
Tinnitus Perception in Light of a Parietal Operculo–Insular Involvement: A Review

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Microscopic Scale
3. Mesoscopic Scale
3.1. Intracerebral Recordings of the Operculo–Insular Cortex
3.2. Perceptions Mapping in the Operculo–Insular Cortex Induced by DES
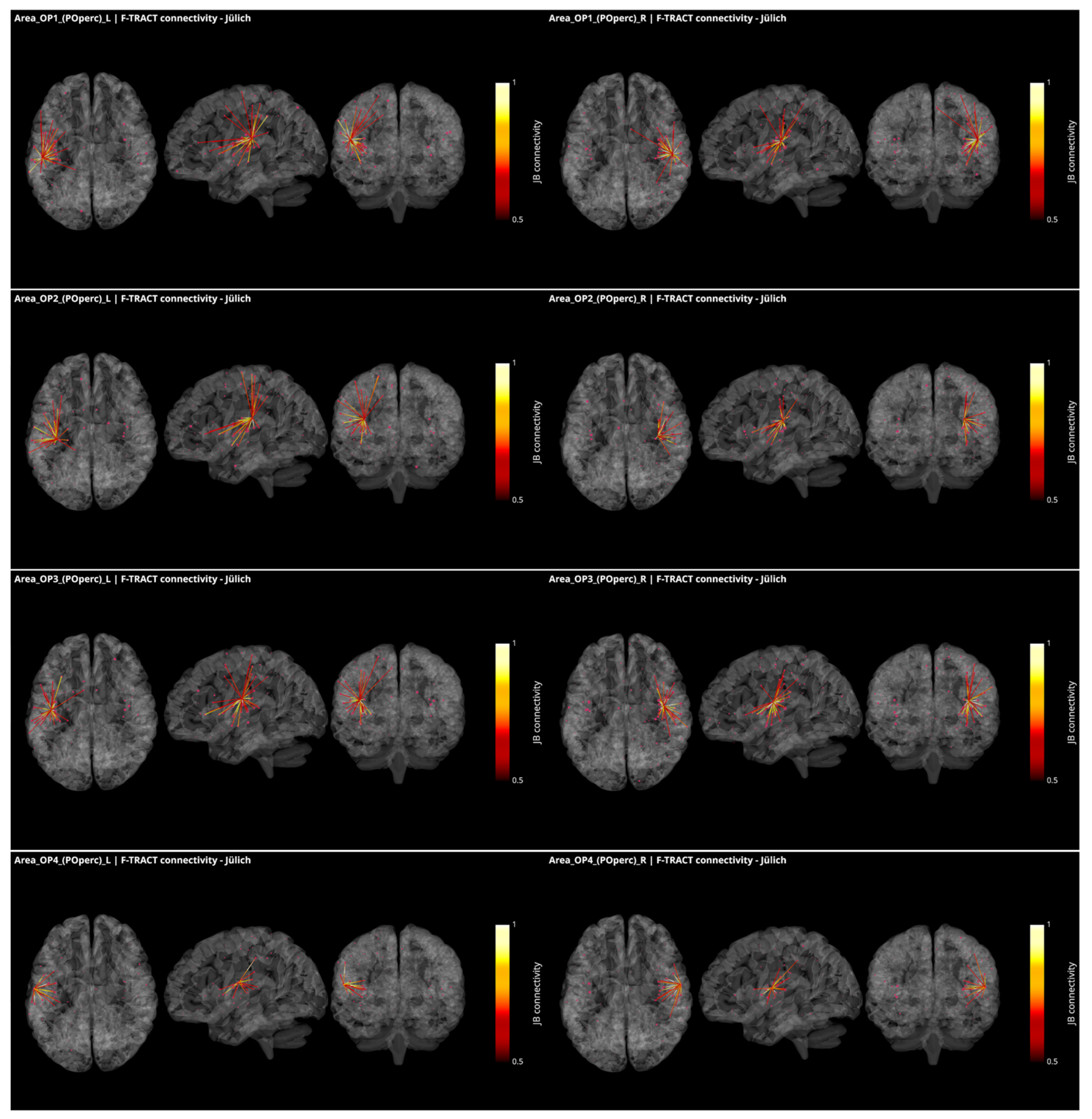
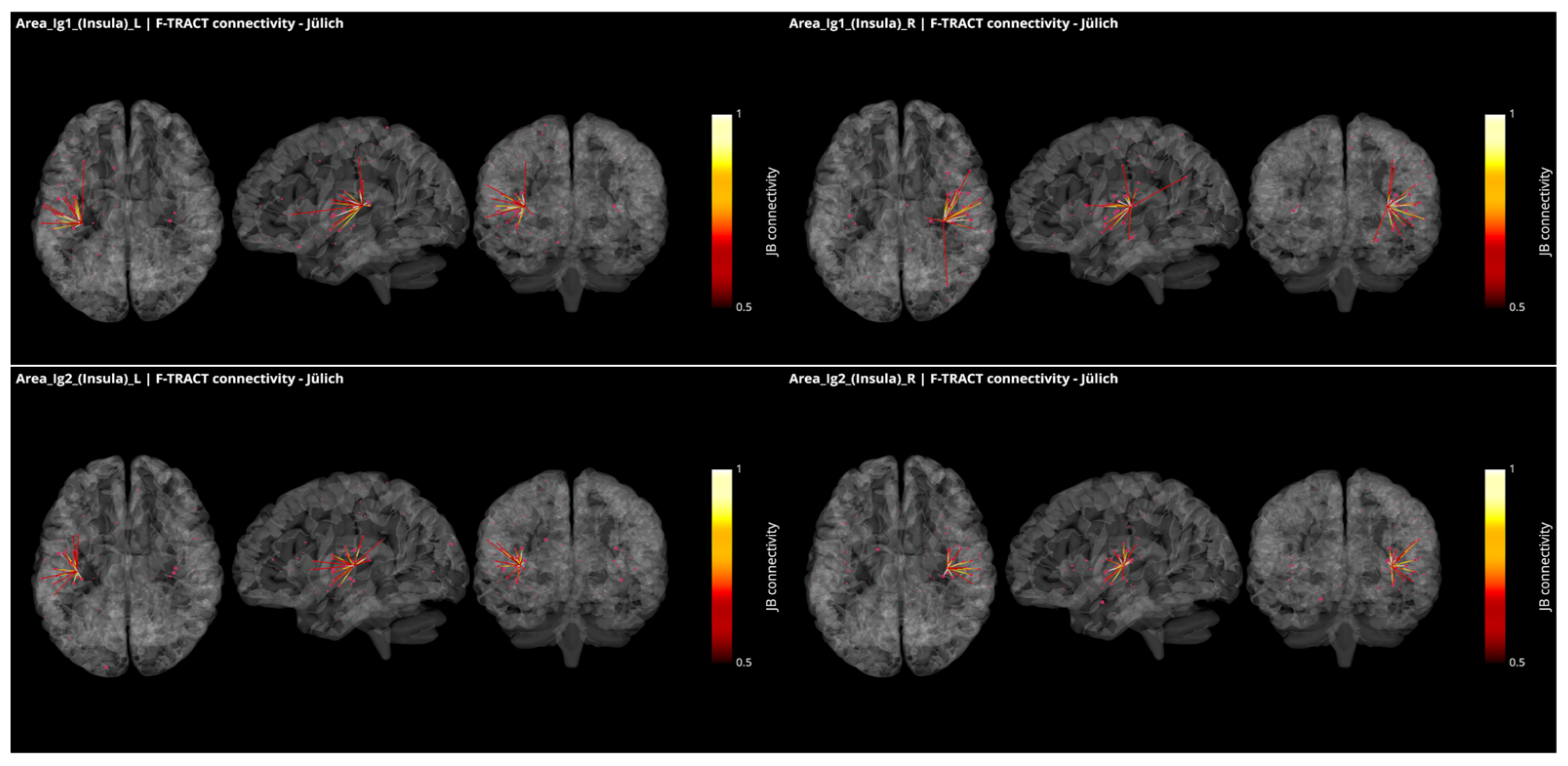
3.3. Operculo–Insular Connectivity
4. Macroscopic Scale
4.1. Integration of Auditory and Somatosensory Stimuli in the Operculo–Insular Cortex
4.2. Functional and Structural Connectivity of the Parietal Operculum and Insular Cortex
4.3. Encoding of Tinnitus in the Parietal Operculo–Insular Cortex
4.4. Functional and Structural Connectivity with the Parietal Operculum in Tinnitus
5. Perspectives for Treatment of Tinnitus
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Biswas, R.; Hall, D.A. Prevalence, Incidence, and Risk Factors for Tinnitus. Curr. Top. Behav. Neurosci. 2021, 51, 3–28. [Google Scholar] [PubMed]
- Baguley, D.; McFerran, D.; Hall, D. Tinnitus. Lancet 2013, 382, 1600–1607. [Google Scholar] [CrossRef] [Green Version]
- Gallus, S.; Lugo, A.; Garavello, W.; Bosetti, C.; Santoro, E.; Colombo, P.; Perin, P.; La, V.C.; Langguth, B. Prevalence and Determinants of Tinnitus in the Italian Adult Population. Neuroepidemiology 2015, 45, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ridder, D.; Schlee, W.; Vanneste, S.; Londero, A.; Weisz, N.; Kleinjung, T.; Shekhawat, G.S.; Elgoyhen, A.B.; Song, J.J.; Andersson, G.; et al. Tinnitus and tinnitus disorder: Theoretical and operational definitions (an international multidisciplinary proposal). Prog. Brain Res. 2021, 260, 1–25. [Google Scholar]
- Wineland, A.M.; Burton, H.; Piccirillo, J. Functional connectivity networks in nonbothersome tinnitus. Otolaryngol. Head Neck Surg. 2012, 147, 900–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter-Thompson, J.R.; Schmidt, S.A.; Husain, F.T. Neural Plasticity of Mild Tinnitus: An fMRI Investigation Comparing Those Recently Diagnosed with Tinnitus to Those That Had Tinnitus for a Long Period of Time. Neural Plast. 2015, 2015, 161478. [Google Scholar] [CrossRef]
- Jain, N.; Qi, H.X.; Collins, C.E.; Kaas, J.H. Large-scale reorganization in the somatosensory cortex and thalamus after sensory loss in macaque monkeys. J. Neurosci. 2008, 28, 11042–11060. [Google Scholar] [CrossRef]
- Gagné, M.; Hétu, S.; Reilly, K.T.; Mercier, C. The map is not the territory: Motor system reorganization in upper limb amputees. Hum. Brain Mapp. 2011, 32, 509–519. [Google Scholar] [CrossRef]
- Sanda, N.; Cerliani, L.; Authié, C.N.; Sabbah, N.; Sahel, J.A.; Habas, C.; Safran, A.B.; Thiebaut de Schotten, M. Visual brain plasticity induced by central and peripheral visual field loss. Brain Struct. Funct. 2018, 223, 3473–3485. [Google Scholar] [CrossRef] [Green Version]
- Gannon, M.A.; Long, S.M.; Parks, N.A. Homeostatic plasticity in human extrastriate cortex following a simulated peripheral scotoma. Exp. Brain Res. 2017, 235, 3391–3401. [Google Scholar] [CrossRef]
- Flor, H.; Diers, M.; Andoh, J. The neural basis of phantom limb pain. Trends Cogn. Sci. 2013, 17, 307–308. [Google Scholar] [CrossRef] [PubMed]
- Langers, D.R.M.; de Kleine, E.; van Dijk, P. Tinnitus does not require macroscopic tonotopic map reorganization. Front. Syst. Neurosci. 2012, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazaleh, N.; Zwaag, W.V.; Clarke, S.; Ville, D.V.; Maire, R.; Saenz, M. High-Resolution fMRI of Auditory Cortical Map Changes in Unilateral Hearing Loss and Tinnitus. Brain Topogr. 2017, 30, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Koops, E.A.; Renken, R.J.; Lanting, C.P.; van Dijk, P. Cortical Tonotopic Map Changes in Humans Are Larger in Hearing Loss Than in Additional Tinnitus. J. Neurosci. 2020, 40, 3178–3185. [Google Scholar] [CrossRef] [PubMed]
- Shore, S.E.; Roberts, L.E.; Langguth, B. Maladaptive plasticity in tinnitus–triggers, mechanisms and treatment. Nat. Rev. Neurol. 2016, 12, 150–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michiels, S.; Cardon, E.; Gilles, A.; Goedhart, H.; Vesala, M.; Schlee, W. Somatosensory Tinnitus Diagnosis: Diagnostic Value of Existing Criteria. Ear Hear. 2021, 43, 143–149. [Google Scholar] [CrossRef]
- Leaver, A.M.; Seydell-Greenwald, A.; Rauschecker, J.P. Auditory-limbic interactions in chronic tinnitus: Challenges for neuroimaging research. Hear. Res. 2016, 334, 49–57. [Google Scholar] [CrossRef]
- Job, A. Rifle impulse noise affects middle-ear compliance in soldiers wearing protective earplugs. Int. J. Audiol. 2016, 55, 30–37. [Google Scholar] [CrossRef]
- Job, A.; Paucod, J.C.; O’Beirne, G.A.; Delon-Martin, C. Cortical representation of tympanic membrane movements due to pressure variation: An fMRI study. Hum. Brain Mapp. 2011, 32, 744–749. [Google Scholar] [CrossRef] [Green Version]
- Job, A.; Pons, Y.; Lamalle, L.; Jaillard, A.; Buck, K.; Segebarth, C.; Delon-Martin, C. Abnormal cortical sensorimotor activity during Target sound detection in subjects with acute acoustic trauma sequelae: An fMRI study. Brain Behav. 2012, 2, 187–199. [Google Scholar] [CrossRef]
- Job, A.; Jaroszynski, C.; Kavounoudias, A.; Jaillard, A.; Delon-Martin, C. Functional Connectivity in Chronic Nonbothersome Tinnitus Following Acoustic Trauma: A Seed-Based Resting-State Functional Magnetic Resonance Imaging Study. Brain Connect. 2020, 6, 32458713. [Google Scholar] [CrossRef]
- Job, A.; Jacob, R.; Pons, Y.; Raynal, M.; Kossowski, M.; Gauthier, J.; Lombard, B.; Delon-Martin, C. Specific activation of operculum 3 (OP3) brain region during provoked tinnitus-related phantom auditory perceptions in humans. Brain Struct. Funct. 2016, 221, 913–922. [Google Scholar] [CrossRef]
- Tsukano, H.; Horie, M.; Ohga, S.; Takahashi, K.; Kubota, Y.; Hishida, R.; Takebayashi, H.; Shibuki, K. Reconsidering Tonotopic Maps in the Auditory Cortex and Lemniscal Auditory Thalamus in Mice. Front. Neural Circuits 2017, 11, 14. [Google Scholar] [PubMed] [Green Version]
- Kaas, J.H.; Hackett, T.A. Subdivisions of auditory cortex and processing streams in primates. Proc. Natl. Acad. Sci. USA 2000, 97, 11793–11799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaas, J.H. The functional organization of somatosensory cortex in primates. Ann. Anat. 1993, 175, 509–518. [Google Scholar] [CrossRef]
- Augustine, J.R. Circuitry and functional aspects of the insular lobe in primates including humans. Brain Res. Brain Res. Rev. 1996, 22, 229–244. [Google Scholar] [CrossRef]
- Ogawa, H. Gustatory cortex of primates: Anatomy and physiology. Neurosci. Res. 1994, 20, 1–13. [Google Scholar] [CrossRef]
- Schroeder, C.E.; Lindsley, R.W.; Specht, C.; Marcovici, A.; Smiley, J.F.; Javitt, D.C. Somatosensory input to auditory association cortex in the macaque monkey. J. Neurophysiol. 2001, 85, 1322–1327. [Google Scholar] [CrossRef]
- Hihara, S.; Taoka, M.; Tanaka, M.; Iriki, A. Visual Responsiveness of Neurons in the Secondary Somatosensory Area and its Surrounding Parietal Operculum Regions in Awake Macaque Monkeys. Cereb. Cortex 2015, 25, 4535–4550. [Google Scholar] [CrossRef]
- Cappe, C.; Barone, P. Heteromodal connections supporting multisensory integration at low levels of cortical processing in the monkey. Eur. J. Neurosci. 2005, 22, 2886–2902. [Google Scholar] [CrossRef]
- Smiley, J.F.; Hackett, T.A.; Ulbert, I.; Karmas, G.; Lakatos, P.; Javitt, D.C.; Schroeder, C.E. Multisensory convergence in auditory cortex, I. Cortical connections of the caudal superior temporal plane in macaque monkeys. J. Comp. Neurol. 2007, 502, 894–923. [Google Scholar] [CrossRef] [PubMed]
- Kaas, J.H. The evolution of brains from early mammals to humans. Wiley Interdiscip. Rev. Cogn. Sci. 2013, 4, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Amunts, K.; Mohlberg, H.; Bludau, S.; Zilles, K. Julich-Brain: A 3D probabilistic atlas of the human brain’s cytoarchitecture. Science 2020, 369, 988–992. [Google Scholar] [CrossRef] [PubMed]
- Eickhoff, S.B.; Schleicher, A.; Zilles, K.; Amunts, K. The human parietal operculum. I. Cytoarchitectonic mapping of subdivisions. Cereb. Cortex 2006, 16, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Eickhoff, S.B.; Amunts, K.; Mohlberg, H.; Zilles, K. The human parietal operculum. II. Stereotaxic maps and correlation with functional imaging results. Cereb. Cortex 2006, 16, 268–279. [Google Scholar] [CrossRef]
- Zachlod, D.; Rüttgers, B.; Bludau, S.; Mohlberg, H.; Langner, R.; Zilles, K.; Amunts, K. Four new cytoarchitectonic areas surrounding the primary and early auditory cortex in human brains. Cortex 2020, 128, 1–21. [Google Scholar] [CrossRef]
- Donos, C.; Mălîia, M.D.; Mîndruţă, I.; Popa, I.; Ene, M.; Bălănescu, B.; Ciurea, A.; Barborica, A. A connectomics approach combining structural and effective connectivity assessed by intracranial electrical stimulation. Neuroimage 2016, 132, 344–358. [Google Scholar] [CrossRef]
- Trebaul, L.; Deman, P.; Tuyisenge, V.; Jedynak, M.; Hugues, E.; Rudrauf, D.; Bhattacharjee, M.; Tadel, F.; Chanteloup-Foret, B.; Saubat, C.; et al. Probabilistic functional tractography of the human cortex revisited. Neuroimage 2018, 181, 414–429. [Google Scholar] [CrossRef]
- Lemaréchal, J.-D.; Jedynak, M.; Trebaul, L.; Boyer, A.; Tadel, F.; Bhattacharjee, M.; Deman, P.; Tuyisenge, V.; Ayoubian, L.; Hugues, E.; et al. A brain atlas of axonal and synaptic delays based on modelling of cortico-cortical evoked potentials. Brain J. Neurol. 2021, awab362. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, W.; Wang, S.; Zhou, Q.; Wang, H.; Zhang, B.; Huang, J.; Hong, B.; Wang, X. The Roles of Subdivisions of Human Insula in Emotion Perception and Auditory Processing. Cereb. Cortex 2019, 29, 517–528. [Google Scholar] [CrossRef]
- Augustine, J.R. The insular lobe in primates including humans. Neurol. Res. 1985, 7, 2–10. [Google Scholar] [CrossRef]
- Bamiou, D.E.; Musiek, F.E.; Luxon, L.M. The insula (Island of Reil) and its role in auditory processing. Literature review. Brain Res. Brain Res. Rev. 2003, 42, 143–154. [Google Scholar] [CrossRef]
- Wong, P.C.; Parsons, L.M.; Martinez, M.; Diehl, R.L. The role of the insular cortex in pitch pattern perception: The effect of linguistic contexts. J. Neurosci. 2004, 24, 9153–9160. [Google Scholar] [CrossRef] [PubMed]
- Bestelmeyer, P.E.; Maurage, P.; Rouger, J.; Latinus, M.; Belin, P. Adaptation to vocal expressions reveals multistep perception of auditory emotion. J. Neurosci. 2014, 34, 8098–8105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzola, L.; Mauguière, F.; Isnard, J. Electrical Stimulations of the Human Insula: Their Contribution to the Ictal Semiology of Insular Seizures. J. Clin. Neurophysiol. 2017, 34, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Trébuchon, A.; Alario, F.X.; Liégeois-Chauvel, C. Functional Topography of Auditory Areas Derived from the Combination of Electrophysiological Recordings and Cortical Electrical Stimulation. Front. Hum. Neurosci. 2021, 15, 702773. [Google Scholar] [CrossRef]
- Mălîia, M.D.; Donos, C.; Barborica, A.; Popa, I.; Ciurea, J.; Cinatti, S.; Mîndruţă, I. Functional mapping and effective connectivity of the human operculum. Cortex 2018, 109, 303–321. [Google Scholar] [CrossRef]
- Mazzola, L.; Mauguière, F.; Isnard, J. Functional mapping of the human insula: Data from electrical stimulations. Rev. Neurol. 2019, 175, 150–156. [Google Scholar] [CrossRef]
- Yu, K.; Yu, T.; Qiao, L.; Liu, C.; Wang, X.; Zhou, X.; Ni, D.; Zhang, G.; Li, Y. Electrical stimulation of the insulo-opercular region: Visual phenomena and altered body-ownership symptoms. Epilepsy Res. 2018, 148, 96–106. [Google Scholar] [CrossRef]
- Jaroszynski, C.; Amorim-Leite, R.; Deman, P.; Perrone-Bertolotti, M.; Chabert, F.; Job-Chapron, A.S.; Minotti, L.; Hoffmann, D.; David, O.; Kahane, P. Brain mapping of auditory hallucinations and illusions induced by Direct Intracortical Electrical stimulation. Neurology 2022. submitted. [Google Scholar]
- Matsumoto, R.; Kunieda, T.; Nair, D. Single pulse electrical stimulation to probe functional and pathological connectivity in epilepsy. Seizure 2017, 44, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trebaul, L.; Rudrauf, D.; Job, A.S.; Mălîia, M.D.; Popa, I.; Barborica, A.; Minotti, L.; Mîndruţă, I.; Kahane, P.; David, O. Stimulation artifact correction method for estimation of early cortico-cortical evoked potentials. J. Neurosci. Methods 2016, 264, 94–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friston, K.J. Functional and effective connectivity in neuroimaging: A synthesis. Hum. Brain Mapp. 1994, 2, 56–78. [Google Scholar] [CrossRef]
- Lemaréchal, J.D.; George, N.; David, O. Comparison of two integration methods for dynamic causal modeling of electrophysiological data. Neuroimage 2018, 173, 623–631. [Google Scholar] [CrossRef]
- Albouy, P.; Benjamin, L.; Morillon, B.; Zatorre, R.J. Distinct sensitivity to spectrotemporal modulation supports brain asymmetry for speech and melody. Science 2020, 367, 1043–1047. [Google Scholar] [CrossRef]
- Sepulcre, J. An OP4 functional stream in the language-related neuroarchitecture. Cereb. Cortex 2015, 25, 658–666. [Google Scholar] [CrossRef] [Green Version]
- Sepulcre, J.; Sabuncu, M.R.; Yeo, T.B.; Liu, H.; Johnson, K.A. Stepwise connectivity of the modal cortex reveals the multimodal organization of the human brain. J. Neurosci. 2012, 32, 10649–10661. [Google Scholar] [CrossRef]
- Tanaka, S.; Kirino, E. The parietal opercular auditory-sensorimotor network in musicians: A resting-state fMRI study. Brain Cogn. 2018, 120, 43–47. [Google Scholar] [CrossRef]
- Koelsch, S.; Cheung, V.K.M.; Jentschke, S.; Haynes, J.D. Neocortical substrates of feelings evoked with music in the ACC, insula, and somatosensory cortex. Sci. Rep. 2021, 11, 10119. [Google Scholar] [CrossRef]
- Pérez-Bellido, A.; Anne, B.K.; Crommett, L.E.; Yau, J.M. Auditory Frequency Representations in Human Somatosensory Cortex. Cereb. Cortex 2018, 28, 3908–3921. [Google Scholar] [CrossRef] [Green Version]
- Wager, T.D.; Atlas, L.Y.; Lindquist, M.A.; Roy, M.; Woo, C.W.; Kross, E. An fMRI-based neurologic signature of physical pain. N. Engl. J. Med. 2013, 368, 1388–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, L.Q.; Nomi, J.S.; Hébert-Seropian, B.; Ghaziri, J.; Boucher, O. Structure and Function of the Human Insula. J. Clin. Neurophysiol. 2017, 34, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Kurth, F.; Zilles, K.; Fox, P.T.; Laird, A.R.; Eickhoff, S.B. A link between the systems: Functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct. Funct. 2010, 214, 519–534. [Google Scholar] [CrossRef]
- Di Stefano, V.; De Angelis, M.V.; Montemitro, C.; Russo, M.; Carrarini, C.; di Giannantonio, M.; Brighina, F.; Onofrj, M.; Werring, D.J.; Simister, R. Clinical presentation of strokes confined to the insula: A systematic review of literature. Neurol. Sci. 2021, 42, 1697–1704. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Jbabdi, S.; Caspers, S.; Laird, A.R.; Fox, P.T.; Zilles, K.; Behrens, T.E. Anatomical and functional connectivity of cytoarchitectonic areas within the human parietal operculum. J. Neurosci. 2010, 30, 6409–6421. [Google Scholar] [CrossRef]
- Zu Eulenburg, P.; Caspers, S.; Roski, C.; Eickhoff, S.B. Meta-analytical definition and functional connectivity of the human vestibular cortex. Neuroimage 2012, 60, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.M.; Greenlee, M.W. The parieto-insular vestibular cortex in humans: More than a single area? J. Neurophysiol. 2018, 120, 1438–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göttlich, M.; Jandl, N.M.; Wojak, J.F.; Sprenger, A.; von der Gablentz, J.; Münte, T.F.; Krämer, U.M.; Helmchen, C. Altered resting-state functional connectivity in patients with chronic bilateral vestibular failure. Neuroimage Clin. 2014, 4, 488–499. [Google Scholar] [CrossRef] [Green Version]
- Veldhuizen, M.G.; Albrecht, J.; Zelano, C.; Boesveldt, S.; Breslin, P.; Lundström, J.N. Identification of human gustatory cortex by activation likelihood estimation. Hum. Brain Mapp. 2011, 32, 2256–2266. [Google Scholar] [CrossRef] [Green Version]
- Kober, S.E.; Grössinger, D.; Wood, G. Effects of Motor Imagery and Visual Neurofeedback on Activation in the Swallowing Network: A Real-Time fMRI Study. Dysphagia 2019, 34, 879–895. [Google Scholar] [CrossRef]
- Kumar, V.; Croxson, P.L.; Simonyan, K. Structural Organization of the Laryngeal Motor Cortical Network and Its Implication for Evolution of Speech Production. J. Neurosci. 2016, 36, 4170–4181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattendorf, E.; Westermann, B.; Fiedler, K.; Kaza, E.; Lotze, M.; Celio, M.R. Exploration of the neural correlates of ticklish laughter by functional magnetic resonance imaging. Cereb. Cortex 2013, 23, 1280–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotze, M.; Lucas, C.; Domin, M.; Kordass, B. The cerebral representation of temporomandibular joint occlusion and its alternation by occlusal splints. Hum. Brain Mapp. 2012, 33, 2984–2993. [Google Scholar] [CrossRef] [PubMed]
- Glasser, M.F.; Coalson, T.S.; Robinson, E.C.; Hacker, C.D.; Harwell, J.; Yacoub, E.; Ugurbil, K.; Andersson, J.; Beckmann, C.F.; Jenkinson, M.; et al. A multi-modal parcellation of human cerebral cortex. Nature 2016, 536, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Baker, C.M.; Burks, J.D.; Briggs, R.G.; Conner, A.K.; Glenn, C.A.; Robbins, J.M.; Sheets, J.R.; Sali, G.; McCoy, T.M.; Battiste, J.D.; et al. A Connectomic Atlas of the Human Cerebrum-Chapter 5: The Insula and Opercular Cortex. Oper. Neurosurg. 2018, 15, S175–S244. [Google Scholar] [CrossRef]
- Ghaziri, J.; Tucholka, A.; Girard, G.; Boucher, O.; Houde, J.C.; Descoteaux, M.; Obaid, S.; Gilbert, G.; Rouleau, I.; Nguyen, D.K. Subcortical structural connectivity of insular subregions. Sci. Rep. 2018, 8, 8596. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Lee, S.H. Perspective of functional magnetic resonance imaging in middle ear research. Hear. Res. 2013, 301, 183–192. [Google Scholar] [CrossRef]
- Kavounoudias, A.; Roll, J.P.; Anton, J.L.; Nazarian, B.; Roth, M.; Roll, R. Proprio-tactile integration for kinesthetic perception: An fMRI study. Neuropsychologia 2008, 46, 567–575. [Google Scholar] [CrossRef]
- Skog, C.; Fjellner, J.; Ekberg, E.; Häggman-Henrikson, B. Tinnitus as a comorbidity to temporomandibular disorders—A systematic review. J. Oral Rehabil. 2019, 46, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, T.G.; Rocha, C.B. Diagnosis and management of somatosensory tinnitus: Review article. Clinics 2011, 66, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Levine, R.A.; Abel, M.; Cheng, H. CNS somatosensory-auditory interactions elicit or modulate tinnitus. Exp. Brain Res. 2003, 153, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Shahsavarani, S.; Khan, R.A.; Husain, F.T. Tinnitus and the Brain: A Review of Functional and Anatomical Magnetic Resonance Imaging Studies. Perspect. ASHA Spec. Interest Groups 2019, 4, 896–909. [Google Scholar] [CrossRef]
- Khan, R.A.; Sutton, B.P.; Tai, Y.; Schmidt, S.A.; Shahsavarani, S.; Husain, F.T. A large-scale diffusion imaging study of tinnitus and hearing loss. Sci. Rep. 2021, 11, 23395. [Google Scholar] [CrossRef] [PubMed]
- Jaroszynski, C.; Attyé, A.; Job, A.; Delon-Martin, C. Tracking white-matter brain modifications in chronic non-bothersome acoustic trauma tinnitus. Neuroimage Clin. 2021, 31, 102696. [Google Scholar] [CrossRef]
- Jastreboff, P.J. 25 years of tinnitus retraining therapy. HNO 2015, 63, 307–311. [Google Scholar] [CrossRef]
- Simonetti, P.; Ono, C.R.; Godoi, C.C.; Ali, K.R.; Shahsavarani, S.; Husain, F.T.; Oiticica, J. Evaluating the efficacy of hearing aids for tinnitus therapy—A Positron emission tomography study. Brain Res. 2022, 1775, 147728. [Google Scholar] [CrossRef]
- Wei, X.; Lv, H.; Chen, Q.; Wang, Z.; Liu, C.; Zhao, P.; Gong, S.; Yang, Z.; Wang, Z. Cortical Thickness Alterations in Patients with Tinnitus Before and After Sound Therapy: A Surface-Based Morphometry Study. Front. Neurosci. 2021, 15, 633364. [Google Scholar] [CrossRef]
- Chen, Q.; Lv, H.; Wang, Z.; Wei, X.; Zhao, P.; Yang, Z.; Gong, S.; Wang, Z. Brain Structural and Functional Reorganization in Tinnitus Patients without Hearing Loss After Sound Therapy: A Preliminary Longitudinal Study. Front. Neurosci. 2021, 15, 573858. [Google Scholar] [CrossRef]
- Wei, X.; Lv, H.; Chen, Q.; Wang, Z.; Zhao, P.; Liu, C.; Gong, S.; Yang, Z.; Wang, Z. Surface-Based Amplitude of Low-Frequency Fluctuation Alterations in Patients with Tinnitus Before and After Sound Therapy: A Resting-State Functional Magnetic Resonance Imaging Study. Front. Neurosci. 2021, 15, 709482. [Google Scholar] [CrossRef]
- Krick, C.M.; Grapp, M.; Daneshvar-Talebi, J.; Reith, W.; Plinkert, P.K.; Bolay, H.V. Cortical reorganization in recent-onset tinnitus patients by the Heidelberg Model of Music Therapy. Front. Neurosci. 2015, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Lefaucheur, J.-P.; Aleman, A.; Baeken, C.; Benninger, D.H.; Brunelin, J.; Di Lazzaro, V.; Filipović, S.R.; Grefkes, C.; Hasan, A.; Hummel, F.C.; et al. Evidence-based guidelines on the therapeutic use of repetitive transcranial magnetic stimulation (rTMS): An update (2014–2018). Clin. Neurophysiol. 2020, 131, 474–528. [Google Scholar] [CrossRef] [PubMed]
- Jacquemin, L.; Mertens, G.; Shekhawat, G.S.; Van de Heyning, P.; Vanderveken, O.M.; Topsakal, V.; De Hertogh, W.; Michiels, S.; Beyers, J.; Moyaert, J.; et al. High Definition transcranial Direct Current Stimulation (HD-tDCS) for chronic tinnitus: Outcomes from a prospective longitudinal large cohort study. Prog. Brain Res. 2021, 263, 137–152. [Google Scholar] [PubMed]
- Rammo, R.; Ali, R.; Pabaney, A.; Seidman, M.; Schwalb, J. Surgical Neuromodulation of Tinnitus: A Review of Current Therapies and Future Applications. Neuromodulation 2019, 22, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Yakunina, N.; Kim, S.S.; Nam, E.C. BOLD fMRI effects of transcutaneous vagus nerve stimulation in patients with chronic tinnitus. PLoS ONE 2018, 13, e0207281. [Google Scholar] [CrossRef] [Green Version]
- Yakunina, N.; Nam, E.C. Direct and Transcutaneous Vagus Nerve Stimulation for Treatment of Tinnitus: A Scoping Review. Front. Neurosci. 2021, 15, 680590. [Google Scholar] [CrossRef]
- Conlon, B.; Langguth, B.; Hamilton, C.; Hughes, S.; Meade, E.; Connor, C.O.; Schecklmann, M.; Hall, D.A.; Vanneste, S.; Leong, S.L.; et al. Bimodal neuromodulation combining sound and tongue stimulation reduces tinnitus symptoms in a large randomized clinical study. Sci. Transl. Med. 2020, 12, eabb2830. [Google Scholar] [CrossRef]
- Balmer, T.S.; Trussell, L.O. Trigeminal Contributions to the Dorsal Cochlear Nucleus in Mouse. Front. Neurosci. 2021, 15, 715954. [Google Scholar] [CrossRef]
- Shore, S.E.; Wu, C. Mechanisms of noise-induced tinnitus: Insights from cellular studies. Neuron 2019, 103, 8–20. [Google Scholar] [CrossRef]
- Lee, S.Y.; Rhee, J.; Shim, Y.J.; Kim, Y.; Koo, J.W.; De Ridder, D.; Vanneste, S.; Song, J.J. Changes in the Resting-State Cortical Oscillatory Activity 6 Months After Modified Tinnitus Retraining Therapy. Front. Neurosci. 2019, 13, 1123. [Google Scholar] [CrossRef]
- Zimmerman, B.; Finnegan, M.; Paul, S.; Schmidt, S.; Tai, Y.; Roth, K.; Chen, Y.; Husain, F.T. Functional Brain Changes During Mindfulness-Based Cognitive Therapy Associated with Tinnitus Severity. Front. Neurosci. 2019, 13, 747. [Google Scholar] [CrossRef]
- Emmert, K.; Kopel, R.; Koush, Y.; Maire, R.; Senn, P.; Van De Ville, D.; Haller, S. Continuous vs. intermittent neurofeedback to regulate auditory cortex activity of tinnitus patients using real-time fMRI—A pilot study. Neuroimage Clin. 2017, 14, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, M.S.; Parker, J.G.; Diller, E.E.; Ganapathy, S.; Bennett, K.B.; Esquivel, C.R.; Nelson, J.T. Self-directed down-regulation of auditory cortex activity mediated by real-time fMRI neurofeedback augments attentional processes, resting cerebral perfusion, and auditory activation. Neuroimage 2019, 195, 475–489. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaroszynski, C.; Job, A.; Jedynak, M.; David, O.; Delon-Martin, C. Tinnitus Perception in Light of a Parietal Operculo–Insular Involvement: A Review. Brain Sci. 2022, 12, 334. https://doi.org/10.3390/brainsci12030334
Jaroszynski C, Job A, Jedynak M, David O, Delon-Martin C. Tinnitus Perception in Light of a Parietal Operculo–Insular Involvement: A Review. Brain Sciences. 2022; 12(3):334. https://doi.org/10.3390/brainsci12030334
Chicago/Turabian StyleJaroszynski, Chloé, Agnès Job, Maciej Jedynak, Olivier David, and Chantal Delon-Martin. 2022. "Tinnitus Perception in Light of a Parietal Operculo–Insular Involvement: A Review" Brain Sciences 12, no. 3: 334. https://doi.org/10.3390/brainsci12030334
APA StyleJaroszynski, C., Job, A., Jedynak, M., David, O., & Delon-Martin, C. (2022). Tinnitus Perception in Light of a Parietal Operculo–Insular Involvement: A Review. Brain Sciences, 12(3), 334. https://doi.org/10.3390/brainsci12030334