

On the Application of Developmental Cognitive Neuroscience in Educational Environments


Abstract
1. Introduction
Cognitive and Educational Neurosciences in Co-Development: Seeking a Common Language
2. Neuroanatomical and Neurophysiological Relevancies and Irrelevancies in Learning and Classroom Settings
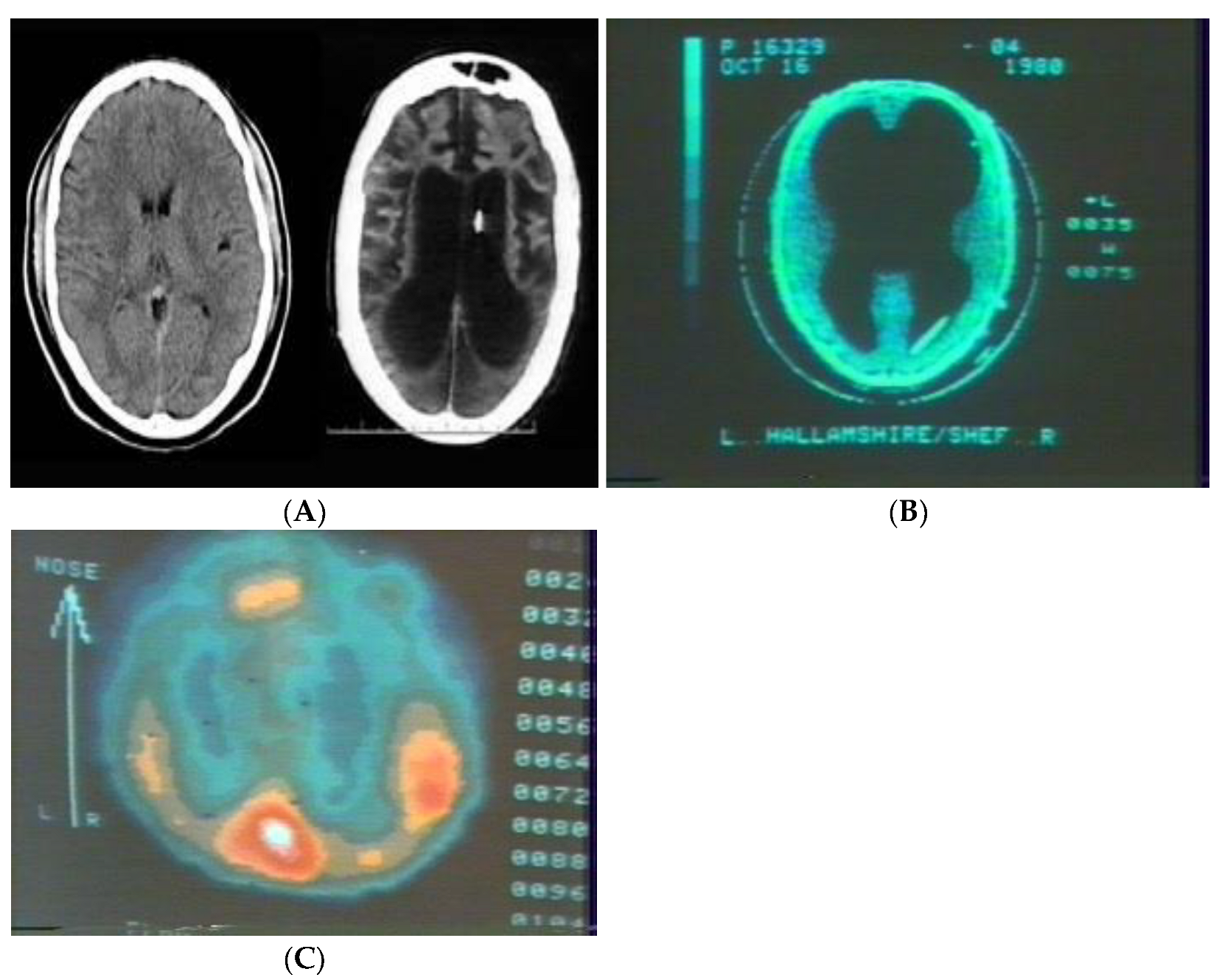
2.1. Brain Anatomy Is Irrelevant to Educational Practice but Not Functional Connectivity
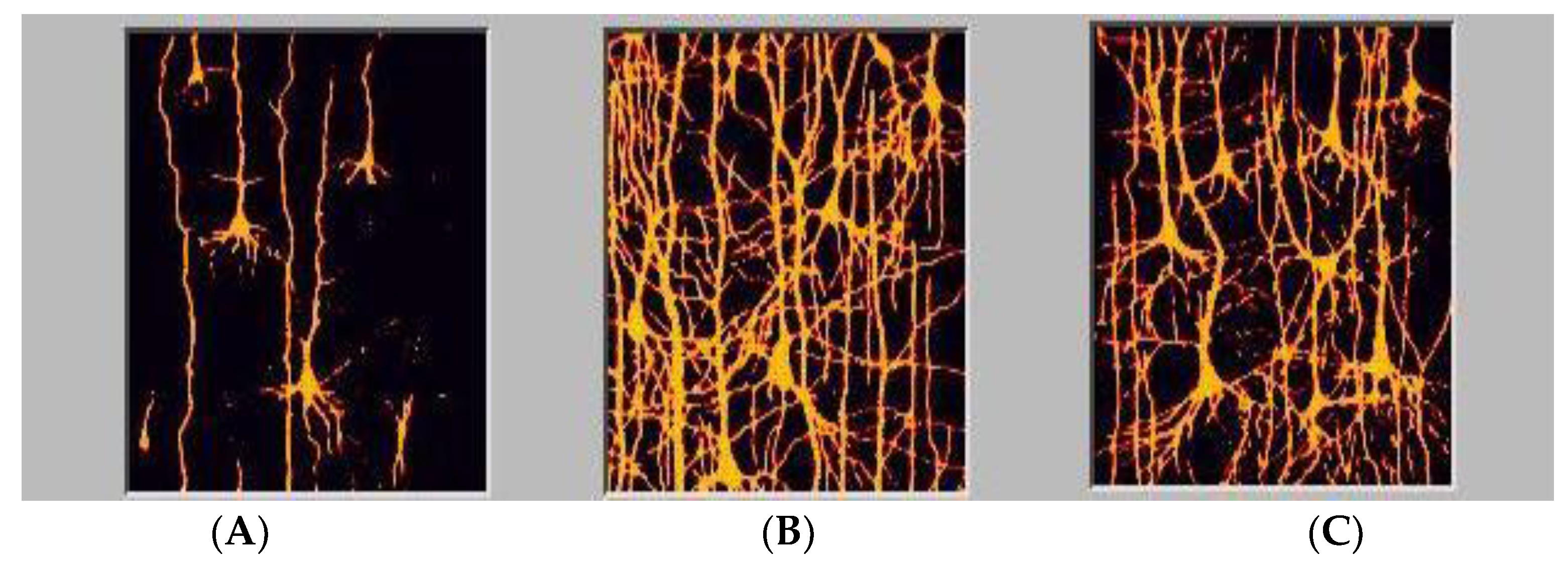
2.2. Plasticity and Connectivity in Brain Networking: A Basis for Child and Adult Education
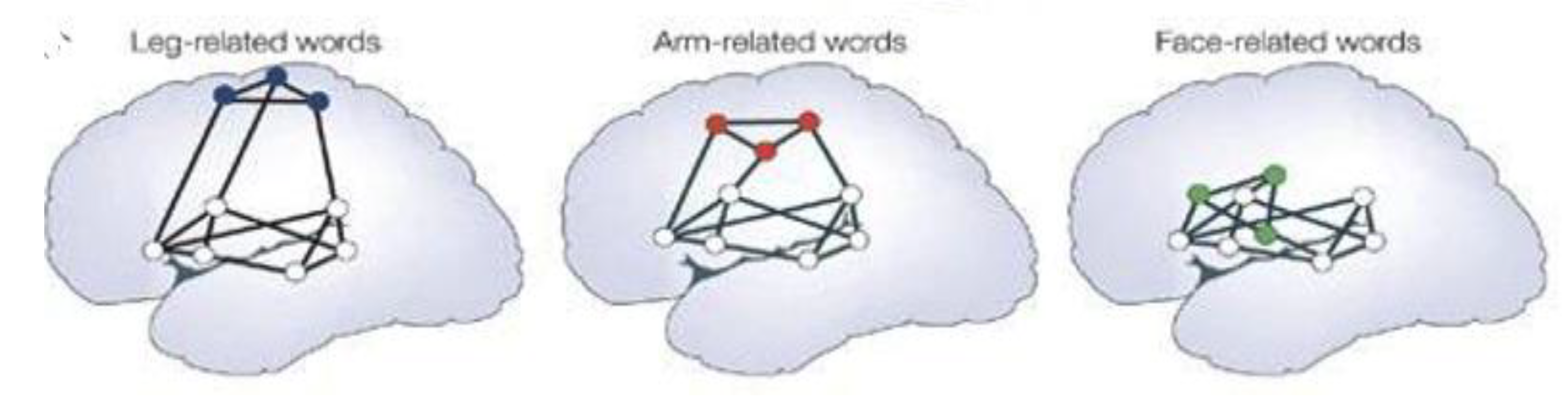
2.3. Functional Connectivities Creating Efficiencies of Response: Creating Associative Educational Networks
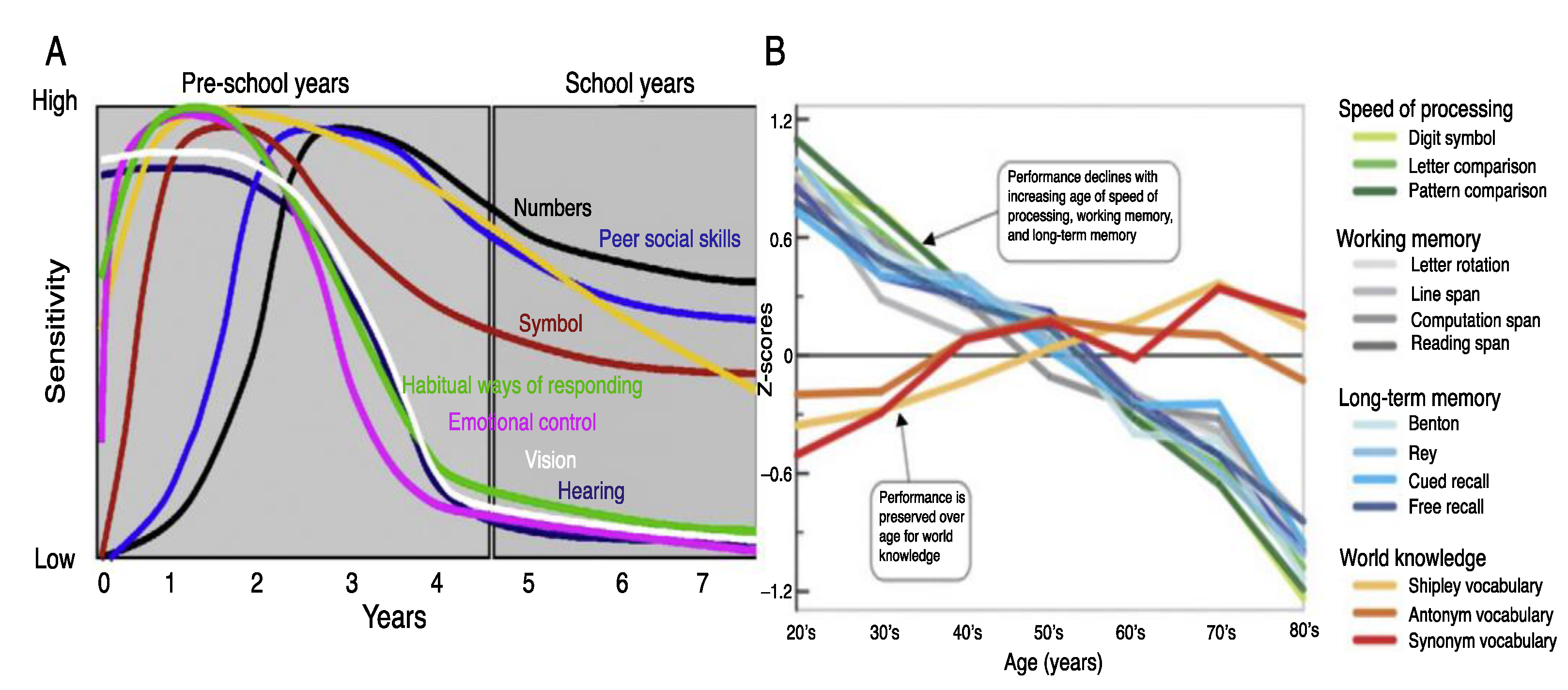
2.4. The Timing of Network Building: Critical and Sensitive Periods in Neurocognitive Development during Preschool and School Years

2.5. Timing, Associations, and the Building of Learning Networks
3. Facilitating Neuronal Connections in Classroom Learning
3.1. The Importance of Connections: Learning by Association and Context
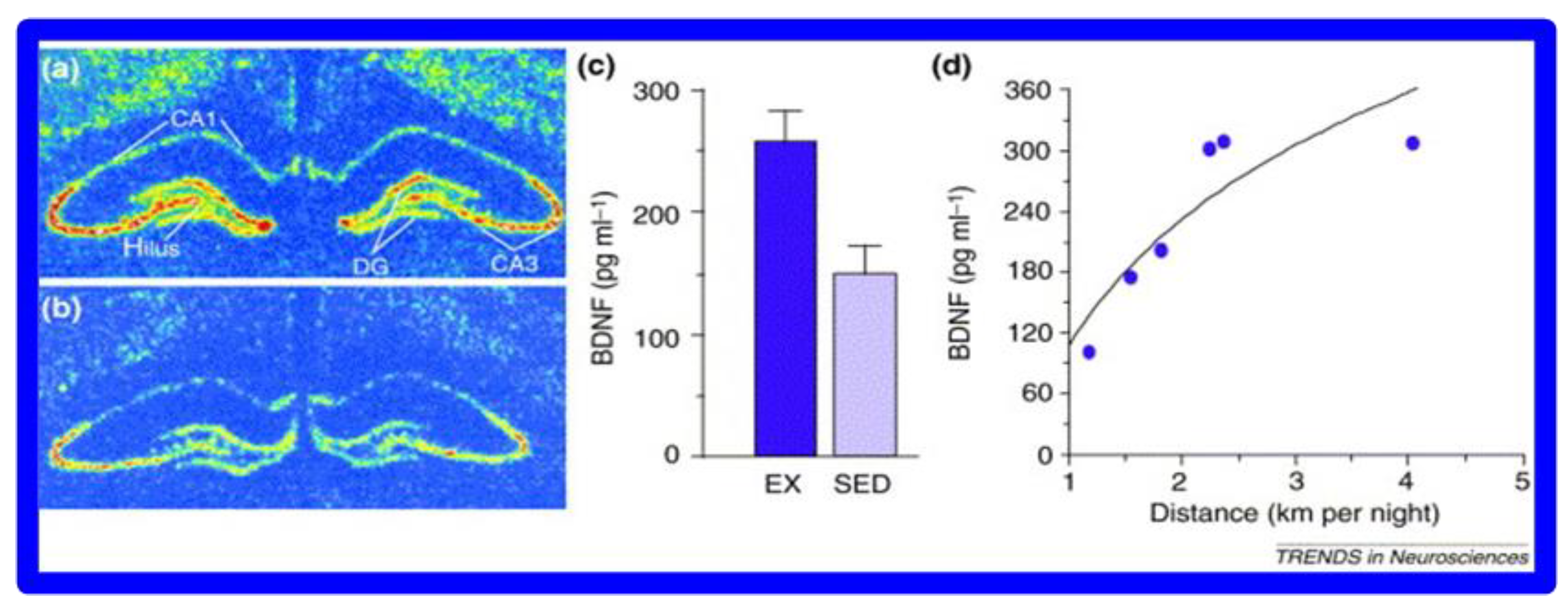
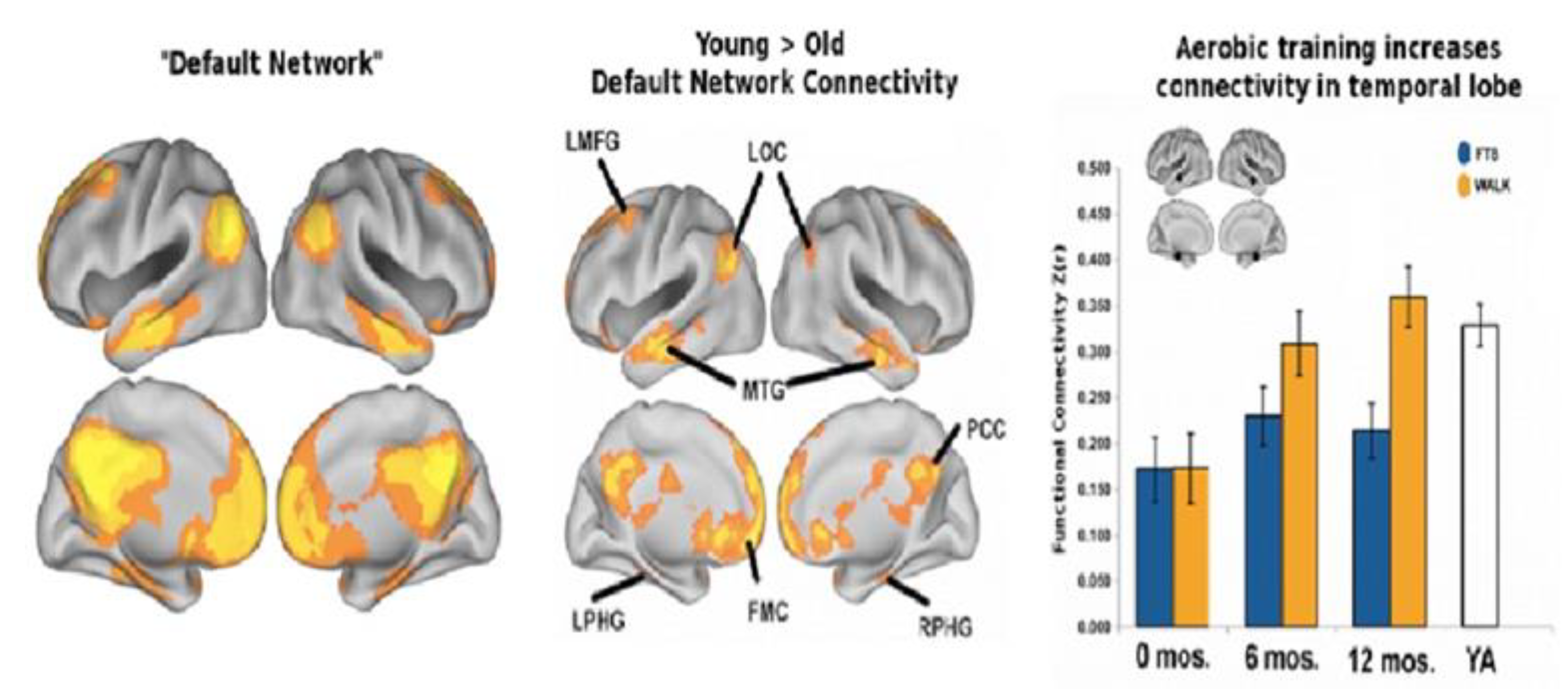
3.2. Physical Activity and Cognitive Relationships
3.3. Implications of the Development and Plasticity of the Neural Connectivities for Learning and Instruction
3.3.1. State of Our Understanding of Brain-Based Learning
3.3.2. More Comprehensive Understanding of the Power behind the Spinal Cord and Lower Parts of the Brain, Especially the Cerebellum in Classroom Learning
4. Neuronal Systems Known to Facilitate Classroom Learning
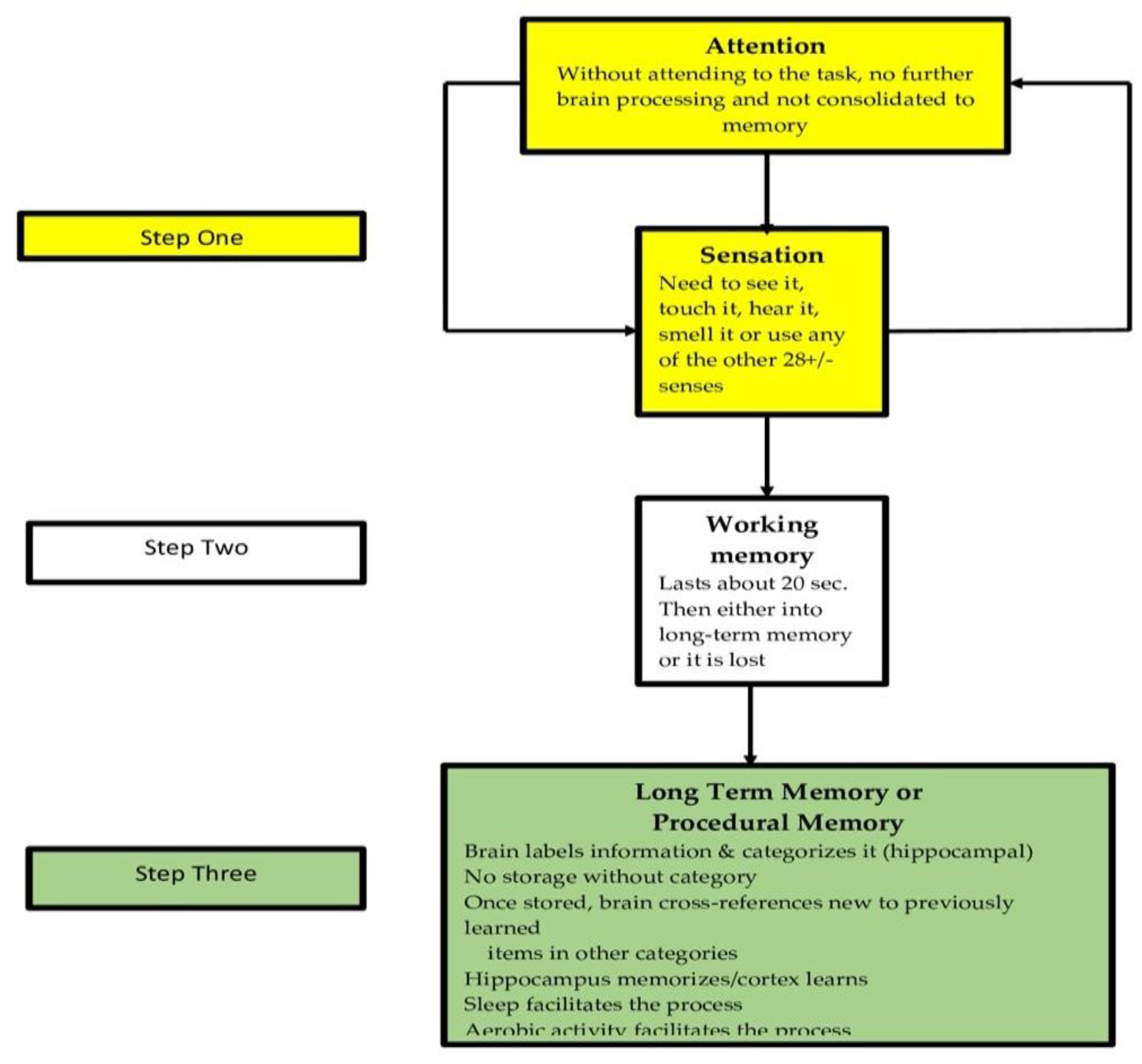
4.1. Attention
4.2. Emotional Hooks in Learning
4.3. Complexity v. Difficulty
4.4. Creativity: A Principal Goal of Instruction Is to Be Able to Use Knowledge in Different Settings
4.5. Memory Considerations in School-Based Learning
5. Discussion
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ansari, D.; Coch, D. Bridges over troubled waters: Education and cognitive neuroscience. Trends Cogn. Sci. 2006, 10, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Sigman, M.; Peña, M.; Goldin, A.P.; Ribeiro, S. Neuroscience and education: Prime time to build the bridge. Nat. Neurosci. 2014, 17, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, G.; Morett, L.M.; Hawes, Z.; Dommett, E.J. Why educational neuroscience needs educational and school psychology to effectively translate neuroscience to educational practice. Front. Psychol. 2021, 11, 618449. [Google Scholar] [CrossRef]
- Leisman, G. Coherence of hemispheric function in developmental dyslexia. Brain Cogn. 2002, 48, 425–431. [Google Scholar]
- Shaywitz, S.E.; Shaywitz, B.A. Paying attention to reading: The neurobiology of reading and dyslexia. Dev. Psychopathol. 2008, 20, 1329–1349. [Google Scholar] [CrossRef] [PubMed]
- Goswami, U. Neuroscience and education: From research to practice? Nat. Rev. Neurosci. 2006, 7, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Carew, T.J.; Magsamen, S.H. Neuroscience and education: An ideal partnership for producing evidence-based solutions to guide 21st century learning. Neuron 2010, 67, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Willingham, D.T. Three problems in the marriage of neuroscience and education. Cortex 2009, 45, 544. [Google Scholar] [CrossRef]
- Devonshire, I.M.; Dommett, E.J. Neuroscience: Viable applications in education? Neurosci. 2010, 16, 349–356. [Google Scholar] [CrossRef]
- Chiu, Y.C.; Egner, T. Cortical and subcortical contributions to context-control learning. Neurosci. Biobehav. Rev. 2019, 99, 33–41. [Google Scholar] [CrossRef]
- Kindell, C. Self-Directed Pedagogy and Visual Learning. Doctoral Dissertation, Florida Southern College, Lakeland, FL, USA, 2019. Available online: https://repository.flsouthern.edu/handle/11416/471 (accessed on 11 September 2022).
- Lindsley, D.B. Neural mechanisms of arousal, attention, and information processing. In Neuropsychology after Lashley; Routledge: London, UK, 2018; pp. 315–408. [Google Scholar]
- Osher, D.; Cantor, P.; Berg, J.; Steyer, L.; Rose, T. Drivers of human development: How relationships and context shape learning and development1. Appl. Dev. Sci. 2020, 24, 6–36. [Google Scholar] [CrossRef]
- Sousa, D.A. How the Brain Learns; Corwin Press: Thousand Oaks, CA, USA, 2016; p. 28. [Google Scholar]
- Cohen, S.S.; Madsen, J.; Touchan, G.; Robles, D.; Lima, S.F.; Henin, S.; Parra, L.C. Neural engagement with online educational videos predicts learning performance for individual students. Neurobiol. Learn. Mem. 2018, 155, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Feldhacker, D.R.; Cosgrove, R.; Feiten, B.; Schmidt, K.; Stewart, M. The Correlation between Retained Primitive Reflexes and Scholastic Performance among Early Elementary Students. J. Occup. Ther. Sch. Early Interv. 2021, 15, 288–301. [Google Scholar] [CrossRef]
- Russo, N.; Kaplan-16 Kahn, E.A.; Wilson, J.; Criss, A.; Burack, J.A. Choices, challenges, and constraints: A pragmatic examination of the limits of mental age matching in empirical research. Dev. Psychopathol. 2021, 33, 727–738. [Google Scholar] [CrossRef]
- Borghans, L.; Golsteyn, B.H.; Heckman, J.J.; Humphries, J.E. What grades and achievement tests measure. Proc. Natl. Acad. Sci. USA 2016, 113, 13354–13359. [Google Scholar] [CrossRef] [PubMed]
- Cohen, L.; Manion, L.; Morrison, K. Tests. In Research Methods in Education; Routledge: London, UK, 2017; pp. 563–585. [Google Scholar]
- Johnson, M.H. Interactive specialization: A domain-general framework for human functional brain development? Dev. Cogn. Neurosci. 2011, 1, 7–21. [Google Scholar] [CrossRef]
- Leisman, G.; Mualem, R.; Mughrabi, S.K. The neurological development of the child with the educational enrichment in mind. Psicol. Educ. 2015, 21, 79–96. [Google Scholar] [CrossRef]
- Melillo, R.; Leisman, G. Neurobehavioral Disorders of Childhood: An Evolutionary Perspective; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Ertl, J.P.; Schafer, E.W. Brain response correlates of psychometric intelligence. Nature 1969, 223, 421–422. [Google Scholar] [CrossRef]
- Grabner, R.H.; Stern, E.; Neubauer, A.C. When intelligence loses its impact: Neural efficiency during reasoning in a familiar area. Int. J. Psychophysiol. 2003, 49, 89–98. [Google Scholar] [CrossRef]
- Neubauer, A.C.; Grabner, R.H.; Freudenthaler, H.H.; Beckmann, J.F.; Guthke, J. Intelligence and individual differences in becoming neurally efficient. Acta Psychol. 2004, 116, 55–74. [Google Scholar] [CrossRef]
- Rypma, B.; Berger, J.S.; Prabhakaran, V.; Bly, B.M.; Kimberg, D.Y.; Biswal, B.B.; D’Esposito, M. Neural correlates of cognitive efficiency. Neuroimage 2006, 33, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, B. Cognitive efficiency: A conceptual and methodological comparison. Learn. Instr. 2012, 22, 133–144. [Google Scholar] [CrossRef]
- Neubauer, A.C.; Fink, A. Intelligence and neural efficiency. Neurosci. Biobehav. Rev. 2009, 33, 1004–1023. [Google Scholar] [CrossRef]
- Parks, R.W.; Loewenstein, D.A.; Dodrill, K.L.; Barker, W.W.; Yoshii, F.; Chang, J.Y.; Emran, A.; Apicella, A.; Sheramata, W.A.; Duara, R. Cerebral metabolic effects of a verbal fluency test: A PET scan study. J. Clin. Exp. Neuropsychol. 1988, 10, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Schultz, D.H.; Cole, M.W. Higher intelligence is associated with less task-related brain network reconfiguration. J. Neurosci. 2016, 36, 8551–8561. [Google Scholar] [CrossRef] [PubMed]
- Causse, M.; Chua, Z.; Peysakhovich, V.; Del Campo, N.; Matton, N. Mental workload and neural efficiency quantified in the prefrontal cortex using fNIRS. Sci. Rep. 2017, 7, 5222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gan, J.Q.; Wang, H. Neurocognitive mechanisms of mathematical giftedness: A literature review. Appl. Neuropsychol. Child 2017, 6, 79–94. [Google Scholar] [CrossRef]
- Collell, G.; Fauquet, J. Brain activity and cognition: A connection from thermodynamics and information theory. Front. Psychol. 2015, 6, 818. [Google Scholar] [CrossRef]
- Manza, P.; Wiers, C.E.; Shokri-Kojori, E.; Kroll, D.; Feldman, D.; Schwandt, M.; Wang, G.J.; Tomasi, D.; Volkow, N.D. Brain network segregation and glucose energy utilization: Relevance for age-related differences in cognitive function. Cereb. Cortex 2020, 30, 5930–5942. [Google Scholar] [CrossRef]
- Grabner, R.H.; Fink, A.; Stipacek, A.; Neuper, C.; Neubauer, A.C. Intelligence and working memory systems: Evidence of neural efficiency in alpha band ERD. Cogn. Brain Res. 2004, 20, 212–225. [Google Scholar] [CrossRef]
- Duchowny, M.; Jayakar, P.; Harvey, A.S.; Resnick, T.; Alvarez, L.; Dean, P.; Levin, B. Language cortex representation: Effects of developmental versus acquired pathology. Ann. Neurol. 1996, 40, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Olulade, O.A.; Seydell-Greenwald, A.; Chambers, C.E.; Turkeltaub, P.E.; Dromerick, A.W.; Berl, M.M.; Gaillard, W.D.; Newport, E.L. The neural basis of language development: Changes in laterali- zation over age. Proc. Natl. Acad. Sci. USA 2020, 117, 23477–23483. [Google Scholar] [CrossRef] [PubMed]
- Josselyn, S.A.; Tonegawa, S. Memory engrams: Recalling the past and imagining the future. Science 2020, 367, eaaw4325. [Google Scholar] [CrossRef] [PubMed]
- Palmiero, M.; Piccardi, L. Is visual creativity embodied? Thinking aloud while performing the creative mental synthesis task. Brain Sci. 2020, 10, 455. [Google Scholar] [CrossRef]
- Glover, S.; Baran, M. The motor-cognitive model of motor imagery: Evidence from timing errors in simulated reaching and grasping. J. Exp. Psychol. Hum. Percept. Perform. 2017, 43, 1359. [Google Scholar] [CrossRef]
- Karklinsky, M.; Flash, T. Timing of continuous motor imagery: The two-thirds power law originates in trajectory planning. J. Neurophysiol. 2015, 113, 2490–2499. [Google Scholar] [CrossRef]
- Kaller, M.S.; Lazari, A.; Blanco-Duque, C.; Sampaio-Baptista, C.; Johansen-Berg, H. Myelin plas- ticity and behaviour—Connecting the dots. Curr. Opin. Neurobiol. 2017, 47, 86–92. [Google Scholar] [CrossRef]
- Koch, P.; Leisman, G. Effect of local synaptic strengthening on global activity- wave growth in the hippocampus. J. Neurosci. 2001, 108, 127–146. [Google Scholar] [CrossRef]
- Koch, P.; Leisman, G. Cortical Activity Waves are the Physical Carriers of Memory and Thought. In Proceedings of the 7th Annual IEEE Engineering in Medicine and Biology Society Neural Engineering Conference, Montpellier, France, 22–24 April 2015. [Google Scholar] [CrossRef]
- Schaefers, A.T.; Teuchert-Noodt, G. Developmental neuroplasticity and the origin of neuro-degenerative diseases. World J. Biol. Psychiatry 2016, 17, 587–599. [Google Scholar]
- Costandi, M. Neuroplasticity; MIT Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Gelfo, F. Does experience enhance cognitive flexibility? An overview of the evidence provided by the environmental enrichment studies. Front. Behav. Neurosci. 2019, 13, 150. [Google Scholar] [CrossRef]
- Dehorter, N.; Del Pino, I. Shifting developmental trajectories during critical periods of brain formation. Front. Cell. Neurosci. 2020, 14, 283. [Google Scholar] [CrossRef] [PubMed]
- Khazipov, R.; Luhmann, H.J. Early patterns of electrical activity in the developing cerebral cortex of humans and rodents. Trends Neurosci. 2006, 29, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Schlaug, G.; Jäncke, L.; Huang, Y.; Staiger, J.F.; Steinmetz, H. Increased corpus callosum size in musicians. Neuropsychologia 1995, 33, 1047–1055. [Google Scholar] [CrossRef]
- Flohr, J.W. Best practices for young children’s music education: Guidance from brain research. Gen. Music. Today 2010, 23, 13–19. [Google Scholar] [CrossRef]
- Flohr, J.W.; Trevarthen, C. Music learning in childhood–Early developments of a musical brain body In Neurosciences in Music Pedagogy; Gruhn, W., Rauscher, F., Eds.; Nova Science Publishers: Hauppage, NY, USA, 2008; pp. 53–99. [Google Scholar]
- Barnett, S.M.; Ceci, S.J. When and where do we apply what we learn?: A taxonomy for far transfer. Psychol. Bull. 2002, 128, 612. [Google Scholar] [CrossRef]
- Bremner, A.J.; Lewkowicz, D.J.; Spence, C. (Eds.) Multisensory Development; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Lewkowicz, D.J. Early experience and multisensory perceptual narrowing. Dev. Psychobiol. 2014, 56, 292–315. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.M.; Lewkowicz, D.J.; Amedi, A.; Wallace, M.T. Multisensory processes: A balancing act across the lifespan. Trends Neurosci. 2016, 39, 567–579. [Google Scholar] [CrossRef] [PubMed]
- Power, J.D.; Schlaggar, B.L. Neural plasticity across the lifespan. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6, e216. [Google Scholar] [CrossRef]
- Zenke, F.; Gerstner, W.; Ganguli, S. The temporal paradox of Hebbian learning and homeostatic plasticity. Curr. Opin. Neurobiol. 2017, 42, 166–176. [Google Scholar] [CrossRef]
- Kiyota, T. Neurogenesis and brain repair. In Neuroimmune Pharmacology; Springer: Cham, Switzerland, 2017; pp. 575–597. [Google Scholar]
- Quadrato, G.; Elnaggar, M.Y.; Di Giovanni, S. Adult neurogenesis in brain repair: Cellular plasticity vs. cellular replacement. Front. Neurosci. 2014, 8, 17. [Google Scholar] [CrossRef]
- Hartshorne, J.K. How massive online experiments (MOEs) can illuminate critical and sensitive periods in development. Curr. Opin. Behav. Sci. 2020, 36, 135–143. [Google Scholar] [CrossRef]
- Thompson, A.; Steinbeis, N. Sensitive periods in executive function development. Curr. Opin. Behav. Sci. 2020, 36, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Silvano, E.; Bedny, M. Sensitive periods in cortical specialization for language: Insights from studies with deaf and blind individuals. Curr. Opin. Behav. Sci. 2020, 36, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Hebb, D.O. The Organization of Behavior: A Neuropsychological Theory; Psychology Press: London, UK, 2005. [Google Scholar]
- Van Den Heuvel, M.P.; Stam, C.J.; Kahn, R.S.; Pol, H.E. Efficiency of functional brain networks and intellectual performance. J. Neurosci. 2009, 29, 7619–7624. [Google Scholar] [CrossRef]
- Masís-Obando, R.; Norman, K.A.; Baldassano, C. Schema representations in distinct brain networks support narrative memory during encoding and retrieval. Elife 2022, 11, e70445. [Google Scholar] [CrossRef]
- Posner, M.I.; Rothbart, M.K. Influencing brain networks: Implications for education. Trends Cogn. Sci. 2005, 9, 99–103. [Google Scholar] [CrossRef]
- Quiroga, R.Q.; Reddy, L.; Kreiman, G.; Koch, C.; Fried, I. Invariant visual representation by single neurons in the human brain. Nature 2005, 435, 1102–1107. [Google Scholar] [CrossRef]
- Martin, C.B.; Douglas, D.; Newsome, R.N.; Man, L.L.; Barense, M.D. Integrative and distinctive coding of visual and conceptual object features in the ventral visual stream. Elife 2018, 7, e31873. [Google Scholar] [CrossRef]
- Rissman, J.; Wagner, A.D. Distributed representations in memory: Insights from functional brain imaging. Annu. Rev. Psychol. 2012, 63, 101. [Google Scholar] [CrossRef]
- Chen, Y.; Shimotake, A.; Matsumoto, R.; Kunieda, T.; Kikuchi, T.; Miyamoto, S.; Fukuyama, H.; Takahashi, R.; Ikeda, A.; Ralph, M.L. The ‘when’ and ‘where’ of semantic coding in the anterior temporal lobe: Temporal representational similarity analysis of electrocorticogram data. Cortex 2016, 79, 1–13. [Google Scholar] [CrossRef]
- Gao, Z.; Zheng, L.; Chiou, R.; Gouws, A.; Krieger-Redwood, K.; Wang, X.; Varga, D.; Ralph, M.A.; Smallwood, J.; Jefferies, E. Distinct and common neural coding of semantic and non-semantic control demands. NeuroImage 2021, 236, 118230. [Google Scholar] [CrossRef] [PubMed]
- Breedlove, J.L.; St-Yves, G.; Olman, C.A.; Naselaris, T. Generative feedback explains distinct brain activity codes for seen and mental images. Curr. Biol. 2020, 30, 2211–2224. [Google Scholar] [CrossRef] [PubMed]
- Gallese, V. A neuroscientific grasp of concepts: From control to representation. Philosophical Transactions of the Royal Society of London. Ser. B Biol. Sci. 2003, 358, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Solé, R.V.; Corominas-Murtra, B.; Valverde, S.; Steels, L. Language networks: Their structure, function, and evolution. Complexity 2010, 15, 20–26. [Google Scholar] [CrossRef]
- Willis, J. Review of research: Brain-based teaching strategies for improving students' memory, learning, and test-taking success. Child. Educ. 2007, 83, 310–315. [Google Scholar] [CrossRef]
- Leshinskaya, A.; Caramazza, A. For a cognitive neuroscience of concepts: Moving beyond the grounding issue. Psychon. Bull. Rev. 2016, 23, 991–1001. [Google Scholar]
- Newman, M.E.J. Networks, 2nd ed.; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Cotman, C.W.; Berchtold, N.C. Exercise: A behavioral intervention to enhance brain health and plasticity. Trends Neurosci. 2002, 25, 295–301. [Google Scholar] [CrossRef]
- Voss, M.W.; Prakash, R.S.; Erickson, K.I.; Basak, C.; Chaddock, L.; Kim, J.S.; Alves, H.; Heo, S.; Szabo, A.N.; White, S.M.; et al. Plasticity of brain networks in a randomized intervention trial of exercise training in older adults. Front. Aging Neurosci. 2010, 2, 32. [Google Scholar] [CrossRef]
- Quartz, S.R.; Sejnowski, T.J. The neural basis of cognitive development: A constructivist manifesto. Behav. Brain Sci. 1997, 20, 537–556. [Google Scholar] [CrossRef]
- Shin, D.D.; Lee, M.; Bong, M. Beyond Left and Right: Learning Is a Whole-Brain Process. Theory Pract. 2022, 61, 347–357. [Google Scholar] [CrossRef]
- Fişek, M.; Häusser, M. Are Human Dendrites Different? Trends Cogn. Sci. 2020, 24, 411–412. [Google Scholar] [CrossRef] [PubMed]
- Maquet, P. The role of sleep in learning and memory. Science 2001, 294, 1048–1052. [Google Scholar] [CrossRef] [PubMed]
- Curcio, G.; Ferrara, M.; De Gennaro, L. Sleep loss, learning capacity and academic performance. Sleep Med. Rev. 2006, 10, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.O.; Ghosh, A. Activity-dependent regulation of dendritic growth and patterning. Nat. Rev. Neurosci. 2002, 3, 803–812. [Google Scholar] [CrossRef]
- Biernaskie, J.; Corbett, D. Enriched rehabilitative training promotes improved forelimb motor function and enhanced dendritic growth after focal ischemic injury. J. Neurosci. 2001, 21, 5272–5280. [Google Scholar] [CrossRef]
- Rushton, S.; Juola-Rushton, A.; Larkin, E. Neuroscience, play and early childhood education: Connections, implications and assessment. Early Child. Educ. J. 2010, 37, 351–361. [Google Scholar] [CrossRef]
- Robison, L.S.; Francis, N.; Popescu, D.L.; Anderson, M.E.; Hatfield, J.; Xu, F.; Anderson, B.J.; Van Nostrand, W.E.; Robinson, J.K. Environmental enrichment: Disentangling the influence of novelty, social, and physical activity on cerebral amyloid angiopathy in a transgenic mouse model. Int. J. Mol. Sci. 2020, 21, 843. [Google Scholar] [CrossRef]
- Evans-Martin, F.F. Emotion and Stress; Infobase Publishing: New York, NY, USA, 2009. [Google Scholar]
- Riccomagno, M.M.; Kolodkin, A.L. Sculpting neural circuits by axon and dendrite pruning. Annu. Rev. Cell Dev. Biol. 2015, 31, 779. [Google Scholar] [CrossRef]
- Ho, T.C.; King, L.S. Mechanisms of neuroplasticity linking early adversity to depression: Developmental considerations. Transl. Psychiatry 2021, 11, 517. [Google Scholar] [CrossRef]
- Navlakha, S.; Bar-Joseph, Z.; Barth, A.L. Network design and the brain. Trends Cogn. Sci. 2018, 22, 64–78. [Google Scholar] [CrossRef]
- Price, R.B.; Duman, R. Neuroplasticity in cognitive and psychological mechanisms of depression: An integrative model. Mol. Psychiatry 2020, 25, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Llinas, R.R.; Walton, K.D.; Lang, E.J. Chapter 7 Cerebellum. In The Synaptic Organization of the Brain; Shepherd, G.M., Ed.; Oxford University Press: New York, NY, USA, 2004. [Google Scholar]
- Doya, K. Complementary roles of basal ganglia and cerebellum in learning and motor control. Curr. Opin. Neurobiol. 2000, 10, 732–739. [Google Scholar] [CrossRef]
- Rapp, B. Handbook of Cognitive Neuropsychology: What Deficits Reveal about the Human Mind; Psychology Press: London, UK, 2015. [Google Scholar]
- Striemer, C.L.; Chouinard, P.A.; Goodale, M.A.; De Ribaupierre, S. Overlapping neural circuits for visual attention and eye movements in the human cerebellum. Neuropsychologia 2015, 69, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Goetz, M.; Schwabova, J.P.; Hlavka, Z.; Ptacek, R.; Surman, C.B. Dynamic balance in children with attention-deficit hyperactivity disorder and its relationship with cognitive functions and cerebellum. Neuropsychiatr. Dis. Treat. 2017, 13, 873. [Google Scholar] [CrossRef]
- Argyropoulos, G.P. The cerebellum, internal models and prediction in ‘non-motor’aspects of language: A critical review. Brain Lang. 2016, 161, 4–17. [Google Scholar] [CrossRef]
- Runnqvist, E.; Bonnard, M.; Gauvin, H.S.; Attarian, S.; Trébuchon, A.; Hartsuiker, R.J.; Alario, F.X. Internal modeling of upcoming speech: A causal role of the right posterior cerebellum in non-motor aspects of language production. Cortex 2016, 81, 203–214. [Google Scholar] [CrossRef]
- Tian, X.; Zarate, J.M.; Poeppel, D. Mental imagery of speech implicates two mechanisms of perceptual reactivation. Cortex 2016, 77, 1–12. [Google Scholar] [CrossRef]
- Kilteni, K.; Andersson, B.J.; Houborg, C.; Ehrsson, H.H. Motor imagery involves predicting the sensory consequences of the imagined movement. Nat. Commun. 2018, 9, 1617. [Google Scholar] [CrossRef]
- Xue, A.; Kong, R.; Yang, Q.; Eldaief, M.C.; Angeli, P.A.; DiNicola, L.M.; Braga, R.M.; Buckner, R.L.; Yeo, B.T. The detailed organization of the human cerebellum estimated by intrinsic functional connectivity within the individual. J. Neurophysiol. 2021, 125, 358–384. [Google Scholar] [CrossRef]
- Hernáez-Goñi, P.; Tirapu-Ustárroz, J.; Iglesias-Fernández, L.; Luna-Lario, P. Participación del cerebelo en la regulación del afecto, la emoción y la conducta. Rev. Neurol. 2010, 51, 597–609. [Google Scholar] [CrossRef]
- Turner, B.M.; Paradiso, S.; Marvel, C.L.; Pierson, R.; Ponto, L.L.; Hichwa, R.D.; Robinson, R.G. The cerebellum and emotional experience. Neuropsychologia 2007, 45, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D. The cerebellum and cognition. Neurosci. Lett. 2019, 688, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Van Overwalle, F.; Manto, M.; Cattaneo, Z.; Clausi, S.; Ferrari, C.; Gabrieli, J.D.; Guell, X.; Heleven, E.; Lupo, M.; Ma, Q.; et al. Consensus paper: Cerebellum and social cognition. Cerebellum 2020, 19, 833–868. [Google Scholar] [CrossRef] [PubMed]
- Marvel, C. From Motor Systems to Working Memory: The Origins of Stone Tools, Language, Culture, and Rise of Homo. In The New Revolution in Psychology and the Neurosciences; Springer: Cham, Switzerland, 2022; pp. 93–103. [Google Scholar]
- Ernst, D.; Becker, S.; Horstmann, G. Novelty competes with saliency for attention. Vis. Res. 2020, 168, 42–52. [Google Scholar] [CrossRef]
- Bolkan, S.; Griffin, D.J.; Goodboy, A.K. Humor in the classroom: The effects of integrated humor on student learning. Commun. Educ. 2018, 67, 144–164. [Google Scholar] [CrossRef]
- Watson, K.K.; Matthews, B.J.; Allman, J.M. Brain activation during sight gags and language-dependent humor. Cereb. Cortex 2007, 17, 314–324. [Google Scholar] [CrossRef]
- Schomaker, J.; Meeter, M. Short-and long-lasting consequences of novelty, deviance and surprise on brain and cognition. Neurosci. Biobehav. Rev. 2015, 55, 268–279. [Google Scholar] [CrossRef]
- McDermott, K.B.; Roediger, H.L. Memory (Encoding, Storage, Retrieval); General Psychology FA2018; Noba Project: Milwaukie, OR, USA; DEF Publishers: Champaign, IL, USA, 2018; pp. 117–153. [Google Scholar]
- Faraji, M.; Preuschoff, K.; Gerstner, W. Balancing new against old information: The role of puzzlement surprise in learning. Neural Comput. 2018, 30, 34–83. [Google Scholar] [CrossRef]
- Albrecht, J.R.; Karabenick, S.A. Relevance for learning and motivation in education. J. Exp. Educ. 2018, 86, 1–10. [Google Scholar] [CrossRef]
- Panksepp, J. Affective Neuroscience: The Foundations of Human and Animal Emotions; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Tyng, C.M.; Amin, H.U.; Saad, M.N.M.; Malik, A.S. The Influences of Emotion on Learning and Memory. Front. Psychol. 2017, 8, 1454. [Google Scholar] [CrossRef]
- Illeris, K. A comprehensive understanding of human learning. In Contemporary Theories of Learning; Routledge: London, UK, 27 March 2018; pp. 1–14. [Google Scholar]
- Matthews, G.; Wells, A. Attention and Emotion: A Clinical Perspective; Psychology Press: London, UK, 2016. [Google Scholar]
- Pessoa, L. Précis on the cognitive-emotional brain. Behav. Brain Sci. 2015, 38, E71. [Google Scholar] [CrossRef] [PubMed]
- Johnston, E.; Olson, L. The Feeling Brain: The Biology and Psychology of Emotions; WW Norton & Company: New York, NY, USA, 11 May 2015. [Google Scholar]
- Rolls, E.T. The cingulate cortex and limbic systems for emotion, action, and memory. Brain Struct. Funct. 2019, 224, 3001–3018. [Google Scholar] [CrossRef] [PubMed]
- Umbreit, J.; Lane, K.L.; Dejud, C. Improving classroom behavior by modifying task difficulty: Effects of increasing the difficulty of too-easy tasks. J. Posit. Behav. Interv. 2004, 6, 13–20. [Google Scholar] [CrossRef]
- Lynch, R.; Patten, J.V.; Hennessy, J. The impact of task difficulty and performance scores on student engagement and progression. Educ. Res. 2013, 55, 291–303. [Google Scholar] [CrossRef]
- Francis, E.W. Grade level and task difficulty in learning by discovery and verbal reception methods. J. Educ. Psychol. 1975, 67, 146. [Google Scholar] [CrossRef]
- Tavakoli, P. Investigating task difficulty: Learners’ and teachers’ perceptions. Int. J. Appl. Linguist. 2009, 19, 1–25. [Google Scholar] [CrossRef]
- Collins, R. Skills for the 21st Century: Teaching higher-order thinking. Curric. Lead. Ship J. 2014, 12, 10. [Google Scholar]
- Yen, T.S.; Halili, S.H. Effective teaching of higher order thinking (HOT) in education. Online J. Distance Educ. e-Learn. 2015, 3, 41–47. [Google Scholar]
- Larsen, B.; Luna, B. Adolescence as a neurobiological critical period for the development of higher-order cognition. Neurosci. Biobehav. Rev. 2018, 94, 179–195. [Google Scholar] [CrossRef]
- Fuhrmann, D.; Knoll, L.J.; Blakemore, S.J. Adolescence as a sensitive period of brain development. Trends Cogn. Sci. 2015, 19, 558–566. [Google Scholar] [CrossRef]
- Zull, J.E. The Art of Changing the Brain: Enriching the Practice of Teaching by Exploring the Biology of Learning; Stylus Publishing, LLC: Sterling, VA, USA, 28 May 2020. [Google Scholar]
- Oberauer, K. Is rehearsal an effective maintenance strategy for working memory? Trends Inmcognitive Sci. 2019, 23, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Himmer, L.; Schönauer, M.; Heib, D.P.; Schabus, M.; Gais, S. Rehearsal initiates systems memory consolidation, sleep makes it last. Sci. Adv. 2019, 5, eaav1695. [Google Scholar] [CrossRef] [PubMed]
- Wulf, G. Attentional focus and motor learning: A review of 15 years. Int. Rev. Sport Exerc. Psychol. 2013, 6, 77–104. [Google Scholar] [CrossRef]
- Ward, G. Rehearsal Processes. In Oxford Handbook of Human Memory; Oxford University Press: Oxford, UK, 13 July 2022. [Google Scholar]
- Florea, N.M.; Hurjui, E. Critical thinking in elementary school children. Procedia-Soc. Behav. Sci. 2015, 180, 565–572. [Google Scholar] [CrossRef]
- Stan, M.M. The role of learning style in the relationship between cognitive ability and aca- demic performance in primary schooling. In Studies and Current Trends in Science of Education; Editura Lumen, Asociatia Lumen: Iași, Romania, 20 August 2017; pp. 370–378. [Google Scholar]
- Meyer, H.C.; Bucci, D.J. Neural and behavioral mechanisms of proactive and reactive inhibition. Learn. Mem. 2016, 23, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, J.; Saiki, A.; Soma, S.; Yamanaka, K.; Nonomura, S.; Ríos, A.; Kawabata, M.; Kimura, M.; Sakai, Y.; Isomura, Y. Area-specific modulation of functional cortical activity during block-based and trial-based proactive inhibition. Neuroscience 2018, 388, 297–316. [Google Scholar] [CrossRef]
- Solange, D.; Christensen, A.P.; Kenett, Y.N.; Beaty, R.E. Education shapes the structure of semantic memory and impacts creative thinking. NPJ Sci. Learn. 2021, 6, 35. [Google Scholar]
- Takashima, A.; Bakker, I.; Van Hell, J.G.; Janzen, G.; McQueen, J.M. Interaction between episodic and semantic memory networks in the acquisition and consolidation of novel spoken words. Brain Lang. 2017, 167, 44–60. [Google Scholar] [CrossRef]
- Fandakova, Y.; Bunge, S.A. What connections can we draw between research on long-term memory and student learning? Mind Brain Educ. 2016, 10, 135–141. [Google Scholar] [CrossRef]
- Blair, C. Executive function and early childhood education. Curr. Opin. Behav. Sci. 2016, 10, 102–107. [Google Scholar] [CrossRef]
- Sala, G.; Gobet, F. Working memory training in typically developing children: A meta-analysis of the available evidence. Dev. Psychol. 2017, 53, 671. [Google Scholar] [CrossRef] [PubMed]
- Hainselin, M.; Picard, L.; Manolli, P.; Vankerkore-Candas, S.; Bourdin, B. Hey teacher, don’t leave them kids alone: Action is better for memory than reading. Front. Psychol. 2017, 8, 325. [Google Scholar] [CrossRef] [PubMed]
- McLaren, B.M.; Bollen, L.; Walker, E.; Harrer, A.; Sewall, J. Cognitive tutoring of collaboration: Developmental and empirical steps towards realization. In Computer Supported Collaborative Learning 2005: The Next 10 Years! Routledge: London, UK, 3 October 2017; pp. 418–422. [Google Scholar]
- Minges, K.E.; Redeker, N.S. Delayed school start times and adolescent sleep: A systematic review of the experimental evidence. Sleep Med. Rev. 2016, 28, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H. Spaced repetition promotes efficient and effective learning: Policy implications for instruction. Policy Insights Behav. Brain Sci. 2016, 3, 12–19. [Google Scholar] [CrossRef]
- Jonassen, D.H.; Hung, W. All problems are not equal: Implications for problem-based learning. Essent. Read. Probl.-Based Learn. 2015, 15, 17–42. [Google Scholar]
- Kristanto, D.; Liu, M.; Liu, X.; Sommer, W.; Zhou, C. Predicting reading ability from brain anatomy and function: From areas to connections. NeuroImage 2020, 218, 116966. [Google Scholar] [CrossRef]
- Anand, J.; Mulotte, L.; Ren, C.R. Does experience imply learning? Strateg. Manag. J. 2016, 37, 1395–1412. [Google Scholar] [CrossRef]
- Donnelly, J.E.; Hillman, C.H.; Castelli, D.; Etnier, J.L.; Lee, S.; Tomporowski, P.; Lambourne, K.; Szabo-Reed, A.N. Physical activity, fitness, cognitive function, and academic achievement in children: A systematic review. Med. Sci. Sport. Exerc. 2016, 48, 1197. [Google Scholar] [CrossRef]
- Ansari, D.; De Smedt, B.; Grabner, R.H. Neuroeducation–a critical overview of an emerging field. Neuroethics 2012, 5, 105–117. [Google Scholar] [CrossRef]
- Jolles, J.; Jolles, D.D. On Neuroeducation: Why and How to Improve Neuroscientific Literacy in Educational Professionals. Front. Psychol. 2021, 12, 752151. [Google Scholar] [CrossRef]
- Leisman, G. Children’s Language Production: How Cognitive Neuroscience and Industrial Engineering Can Inform Public Education Policy and Practice. In Forum on Public Policy; Oxford Round Table: Urbana, IL, USA, 2012; Volume 2012. [Google Scholar]
- Horvath, J.C.; Donoghue, G.M. A bridge too far–revisited: Reframing bruer’s neuroeducation argument for modern science of learning practitioners. Front. Psychol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, D.L. The biological basis of learning: Neuroeducation through simulation. Simul. Gaming 2015, 46, 175–186. [Google Scholar]
- Grant, J.G.; Siegel, L.S.; D’Angiulli, A. From schools to scans: A neuroeducational approach to comorbid math and reading disabilities. Front. Public Health 2020, 8, 469. [Google Scholar] [CrossRef] [PubMed]
- Sack, K. Georgia’s governor seeks musical start for babies. N. Y. Times 1998, 15, 1998. [Google Scholar]
- Rauscher, F.H.; Shaw, G.L.; Ky, K.N. Listening to Mozart enhances spatial-temporal reasoning: Towards a neurophysiological basis. Neurosci. Lett. 1995, 185, 44–47. [Google Scholar] [CrossRef]
- Rauscher, F.H.; Shaw, G.L.; Ky, C.N. Music and spatial task performance. Nature 1993, 365, 611. [Google Scholar] [CrossRef]
- Schlaug, G.; Norton, A.; Overy, K.; Winner, E. Effects of music training on the child's brain and cognitive development. Ann. N. Y. Acad. Sci. 2005, 1060, 219–230. [Google Scholar] [CrossRef]
- Steele, K.M.; Bass, K.E.; Crook, M.D. The mystery of the Mozart effect: Failure to replicate. Psychol. Sci. 1999, 10, 366–369. [Google Scholar] [CrossRef]
- McCutcheon, L.E. Another failure to generalize the Mozart effect. Psychol. Rep. 2000, 87, 325–330. [Google Scholar] [CrossRef]
- Pietschnig, J.; Voracek, M.; Formann, A.K. Mozart effect–Shmozart effect: A meta-analysis. Intelligence 2010, 38, 314–323. [Google Scholar] [CrossRef]
- Legislative Analysts Office, State of California. Education Mandates: Overhauling a Broken System. 2 February 2010. Available online: https://lao.ca.gov/reports/2010/edu/educ_mandates/ed_mandates_020210.aspx (accessed on 11 September 2022).
- Alferink, L.A.; Farmer-Dougan, V. Brain-(not) based education: Dangers of misunderstanding and misapplication of neuroscience research. Exceptionality 2010, 18, 42–52. [Google Scholar] [CrossRef]
- Bruner, J.S. “The Process of Education” Revisited. Phi Delta Kappan 1971, 53, 18–21. [Google Scholar]
- Stigler, J.W.; Lee, S.Y.; Stevenson, H.W. Mathematics classrooms in Japan, Taiwan, and the United States. Child Dev. 1987, 58, 1272–1285. [Google Scholar] [CrossRef]
- Spiegel, A. Struggle for Smarts? How Eastern and Western Cultures Tackle Learning. National Public Radio. 12 November 2012. Available online: https://www.npr.org/sections/health-shots/2012/11/12/164793058/struggle-for-smarts-how-eastern-and-westerncultures-tackle-learning (accessed on 11 September 2022).
- Ten Houten, W.D. Cognitive Styles and the Social Order. Final Report, Part II. OEO Study B00-5135; University of California: Los Angeles, CA, USA, 1971. [Google Scholar]
- Weingartner, C. Educational Research: The Romance of Quantification. Cetera 2017, 74, 381–402. [Google Scholar]
- De Boer, A.L.; Steyn, T.; Du Toit, P.H. A whole brain approach to teaching and learning in higher education. S. Afr. J. High. Educ. 2001, 15, 185–193. [Google Scholar] [CrossRef][Green Version]
- Sperry, R.W. Left-brain, right-brain. Saturday Rev. 1975, 2, 30–32. [Google Scholar]
- Aunio, P.; Räsänen, P. Core numerical skills for learning mathematics in children aged five to eight years–a working model for educators. Eur. Early Child. Educ. Res. J. 2016, 24, 684–704. [Google Scholar] [CrossRef]
- Zhang, X.; Räsänen, P.; Koponen, T.; Aunola, K.; Lerkkanen, M.K.; Nurmi, J.E. Knowing, applying, and reasoning about arithmetic: Roles of domain-general and numerical skills in multiple domains of arithmetic learning. Dev. Psychol. 2017, 53, 2304. [Google Scholar] [CrossRef]
- Olatoye, R.A.; Aderogba, A.A. Performance of senior secondary school science students in aptitude test: The role of student verbal and numerical abilities. J. Emerg. Trends Educ. Res. Policy Stud. 2011, 2, 431–435. [Google Scholar]
- Wiliams, D. Principled Curriculum Design; SSAT (The Schools Network) Limited: London, UK, 2013. [Google Scholar]
- Asttle, J. Do Schools Really Kill Creativity. Available online: https://www.thersa.org/blog/2018/04/do-schools-kill-creativity (accessed on 12 September 2022).
- Woodward, A. Beyond Textbooks in Elementary Social Studies. Soc. Educ. 1986, 50, 50–53. [Google Scholar]
- Chang, C.C.; Silalahi, S.M. A review and content analysis of mathematics textbooks in educational research. Probl. Educ. 21st Century 2017, 75, 235. [Google Scholar] [CrossRef]
- Fitzgerald, J.; Elmore, J.; Relyea, J.E.; Stenner, A.J. Domain-specific academic vocabulary network development in elementary grades core disciplinary textbooks. J. Educ. Psychol. 2020, 112, 855. [Google Scholar] [CrossRef]
- Michaud, A. The Mechanics of Conceptual Thinking. Creat. Educ. 2019, 10, 353. [Google Scholar] [CrossRef]
- Lohbeck, A.; Möller, J. Social and dimensional comparison effects on math and reading self- concepts of elementary school children. Learn. Individ. Differ. 2017, 54, 73–81. [Google Scholar] [CrossRef]
- Gazzaniga, M.S. Review of the split brain. J. Neurol. 1975, 209, 75–79. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Meaning, Emotions and Learning | However, When Individuals Feel Helpless and Anxious… | However, There Must Be Some “Challenge” |
|---|---|---|
|
|
|
| Learning | Memory | Retention |
|---|---|---|
| How our brain acquires new information and skills | How and where our brain stores learned information and skills | How long-term memory preserves learning so that it can locate, identify, and retrieve it accurately |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leisman, G. On the Application of Developmental Cognitive Neuroscience in Educational Environments. Brain Sci. 2022, 12, 1501. https://doi.org/10.3390/brainsci12111501
Leisman G. On the Application of Developmental Cognitive Neuroscience in Educational Environments. Brain Sciences. 2022; 12(11):1501. https://doi.org/10.3390/brainsci12111501
Chicago/Turabian StyleLeisman, Gerry. 2022. "On the Application of Developmental Cognitive Neuroscience in Educational Environments" Brain Sciences 12, no. 11: 1501. https://doi.org/10.3390/brainsci12111501
APA StyleLeisman, G. (2022). On the Application of Developmental Cognitive Neuroscience in Educational Environments. Brain Sciences, 12(11), 1501. https://doi.org/10.3390/brainsci12111501