687. [Google Scholar] [CrossRef] [PubMed]
- Miyake, H.; Mori, D.; Katayama, T.; Fujiwara, S.; Sato, Y.; Azuma, K.; Kubo, K.-Y. Novel stress increases hypothalamic-pituitary-adrenal activity in mice with a raised bite. Arch. Oral Biol. 2016, 68, 55–60. [Google Scholar] [CrossRef]
- Papp, M. Models of Affective Illness: Chronic Mild Stress in the Rat. Curr. Protoc. Pharm. 2012, 57, 5.9.1–5.9.11. [Google Scholar] [CrossRef]
- Ekuni, D.; Furuta, M.; Irie, K.; Azuma, T.; Tomofuji, T.; Murakami, T.; Yamashiro, T.; Ogura, T.; Morita, M. Relationship between impacts attributed to malocclusion and psychological stress in young Japanese adults. Eur. J. Orthod. 2011, 33, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Budtz-Jørgensen, E. Occlusal dysfunction and stress. An experimental study in macaque monkeys. J. Oral Rehabil. 2010, 8, 1–9. [Google Scholar] [CrossRef]
- Enkling, N.; Heussner, S.; Nicolay, C.; Bayer, S.; Mericske-Stern, R.; Utz, K.H. Tactile sensibility of single-tooth implants and natural teeth under local anesthesia of the natural antagonistic teeth. Clin. Implant. Dent. Relat. Res. 2012, 14, 273–280. [Google Scholar] [CrossRef]
- Nose-Ishibashi, K.; Watahiki, J.; Yamada, K.; Maekawa, M.; Watanabe, A.; Yamamoto, G.; Enamoto, A.; Nampo, M.T.; Taguchi, T.; Ichikawa, Y.; et al. Soft-diet feeding after weaning affects behavior in mice: Potential increase in vulnerability to mental disorders. Neuroscience 2014, 263, 257–268. [Google Scholar] [CrossRef]
- Moreau, J.; Jenck, F.; Martin, J.; Mortas, P.; Haefely, W.J.P. Effects of Moclobemide, a New Generation Reversible Mao-A Inhibitor, in a Novel Animal Model of Depression. Pharmacopsychiatry 1993, 26, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalam-ic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar] [PubMed]
- Gold, P.W.; Chrousos, G.P. Organization of the stress system and its dysregulation in melancholic and atypical depression: High vs low CRH/NE states. Mol. Psychiatry 2002, 7, 254–275. [Google Scholar] [CrossRef]
- Herman, J.P.; Figueiredo, H.; Mueller, N.K.; Ulrich-Lai, Y.; Ostrander, M.M.; Choi, D.C.; Cullinan, W.E. Central mechanisms of stress integration: Hierarchical circuitry controlling hypothalamo-pituitary-adrenocortical responsiveness. Front. Neuroendocrinol. 2003, 24, 151–180. [Google Scholar] [CrossRef]
- Popova, N.K.; Naumenko, V.S. Neuronal and behavioral plasticity: The role of serotonin and BDNF systems tandem. Expert Opin. Targets 2019, 23, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Artigas, F. Serotonin receptors involved in antidepressant effects. Pharm. Ther. 2013, 137, 119–131. [Google Scholar] [CrossRef]
- Bel, N.; Artigas, F. Chronic treatment with fluvoxamine increases extracellular serotonin in frontal cortex but not in raphe nu-clei. Synapse 1993, 15, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Yohn, C.N.; Gergues, M.M.; Samuels, B.A. The role of 5-HT receptors in depression. Mol. Brain 2017, 10, 28. [Google Scholar] [CrossRef]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Hongli, H.; Juan, Z.; Lin, J.; Xie, Y.; Xia, Y.; Lv, R.; Dong, L. Paeoniflorin improves menopause depression in ovariectomized rats under chronic unpredictable mild stress. Int. J. Clin. Exp. Med. 2015, 8, 5103–5111. [Google Scholar]
- Popova, N.K.; Ilchibaeva, T.V.; Antonov, E.V.; Pershina, A.V.; Bazovkina, D.V.; Naumenko, V.S. On the interaction between BDNF and serotonin systems: The effects of long-term ethanol consumption in mice. Alcohol 2020, 87, 1–15. [Google Scholar] [CrossRef]
- Mastitskaya, S.; Turovsky, E.; Marina, N.; Theparambil, S.M.; Hadjihambi, A.; Kasparov, S.; Teschemache, A.G.; Ramage, A.G.; Gourine, A.V.; Hosford, P.S. Astrocytes Modulate Baroreflex Sensitivity at the Level of the Nucleus of the Solitary Tract. J. Neurosci. 2020, 40, 3052–3062. [Google Scholar] [CrossRef] [PubMed]
- Kurhe, Y.; Mahesh, R.; Devadoss, T. QCM-4, a 5-HT(3) receptor antagonist ameliorates plasma HPA axis hyperactivity, leptin resistance and brain oxidative stress in depression and anxiety-like behavior in obese mice. Biochem. Biophys. Res. Commun. 2015, 456, 74–79. [Google Scholar] [CrossRef]
- Blier, P.; de Montigny, C. Current advances and trends in the treatment of depression. Trends Pharm. Sci. 1994, 15, 220–226. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Popova, N.K.; Lacivita, E.; Leopoldo, M.; Ponimaskin, E.G. Interplay between serotonin 5-HT1A and 5-HT7 receptors in depressive disorders. CNS Neurosci. Ther. 2014, 20, 582–590. [Google Scholar] [CrossRef]
- Cavalcante, A.L.; Siqueira, R.M.; Araujo, J.C.; Gondim, D.V.; Ribeiro, R.A.; Quetz, J.S.; Havt, A.; Lima, A.A.M.; Vale, M.L. Role of NMDA receptors in the trigeminal pathway, and the modulatory effect of magnesium in a model of rat temporomandibular joint arthritis. Eur. J. Oral Sci. 2013, 121, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Renner, U.; Zeug, A.; Woehler, A.; Niebert, M.; Dityatev, A.; Dityateva, G.; Gorinski, N.; Guseva, D.; Abdel-Galil, D.; Fröhlich, M.; et al. Heterodimerization of serotonin receptors 5-HT1A and 5-HT7 differentially regulates receptor signalling and trafficking. J. Cell Sci. 2012, 125, 2486–2499. [Google Scholar] [CrossRef]
- Ono, Y.; Yamamoto, T.; Kubo, K.Y.; Onozuka, M. Occlusion and brain function: Mastication as a prevention of cognitive dysfunction. J. Oral Rehabil. 2010, 37, 624–640. [Google Scholar] [CrossRef]
- Oh, D.R.; Yoo, J.S.; Kim, Y.; Kang, H.; Lee, H.; Lm, S.J.; Choi, E.-J.; Jung, M.-A.; Bae, D.; Oh, K.-N.; et al. Vaccinium bracteatum Leaf Extract Reverses Chronic Restraint Stress-Induced Depression-Like Behavior in Mice: Regulation of Hypothalamic-Pituitary-Adrenal Axis, Serotonin Turnover Systems, and ERK/Akt Phosphorylation. Front. Pharmacol. 2018, 9, 604. [Google Scholar] [CrossRef]
- Piancino, M.G.; Tortarolo, A.; Polimeni, A.; Cannavale, R.; Tonni, I.; Deregibus, A. Adverse effects of the bite-raised condition in animal studies: A systematic review. Arch. Oral Biol. 2019, 107, 104516. [Google Scholar] [CrossRef] [PubMed]
- Porter, R.J.; Gallagher, P.; Watson, S.; Young, A.H. Corticosteroid-serotonin interactions in depression: A review of the human evidence. Psychopharmacology 2004, 173, 1–17. [Google Scholar] [CrossRef]
- McEuen, J.G.; Beck, S.G.; Bale, T.L. Failure to mount adaptive responses to stress results in dysregulation and cell death in the midbrain raphe. J. Neurosci. 2008, 28, 8169–8177. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; Tasker, J.G. Paraventricular Hypothalamic Mechanisms of Chronic Stress Adaptation. Front. Endocrinol. 2016, 7, 137. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Wu, L.; Zhang, M.; He, K.; Wang, X.; Lin, Y.; Li, S.; Chen, J. Occlusal Disharmony—A Potential Factor Promoting Depression in a Rat Model. Brain Sci. 2022, 12, 747. https://doi.org/10.3390/brainsci12060747
Zhang S, Wu L, Zhang M, He K, Wang X, Lin Y, Li S, Chen J. Occlusal Disharmony—A Potential Factor Promoting Depression in a Rat Model. Brain Sciences. 2022; 12(6):747. https://doi.org/10.3390/brainsci12060747
Chicago/Turabian StyleZhang, Sihui, Ling Wu, Mi Zhang, Kaixun He, Xudong Wang, Yuxuan Lin, Shuxian Li, and Jiang Chen. 2022. "Occlusal Disharmony—A Potential Factor Promoting Depression in a Rat Model" Brain Sciences 12, no. 6: 747. https://doi.org/10.3390/brainsci12060747
APA StyleZhang, S., Wu, L., Zhang, M., He, K., Wang, X., Lin, Y., Li, S., & Chen, J. (2022). Occlusal Disharmony—A Potential Factor Promoting Depression in a Rat Model. Brain Sciences, 12(6), 747. https://doi.org/10.3390/brainsci12060747