
Functional Neuroimaging of Human Hypothalamus in Socioemotional Behavior: A Systematic Review


Abstract
1. Introduction
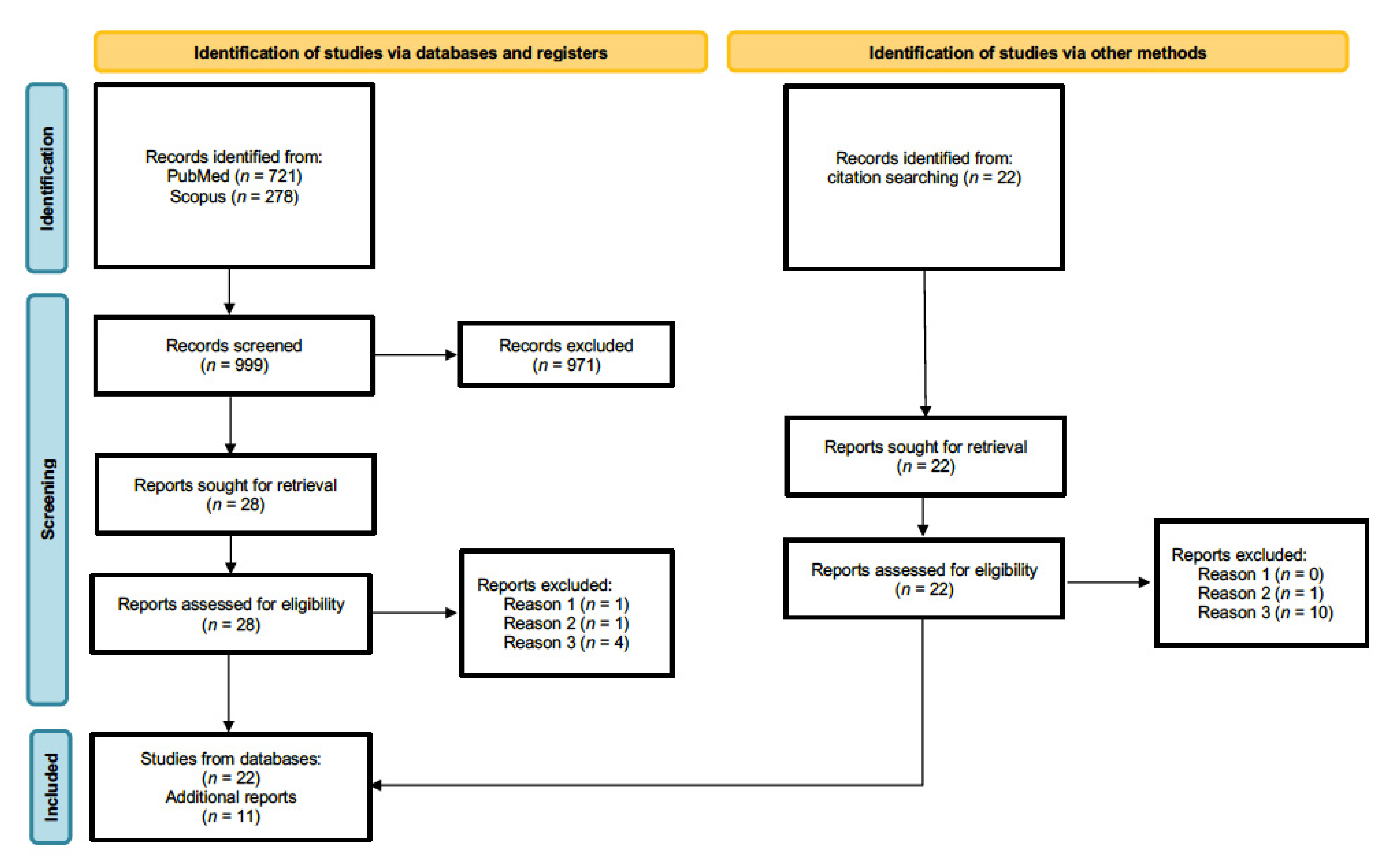
2. Methods
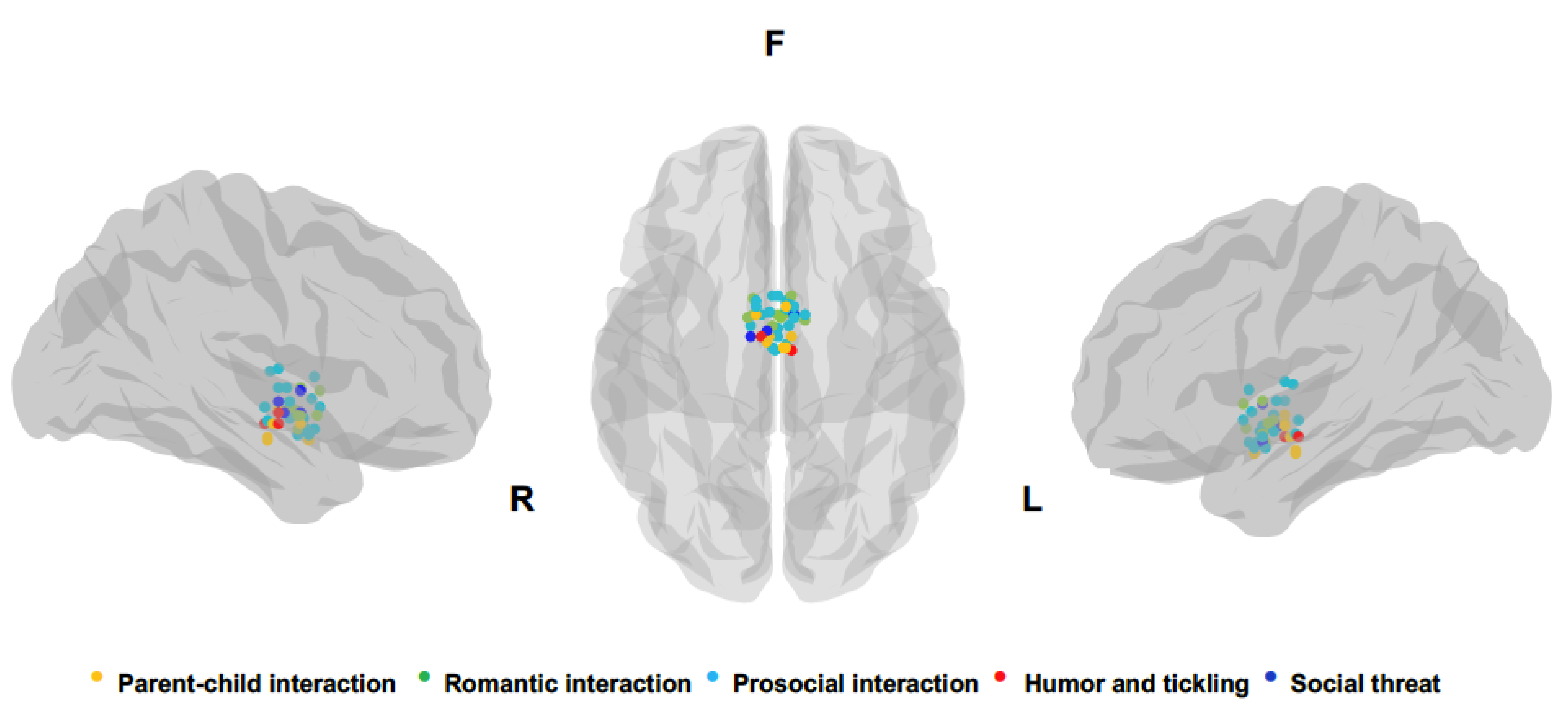
3. Results
3.1. Parental Interactions
3.2. Pair-Bonding
3.3. Other Prosocial Interactions
3.4. Laughter
3.5. Social Threat
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Reeves, A.G.; Plum, F. Hyperphagia, rage, and dementia accompanying a ventromedial hypothalamic neoplasm. Arch. Neurol. 1969, 20, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Gorman, D.G.; Cummings, J.L. Hypersexuality following septal injury. Arch. Neurol. 1992, 49, 308–310. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.; Franzini, A.; Giannicola, G.; Messina, G.; Altamura, A.C.; Priori, A. Hypothalamic oscillations in human pathological aggressiveness. Biol. Psychiatry 2012, 72, e33–e35. [Google Scholar] [CrossRef] [PubMed]
- Bejjani, B.P.; Houeto, J.L.; Hariz, M.; Yelnik, J.; Mesnage, V.; Bonnet, A.M.; Pidoux, B.; Dormont, D.; Cornu, P.; Agid, Y. Aggressive behavior induced by intraoperative stimulation in the triangle of Sano. Neurology 2002, 59, 1425–1427. [Google Scholar] [CrossRef] [PubMed]
- Giustina, A.; Braunstein, G.D. Hypothalamic Syndromes. In Endocrinology: Adult and Pediatric; Elsevier: Amsterdam, The Netherlands, 2016; pp. 174–187. [Google Scholar]
- Kunwar, P.S.; Zelikowsky, M.; Remedios, R.; Cai, H.; Yilmaz, M.; Meister, M.; Anderson, D.J. Ventromedial hypothalamic neurons control a defensive emotion state. eLife 2015, 4, e06633. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.A.; Mattucci, C.; Krzywkowski, P.; Murana, E.; Illarionova, A.; Grinevich, V.; Canteras, N.S.; Ragozzino, D.; Gross, C.T. Independent hypothalamic circuits for social and predator fear. Nat. Neurosci. 2013, 16, 1731–1733. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, I.Z.; Lin, D. Collateral pathways from the ventromedial hypothalamus mediate defensive behaviors. Neuron 2015, 85, 1344–1358. [Google Scholar] [CrossRef]
- Mangieri, L.R.; Jiang, Z.; Lu, Y.; Xu, Y.; Cassidy, R.M.; Justice, N.; Xu, Y.; Arenkiel, B.R.; Tong, Q. Defensive Behaviors Driven by a Hypothalamic-Ventral Midbrain Circuit. eNeuro 2019, 6, 1–19. [Google Scholar] [CrossRef]
- Lin, D.; Boyle, M.P.; Dollar, P.; Lee, H.; Lein, E.S.; Perona, P.; Anderson, D.J. Functional identification of an aggression locus in the mouse hypothalamus. Nature 2011, 470, 221–226. [Google Scholar] [CrossRef]
- Meyer-Lindenberg, A.; Domes, G.; Kirsch, P.; Heinrichs, M. Oxytocin and vasopressin in the human brain: Social neuropeptides for translational medicine. Nat. Rev. Neurosci. 2011, 12, 524–538. [Google Scholar] [CrossRef]
- Quattrocki, E.; Friston, K. Autism, oxytocin and interoception. Neurosci. Biobehav. Rev. 2014, 47, 410–430. [Google Scholar] [CrossRef]
- Hammock, E.; Veenstra-VanderWeele, J.; Yan, Z.; Kerr, T.M.; Morris, M.; Anderson, G.M.; Carter, C.S.; Cook, E.H.; Jacob, S. Examining autism spectrum disorders by biomarkers: Example from the oxytocin and serotonin systems. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 712–721.e11. [Google Scholar] [CrossRef]
- Torres, N.; Martins, D.; Santos, A.J.; Prata, D.; Verissimo, M. How do hypothalamic nonapeptides shape youth’s sociality? A systematic review on oxytocin, vasopressin and human socio-emotional development. Neurosci. Biobehav. Rev. 2018, 90, 309–331. [Google Scholar] [CrossRef]
- Wang, D.; Xue, S.W.; Tan, Z.; Wang, Y.; Lian, Z.; Sun, Y. Altered hypothalamic functional connectivity patterns in major depressive disorder. Neuroreport 2019, 30, 1115–1120. [Google Scholar] [CrossRef]
- Schindler, S.; Geyer, S.; Strauss, M.; Anwander, A.; Hegerl, U.; Turner, R.; Schonknecht, P. Structural studies of the hypothalamus and its nuclei in mood disorders. Psychiatry Res. 2012, 201, 1–9. [Google Scholar] [CrossRef]
- Caria, A.; Ciringione, L.; Falco, S. Morphofunctional Alterations of the Hypothalamus and Social Behavior in Autism Spectrum Disorders. Brain Sci. 2020, 10, 435. [Google Scholar] [CrossRef]
- Guastella, A.J.; Mitchell, P.B.; Dadds, M.R. Oxytocin increases gaze to the eye region of human faces. Biol. Psychiatry 2008, 63, 3–5. [Google Scholar] [CrossRef]
- Rimmele, U.; Hediger, K.; Heinrichs, M.; Klaver, P. Oxytocin makes a face in memory familiar. J. Neurosci. 2009, 29, 38–42. [Google Scholar] [CrossRef]
- Savaskan, E.; Ehrhardt, R.; Schulz, A.; Walter, M.; Schachinger, H. Post-learning intranasal oxytocin modulates human memory for facial identity. Psychoneuroendocrinology 2008, 33, 368–374. [Google Scholar] [CrossRef]
- Domes, G.; Heinrichs, M.; Michel, A.; Berger, C.; Herpertz, S.C. Oxytocin improves “mind-reading” in humans. Biol. Psychiatry 2007, 61, 731–733. [Google Scholar] [CrossRef]
- Kosfeld, M.; Heinrichs, M.; Zak, P.J.; Fischbacher, U.; Fehr, E. Oxytocin increases trust in humans. Nature 2005, 435, 673–676. [Google Scholar] [CrossRef]
- Bethlehem, R.A.; van Honk, J.; Auyeung, B.; Baron-Cohen, S. Oxytocin, brain physiology, and functional connectivity: A review of intranasal oxytocin fMRI studies. Psychoneuroendocrinology 2013, 38, 962–974. [Google Scholar] [CrossRef]
- Gordon, I.; Vander Wyk, B.C.; Bennett, R.H.; Cordeaux, C.; Lucas, M.V.; Eilbott, J.A.; Zagoory-Sharon, O.; Leckman, J.F.; Feldman, R.; Pelphrey, K.A. Oxytocin enhances brain function in children with autism. Proc. Natl. Acad. Sci. USA 2013, 110, 20953–20958. [Google Scholar] [CrossRef]
- Kirsch, P.; Esslinger, C.; Chen, Q.; Mier, D.; Lis, S.; Siddhanti, S.; Gruppe, H.; Mattay, V.S.; Gallhofer, B.; Meyer-Lindenberg, A. Oxytocin modulates neural circuitry for social cognition and fear in humans. J. Neurosci. 2005, 25, 11489–11493. [Google Scholar] [CrossRef]
- Riem, M.M.; van, I.M.H.; Tops, M.; Boksem, M.A.; Rombouts, S.A.; Bakermans-Kranenburg, M.J. Oxytocin effects on complex brain networks are moderated by experiences of maternal love withdrawal. Eur. Neuropsychopharmacol. 2013, 23, 1288–1295. [Google Scholar] [CrossRef]
- Wittfoth-Schardt, D.; Grunding, J.; Wittfoth, M.; Lanfermann, H.; Heinrichs, M.; Domes, G.; Buchheim, A.; Gundel, H.; Waller, C. Oxytocin modulates neural reactivity to children’s faces as a function of social salience. Neuropsychopharmacology 2012, 37, 1799–1807. [Google Scholar] [CrossRef]
- Domes, G.; Heinrichs, M.; Glascher, J.; Buchel, C.; Braus, D.F.; Herpertz, S.C. Oxytocin attenuates amygdala responses to emotional faces regardless of valence. Biol. Psychiatry 2007, 62, 1187–1190. [Google Scholar] [CrossRef]
- Ne’eman, R.; Perach-Barzilay, N.; Fischer-Shofty, M.; Atias, A.; Shamay-Tsoory, S.G. Intranasal administration of oxytocin increases human aggressive behavior. Horm. Behav. 2016, 80, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Harari-Dahan, O.; Bernstein, A. A general approach-avoidance hypothesis of oxytocin: Accounting for social and non-social effects of oxytocin. Neurosci. Biobehav. Rev. 2014, 47, 506–519. [Google Scholar] [CrossRef] [PubMed]
- Mierop, A.; Mikolajczak, M.; Stahl, C.; Bena, J.; Luminet, O.; Lane, A.; Corneille, O. How Can Intranasal Oxytocin Research Be Trusted? A Systematic Review of the Interactive Effects of Intranasal Oxytocin on Psychosocial Outcomes. Perspect. Psychol. Sci. 2020, 15, 1228–1242. [Google Scholar] [CrossRef] [PubMed]
- Andari, E.; Richard, N.; Leboyer, M.; Sirigu, A. Adaptive coding of the value of social cues with oxytocin, an fMRI study in autism spectrum disorder. Cortex 2016, 76, 79–88. [Google Scholar] [CrossRef]
- Aoki, Y.; Yahata, N.; Watanabe, T.; Takano, Y.; Kawakubo, Y.; Kuwabara, H.; Iwashiro, N.; Natsubori, T.; Inoue, H.; Suga, M.; et al. Oxytocin improves behavioural and neural deficits in inferring others’ social emotions in autism. Brain 2014, 137, 3073–3086. [Google Scholar] [CrossRef]
- Aoki, Y.; Yamasue, H. Reply: Does imitation act as an oxytocin nebulizer in autism spectrum disorder? Brain 2015, 138, e361. [Google Scholar] [CrossRef]
- Watanabe, T.; Abe, O.; Kuwabara, H.; Yahata, N.; Takano, Y.; Iwashiro, N.; Natsubori, T.; Aoki, Y.; Takao, H.; Kawakubo, Y.; et al. Mitigation of sociocommunicational deficits of autism through oxytocin-induced recovery of medial prefrontal activity: A randomized trial. JAMA Psychiatry 2014, 71, 166–175. [Google Scholar] [CrossRef]
- Watanabe, T.; Kuroda, M.; Kuwabara, H.; Aoki, Y.; Iwashiro, N.; Tatsunobu, N.; Takao, H.; Nippashi, Y.; Kawakubo, Y.; Kunimatsu, A.; et al. Clinical and neural effects of six-week administration of oxytocin on core symptoms of autism. Brain 2015, 138, 3400–3412. [Google Scholar] [CrossRef]
- Kanat, M.; Spenthof, I.; Riedel, A.; van Elst, L.T.; Heinrichs, M.; Domes, G. Restoring effects of oxytocin on the attentional preference for faces in autism. Transl. Psychiatry 2017, 7, e1097. [Google Scholar] [CrossRef]
- Parker, K.J.; Oztan, O.; Libove, R.A.; Mohsin, N.; Karhson, D.S.; Sumiyoshi, R.D.; Summers, J.E.; Hinman, K.E.; Motonaga, K.S.; Phillips, J.M.; et al. A randomized placebo-controlled pilot trial shows that intranasal vasopressin improves social deficits in children with autism. Sci. Transl. Med. 2019, 11, eaau7356. [Google Scholar] [CrossRef]
- Mobbs, D.; Greicius, M.D.; Abdel-Azim, E.; Menon, V.; Reiss, A.L. Humor modulates the mesolimbic reward centers. Neuron 2003, 40, 1041–1048. [Google Scholar] [CrossRef]
- Provine, R.R. Laughing, Tickling, and the Evolution of Speech and Self. Curr. Dir. Psychol. Sci. 2004, 13, 215–218. [Google Scholar] [CrossRef]
- Wattendorf, E.; Westermann, B.; Fiedler, K.; Ritz, S.; Redmann, A.; Pfannmoller, J.; Lotze, M.; Celio, M.R. Laughter is in the air: Involvement of key nodes of the emotional motor system in the anticipation of tickling. Soc. Cogn. Affect. Neurosci. 2019, 14, 837–847. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Lenzi, D.; Trentini, C.; Tambelli, R.; Pantano, P. Neural basis of attachment-caregiving systems interaction: Insights from neuroimaging studies. Front. Psychol. 2015, 6, 1241. [Google Scholar] [CrossRef]
- De Boer, A.; van Buel, E.M.; Ter Horst, G.J. Love is more than just a kiss: A neurobiological perspective on love and affection. Neuroscience 2012, 201, 114–124. [Google Scholar] [CrossRef]
- Mercado, E.; Hibel, L.C. I love you from the bottom of my hypothalamus: The role of stress physiology in romantic pair bond formation and maintenance. Soc. Pers. Psychol. Compass 2017, 11, 1–12. [Google Scholar] [CrossRef]
- Swain, J.E. The human parental brain: In vivo neuroimaging. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 1242–1254. [Google Scholar] [CrossRef]
- Wild, B.; Rodden, F.A.; Grodd, W.; Ruch, W. Neural correlates of laughter and humour. Brain 2003, 126, 2121–2138. [Google Scholar] [CrossRef]
- Mashek, D.; Aron, A.; Fisher, H.E. Identifying, evoking, and measuring intense feelings of romantic love. Rep. Res. Soc. Psychol. 2000, 24, 48–55. [Google Scholar]
- Koscik, T.R.; Tranel, D. The human amygdala is necessary for developing and expressing normal interpersonal trust. Neuropsychologia 2011, 49, 602–611. [Google Scholar] [CrossRef]
- Conner, O.L.; Siegle, G.J.; McFarland, A.M.; Silk, J.S.; Ladouceur, C.D.; Dahl, R.E.; Coan, J.A.; Ryan, N.D. Mom-it helps when you’re right here! Attenuation of neural stress markers in anxious youths whose caregivers are present during fMRI. PLoS ONE 2012, 7, e50680. [Google Scholar] [CrossRef]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A network visualization tool for human brain connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef]
- Lorberbaum, J.P.; Newman, J.D.; Horwitz, A.R.; Dubno, J.R.; Lydiard, R.B.; Hamner, M.B.; Bohning, D.E.; George, M.S. A potential role for thalamocingulate circuitry in human maternal behavior. Biol. Psychiatry 2002, 51, 431–445. [Google Scholar] [CrossRef]
- Bartels, A.; Zeki, S. The neural correlates of maternal and romantic love. Neuroimage 2004, 21, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Strathearn, L.; Li, J.; Fonagy, P.; Montague, P.R. What’s in a smile? Maternal brain responses to infant facial cues. Pediatrics 2008, 122, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Noriuchi, M.; Kikuchi, Y.; Senoo, A. The functional neuroanatomy of maternal love: Mother’s response to infant’s attachment behaviors. Biol. Psychiatry 2008, 63, 415–423. [Google Scholar] [CrossRef]
- Strathearn, L.; Fonagy, P.; Amico, J.; Montague, P.R. Adult attachment predicts maternal brain and oxytocin response to infant cues. Neuropsychopharmacology 2009, 34, 2655–2666. [Google Scholar] [CrossRef]
- Moll, J.; Bado, P.; de Oliveira-Souza, R.; Bramati, I.E.; Lima, D.O.; Paiva, F.F.; Sato, J.R.; Tovar-Moll, F.; Zahn, R. A neural signature of affiliative emotion in the human septohypothalamic area. J. Neurosci. 2012, 32, 12499–12505. [Google Scholar] [CrossRef]
- Ho, S.S.; Konrath, S.; Brown, S.; Swain, J.E. Empathy and stress related neural responses in maternal decision making. Front. Neurosci. 2014, 8, 152. [Google Scholar] [CrossRef]
- Li, T.; Horta, M.; Mascaro, J.S.; Bijanki, K.; Arnal, L.H.; Adams, M.; Barr, R.G.; Rilling, J.K. Explaining individual variation in paternal brain responses to infant cries. Physiol. Behav. 2018, 193, 43–54. [Google Scholar] [CrossRef]
- Najib, A.; Lorberbaum, J.P.; Kose, S.; Bohning, D.E.; George, M.S. Regional brain activity in women grieving a romantic relationship breakup. Am. J. Psychiatry 2004, 161, 2245–2256. [Google Scholar] [CrossRef]
- Karremans, J.C.; Heslenfeld, D.J.; van Dillen, L.F.; Van Lange, P.A. Secure attachment partners attenuate neural responses to social exclusion: An fMRI investigation. Int. J. Psychophysiol. 2011, 81, 44–50. [Google Scholar] [CrossRef]
- Xu, X.; Aron, A.; Brown, L.; Cao, G.; Feng, T.; Weng, X. Reward and motivation systems: A brain mapping study of early-stage intense romantic love in Chinese participants. Hum. Brain Mapp. 2011, 32, 249–257. [Google Scholar] [CrossRef]
- Acevedo, B.P.; Poulin, M.J.; Brown, L.L. Beyond romance: Neural and genetic correlates of altruism in pair-bonds. Behav. Neurosci. 2019, 133, 18–31. [Google Scholar] [CrossRef]
- Heijne, A.; Rossi, F.; Sanfey, A.G. Why we stay with our social partners: Neural mechanisms of stay/leave decision-making. Soc. Neurosci. 2018, 13, 667–679. [Google Scholar] [CrossRef]
- Coan, J.A.; Schaefer, H.S.; Davidson, R.J. Lending a hand: Social regulation of the neural response to threat. Psychol. Sci. 2006, 17, 1032–1039. [Google Scholar] [CrossRef]
- Moll, J.; Krueger, F.; Zahn, R.; Pardini, M.; de Oliveira-Souza, R.; Grafman, J. Human fronto-mesolimbic networks guide decisions about charitable donation. Proc. Natl. Acad. Sci. USA 2006, 103, 15623–15628. [Google Scholar] [CrossRef]
- Eisenberger, N.I.; Lieberman, M.D.; Williams, K.D. Does rejection hurt? An FMRI study of social exclusion. Science 2003, 302, 290–292. [Google Scholar] [CrossRef]
- Immordino-Yang, M.H.; McColl, A.; Damasio, H.; Damasio, A. Neural correlates of admiration and compassion. Proc. Natl. Acad. Sci. USA 2009, 106, 8021–8026. [Google Scholar] [CrossRef]
- Wolfe, F.H.; Deruelle, C.; Chaminade, T. Are friends really the family we choose? Local variations of hypothalamus activity when viewing personally known faces. Soc. Neurosci. 2018, 13, 289–300. [Google Scholar] [CrossRef]
- Yu, H.; Cai, Q.; Shen, B.; Gao, X.; Zhou, X. Neural substrates and social consequences of interpersonal gratitude: Intention matters. Emotion 2017, 17, 589–601. [Google Scholar] [CrossRef]
- Lopez-Sola, M.; Geuter, S.; Koban, L.; Coan, J.A.; Wager, T.D. Brain mechanisms of social touch-induced analgesia in females. Pain 2019, 160, 2072–2085. [Google Scholar] [CrossRef]
- Rauchbauer, B.; Nazarian, B.; Bourhis, M.; Ochs, M.; Prevot, L.; Chaminade, T. Brain activity during reciprocal social interaction investigated using conversational robots as control condition. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180033. [Google Scholar] [CrossRef]
- Bortolini, T.; Melo, B.; Basilio, R.; Fischer, R.; Zahn, R.; de Oliveira-Souza, R.; Knutson, B.; Moll, J. Striatal and septo-hypothalamic responses to anticipation and outcome of affiliative rewards. Neuroimage 2021, 243, 118474. [Google Scholar] [CrossRef]
- Karlsson, K.A.; Windischberger, C.; Gerstl, F.; Mayr, W.; Siegel, J.M.; Moser, E. Modulation of hypothalamus and amygdalar activation levels with stimulus valence. Neuroimage 2010, 51, 324–328. [Google Scholar] [CrossRef][Green Version]
- Pichon, S.; de Gelder, B.; Grezes, J. Emotional modulation of visual and motor areas by dynamic body expressions of anger. Soc. Neurosci. 2008, 3, 199–212. [Google Scholar] [CrossRef]
- Hermans, E.J.; Ramsey, N.F.; van Honk, J. Exogenous testosterone enhances responsiveness to social threat in the neural circuitry of social aggression in humans. Biol. Psychiatry 2008, 63, 263–270. [Google Scholar] [CrossRef]
- Sinke, C.B.; Sorger, B.; Goebel, R.; de Gelder, B. Tease or threat? Judging social interactions from bodily expressions. Neuroimage 2010, 49, 1717–1727. [Google Scholar] [CrossRef]
- Pichon, S.; de Gelder, B.; Grezes, J. Threat prompts defensive brain responses independently of attentional control. Cereb. Cortex 2012, 22, 274–285. [Google Scholar] [CrossRef]
- George, C.; Kaplan, N.; Main, M. Adult Attachment Interview; Unpublished manuscript; University of California: Berkeley, CA, USA, 1985. [Google Scholar]
- Singewald, G.M.; Rjabokon, A.; Singewald, N.; Ebner, K. The modulatory role of the lateral septum on neuroendocrine and behavioral stress responses. Neuropsychopharmacology 2011, 36, 793–804. [Google Scholar] [CrossRef]
- Atzil, S.; Hendler, T.; Feldman, R. Specifying the neurobiological basis of human attachment: Brain, hormones, and behavior in synchronous and intrusive mothers. Neuropsychopharmacology 2011, 36, 2603–2615. [Google Scholar] [CrossRef]
- Carter, C.S. Neuroendocrine perspectives on social attachment and love. Psychoneuroendocrinology 1998, 23, 779–818. [Google Scholar] [CrossRef]
- Bartels, A.; Zeki, S. The neural basis of romantic love. Neuroreport 2000, 11, 3829–3834. [Google Scholar] [CrossRef] [PubMed]
- Hazan, C.; Shaver, P. Romantic love conceptualized as an attachment process. J. Pers. Soc. Psychol. 1987, 52, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Buffone, A.E.; Poulin, M.J. Empathy, target distress, and neurohormone genes interact to predict aggression for others-even without provocation. Pers. Soc. Psychol. Bull. 2014, 40, 1406–1422. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.M.; Saslow, L.R.; Garcia, N.; John, O.P.; Keltner, D. Oxytocin receptor genetic variation relates to empathy and stress reactivity in humans. Proc. Natl. Acad. Sci. USA 2009, 106, 21437–21441. [Google Scholar] [CrossRef]
- Uzefovsky, F.; Shalev, I.; Israel, S.; Edelman, S.; Raz, Y.; Mankuta, D.; Knafo-Noam, A.; Ebstein, R.P. Oxytocin receptor and vasopressin receptor 1a genes are respectively associated with emotional and cognitive empathy. Horm. Behav. 2015, 67, 60–65. [Google Scholar] [CrossRef]
- De Vries, A.C.; Glasper, E.R.; Detillion, C.E. Social modulation of stress responses. Physiol. Behav. 2003, 79, 399–407. [Google Scholar] [CrossRef]
- Brown, C.L.; Beckes, L.; Allen, J.P.; Coan, J.A. Subjective General Health and the Social Regulation of Hypothalamic Activity. Psychosom. Med. 2017, 79, 670–673. [Google Scholar] [CrossRef]
- Takahashi, H.; Matsuura, M.; Yahata, N.; Koeda, M.; Suhara, T.; Okubo, Y. Men and women show distinct brain activations during imagery of sexual and emotional infidelity. Neuroimage 2006, 32, 1299–1307. [Google Scholar] [CrossRef]
- Eisenberger, N.I.; Taylor, S.E.; Gable, S.L.; Hilmert, C.J.; Lieberman, M.D. Neural pathways link social support to attenuated neuroendocrine stress responses. Neuroimage 2007, 35, 1601–1612. [Google Scholar] [CrossRef]
- Krueger, F.; McCabe, K.; Moll, J.; Kriegeskorte, N.; Zahn, R.; Strenziok, M.; Heinecke, A.; Grafman, J. Neural correlates of trust. Proc. Natl. Acad. Sci. USA 2007, 104, 20084–20089. [Google Scholar] [CrossRef]
- Chaminade, T.; Da Fonseca, D.; Rosset, D.; Cheng, G.; Deruelle, C. Atypical modulation of hypothalamic activity by social context in ASD. Res. Autism Spectr. Disord. 2015, 10, 41–50. [Google Scholar] [CrossRef]
- Provine, R.R. Funny science: Review: Ha! The science of when we laugh and why and the humor code: A global search for what makes things funny. Cerebrum 2014, 2014, 10. [Google Scholar]
- Panksepp, J.; Burgdorf, J. “Laughing” rats and the evolutionary antecedents of human joy? Physiol. Behav. 2003, 79, 533–547. [Google Scholar] [CrossRef]
- Wattendorf, E.; Westermann, B.; Fiedler, K.; Kaza, E.; Lotze, M.; Celio, M.R. Exploration of the neural correlates of ticklish laughter by functional magnetic resonance imaging. Cereb. Cortex 2013, 23, 1280–1289. [Google Scholar] [CrossRef]
- Dezecache, G.D.; Dunbar, R.I.M. Sharing the joke: The size of natural laughter groups. Evol. Hum. Behav. 2012, 33, 775–779. [Google Scholar] [CrossRef]
- Provine, R.R. Laughing, grooming, and pub science. Trends Cogn. Sci. 2013, 17, 9–10. [Google Scholar] [CrossRef]
- Provine, R.R. Laughter: A Scientific Investigation; Penguin Press: London, UK, 2001. [Google Scholar]
- Martin, R.A. The Psychology of Humor: An Integrative Approach; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]



{kind=link}
{kind=link}
| Year | Authors | Socioemotional Context | Sample Size | Sex | Main Stimuli Comparison | Hypothalamic Region | MNI Coordinates (Cluster Peak) |
|---|---|---|---|---|---|---|---|
| 2002 | Lorberbaum, et al. [52] | Mother-infant interaction | N = 10 | M = 0, F = 10 | Cry stimuli vs control noise | Hypothalamus | 0, −6, −7 0, −10, −3 |
| 2004 | Bartels & Zeki [53] | Romantic and maternal interactions | N = 20 | M = 0, F = 20 | Own childs photographs vs other childs photographs | L hypothalamus | −3, −12, −17 |
| 2008 | Strathearn, et al. [54] | Mother-infant interaction | N = 28 | M = 0, F = 28 | Own baby’s face vs unknown baby face | Bilateral hypothalamus | 3, −8, −7 −5, −8, −8 |
| 2008 | Noriuchi, et al. [55] | Mother-infant interaction | N = 13 | M = 0, F = 13 | Video clips of own infant vs unknown infants in two situation: play vs separation | R hypothalamus | 6, −8, −4 4, −10, −12 |
| 2009 | Strathearn, et al. [56] | Mother-infant interaction | N = 30 | M = 0, F = 30 | Own infant vs unknown infant pictures | L hypothalamus | −3, 3, −18 |
| 2012 | Moll, et al. [57] | Parent-child interaction | N = 30 | M = 14, F = 16 | Affiliative-positive and negative vs non affiliative positive and negative | L septal/preoptic–anterior hypothalamic area | −3, 2, −14 |
| 2014 | Ho, et al. [58] | Mother-infant interaction | N = 14 | M = 0, F = 14 | Own infant vs unknown infants pictures | R septal-hypothalamic area | 8, 0, −12 |
| 2018 | Li, et al. [59] | Father-infant interaction | N = 39 | M = 39, F = 0 | own infant cry vs unknown infant cry | L hypothalamus | −2, −12, −18 |
| 2004 | Najib, et al. [60] | Romantic interaction | N = 11 | M = 0, F = 11 | Recalling of sad vs neutral thoughts | Hypothalamus | 0, 0, 1 |
| 2011 | Karremans, et al. [61] | Romantic interaction | N = 15 | M = 5, F = 10 | Attachment figure’s name vs non-attachment figure’s name | L hypothalamus | −1, −1, −14 |
| 2011 | Xu, et al. [62] | Romantic interaction | N = 18 | M = 8, F = 10 | Romantic partner VS familiar acquaintance photographs | L hypothalamus | −2, 0, −11 |
| 2012 | Acevedo, et al. [63] | Romantic interaction | N = 17 | M = 7, F = 10 | Partner vs highly familiar acquaintance images; conjunction partner and close friend | Bilateral hypothalamus | −10, −2, −7 2, −4, −6 11, −1, −9 |
| 2017 | Heijne, et al. [64] | Romantic interaction | N = 26 | M = 13, F = 13 | Social vs nonsocial “stay” decisions | L septo-hypothalamic region | −5, 7, 0 |
| 2019 | Acevedo, et al. [63] | Romantic interaction | N = 18 | M = 11, F = 7 | Partner vs familiar face | R hypothalamus | 9, 6, −9 9, 0, −9 |
| 2006 | Coan, et al. [65] | Prosocial interaction | N =16 | M = 0, F = 16 | Husband’s hand-holding vs anonymous male experimenter’ hand-holding | R hypothalamus | 1, −13, −6 |
| 2006 | Moll, et al. [66] | Prosocial interaction | N = 19 | M = 10, F = 9 | Decisions to donate vs pure monetary reward | R septo-hypothalamic region | Not available |
| 2007 | Eisenberger, et al. [67] | Prosocial interaction | N = 32 | M = 13, F = 19 | Social exclusion vs inclusion during a virtual ball tossinggame | Bilateral hypothalamus | 10, −4, −4 6, 0, −8 −10, 0, −12 |
| 2007 | Krueger, et al. [66]. | Prosocial interaction | N = 44 | M = 22, F = 22 | Trust during social reciprocal trust game vs control game | L septo-hypothalamic region | −4, 4,−3 |
| 2009 | Immordino-Yang, et al. [68] | Prosocial interaction | N = 13 | M = 7, F = 9 | Emotional vs non emotional narratives - Admiration for virtue & Admiration for skill & Compassion for social pain & Compassion for physical pain vs control; admiration for virtue and compassion for social/psychological pain vs control | Bilateral hypothalamus | 0, −5, 1 −3, −11, 7 −0, −8, 8 3, −8, 1 −3, 5, 5 |
| 2017 | Wolfe, et al. [69]. | Prosocial interaction | N = 20 | M = 6, F = 14 | Several contrasts among pictures of friend, sibling and celebrity vs unknown | L and R hypothalamic supraoptic and paraventricular nuclei | −6, 3, –16 8, 3, –16 8, 5, –14 −6, –1, –16 3, 1, –10 |
| 2017 | Brown, et al. [58] | Prosocial interaction | N = 75 | M = 41, F = 34 | Partner handholding vs stranger handholding | R hypothalamus | 2, −12, −11 |
| 2017 | Yu, et al. [70]. | Prosocial interaction | N = 27 | M = 11, F = 16 | Sharing vs non-sharing pain stimulation | L hypothalamus | −3, 2, −14 |
| 2019 | Lòpez-Solà, et al. [71] | Prosocial interaction | N = 30 | M = 0, F = 30 | Partner hand-holding vs holding an inert rubber device | Hypothalamus | Not available |
| 2019 | Rauchbauer, et al. [72] | Prosocial interaction | N = 24 | M = 7, F = 17 | Human vs robot agent interaction | Bilateral hypothalamus | Not available |
| 2021 | Bortolini, et al. [73] | Prosocial interaction | N = 23 | M = 9, F = 14 | Videoclips of affiliative vs non affiliative scences with unfamiliar individuals | Bilateral septo-hypothalamic region | −4, −4, −10 −2, 7, −6 0, 7, −6 |
| 2003 | Mobbs, et al. [39] | Humor | N = 16 | M = 7, F = 9 | Funny vs nonfunny cartoons | Bilateral hypothalamus | Not available |
| 2010 | Karlsson, et al. [74] | Humour | N = 20 | M = 16, F = 4 | Funny and sad vs neutral pictures | R hypothalamus | 6, −8, −8 6, −8, −12 |
| 2019 | Wattendorf, et al. [41]. | Tickling | N = 31 | M = 10, F = 21 | Tickling of the foot by a friend/partner vs monotonous foot contact | L posterior lateral hypothalamus | −5, −13, −12 |
| 2008 | Pichon, et al. [75] | Social threat | N = 16 | M = 9, F = 7 | Angry dynamic expressions vs statis stimuli | L hypothalamus | −6, 0, −14 −6, 0, −12 |
| 2008 | Hermans, et al. [76] | Social threat | N = 12 | M = 0, F = 12 | Angry faces vs neutral faces | R hypothalamus | 8, 0, −8 8, 0, 0 |
| 2010 | Sinke, et al. [77] | Social threat | N = 14 | M = 5, F = 9 | Movies of threatening vs teasing interactions | R hypothalamus | 4, −6, −8 |
| 2012 | Pichon, et al. [78] | Social threat | N = 16 | M = 8, F = 8 | Videos of fearful or angry expressions vs neutral expressions | R dorsal hypothalamus | 10, −8, −4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caria, A.; Dall’Ò, G.M. Functional Neuroimaging of Human Hypothalamus in Socioemotional Behavior: A Systematic Review. Brain Sci. 2022, 12, 707. https://doi.org/10.3390/brainsci12060707
Caria A, Dall’Ò GM. Functional Neuroimaging of Human Hypothalamus in Socioemotional Behavior: A Systematic Review. Brain Sciences. 2022; 12(6):707. https://doi.org/10.3390/brainsci12060707
Chicago/Turabian StyleCaria, Andrea, and Ginevra Matilde Dall’Ò. 2022. "Functional Neuroimaging of Human Hypothalamus in Socioemotional Behavior: A Systematic Review" Brain Sciences 12, no. 6: 707. https://doi.org/10.3390/brainsci12060707
APA StyleCaria, A., & Dall’Ò, G. M. (2022). Functional Neuroimaging of Human Hypothalamus in Socioemotional Behavior: A Systematic Review. Brain Sciences, 12(6), 707. https://doi.org/10.3390/brainsci12060707