
Resting-State Neuronal Activity and Functional Connectivity Changes in the Visual Cortex after High Altitude Exposure: A Longitudinal Study

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Participants
2.2. Routine Blood and Behavior Tests
2.3. MRI Data Acquisition
2.4. Resting-State fMRI Data Preprocessing
2.5. Analyses of Resting-State fMRI
2.6. Statistical Analysis
3. Results
3.1. Routine Blood Results
3.2. Behavior Results
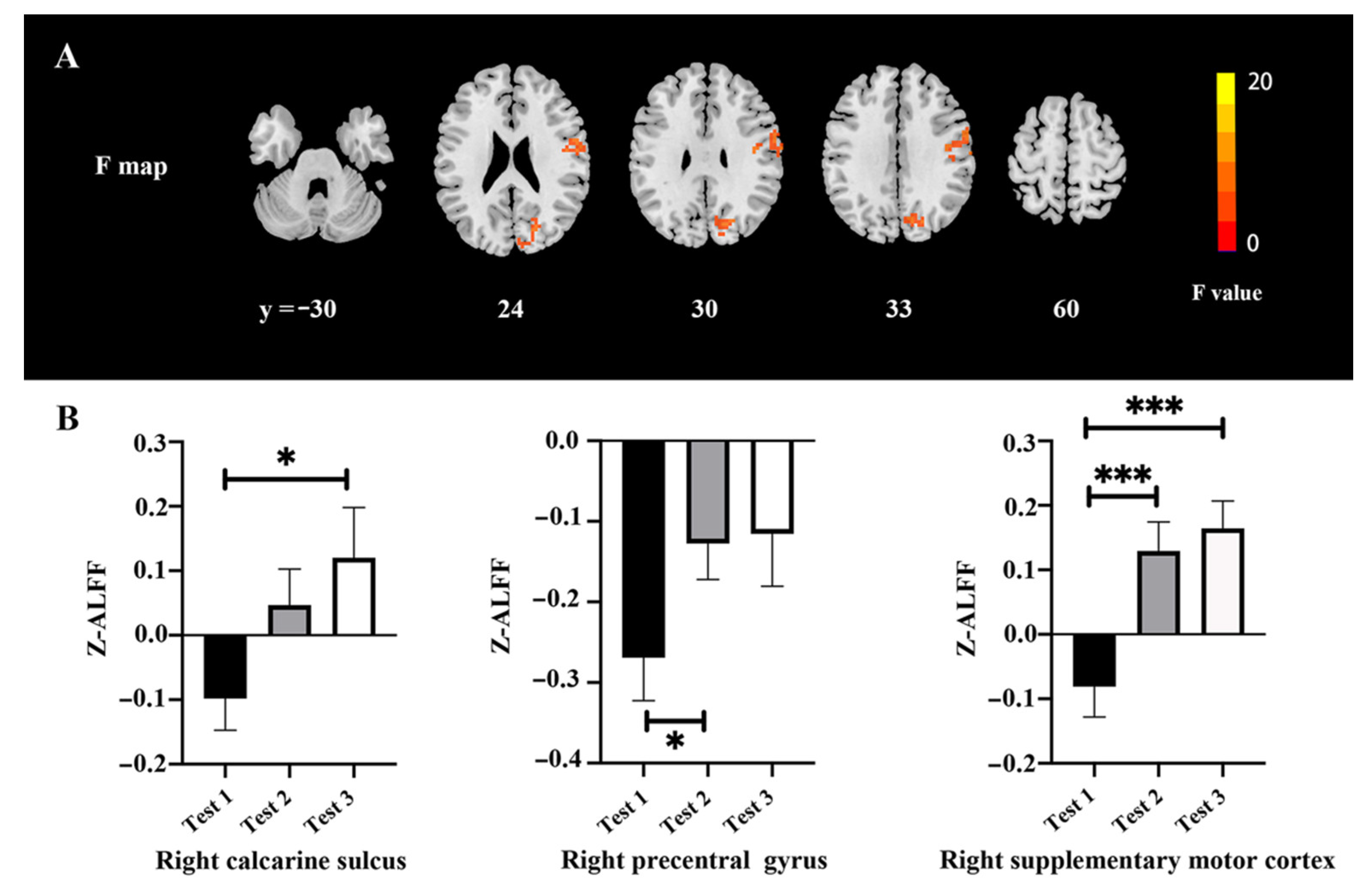
3.3. ALFF Values
3.4. ReHo Values
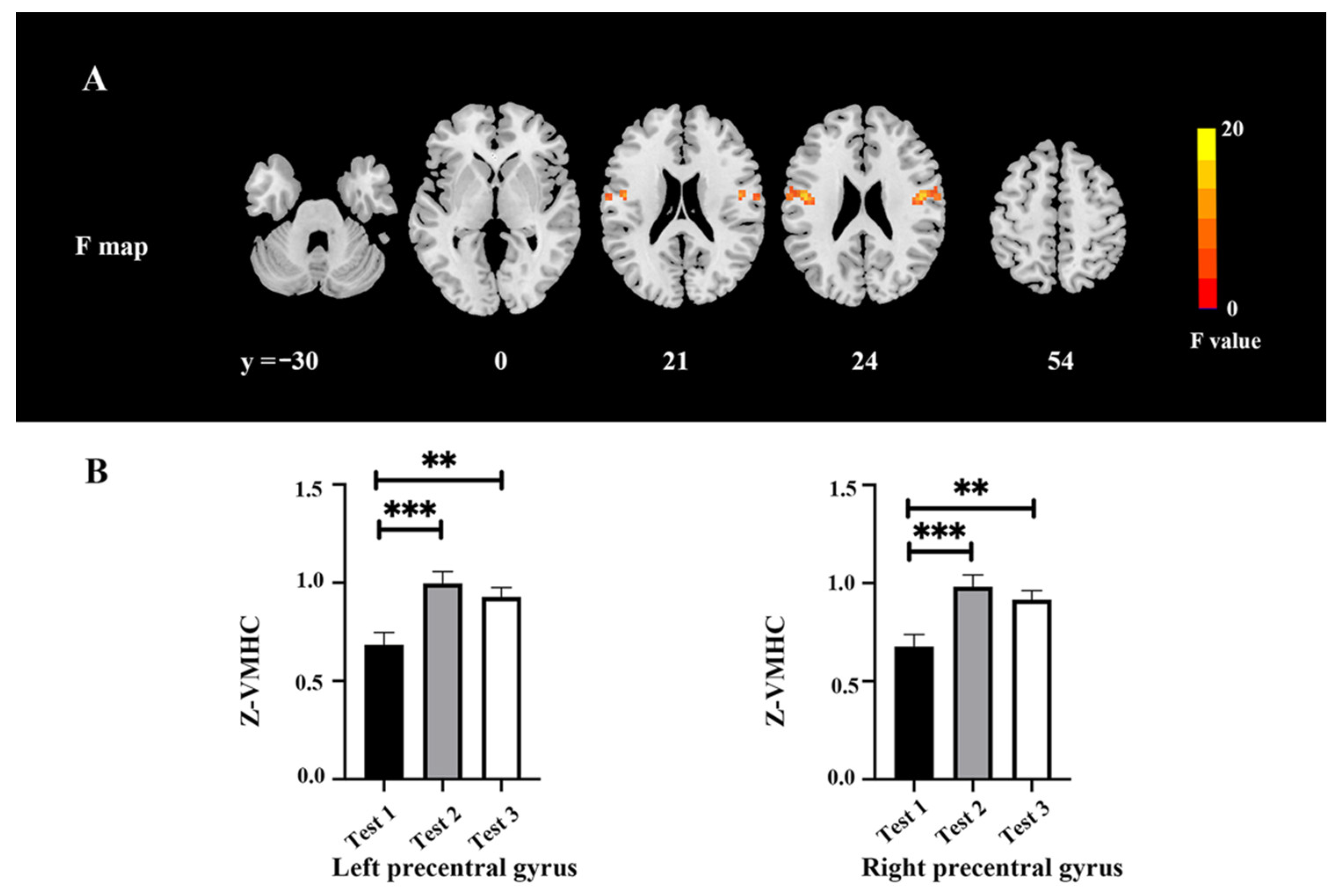
3.5. VMHC Values
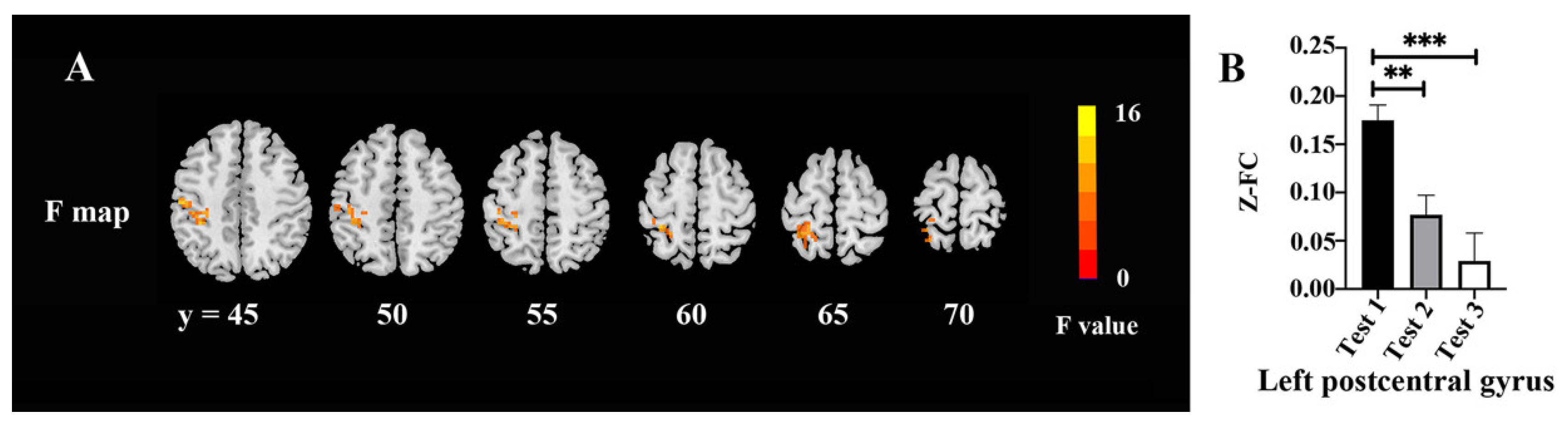
3.6. FC Values
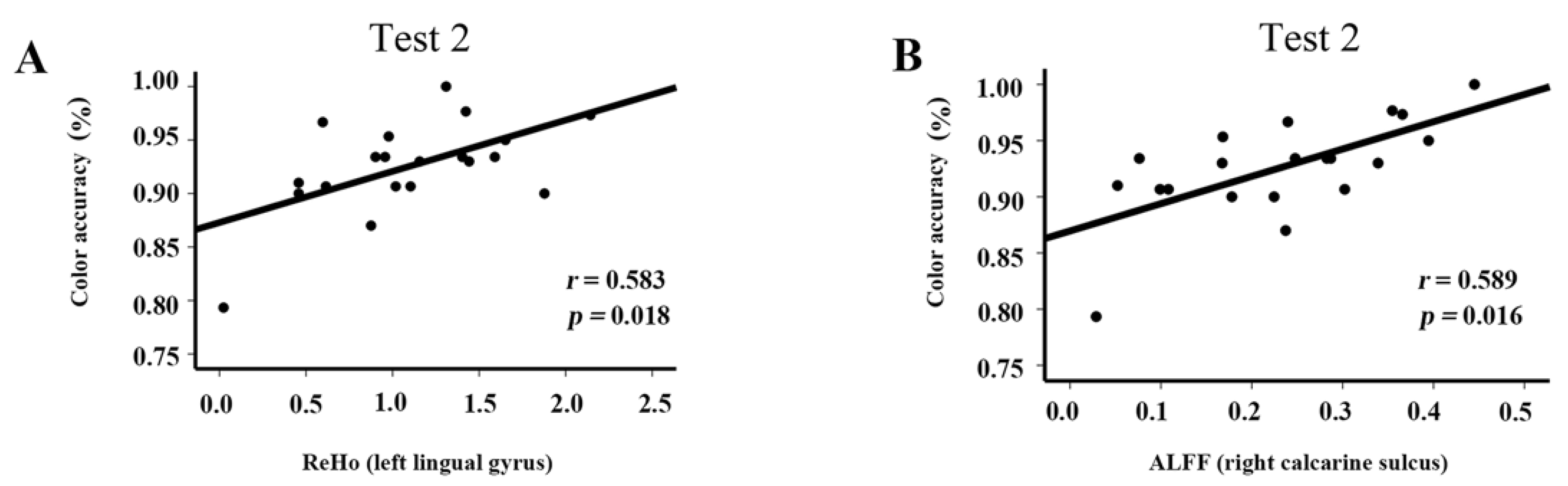
3.7. Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tianyi, W.U.; Kayser, B. High Altitude Adaptation in Tibetans. High Alt. Med. Biol. 2006, 7, 193. [Google Scholar]
- Mujika, I.; Sharma, A.P.; Stellingwerff, T. Contemporary Periodization of Altitude Training for Elite Endurance Athletes: A Narrative Review. Sports Med. 2019, 49, 1651–1669. [Google Scholar] [CrossRef]
- Khodaee, M.; Grothe, H.L.; Seyfert, J.H.; VanBaak, K. Athletes at High Altitude. Sports Health 2016, 8, 126–132. [Google Scholar] [CrossRef] [Green Version]
- Durand, S.; Iyer, R.; Mizuseki, K.; de Vries, S.; Mihalas, S.; Reid, R.C. A Comparison of Visual Response Properties in the Lateral Geniculate Nucleus and Primary Visual Cortex of Awake and Anesthetized Mice. J. Neurosci. 2016, 36, 12144–12156. [Google Scholar] [CrossRef] [Green Version]
- Karakucuk, S.; Mirza, G.E. Ophthalmological effects of high altitude. Ophthalmic Res. 2000, 32, 30–40. [Google Scholar] [CrossRef]
- Wang, Z.X.; Zhang, D.L.; Ma, H.L. The effect of high altitude on human color perception. Sheng Li Xue Bao 2019, 71, 833–838. [Google Scholar]
- Shrestha, A.; Suwal, R.; Shrestha, B. Vitreous Hemorrhage following High-Altitude Retinopathy. Case Rep. Ophthalmol. Med. 2021, 2021, 7076190. [Google Scholar] [CrossRef]
- Gekeler, K.; Schatz, A.; Fischer, M.D.; Schommer, K.; Boden, K.; Bartz-Schmidt, K.U.; Gekeler, F.; Willmann, G. Decreased contrast sensitivity at high altitude. Br. J. Ophthalmol. 2019, 103, 1815–1819. [Google Scholar] [CrossRef]
- Zhang, Q.F.; Li, H.; Chen, M.; Guo, A.; Wen, Y.; Poo, M.M. Functional organization of intrinsic and feedback presynaptic inputs in the primary visual cortex. Proc. Natl. Acad. Sci. USA 2018, 115, E5174–E5182. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.C.; Fahle, M.; Mulder, T.; Trauzettel-Klosinski, S. Adaptation, perceptual learning, and plasticity of brain functions. Graefes Arch. Clin. Exp. Ophthalmol. 2017, 255, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Castaldi, E.; Lunghi, C.; Morrone, M.C. Neuroplasticity in adult human visual cortex. Neurosci. Biobehav. Rev. 2020, 112, 542–552. [Google Scholar] [CrossRef]
- Zhang, J.; Yan, X.; Shi, J.; Gong, Q.; Weng, X.; Liu, Y. Structural modifications of the brain in acclimatization to high-altitude. PLoS ONE 2010, 5, e11449. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Zhang, J.; Gong, Q.; Weng, X. Cerebrovascular reactivity among native-raised high altitude residents: An fMRI study. BMC Neurosci. 2011, 12, 94. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wei, W.; Yuan, F.; Li, S.; Lin, J.; Zhang, J. Regional cerebral blood flow in natives at high altitude: An arterial spin labeled MRI study. J. Magn. Reson. Imaging 2018, 48, 708–717. [Google Scholar] [CrossRef]
- Wei, W.; Wang, X.; Gong, Q.; Fan, M.; Zhang, J. Cortical Thickness of Native Tibetans in the Qinghai-Tibetan Plateau. AJNR Am. J. Neuroradiol. 2017, 38, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, J.; Han, Q.; Lin, J.; Yang, T.; Chen, Z.; Zhang, J. Long-term acclimatization to high-altitude hypoxia modifies interhemispheric functional and structural connectivity in the adult brain. Brain Behav. 2016, 6, e00512. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Fan, C.; Li, J.; Lin, J.; Yang, T.; Fan, M. Alteration of Spontaneous Brain Activity After Hypoxia-Reoxygenation: A Resting-State fMRI Study. High Alt. Med. Biol. 2017, 18, 20–26. [Google Scholar] [CrossRef]
- Zhao, J.P.; Zhang, R.; Yu, Q.; Zhang, J.X. Characteristics of EEG activity during high altitude hypoxia and lowland reoxygenation. Brain Res. 2016, 1648, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Zhao, Y.; Yu, Q.; Yin, W.; Liu, H.; Lin, J.; Yang, T.; Fan, M.; Gesang, L.; Zhang, J. Reversible Brain Abnormalities in People Without Signs of Mountain Sickness During High-Altitude Exposure. Sci. Rep. 2016, 6, 33596. [Google Scholar] [CrossRef] [Green Version]
- Kühn, S.; Gerlach, D.; Noblé, H.J.; Weber, F.; Rittweger, J.; Jordan, J.; Limper, U. An Observational Cerebral Magnetic Resonance Imaging Study Following 7 Days at 4554 m. High Alt. Med. Biol. 2019, 20, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Yu-Feng, Z.; Yong, H.; Chao-Zhe, Z.; Qing-Jiu, C.; Man-Qiu, S.; Meng, L.; Li-Xia, T.; Tian-Zi, J.; Yu-Feng, W. Altered baseline brain activity in children with ADHD revealed by resting-state functional MRI. Brain Dev. 2007, 29, 83–91. [Google Scholar] [CrossRef]
- Zang, Y.; Jiang, T.; Lu, Y.; He, Y.; Tian, L. Regional homogeneity approach to fMRI data analysis. Neuroimage 2004, 22, 394–400. [Google Scholar] [CrossRef]
- Zuo, X.N.; Kelly, C.; Di Martino, A.; Mennes, M.; Margulies, D.S.; Bangaru, S.; Grzadzinski, R.; Evans, A.C.; Zang, Y.F.; Castellanos, F.X.; et al. Growing together and growing apart: Regional and sex differences in the lifespan developmental trajectories of functional homotopy. J. Neurosci. 2010, 30, 15034–15043. [Google Scholar] [CrossRef]
- Biswal, B.; Yetkin, F.Z.; Haughton, V.M.; Hyde, J.S. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn. Reson. Med. 1995, 34, 537–541. [Google Scholar] [CrossRef]
- Petrassi, F.A.; Hodkinson, P.D.; Walters, P.L.; Gaydos, S.J. Hypoxic hypoxia at moderate altitudes: Review of the state of the science. Aviat. Space Environ. Med. 2012, 83, 975–984. [Google Scholar] [CrossRef]
- Zubieta-Calleja, G.R.; Paulev, P.E.; Zubieta-Calleja, L.; Zubieta-Castillo, G. Altitude adaptation through hematocrit changes. J. Physiol. Pharmacol. 2007, 58 (Suppl. 5), 811–818. [Google Scholar]
- Qiu, Q.; Lv, P.; Zhongshen, Y.; Yuan, F.; Zhang, X.; Zhou, X.; Li, S.; Liu, X.; Zhang, J. Electrophysiological mechanisms underlying hypoxia-induced deficits in visual spatial and non-spatial discrimination. Physiol. Rep. 2021, 9, e15036. [Google Scholar] [CrossRef]
- Chao-Gan, Y.; Yu-Feng, Z. DPARSF: A MATLAB Toolbox for “Pipeline” Data Analysis of Resting-State fMRI. Front. Syst. Neurosci. 2010, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.G.; Wang, X.D.; Zuo, X.N.; Zang, Y.F. DPABI: Data Processing & Analysis for (Resting-State) Brain Imaging. Neuroinformatics 2016, 14, 339–351. [Google Scholar] [CrossRef]
- Chen, X.; Lu, B.; Yan, C.G. Reproducibility of R-fMRI metrics on the impact of different strategies for multiple comparison correction and sample sizes. Hum. Brain Mapp. 2018, 39, 300–318. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, M.; Mairbäurl, H.; Livshits, L.; Seide, S.; Hackbusch, M.; Malczyk, M.; Kraut, S.; Gassmann, N.N.; Weissmann, N.; Muckenthaler, M.U. The increase in hemoglobin concentration with altitude varies among human populations. Ann. N. Y. Acad. Sci. 2019, 1450, 204–220. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, M.; Muckenthaler, M.U. Adaptation of iron requirement to hypoxic conditions at high altitude. J. Appl. Physiol. 2015, 119, 1432–1440. [Google Scholar] [CrossRef] [Green Version]
- León-Velarde, F.; Gamboa, A.; Chuquiza, J.A.; Esteba, W.A.; Rivera-Chira, M.; Monge, C.C. Hematological parameters in high altitude residents living at 4355, 4660, and 5500 meters above sea level. High Alt. Med. Biol. 2000, 1, 97–104. [Google Scholar] [CrossRef]
- Miyamoto, O.; Sumitani, K.; Takahashi, M.; Hirakawa, H.; Kusakabe, T.; Hayashida, Y.; Itano, T. Vascular changes in the rat brain during chronic hypoxia in the presence and absence of hypercapnia. Acta Med. Okayama 2005, 59, 135–143. [Google Scholar] [CrossRef]
- Harik, N.; Harik, S.I.; Kuo, N.T.; Sakai, K.; Przybylski, R.J.; LaManna, J.C. Time-course and reversibility of the hypoxia-induced alterations in cerebral vascularity and cerebral capillary glucose transporter density. Brain Res. 1996, 737, 335–338. [Google Scholar] [CrossRef]
- Benderro, G.F.; LaManna, J.C. HIF-1α/COX-2 expression and mouse brain capillary remodeling during prolonged moderate hypoxia and subsequent re-oxygenation. Brain Res. 2014, 1569, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Vigneau-Roy, N.; Bernier, M.; Descoteaux, M.; Whittingstall, K. Regional variations in vascular density correlate with resting-state and task-evoked blood oxygen level-dependent signal amplitude. Hum. Brain Mapp. 2014, 35, 1906–1920. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Li, J.; Chen, J.; Han, Q.; Lin, J.; Yang, T.; Fan, M. Adaptive modulation of adult brain gray and white matter to high altitude: Structural MRI studies. PLoS ONE 2013, 8, e68621. [Google Scholar] [CrossRef]
- Whittington, M.A.; Traub, R.D.; Faulkner, H.J.; Stanford, I.M.; Jefferys, J.G. Recurrent excitatory postsynaptic potentials induced by synchronized fast cortical oscillations. Proc. Natl. Acad. Sci. USA 1997, 94, 12198–12203. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Z.Q.; Huang, Y.L.; Luo, G.L.; Ma, H.L.; Zhang, D.L. Decreased Event-Related Desynchronization of Mental Rotation Tasks in Young Tibetan Immigrants. Front. Hum. Neurosci. 2021, 15, 664039. [Google Scholar] [CrossRef]
- Li, Z.; Xue, X.; Li, X.; Bao, X.; Yu, S.; Wang, Z.; Liu, M.; Ma, H.; Zhang, D. Neuropsychological effect of working memory capacity on mental rotation under hypoxia environment. Int. J. Psychophysiol. 2021, 165, 18–28. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Q.; Wang, J.; Liu, J.; Zhang, W.; Qi, S.; Xu, H.; Li, C.; Zhang, J.; Zhao, H.; et al. Cognitive and neuroimaging changes in healthy immigrants upon relocation to a high altitude: A panel study. Hum. Brain Mapp. 2017, 38, 3865–3877. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Fan, C.; Li, J.; Han, Q.; Lin, J.; Yang, T.; Zhang, J. Increased Intraregional Synchronized Neural Activity in Adult Brain After Prolonged Adaptation to High-Altitude Hypoxia: A Resting-State fMRI Study. High Alt. Med. Biol. 2016, 17, 16–24. [Google Scholar] [CrossRef]
- Essen, D.C.; Zeki, S.M. The topographic organization of rhesus monkey prestriate cortex. J. Physiol. 1978, 277, 193–226. [Google Scholar] [CrossRef]
- Weissman, D.H.; Woldorff, M.G.; Hazlett, C.J.; Mangun, G.R. Effects of practice on executive control investigated with fMRI. Brain Res. Cogn. Brain Res. 2002, 15, 47–60. [Google Scholar] [CrossRef]
- Liu, X.; Yang, X.; Hou, Z.; Ma, M.; Jiang, W.; Wang, C.; Zhang, Y.; Yuan, Y. Increased interhemispheric synchrony underlying the improved athletic performance of rowing athletes by transcranial direct current stimulation. Brain Imaging Behav. 2019, 13, 1324–1332. [Google Scholar] [CrossRef]
- Tanaka, S.; Kirino, E. Dynamic Reconfiguration of the Supplementary Motor Area Network during Imagined Music Performance. Front. Hum. Neurosci. 2017, 11, 606. [Google Scholar] [CrossRef] [Green Version]
- Daly, C.J.; Kelley, G.T.; Krauss, A. Relationship between visual-motor integration and handwriting skills of children in kindergarten: A modified replication study. Am. J. Occup. Ther. 2003, 57, 459–462. [Google Scholar] [CrossRef] [Green Version]
- Kruse, W.; Dannenberg, S.; Kleiser, R.; Hoffmann, K.P. Temporal relation of population activity in visual areas MT/MST and in primary motor cortex during visually guided tracking movements. Cereb. Cortex 2002, 12, 466–476. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Jiang, T.; Yu, C.; Tian, L.; Li, J.; Liu, Y.; Zhou, Y.; Xu, L.; Song, M.; Li, K. Spontaneous activity associated with primary visual cortex: A resting-state FMRI study. Cereb. Cortex 2008, 18, 697–704. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Zhou, F.Q.; Huang, X.; Dan, H.D.; Xie, B.J.; Shen, Y. Altered functional connectivity of primary visual cortex in late blindness. Neuropsychiatr. Dis. Treat. 2018, 14, 3317–3327. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Su, J.; He, X.W.; Ban, S.; Zhu, Q.; Cui, Y.; Zhang, J.; Hu, Y.; Liu, Y.S.; Zhao, R.; et al. Disrupted functional connectivity between sub-regions in the sensorimotor areas and cortex in migraine without aura. J. Headache Pain 2020, 21, 47. [Google Scholar] [CrossRef]

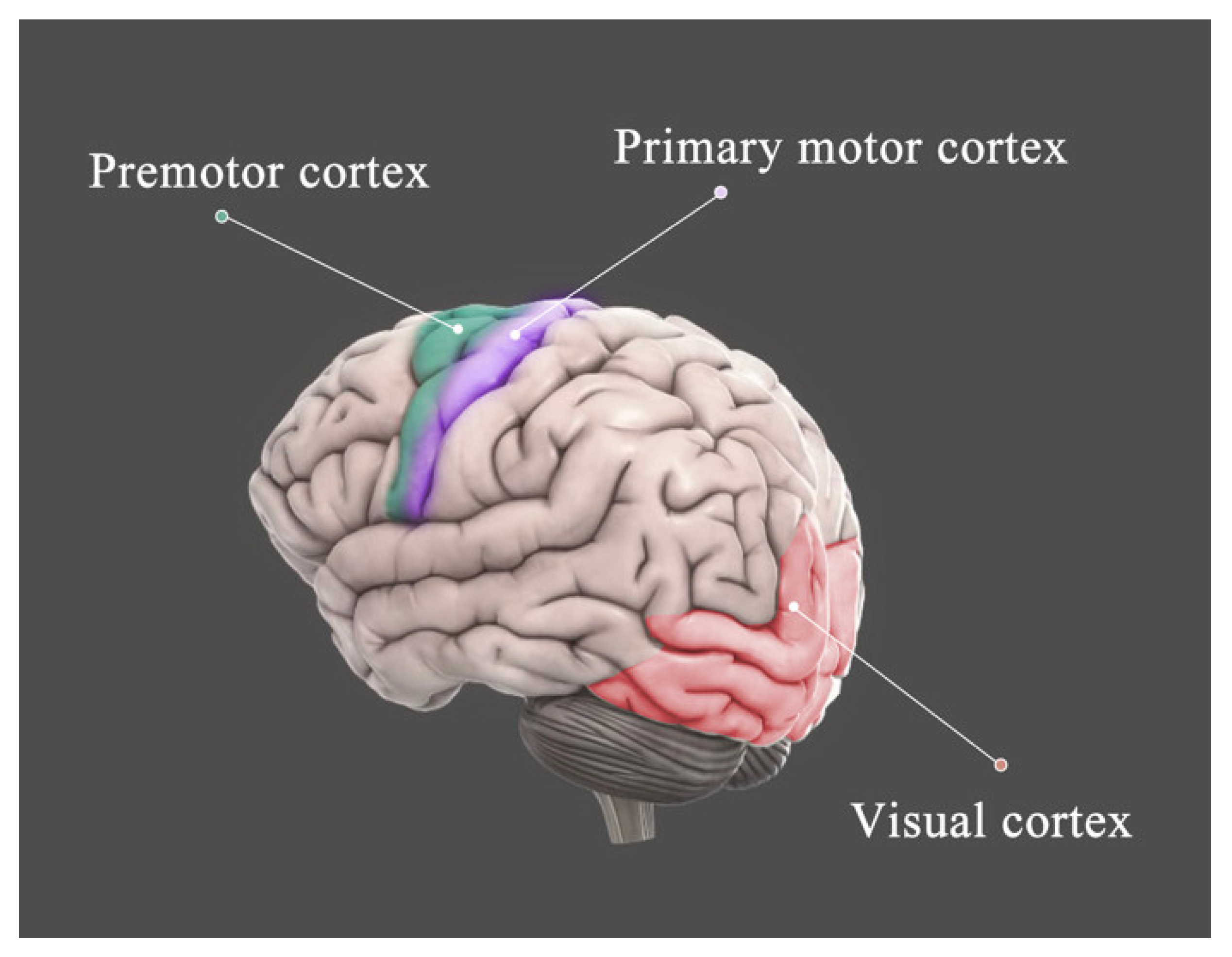

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test 1 | Test 2 | Test 3 | F | p | |
|---|---|---|---|---|---|
| Hematocrit (%) | 40.69 ± 3.17 | 42.87 ± 4.62 | 39.59 ± 3.26 | 13.77 | <0.0001 |
| Hemoglobin (g/L) | 133.10 ± 14.03 | 141.95 ± 18.22 | 132.20 ± 13.78 | 14.92 | <0.0001 |
| Red blood cells count (1012/L) | 4.77 ± 0.50 | 5.02 ± 0.75 | 4.65 ± 0.70 | 16.50 | <0.0001 |
| Angle ACC (%) | Color ACC (%) | Angle RT (ms) | Color RT (ms) | |
|---|---|---|---|---|
| Test 1 | 90.86 ± 5.25 | 94.71 ± 3.14 | 575.35 ± 96.63 | 527.04 ± 81.88 |
| Test 2 | 86.77 ± 4.36 | 92.59 ± 4.46 | 618.52 ± 85.82 | 549.92 ± 92.06 |
| Test 3 | 89.66 ± 6.50 | 92.99 ± 3.58 | 562.87 ± 84.23 | 522.16 ± 98.89 |
| Area | Broadmann Area | Volume (Voxels) | Talairach | F (Peak) | ||
|---|---|---|---|---|---|---|
| X | Y | Z | ||||
| Occipital/Temporal/Calcarine_R/Lingual_R/Precuneus | 17/18/19/37 | 920 | 48 | −57 | 6 | 20.03 |
| Precentral_R | 6/4 | 260 | 60 | 3 | 36 | 11.24 |
| Supp_Motor_Area_R | 6 | 168 | 6 | 3 | 78 | 12.44 |
| Area | Broadmann Area | Volume (Voxels) | Talairach | F (Peak) | ||
|---|---|---|---|---|---|---|
| X | Y | Z | ||||
| Occipital/Lingual_L/Fusiform_L/Calcarine_L | 17/18/19 | 179 | −24 | −93 | 6 | 19.93 |
| Area | Broadmann Area | Volume (Voxels) | Talairach | F (Peak) | ||
|---|---|---|---|---|---|---|
| X | Y | Z | ||||
| Precentral_L/Postcentral_L | 6/4 | 93 | −48 | −46 | 24 | 16.73 |
| Precentral_R/Postcentral_R | 6/4 | 104 | 48 | −46 | 24 | 16.73 |
| Area | Broadmann Area | Volume (Voxels) | Talairach | F (Peak) | ||
|---|---|---|---|---|---|---|
| X | Y | Z | ||||
| Postcentral_L | 6 | 174 | −51 | −18 | 45 | 14.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Kang, T.; Liu, Y.; Yuan, F.; Li, M.; Lin, J.; Zhang, J. Resting-State Neuronal Activity and Functional Connectivity Changes in the Visual Cortex after High Altitude Exposure: A Longitudinal Study. Brain Sci. 2022, 12, 724. https://doi.org/10.3390/brainsci12060724
Zhang X, Kang T, Liu Y, Yuan F, Li M, Lin J, Zhang J. Resting-State Neuronal Activity and Functional Connectivity Changes in the Visual Cortex after High Altitude Exposure: A Longitudinal Study. Brain Sciences. 2022; 12(6):724. https://doi.org/10.3390/brainsci12060724
Chicago/Turabian StyleZhang, Xinjuan, Taishan Kang, Yanqiu Liu, Fengjuan Yuan, Minglu Li, Jianzhong Lin, and Jiaxing Zhang. 2022. "Resting-State Neuronal Activity and Functional Connectivity Changes in the Visual Cortex after High Altitude Exposure: A Longitudinal Study" Brain Sciences 12, no. 6: 724. https://doi.org/10.3390/brainsci12060724
APA StyleZhang, X., Kang, T., Liu, Y., Yuan, F., Li, M., Lin, J., & Zhang, J. (2022). Resting-State Neuronal Activity and Functional Connectivity Changes in the Visual Cortex after High Altitude Exposure: A Longitudinal Study. Brain Sciences, 12(6), 724. https://doi.org/10.3390/brainsci12060724