1. Introduction
One of the core missions for perceptual systems is to provide the brain with reliable information about the environment to enable efficient interaction with nearby objects via goal-directed actions. Sensory estimates need to be accurate and precise in many dimensions as objects (and events) are internally represented within a multidimensional space encompassing many properties, including spatial position, time of occurrence, and numerosity. As these variables often correlate with each other (for example, it takes more time to walk a longer distance or to pick up more cherries from a table), it has been proposed that there exists within the parietal lobe of the human brain a shared magnitude system to process information about space, time, and quantity via a single mechanism. This idea, known as “A theory of Magnitude-ATOM”, has been extensively tested by measuring the extent to which the processing of information in one of the ATOM dimension distorts estimates in another [
1]. For example, several studies have reported that large visual stimuli are perceived to last longer than smaller ones presented for the same physical duration [
2]. Duration estimates are also significantly affected by stimuli numerosity: numerous stimuli are perceived to last longer than less numerous ones [
3].
A shared mechanism processing quantitative information in multiple dimensions may be beneficial in providing a unique interface between the perceptual and the motor systems, sub-serving the transfer of sensory information between them. In line with this idea, voluntary movements performed during the presentation of visual stimuli can affect perceived duration. For example, at the time of saccades, duration is considerably compressed, by about 50% [
4]. Similar (but weaker) compression also occurs when task-unrelated hand movements, either a series of discrete temporal patterns or continuous actions, are made during the presentation of visual stimuli [
5,
6]. Action does not always cause compression: a saccadic movement to a clock’s second hand can induce the illusion of temporal expansion, sometimes referred to as “saccadic chronostasis” [
7].
Recently, a new motor-induced temporal illusion has been reported, where repetitive hand-tapping can distort the perceived duration of stimuli subsequently presented around the tapping area [
8]. The perceived duration of visual stimuli is significantly compressed (by about 30%) after fast tapping and expanded after slow tapping. Motor adaptation of this type does not only distort perceived time but also perceived numerosity [
9] and spatial distance [
10]. However, motor adaptation does not affect the perception of stimulus speed, suggesting that the motor system might selectively interact with the processing of parietal information, without affecting that of earlier levels of visual processing.
A recent study from Lambourne et al. investigated the role of physical exercise on a temporal comparison task and found that visual stimuli presented during sustained aerobic exercise executed with the lower limbs (cycling) induced an expansion of perceived duration of about 15% [
11]. However, the study did not address whether these effects were selective only for temporal estimates or whether they generalize to other quantitative dimensions, as would be suggested by the ATOM Theory. A second question is whether sensory distortions are induced only during the execution of actions or whether the distortion persists after completion of motor activity, indicating a relatively long-term recalibration of the sensory system after physical activity. If the effect induced by self-motion were to disappear immediately after the end of the physical activity, it would indicate that the distortion of perceived duration is related to the movement itself and not to the other physiological variables that are changed during the activity and take time to revert to baseline levels. While both hand-tapping and cycling (or running) can be categorized as “self-motion”, they are very different in terms of effectors (upper or lower limbs) and frequency, and while hand-tapping could more closely resemble the action of counting items scattered on a surface, cycling is a non-goal-directed action. If a different influence of action on numerosity depending on the type of motion was to be found, it would suggest a selective interaction between action and numerosity, with types of motion more closely related to counting being more prone to distortion of numerosity.
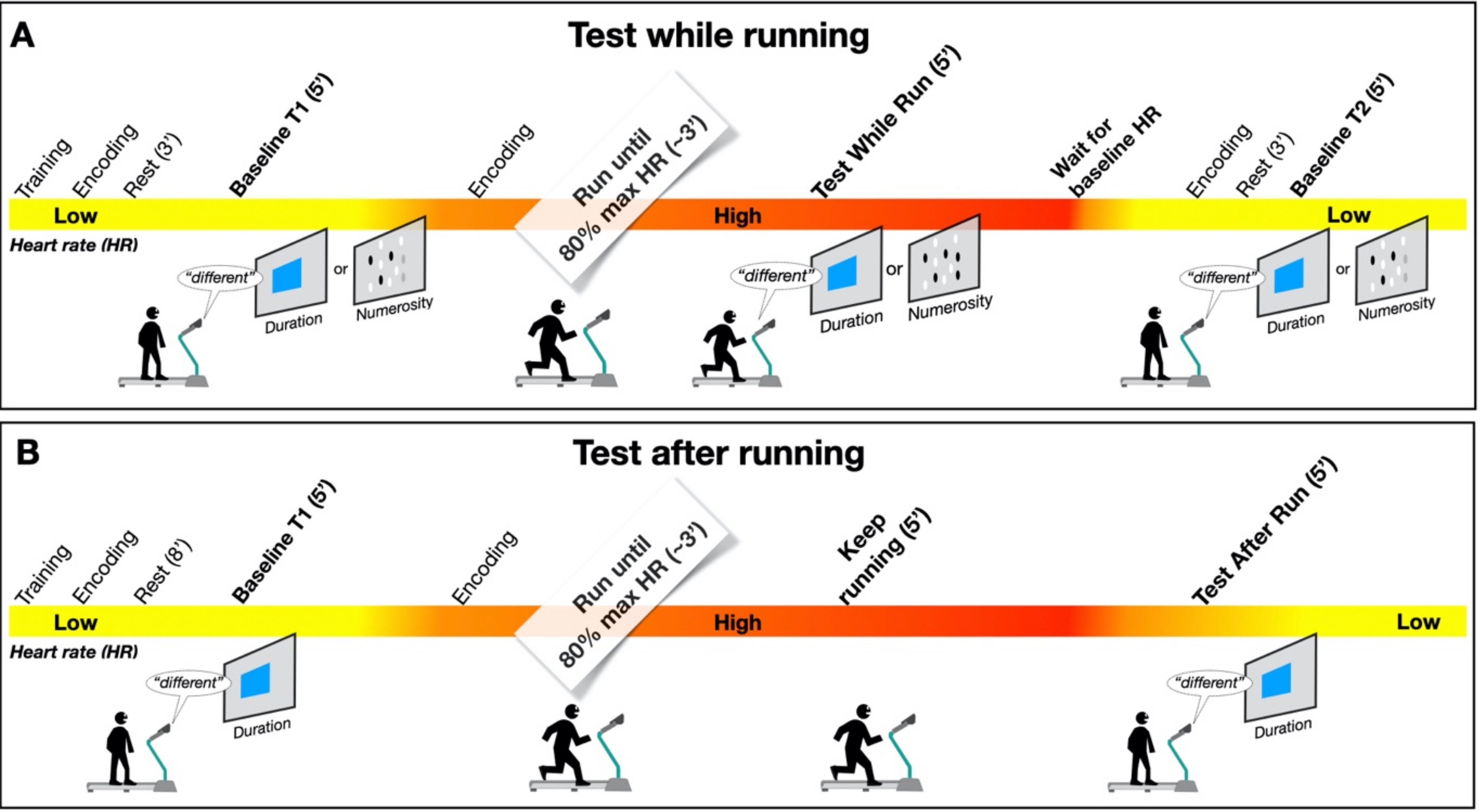
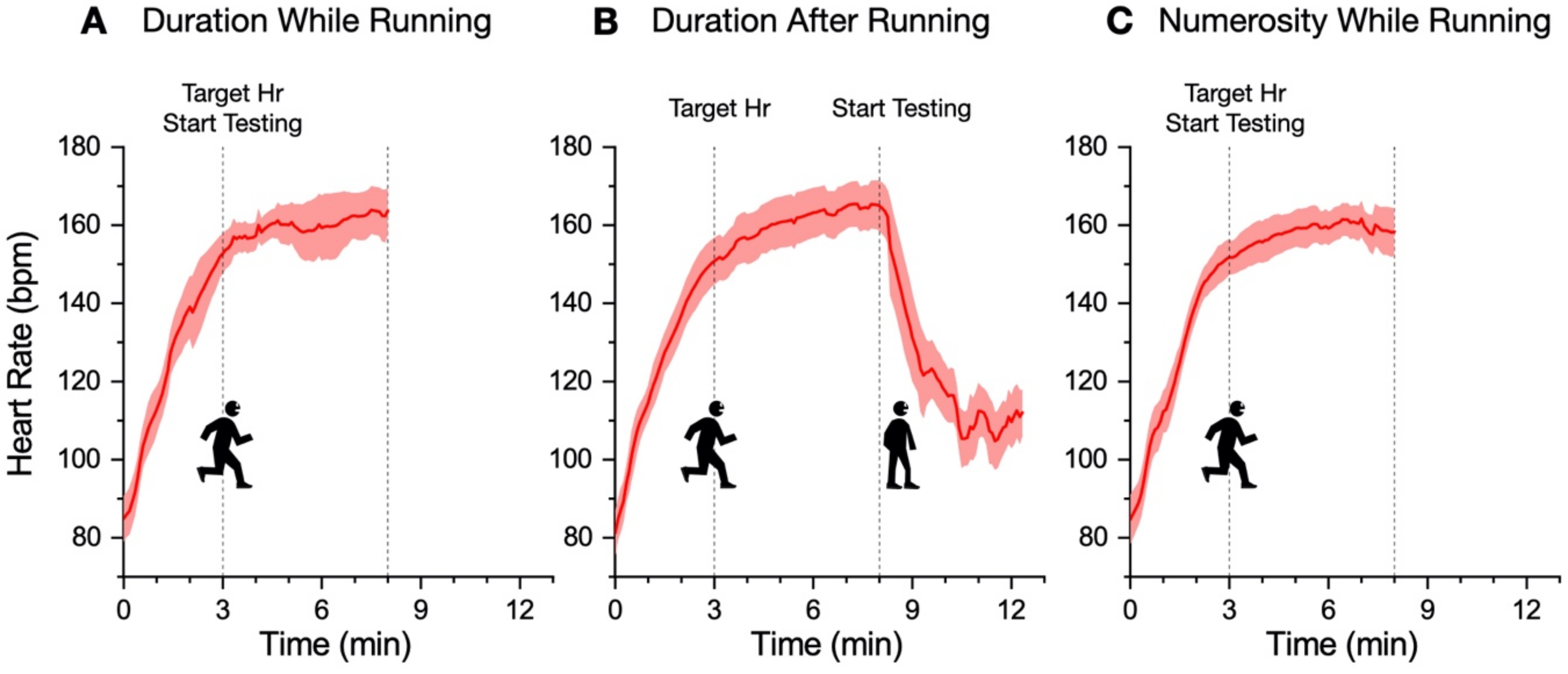
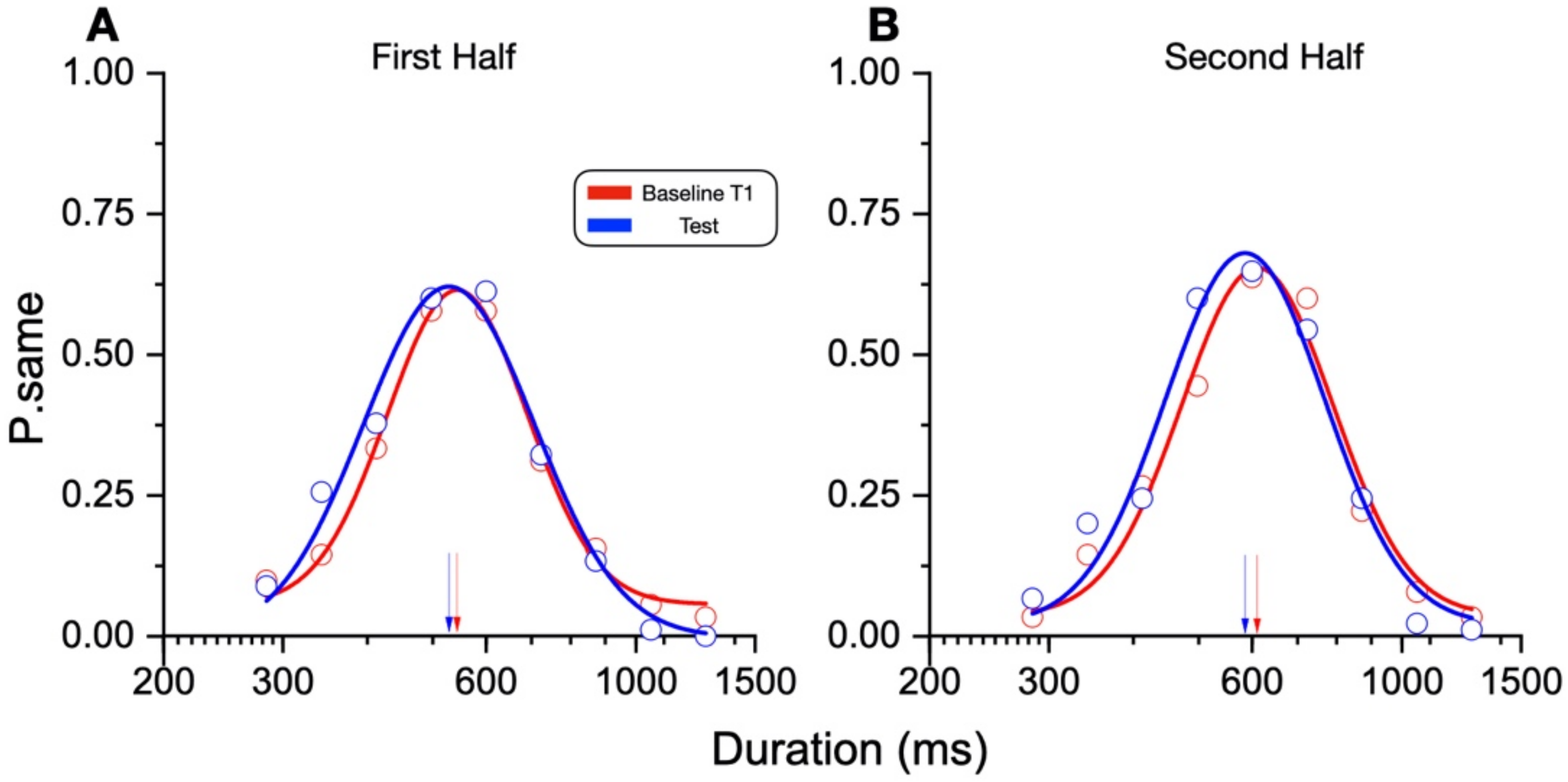
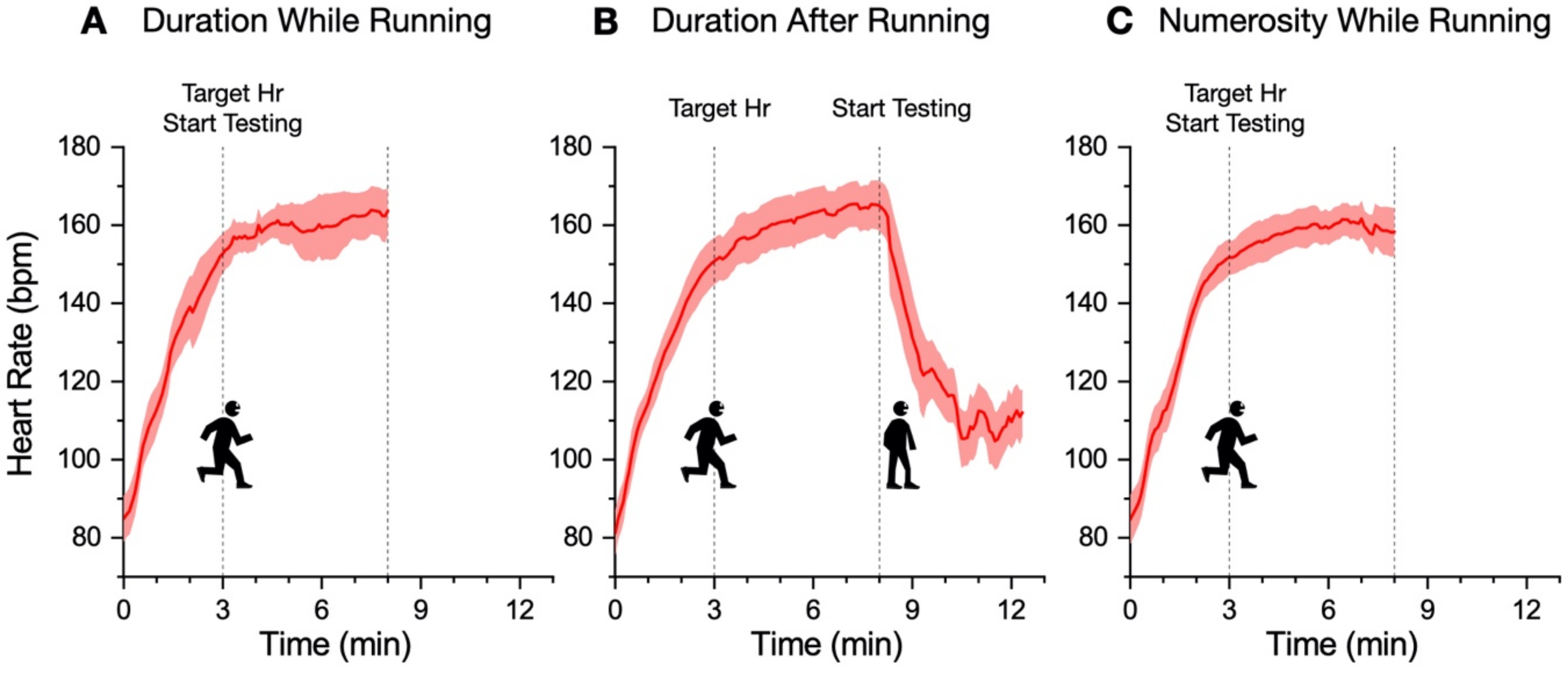
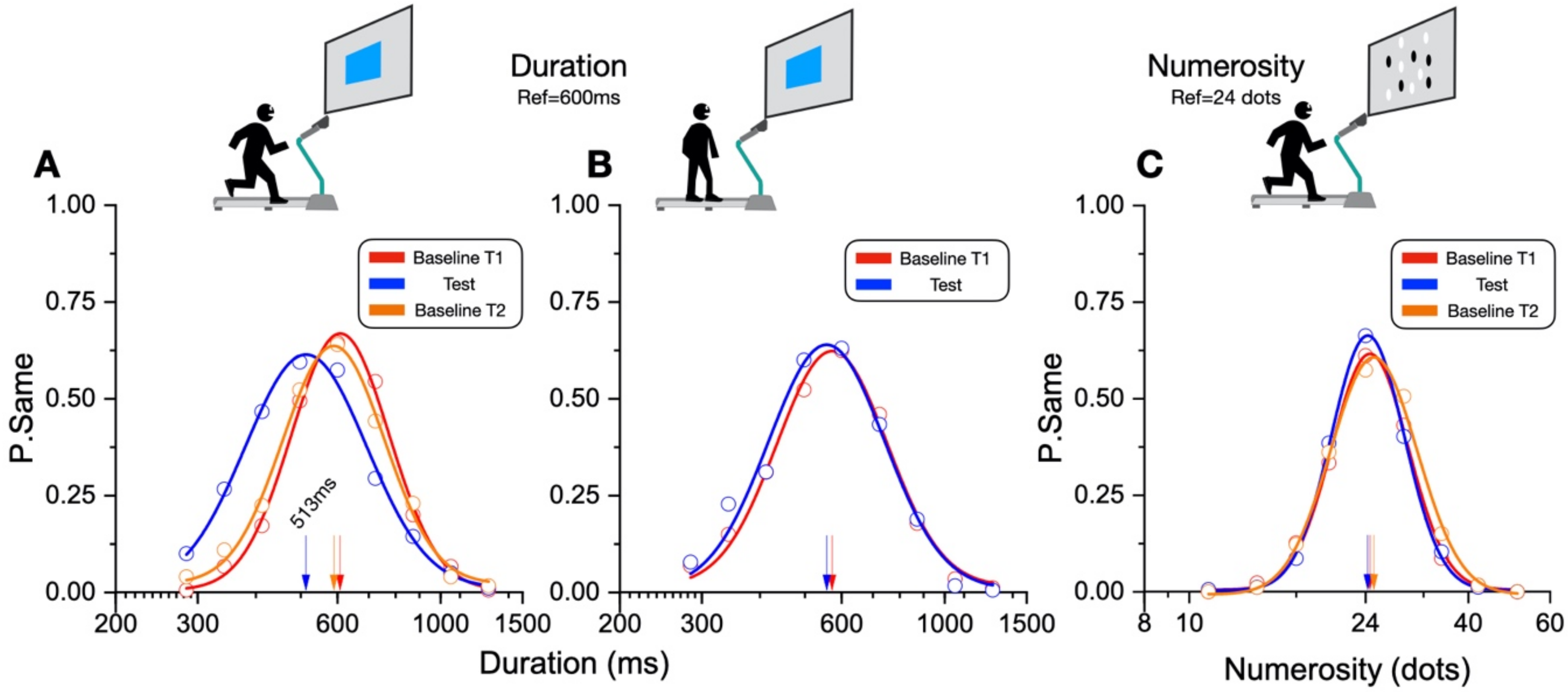
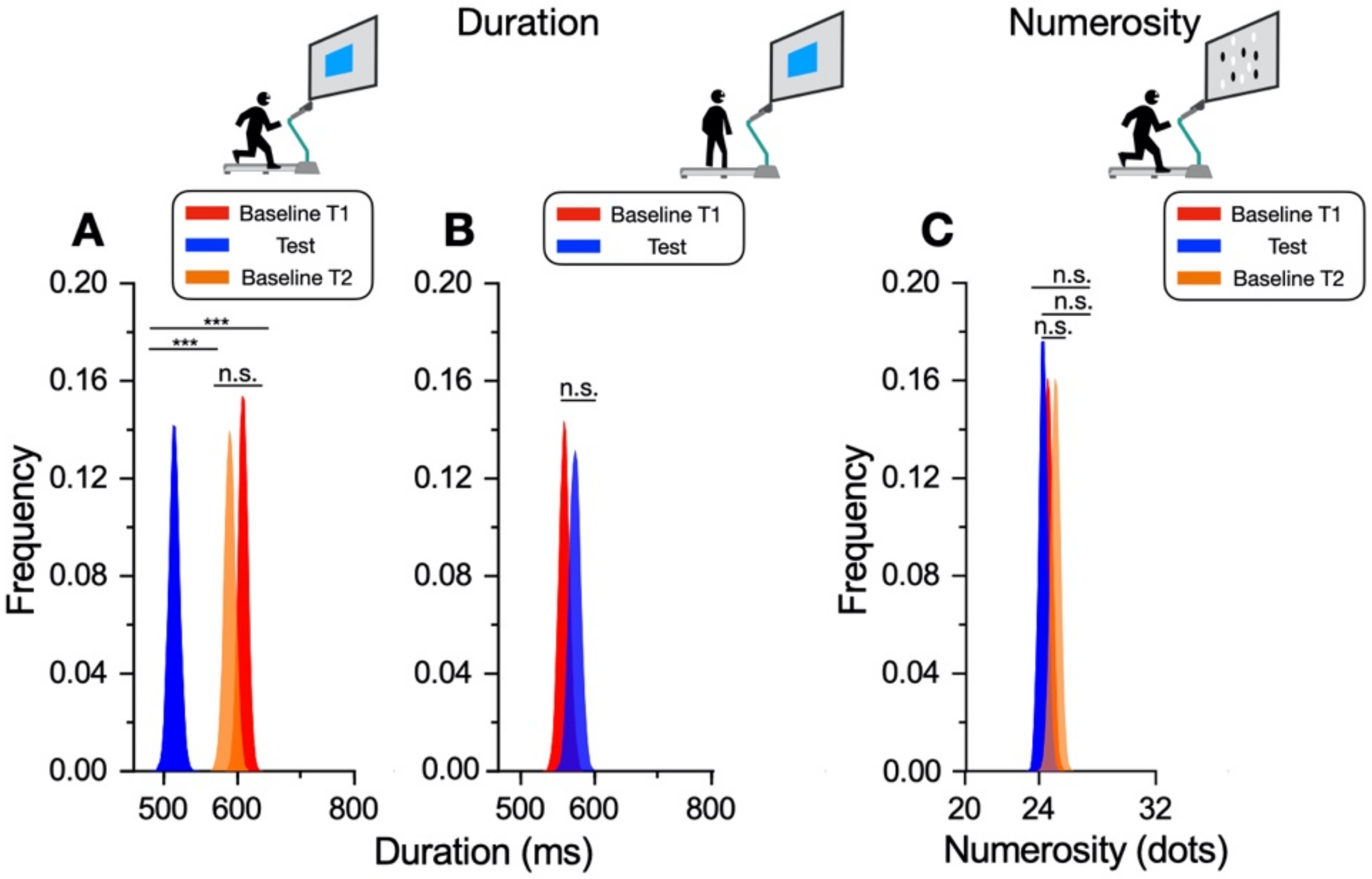
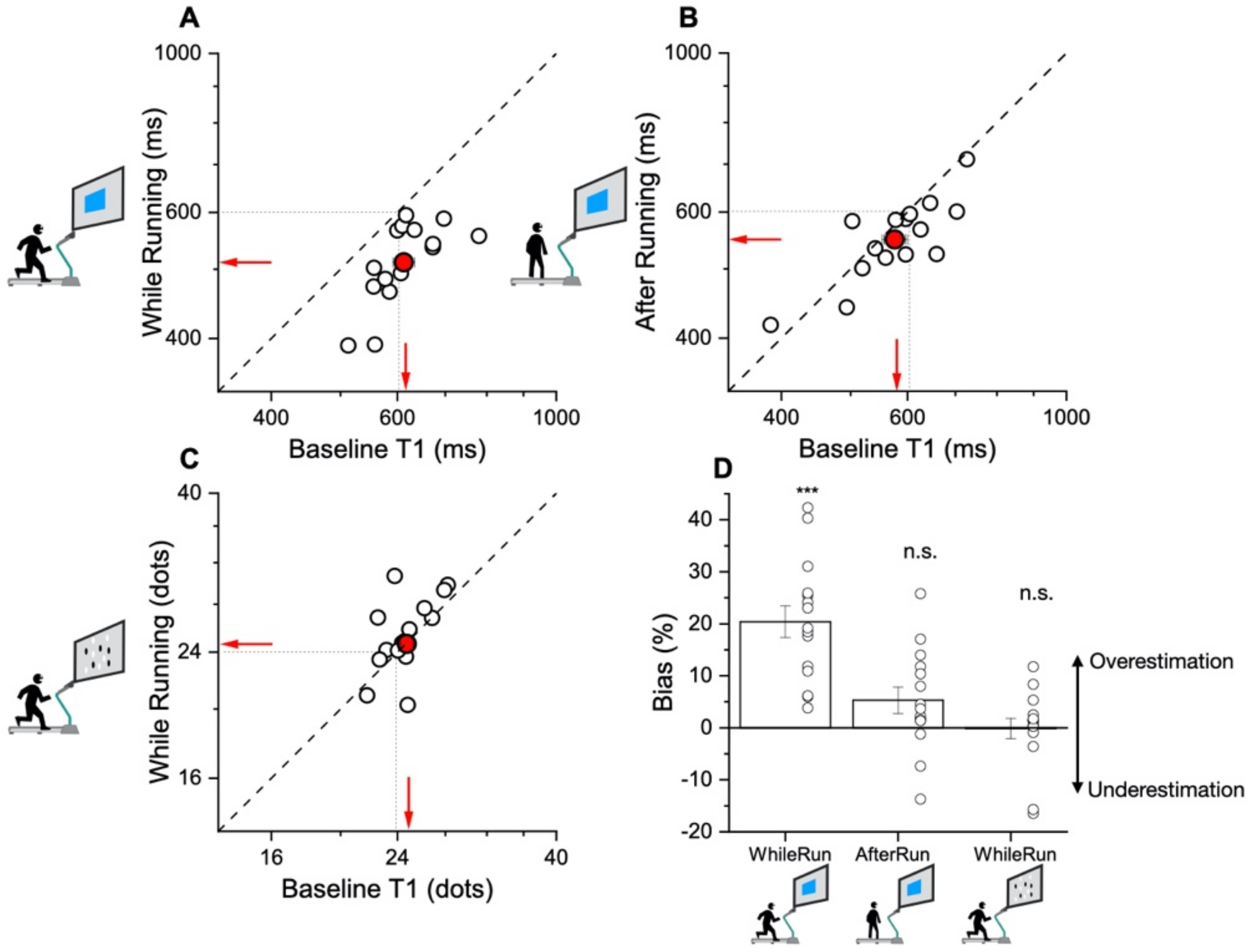
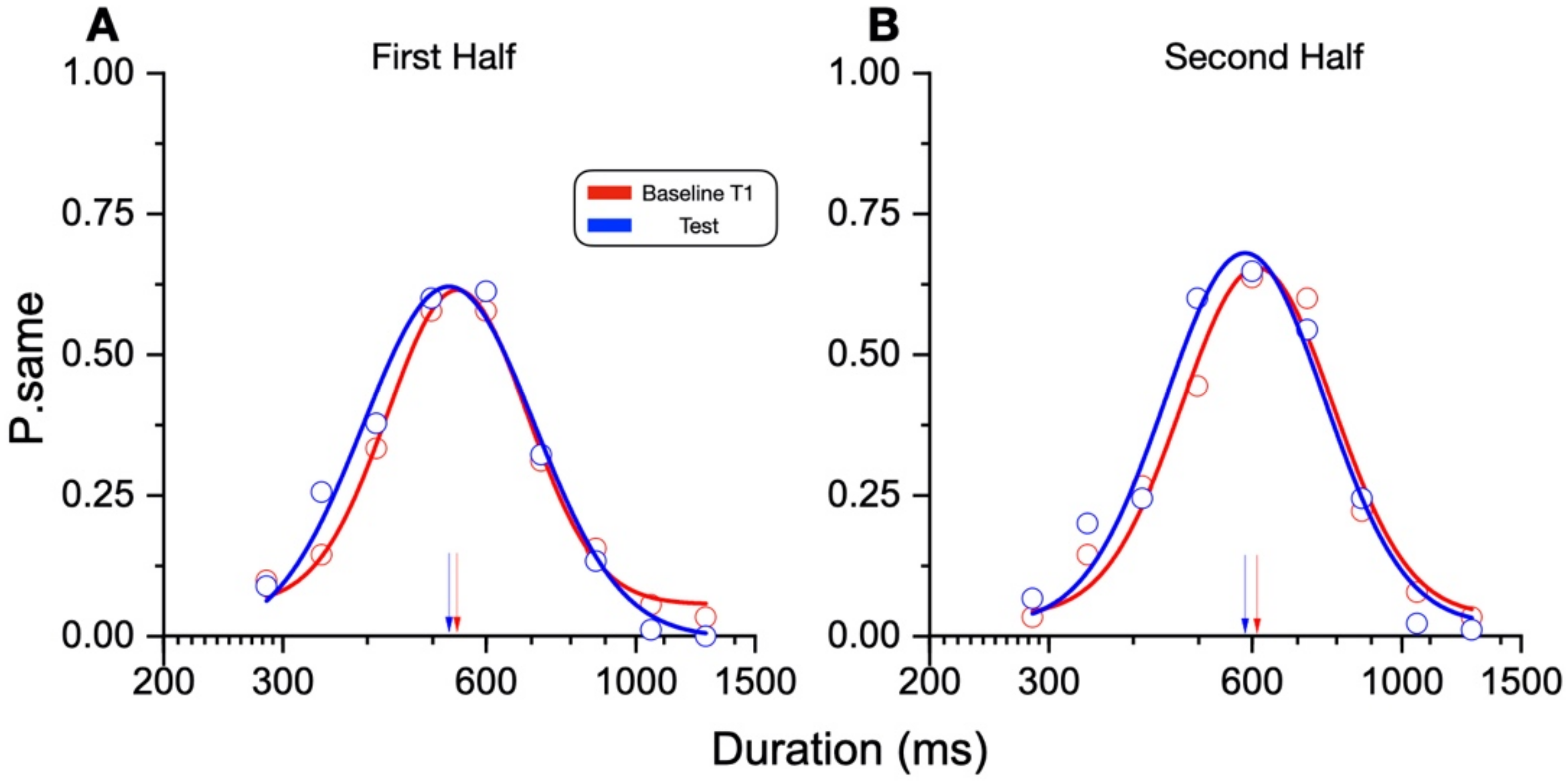
In our new paradigm, the participants made a temporal or numerosity comparison (separate sessions) in three different conditions: at rest, during sustained physical exercise (running on a treadmill), or immediately after the exercise. In line with previous reports, perceived duration was expanded during the motor routines; however, estimates of numerosity were almost completely unaffected. Furthermore, time judgements were not distorted for stimuli presented soon after the end of the physical activity, although several physiological variables, such as heart rate, remained altered relative to the baseline, suggesting that distortions of time occur only during the execution of actions, with rapid recalibration after the activity is completed.
4. Discussion
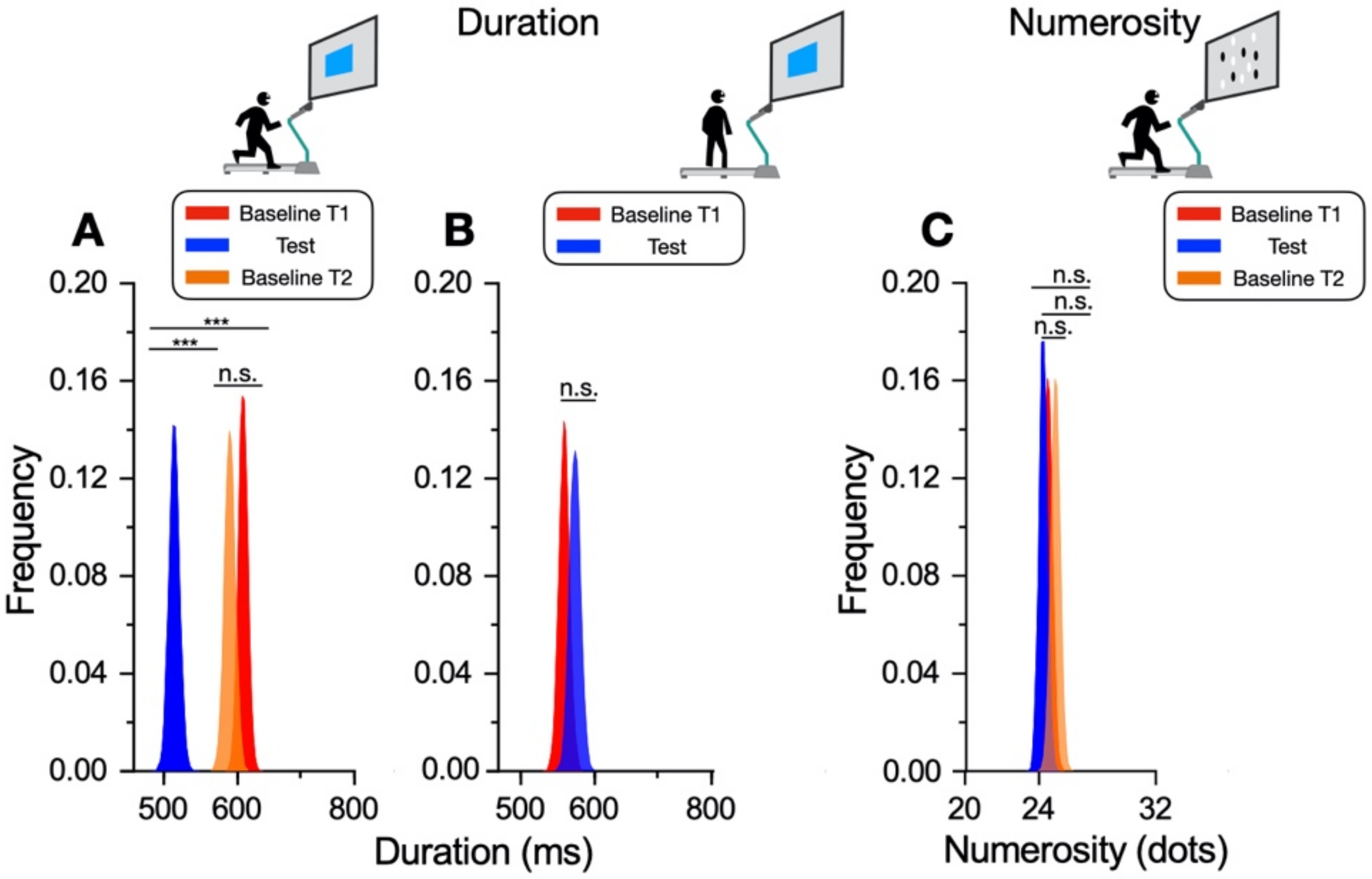
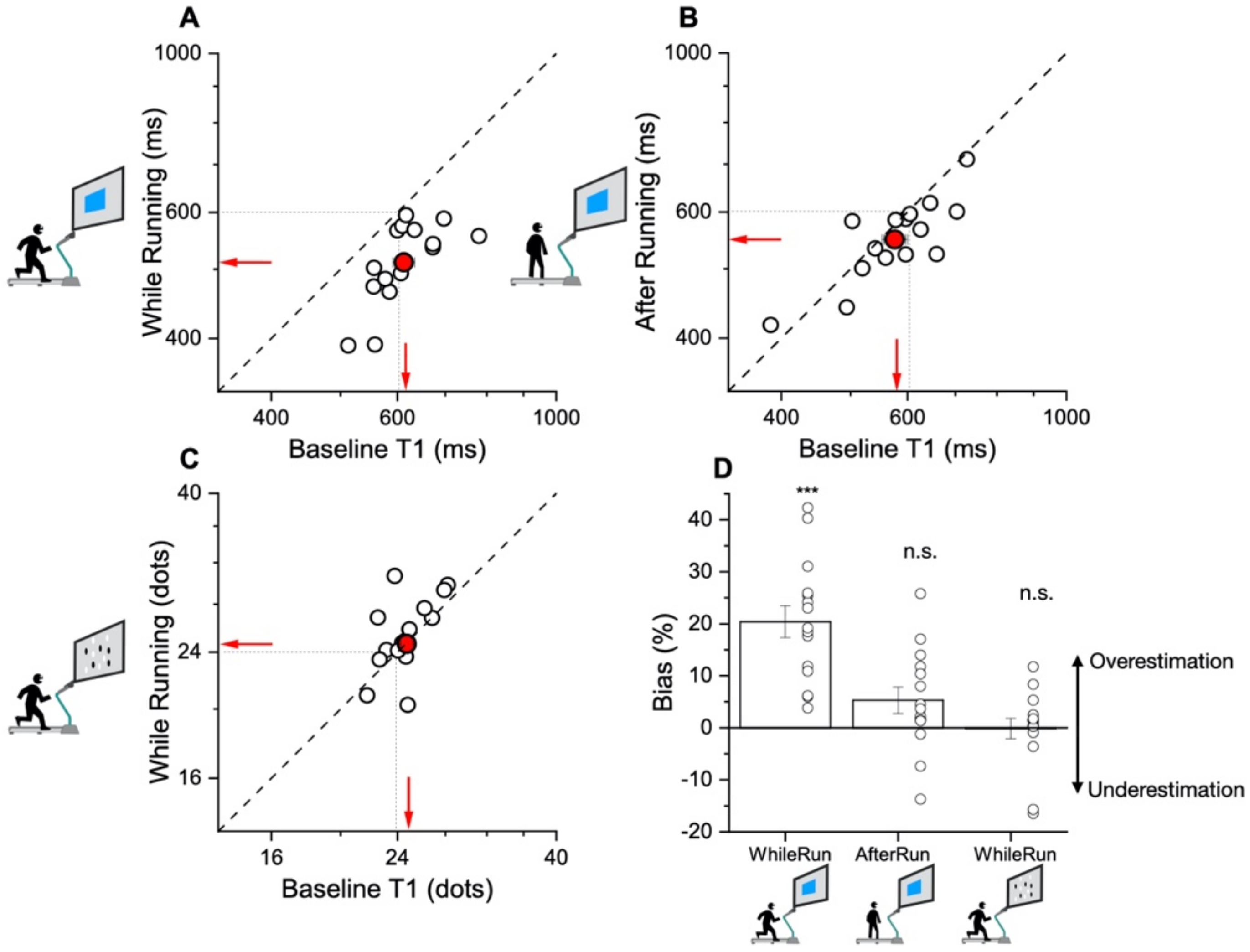
The main result of this study shows that while motor activity can significantly distort the perception of time, it leaves numerosity perception unaffected. The participants compared the duration or numerosity of a test stimulus while running, or having just run, with a standard encoded at rest. The first experiment measuring duration estimation during running supported the previous findings [
11], reporting a systematical overestimation of perceived duration while running. The second experiment, however, showed that the duration measurements made soon after stopping running are veridical, showing that this effect is intrinsically related to the movement itself rather than to other physiological parameters altered by physical activity (such as heart rate). In the final experiment, we showed that the numerosity estimation was unaffected by running.
These findings suggest a magnitude-selective interaction between action and perception that involves only the perceived duration. However, one of the most prominent theories on the perception of magnitude posits that the human brain encodes space, time, and number via a shared mechanism [
1], opening the possibility that the effect of motor activity on the perception of time might also generalize to other magnitudes. Indeed, it has recently been reported that a repetitive motor routine executed with the upper limbs (hand tapping) distorts both perceived duration [
8] and numerosity [
9]. Moreover, motor activity related to eye movements has been reported to significantly distort numerosity [
16,
17].
How can the discrepancy between the present and the previous studies be reconciled? One possibility regards the spatial congruency between the position where the visual stimuli were displayed and the area where the motor activity was directed. The perceived numerosity during saccadic eye movements was distorted only for stimuli displayed between the saccadic starting and ending point, embracing a distance of roughly 20° [
16]. Similarly, hand tapping distorted the perceived numerosity of stimuli presented around the tapping area, with effects that rapidly faded off with the increase in the spatial offset relative to the tapping location and completely vanished for distances higher than 15° [
8].
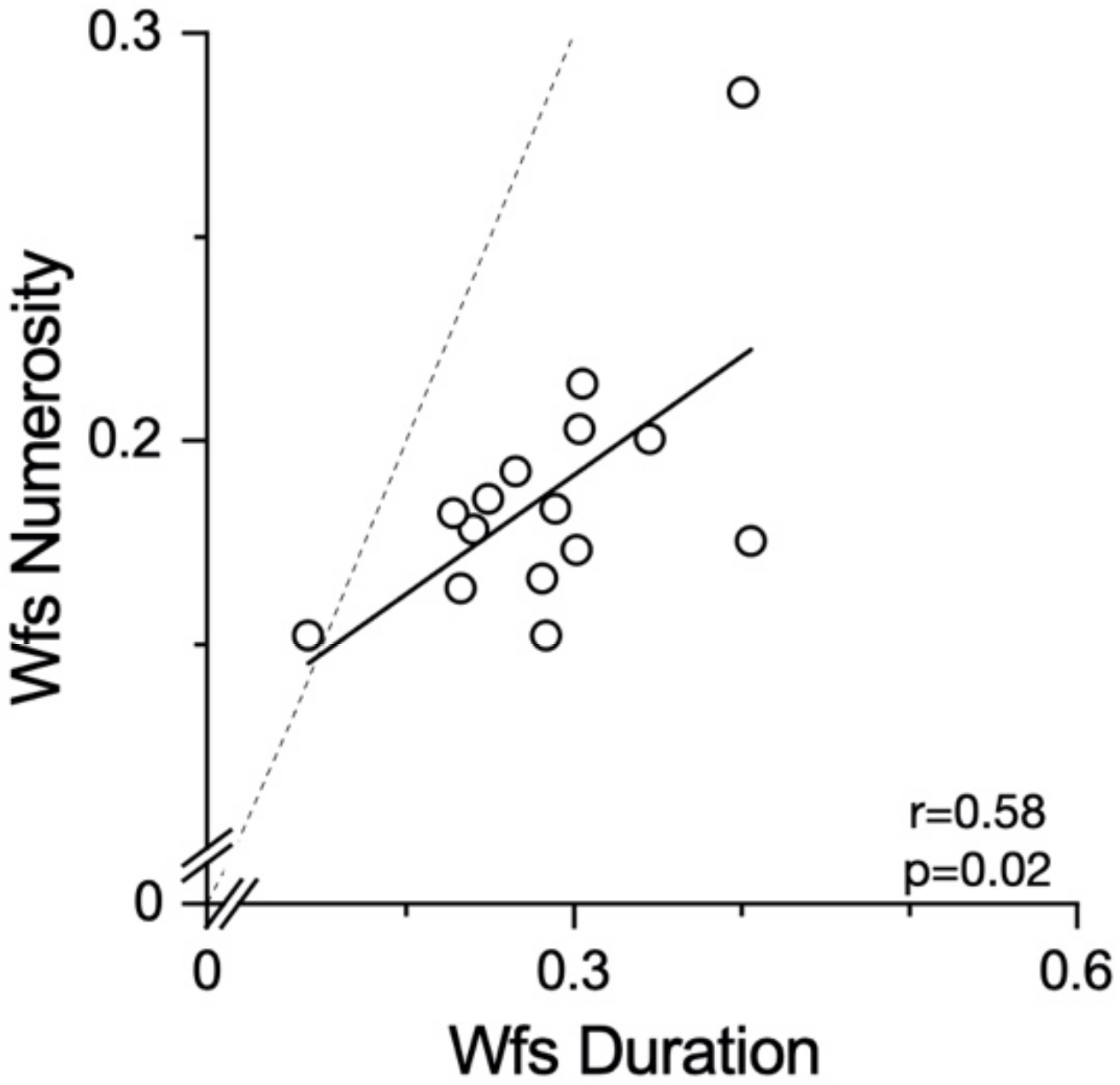
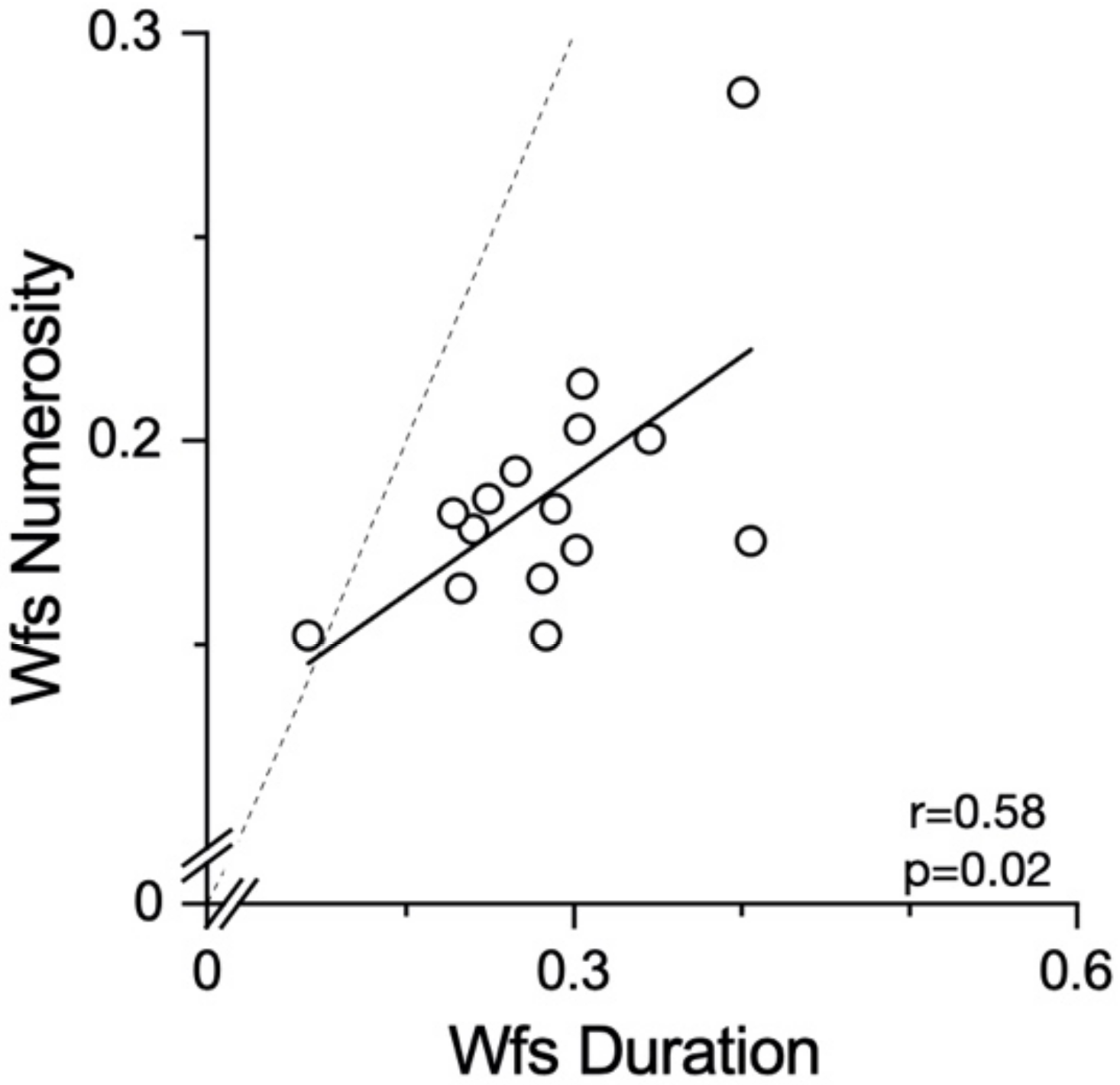
Another possible explanation for the null effect found for numerosity may be the different sensory precision for the two magnitudes. The Weber fractions for the numerosity perception were lower than those for the duration perception. It is possible that the noisier system for duration perception is more prone to distortion by contextual variables, such as running. However, we found no difference in the Weber fractions between the duration judgements during and after running, making it unlikely that this is the general explanation for all lack of effects. A last methodological difference worth discussion is the presentation modality of the stimuli. As the dots in the numerosity task were all presented simultaneously, this might have required a lower involvement of the working memory compared to the duration stimuli. However, as other interactions between self-motion and numerosity were reported for both the sequential and the simultaneous [
9] numerosities, this difference alone is unlikely to have cancelled out an effect of running on numerosity perception.
Interestingly, in many of the previous studies the participants underestimated perceived duration as a consequence of action. In contrast, the participants in the present study showed a tendency to overestimate the durations of visual stimuli presented during the running phase relative to those with the same physical duration perceived at rest. A possible reconciliation of these discrepancies may be the difference in methodologies. In all the previous experiments, the participants were required to compare two intervals presented one after the other or to immediately reproduce a temporal interval that had been just observed. In the paradigm of the present study, the intervals presented during the running phase were compared with the reference encoded before the onset of physical activity. This resulted in a delay between the encoding (at rest) and the test phase (during running) of at least 3 min, a much longer time for which sensory information had to be stored in the short-term working memory.
A second methodological difference regards the duration and the intensity of the motor activity. While many studies on eye and hand movements use transient motor-routines in the sub-second or supra second regime, here participants were engaged in a strenuous physical activity lasting several minutes. Indeed, a previous report in which participants were required to estimate the duration of visual stimuli during a sustained cycling routine lasting several minutes revealed that the duration estimates during physical exercise were robustly lengthened [
11]. Typically, these time-dilation phenomena are accounted for by alterations in the rate of the internal pacemaker: physical activity could accelerate the clock rate, and this, in turn, would induce a perceived dilation of stimulus duration. To reconcile all these results, we might speculate that transient motor activities (such as saccadic eye movements or hand movements) momentarily slow down the rate of the pacemaker, resulting in a compression of perceived time. On the other hand, sustained physical activity may induce an acceleration of the internal clock, yielding the opposite phenomenon of time dilation.
An alternative explanation might be that time processing in different conditions involves several, independent temporal mechanisms in the brain. This idea of multiple clocks in the brain has been demonstrated within the visual domain. A grating drifting at high speed presented in a given location of the visual field strongly compresses the perceived duration of the stimuli subsequently displayed around that area, without affecting those displayed in other locations [
18,
19]. This finding supports the idea of multiple time mechanisms, each responsible for time processing in a well-defined portion of the visual field, violating the well-held belief of a centralized, unique internal clock. Despite previous reports supporting the idea that motor routines can affect time perception, it remains an open question whether such an interaction is prompted by the execution of the movements themselves or by the alteration of other physiological variables that are perturbated during action.
Past reports have proposed that heart rate might be directly related to the internal clock rate; so, an acceleration of heartrate would also speed up the internal clock, leading to an overestimation of perceived time [
20]. To address this possibility, we measured whether time perception was affected not only during the running phase but also immediately after the end of the physical activity. The results indicate that no distortion occurred after completion of the motor routine, even though heartrate had not returned to the baseline level. Even when we took into consideration only the trials immediately after the physical activity, no significant perceived time dilation was observed. This result clearly questions the causal role of heartrate variations in distortions in perceived time and is in line with a previous study reporting significant time dilations when arousal increased but heart rate remained constant, or even decreased [
21]. Can the present results be accounted for by variations of the arousal level induced by motor activity? The lack of temporal distortions when duration estimates were made at the end of the physical activity also questions the arousal hypothesis, given that arousal levels have been reported to be still significantly altered at the end of a 10 min running exercise (see [
22]) in a similar activity to the present study.
Taken together, the results of the present study reinforce previous studies showing that time perception is affected by running, but a similar running regime does not affect numerosity perception. However, a comparison between these findings with those in the literature revealed that this relationship is modulated by movement parameters, such as movement speed, type of effector, spatial proximity between stimuli, and movement location, as well as the time of stimuli presentation relative to the phases of the motor routine [
23]. Future studies should directly investigate each of these issues to provide a full comprehension of the mechanisms and the nature of the interaction between the motor and the sensory systems.
The effect of long-term physical training on time perception should also be investigated. For instance, there have been anecdotical reports from tennis and baseball players that time slows down just before hitting the ball [
24]. This suggests that the target of goal-directed actions can benefit from specialized processing of their temporal features, which might be aimed at maximizing motor performance and be induced by the extreme long-term training characterizing elite athletes. However, while the benefit of time dilation for goal-directed action is evident, the same does not hold for target-free rhythmic actions, such as running. This may suggest that while the observed behavioral effect is the same, it might be caused by different mechanisms. During sustained physical exercise, the fatigue accumulated by the participant may cause an overestimation of perceived time, as has also been reported to occur during sustained rowing exercise [
25]. In the light of this, future studies could investigate the precision and accuracy of temporal perception in elite athletes, such as those engaged in long-distance races (marathons) or hurdling, combining the running routine with a goal-directed, repetitive, transient action aimed at jumping over the obstacles.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}