
Neuroplasticity within and between Functional Brain Networks in Mental Training Based on Long-Term Meditation

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
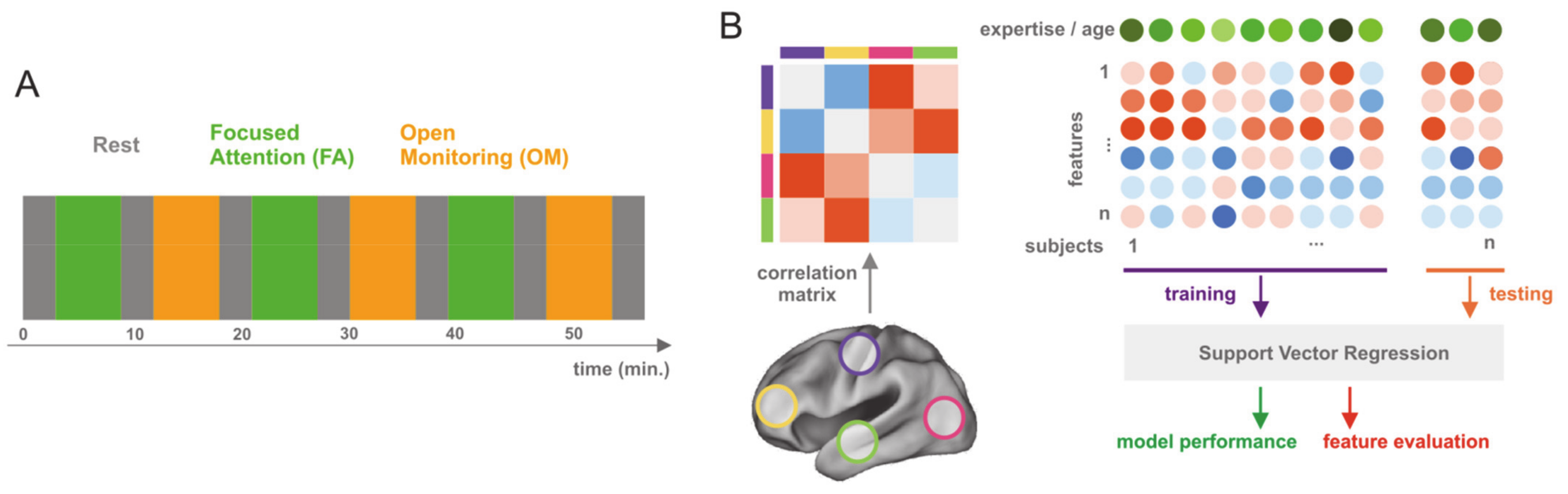
2.2. Experimental Design
2.3. Data Acquisition and Preprocessing
2.4. Functional Connectivity Analysis
2.5. Registration and ROI Definition
2.6. Multivariate Pattern Regression
2.7. Relevant Feature Analysis
2.8. Control Analysis for Confound Effects
2.9. Control Analyses in a Novice Group
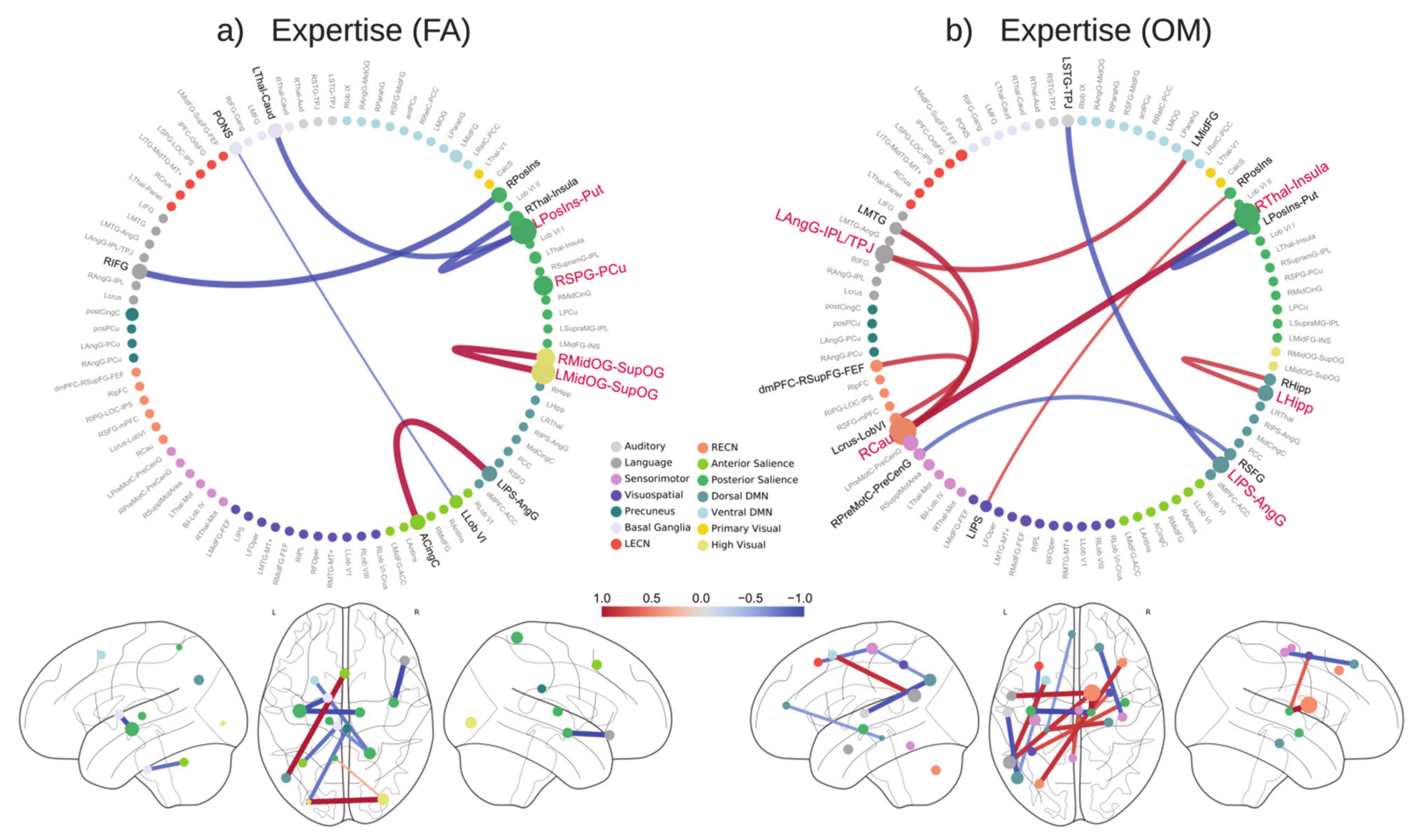
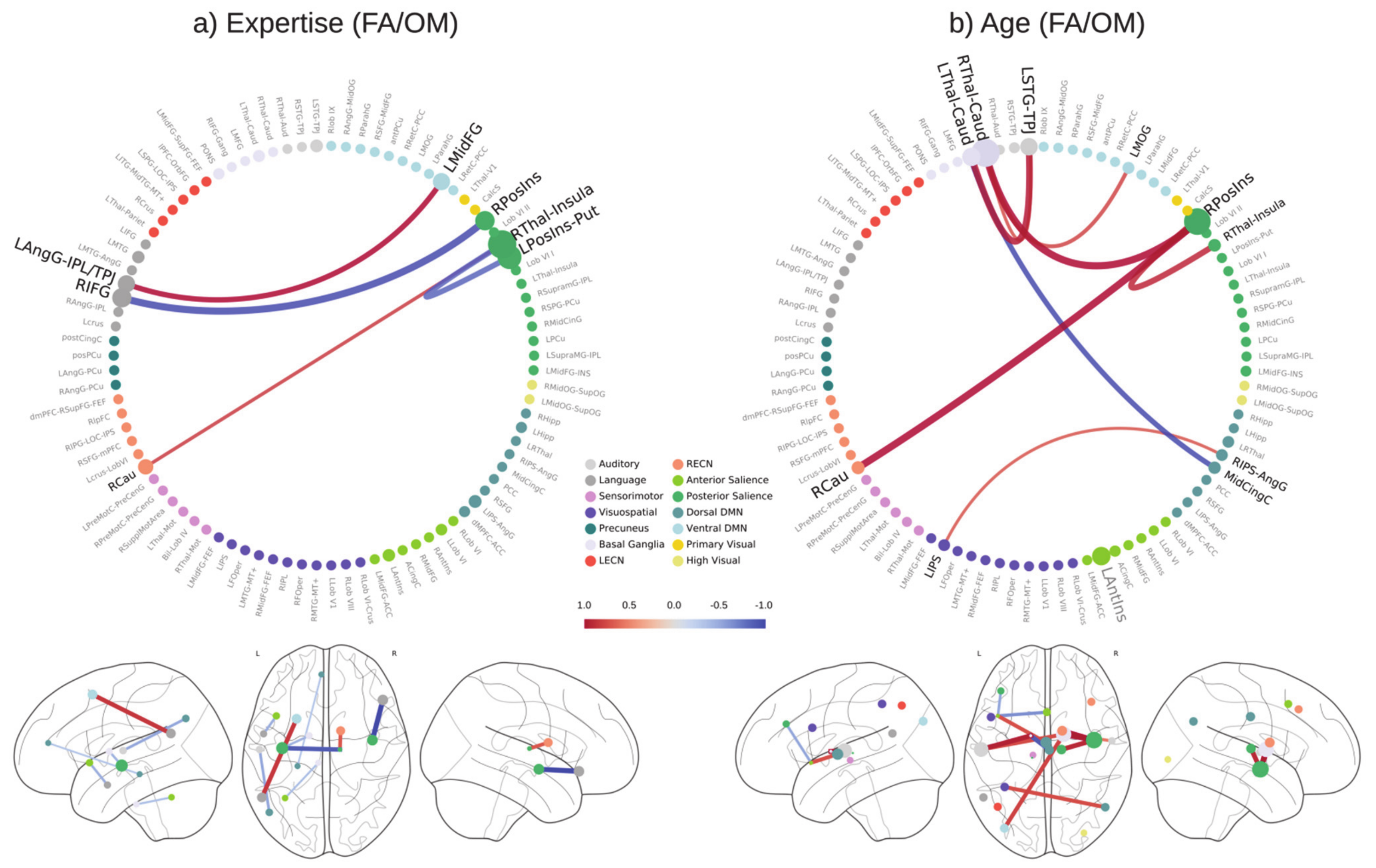
3. Results
3.1. Prediction of Years of Meditation Expertise
3.2. Prediction of Age
3.3. Control Analyses for the Confounding Effect
3.4. Control Analyses in the Control Group
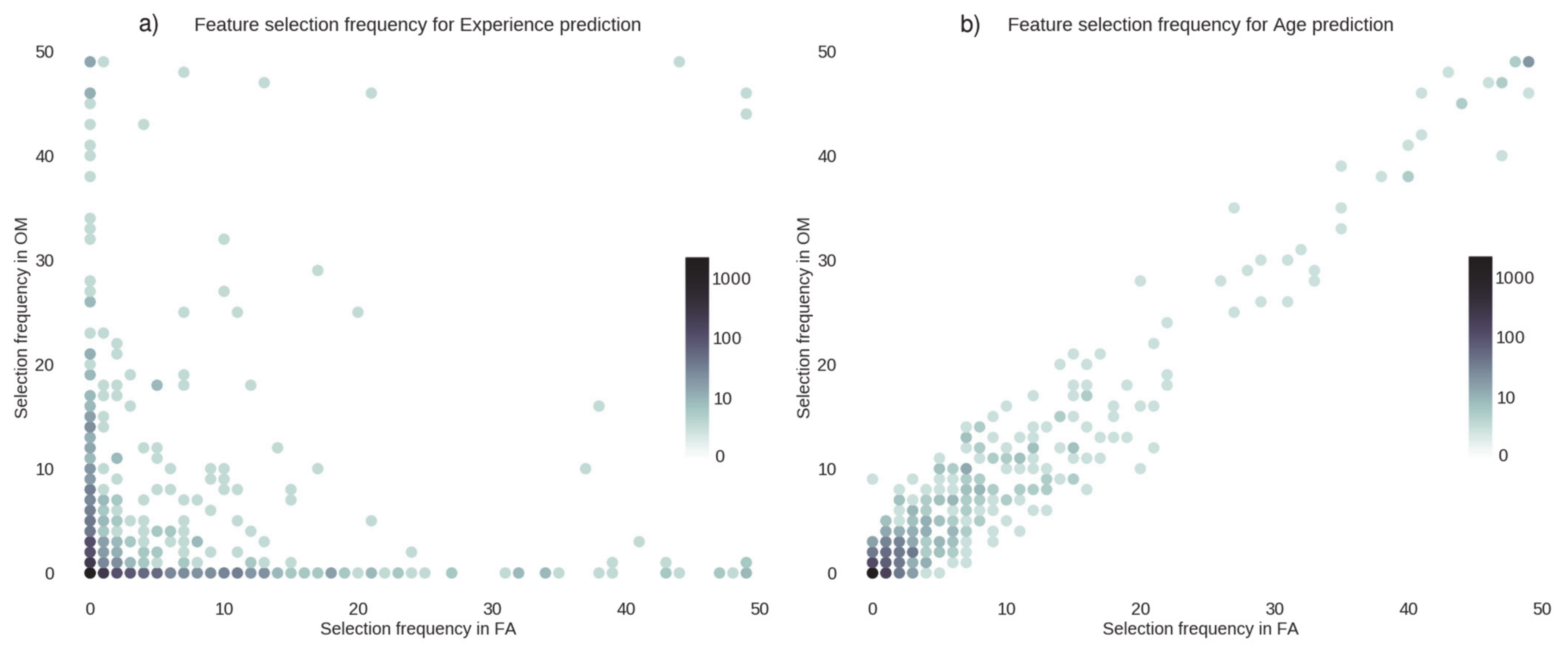
3.5. Feature Selection Frequency
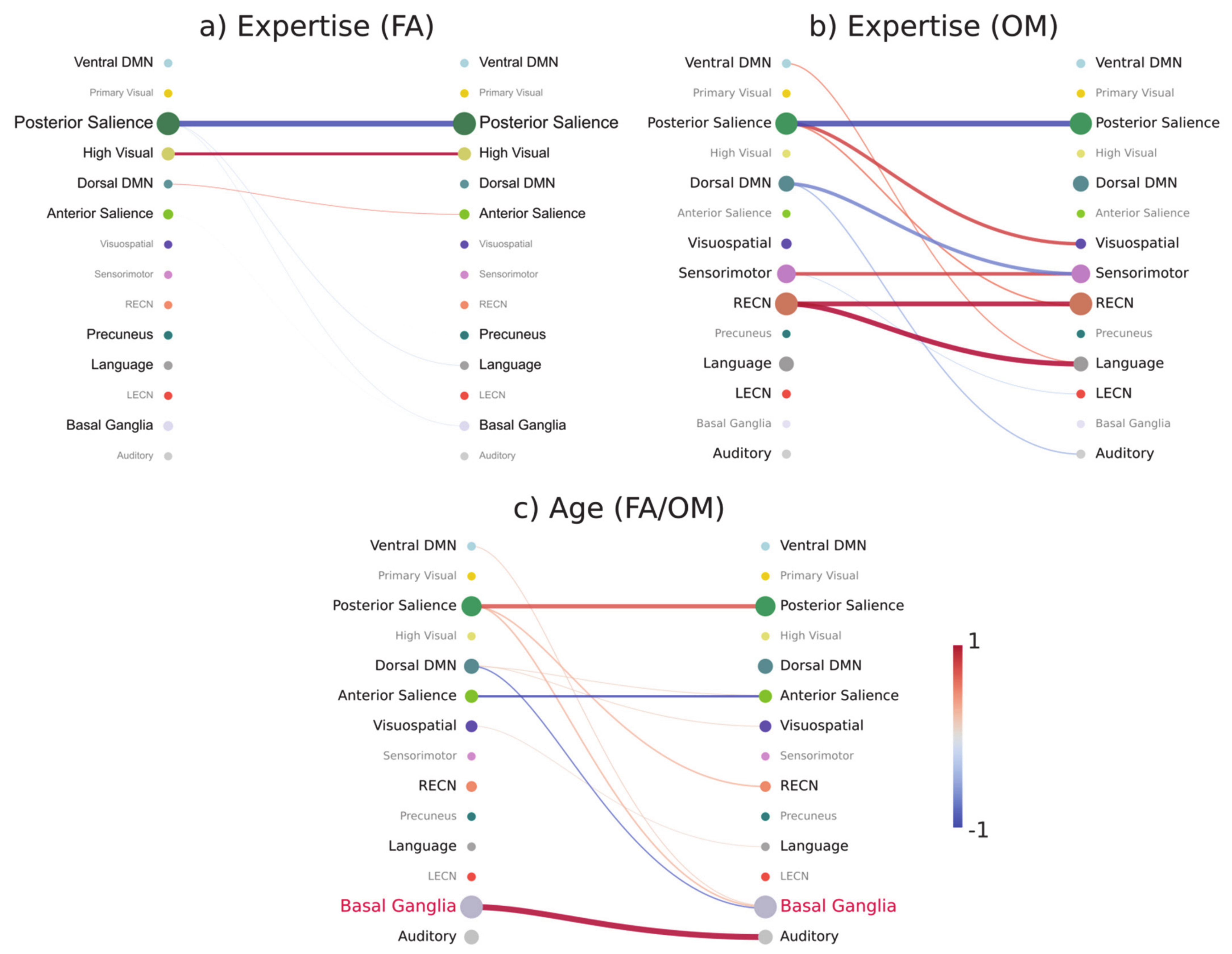
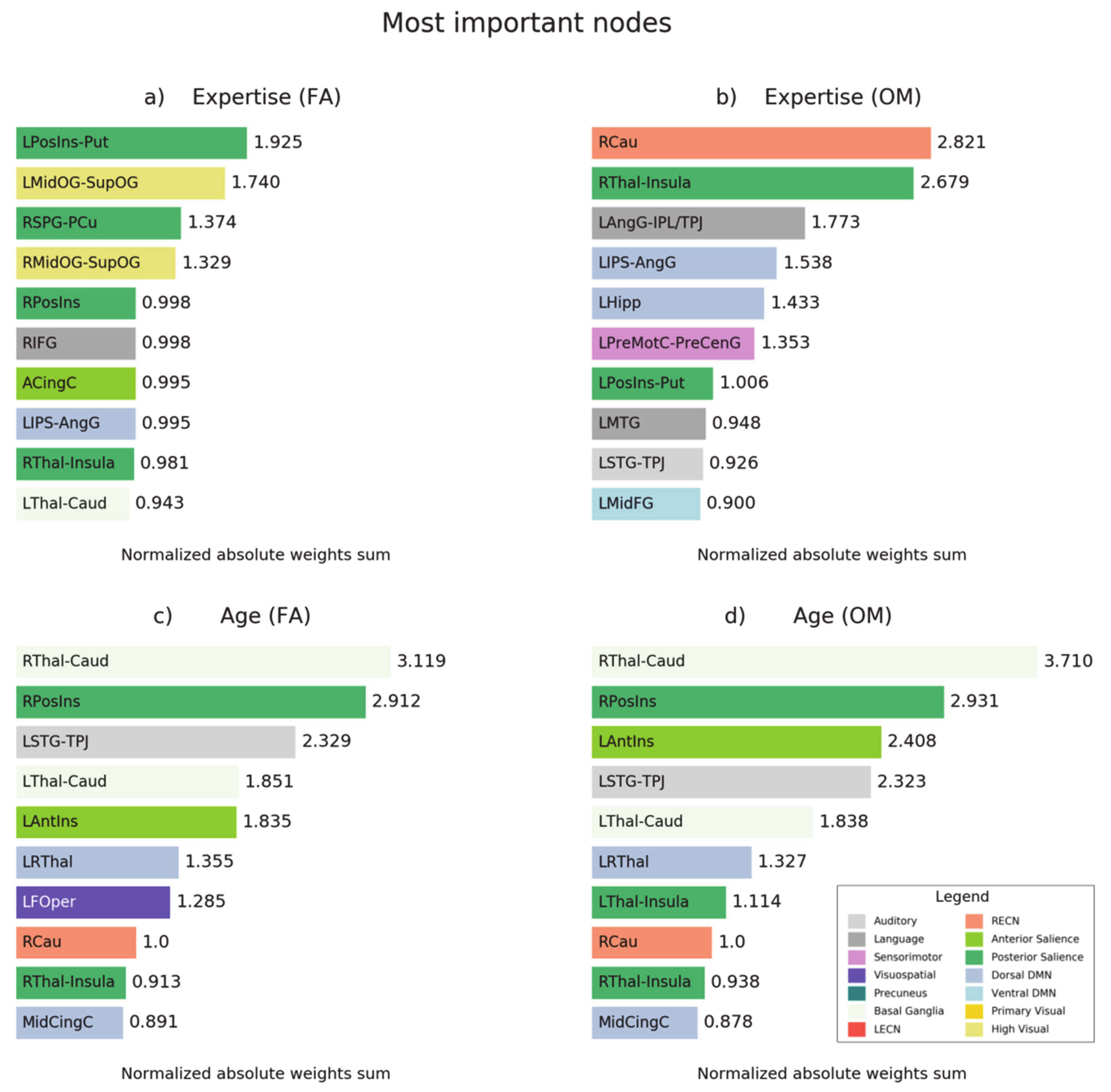
3.6. Prediction-Relevant Feature Weight Analysis
4. Discussion
4.1. Prediction of Meditation Expertise in FA
4.2. Prediction of Meditation Expertise in OM
4.3. Prediction of Age
4.4. Functional Connectivity Patterns at the Brain Network Level
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lutz, A.; Slagter, H.; Dunne, J.D.; Davidson, R.J. Attention regulation and monitoring in meditation. Trends Cogn. Sci. 2008, 12, 163–169. [Google Scholar] [CrossRef]
- Tang, Y.-Y.; Hölzel, B.K.; Posner, M.I. The neuroscience of mindfulness meditation. Nat. Rev. Neurosci. 2015, 16, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Cahn, B.R.; Polich, J. Meditation states and traits: EEG, ERP, and neuroimaging studies. Psychol. Bull. 2006, 132, 180–211. [Google Scholar] [CrossRef]
- Hasenkamp, W.; Wilson-Mendenhall, C.D.; Duncan, E.; Barsalou, L.W. Mind wandering and attention during focused meditation: A fine-grained temporal analysis of fluctuating cognitive states. NeuroImage 2012, 59, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Hölzel, B.K.; Carmody, J.; Vangel, M.; Congleton, C.; Yerramsetti, S.M.; Gard, T.; Lazar, S. Mindfulness practice leads to increases in regional brain gray matter density. Psychiatry Res. Neuroimaging 2011, 191, 36–43. [Google Scholar] [CrossRef]
- Malinowski, P. Neural mechanisms of attentional control in mindfulness meditation. Front. Neurosci. 2013, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Goleman, D.; Davidson, R.J. The Science of Meditation: How to Change Your Brain, Mind and Body, 2nd ed.; . Penguin Life: London, UK, 2018. [Google Scholar]
- Raffone, A.; Marzetti, L.; Del Gratta, C.; Perrucci, M.G.; Romani, G.L.; Pizzella, V. Toward a brain theory of meditation. Prog. Brain Res. 2019, 244, 207–232. [Google Scholar] [CrossRef]
- Lutz, A.; Jha, A.P.; Dunne, J.D.; Saron, C.D. Investigating the phenomenological matrix of mindfulness-related practices from a neurocognitive perspective. Am. Psychol. 2015, 70, 632–658. [Google Scholar] [CrossRef] [PubMed]
- Manna, A.; Raffone, A.; Perrucci, M.G.; Nardo, D.; Ferretti, A.; Tartaro, A.; Londei, A.; Del Gratta, C.; Belardinelli, M.O.; Romani, G.L. Neural correlates of focused attention and cognitive monitoring in meditation. Brain Res. Bull. 2010, 82, 46–56. [Google Scholar] [CrossRef]
- Marzetti, L.; Di Lanzo, C.; Zappasodi, F.; Chella, F.; Raffone, A.; Pizzella, V. Magnetoencephalographic alpha band connectivity reveals differential default mode network interactions during focused attention and open monitoring meditation. Front. Hum. Neurosci. 2014, 8, 832. [Google Scholar] [CrossRef]
- Yordanova, J.; Kolev, V.; Mauro, F.; Nicolardi, V.; Simione, L.; Calabrese, L.; Malinowski, P.; Raffone, A. Common and distinct lateralised patterns of neural coupling during focused attention, open monitoring and loving kindness meditation. Sci. Rep. 2020, 10, 7430. [Google Scholar] [CrossRef]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; Van Essen, D.C.; Raichle, M.E. From The Cover: The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar] [CrossRef]
- Heuvel, M.V.D.; Stam, C.J.; Kahn, R.S.; Pol, H.H. Efficiency of Functional Brain Networks and Intellectual Performance. J. Neurosci. 2009, 29, 7619–7624. [Google Scholar] [CrossRef]
- Bressler, S.L.; Menon, V. Large-scale brain networks in cognition: Emerging methods and principles. Trends Cogn. Sci. 2010, 14, 277–290. [Google Scholar] [CrossRef]
- Raffone, A.; Srinivasan, N. An adaptive workspace hypothesis about the neural correlates of consciousness: Insights from neuroscience and meditation studies. In Progress in Brain Research; Elsevier BV: Amsterdam, The Netherlands, 2009; pp. 161–180. [Google Scholar]
- Zanto, T.P.; Gazzaley, A. Attention and Ageing; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Reuter-Lorenz, P.A.; Festini, S.B.; Jantz, T.K. Executive Functions and Neurocognitive Aging. In Handbook of the Psychology of Aging, 8th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2015; p. 245. [Google Scholar]
- Brefczynski-Lewis, J.A.; Lutz, A.; Schaefer, H.S.; Levinson, D.B.; Davidson, R. Neural correlates of attentional expertise in long-term meditation practitioners. Proc. Natl. Acad. Sci. USA 2007, 104, 11483–11488. [Google Scholar] [CrossRef] [PubMed]
- Tomasino, B.; Fregona, S.; Skrap, M.; Fabbro, F. Meditation-related activations are modulated by the practices needed to obtain it and by the expertise: An ALE meta-analysis study. Front. Hum. Neurosci. 2013, 6, 346. [Google Scholar] [CrossRef] [PubMed]
- Kurth, F.; Cherbuin, N.; Luders, E. Promising Links between Meditation and Reduced (Brain) Aging: An Attempt to Bridge Some Gaps between the Alleged Fountain of Youth and the Youth of the Field. Front. Psychol. 2017, 8, 860. [Google Scholar] [CrossRef]
- Malinowski, P.; Shalamanova, L. Meditation and Cognitive Ageing: The Role of Mindfulness Meditation in Building Cognitive Reserve. J. Cogn. Enhanc. 2017, 1, 96–106. [Google Scholar] [CrossRef]
- Haxby, J.V. Multivariate pattern analysis of fMRI: The early beginnings. NeuroImage 2012, 62, 852–855. [Google Scholar] [CrossRef] [PubMed]
- Haynes, J.-D. A Primer on Pattern-Based Approaches to fMRI: Principles, Pitfalls, and Perspectives. Neuron 2015, 87, 257–270. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, A.J.; Jiang, F.; Abdi, H.; Pénard, N.; Dunlop, J.P.; Parent, M.A. Theoretical, Statistical, and Practical Perspectives on Pattern-based Classification Approaches to the Analysis of Functional Neuroimaging Data. J. Cogn. Neurosci. 2007, 19, 1735–1752. [Google Scholar] [CrossRef]
- Knops, A.; Thirion, B.; Hubbard, E.M.; Michel, V.; Dehaene, S. Recruitment of an Area Involved in Eye Movements During Mental Arithmetic. Science 2009, 324, 1583–1585. [Google Scholar] [CrossRef] [PubMed]
- Tosoni, A.; Guidotti, R.; Del Gratta, C.; Committeri, G.; Sestieri, C. Preferential coding of eye/hand motor actions in the human ventral occipito-temporal cortex. Neuropsychologia 2016, 93, 116–127. [Google Scholar] [CrossRef]
- Guidotti, R.; Tosoni, A.; Perrucci, M.G.; Sestieri, C. Choice-predictive activity in parietal cortex during source memory decisions. NeuroImage 2019, 189, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Guidotti, R.; Tosoni, A.; Sestieri, C. Properties and temporal dynamics of choice- and action-predictive signals during item recognition decisions. Brain Struct. Funct. 2020, 225, 2271–2286. [Google Scholar] [CrossRef] [PubMed]
- Dosenbach, N.U.F.; Nardos, B.; Cohen, A.; Fair, D.A.; Power, J.D.; Church, J.; Nelson, S.M.; Wig, G.S.; Vogel, A.C.; Lessov-Schlaggar, C.N.; et al. Prediction of Individual Brain Maturity Using fMRI. Science 2010, 329, 1358–1361. [Google Scholar] [CrossRef]
- Finn, E.S.; Shen, X.; Scheinost, D.; Rosenberg, M.; Huang, J.; Chun, M.; Papademetris, X.; Constable, R. Functional connectome fingerprinting: Identifying individuals using patterns of brain connectivity. Nat. Neurosci. 2015, 18, 1664–1671. [Google Scholar] [CrossRef]
- Cole, J.H.; Franke, K. Predicting Age Using Neuroimaging: Innovative Brain Ageing Biomarkers. Trends Neurosci. 2017, 40, 681–690. [Google Scholar] [CrossRef]
- Woo, C.-W.; Chang, L.; A Lindquist, M.; Wager, T.D. Building better biomarkers: Brain models in translational neuroimaging. Nat. Neurosci. 2017, 20, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Syrjälä, J.J.; Basti, A.; Guidotti, R.; Marzetti, L.; Pizzella, V. Decoding working memory task condition using magnetoencephalography source level long-range phase coupling patterns. J. Neural Eng. 2021, 18, 016027. [Google Scholar] [CrossRef] [PubMed]
- Luders, E.; Cherbuin, N.; Gaser, C. Estimating brain age using high-resolution pattern recognition: Younger brains in long-term meditation practitioners. NeuroImage 2016, 134, 508–513. [Google Scholar] [CrossRef]
- Sato, J.R.; Kozasa, E.H.; Russell, T.A.; Radvany, J.; Mello, L.E.A.M.; Lacerda, S.S.; Amaro, E. Brain Imaging Analysis Can Identify Participants under Regular Mental Training. PLoS ONE 2012, 7, e39832. [Google Scholar] [CrossRef]
- Tang, Y.-Y.; Tang, Y.; Tang, R.; Lewis-Peacock, J.A. Brief Mental Training Reorganizes Large-Scale Brain Networks. Front. Syst. Neurosci. 2017, 11, 6. [Google Scholar] [CrossRef]
- Alakörkkö, T.; Saarimäki, H.; Glerean, E.; Saramäki, J.; Korhonen, O. Effects of spatial smoothing on functional brain networks. Eur. J. Neurosci. 2017, 46, 2471–2480. [Google Scholar] [CrossRef]
- Shirer, W.R.; Ryali, S.; Rykhlevskaia, E.; Menon, V.; Greicius, M.D. Decoding Subject-Driven Cognitive States with Whole-Brain Connectivity Patterns. Cereb. Cortex 2011, 22, 158–165. [Google Scholar] [CrossRef]
- Zhang, Y.; Brady, M.; Smith, S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE Trans. Med Imaging 2001, 20, 45–57. [Google Scholar] [CrossRef]
- Power, J.D.; Barnes, K.A.; Snyder, A.Z.; Schlaggar, B.L.; Petersen, S.E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. NeuroImage 2012, 59, 2142–2154. [Google Scholar] [CrossRef]
- Andersson, J.; Jenkinson, M.; Smith, S. High resolution nonlinear registration with simultaneous modelling of intensities. bioRxiv 2019, 646802. [Google Scholar] [CrossRef]
- Varoquaux, G.; Raamana, P.R.; Engemann, D.A.; Hoyos-Idrobo, A.; Schwartz, Y.; Thirion, B. Assessing and tuning brain decoders: Cross-validation, caveats, and guidelines. NeuroImage 2017, 145, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.; Mitchell, T.; Botvinick, M. Machine learning classifiers and fMRI: A tutorial overview. NeuroImage 2009, 45, S199–S209. [Google Scholar] [CrossRef] [PubMed]
- Nichols, T.E.; Holmes, A.P. Nonparametric permutation tests for functional neuroimaging: A primer with examples. Hum. Brain Mapp. 2002, 15, 1–25. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental algorithms for scientific computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef]
- Haufe, S.; Meinecke, F.; Görgen, K.; Dähne, S.; Haynes, J.-D.; Blankertz, B.; Bießmann, F. On the interpretation of weight vectors of linear models in multivariate neuroimaging. NeuroImage 2014, 87, 96–110. [Google Scholar] [CrossRef]
- Snoek, L.; Miletić, S.; Scholte, H. How to control for confounds in decoding analyses of neuroimaging data. NeuroImage 2019, 184, 741–760. [Google Scholar] [CrossRef]
- Bron, E.E.; Smits, M.; van der Flier, W.; Vrenken, H.; Barkhof, F.; Scheltens, P.; Papma, J.M.; Steketee, R.; Orellana, C.M.; Meijboom, R.; et al. Standardized evaluation of algorithms for computer-aided diagnosis of dementia based on structural MRI: The CADDementia challenge. NeuroImage 2015, 111, 562–579. [Google Scholar] [CrossRef] [PubMed]
- Gratton, C.; Laumann, T.O.; Nielsen, A.N.; Greene, D.; Gordon, E.; Gilmore, A.W.; Nelson, S.M.; Coalson, R.S.; Snyder, A.Z.; Schlaggar, B.L.; et al. Functional Brain Networks Are Dominated by Stable Group and Individual Factors, Not Cognitive or Daily Variation. Neuron 2018, 98, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Botvinick, M.M.; Braver, T.S.; Barch, D.M.; Carter, C.S.; Cohen, J.D. Conflict monitoring and cognitive control. Psychol. Rev. 2001, 108, 624. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.E.; Posner, M.I. The Attention System of the Human Brain: 20 Years After. Annu. Rev. Neurosci. 2012, 35, 73–89. [Google Scholar] [CrossRef]
- Thakral, P.P.; Slotnick, S.D. The role of parietal cortex during sustained visual spatial attention. Brain Res. 2009, 1302, 157–166. [Google Scholar] [CrossRef]
- Lee, T.M.C.; Leung, M.-K.; Hou, W.-K.; Tang, J.C.Y.; Yin, J.; So, K.-F.; Lee, C.-F.; Chan, C.C.H. Distinct Neural Activity Associated with Focused-Attention Meditation and Loving-Kindness Meditation. PLoS ONE 2012, 7, e40054. [Google Scholar] [CrossRef]
- Brewer, J.A.; Garrison, K.A. The posterior cingulate cortex as a plausible mechanistic target of meditation: Findings from neuroimaging. Ann. N. Y. Acad. Sci. 2013, 1307, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Greicius, M.D.; Krasnow, B.; Reiss, A.L.; Menon, V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 253–258. [Google Scholar] [CrossRef]
- Pagnoni, G. Dynamical Properties of BOLD Activity from the Ventral Posteromedial Cortex Associated with Meditation and Attentional Skills. J. Neurosci. 2012, 32, 5242–5249. [Google Scholar] [CrossRef] [PubMed]
- Hasenkamp, W.; Barsalou, L.W. Effects of Meditation Experience on Functional Connectivity of Distributed Brain Networks. Front. Hum. Neurosci. 2012, 6, 38. [Google Scholar] [CrossRef]
- Xu, J.; Vik, A.; Groote, I.R.; Lagopoulos, J.; Holen, A.; Ellingsen, Ø.; Håberg, A.K.; Davanger, S. Nondirective meditation activates default mode network and areas associated with memory retrieval and emotional processing. Front. Hum. Neurosci. 2014, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Berkovich-Ohana, A.; Harel, M.; Hahamy, A.; Arieli, A.; Malach, R. Alterations in task-induced activity and resting-state fluctuations in visual and DMN areas revealed in long-term meditators. NeuroImage 2016, 135, 125–134. [Google Scholar] [CrossRef]
- Vago, D.R.; Zeidan, F. The brain on silent: Mind wandering, mindful awareness, and states of mental tranquility. Ann. N. Y. Acad. Sci. 2016, 1373, 96–113. [Google Scholar] [CrossRef]
- Stoodley, C.J. The Cerebellum and Cognition: Evidence from Functional Imaging Studies. Cerebellum 2012, 11, 352–365. [Google Scholar] [CrossRef]
- Gard, T.; Taquet, M.; Dixit, R.; Hã¶lzel, B.K.; Dickerson, B.C.; Lazar, S.W.; Hölzel, B.K. Greater widespread functional connectivity of the caudate in older adults who practice kripalu yoga and vipassana meditation than in controls. Front. Hum. Neurosci. 2015, 9, 137. [Google Scholar] [CrossRef]
- Graff-Radford, J.; Williams, L.; Jones, D.T.; Benarroch, E.E. Caudate nucleus as a component of networks controlling behavior. Neurology 2017, 89, 2192–2197. [Google Scholar] [CrossRef]
- Eichenbaum, H. Hippocampus. Neuron 2004, 44, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Dudley, L.; Stevenson, R.J. Interoceptive awareness and its relationship to hippocampal dependent processes. Brain Cogn. 2016, 109, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Marchand, W.R. Neural mechanisms of mindfulness and meditation: Evidence from neuroimaging studies. World J. Radiol. 2014, 6, 471–479. [Google Scholar] [CrossRef]
- Kuehn, E.; Mueller, K.; Lohmann, G.; Schuetz-Bosbach, S. Interoceptive awareness changes the posterior insula functional connectivity profile. Brain Struct. Funct. 2016, 221, 1555–1571. [Google Scholar] [CrossRef] [PubMed]
- Seghier, M.L. The Angular Gyrus: Multiple Functions and Multiple Subdivisions. Neuroscientist 2012, 19, 43–61. [Google Scholar] [CrossRef]
- D’Andrea, A.; Chella, F.; Marshall, T.R.; Pizzella, V.; Romani, G.L.; Jensen, O.; Marzetti, L. Alpha and alpha-beta phase synchronization mediate the recruitment of the visuospatial attention network through the Superior Longitudinal Fasciculus. NeuroImage 2019, 188, 722–732. [Google Scholar] [CrossRef]
- Marshall, T.R.; Bergmann, T.O.; Jensen, O. Frontoparietal Structural Connectivity Mediates the Top-Down Control of Neuronal Synchronization Associated with Selective Attention. PLoS Biol. 2015, 13, e1002272. [Google Scholar] [CrossRef]
- Fox, K.C.; Dixon, M.L.; Nijeboer, S.; Girn, M.; Floman, J.L.; Lifshitz, M.; Ellamil, M.; Sedlmeier, P.; Christoff, K. Functional neuroanatomy of meditation: A review and meta-analysis of 78 functional neuroimaging investigations. Neurosci. Biobehav. Rev. 2016, 65, 208–228. [Google Scholar] [CrossRef]
- Davidson, R.J. Emotion and Affective Style: Hemispheric Substrates. Psychol. Sci. 1992, 3, 39–43. [Google Scholar] [CrossRef]
- Davidson, R.J.; Kabat-Zinn, J.; Schumacher, J.; Rosenkranz, M.; Muller, D.; Santorelli, S.F.; Urbanowski, F.; Harrington, A.; Bonus, K.; Sheridan, J. Alterations in Brain and Immune Function Produced by Mindfulness Meditation. Psychosom. Med. 2003, 65, 564–570. [Google Scholar] [CrossRef] [PubMed]
- Kurth, F.; MacKenzie-Graham, A.; Toga, A.W.; Luders, E. Shifting brain asymmetry: The link between meditation and structural lateralization. Soc. Cogn. Affect. Neurosci. 2015, 10, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Elaneri, D.; Eschuster, V.; Edietsche, B.; Ejansen, A.; Eott, U.; Esommer, J. Effects of Long-Term Mindfulness Meditation on Brain’s White Matter Microstructure and its Aging. Front. Aging Neurosci. 2016, 7, 254. [Google Scholar] [CrossRef]
- Farb, N.A.S.; Segal, Z.V.; Anderson, A.K. Mindfulness meditation training alters cortical representations of interoceptive attention. Soc. Cogn. Affect. Neurosci. 2013, 8, 15–26. [Google Scholar] [CrossRef]
- Pickut, B.A.; Van Hecke, W.; Kerckhofs, E.; Mariën, P.; Vanneste, S.; Cras, P.; Parizel, P.M. Mindfulness based intervention in Parkinson’s disease leads to structural brain changes on MRI. Clin. Neurol. Neurosurg. 2013, 115, 2419–2425. [Google Scholar] [CrossRef]
- Segerdahl, A.R.; Mezue, M.; Okell, T.; Farrar, J.T.; Tracey, I. The dorsal posterior insula subserves a fundamental role in human pain. Nat. Neurosci. 2015, 18, 499–500. [Google Scholar] [CrossRef]
- Lehmann, D.; Faber, P.L.; Tei, S.; Pascual-Marqui, R.D.; Milz, P.; Kochi, K. Reduced functional connectivity between cortical sources in five meditation traditions detected with lagged coherence using EEG tomography. NeuroImage 2012, 60, 1574–1586. [Google Scholar] [CrossRef]
- Polak, G. Reexamining Jhāna: Towards a Critical Reconstruction of Early Buddhist Soteriology, 1st ed.; UMCS: Lublin, Poland, 2011. [Google Scholar]
- Giannandrea, A.; Simione, L.; Pescatori, B.; Ferrell, K.; Belardinelli, M.O.; Hickman, S.D.; Raffone, A. Effects of the Mindfulness-Based Stress Reduction Program on Mind Wandering and Dispositional Mindfulness Facets. Mindfulness 2018, 10, 185–195. [Google Scholar] [CrossRef]
- Touroutoglou, A.; Zhang, J.; Andreano, J.M.; Dickerson, B.C.; Barrett, L.F. Dissociable Effects of Aging on Salience Subnetwork Connectivity Mediate Age-Related Changes in Executive Function and Affect. Front. Aging Neurosci. 2018, 10, 410. [Google Scholar] [CrossRef]
- Hinkley, L.B.; Mizuiri, D.; Hong, O.; Nagarajan, S.S.; Cheung, S.W. Increased striatal functional connectivity with auditory cortex in tinnitus. Front. Hum. Neurosci. 2015, 9, 568. [Google Scholar] [CrossRef]
- Cui, Z.; Gong, G. The effect of machine learning regression algorithms and sample size on individualized behavioral prediction with functional connectivity features. NeuroImage 2018, 178, 622–637. [Google Scholar] [CrossRef] [PubMed]
- Naselaris, T.; Allen, E.; Kay, K. Extensive sampling for complete models of individual brains. Curr. Opin. Behav. Sci. 2021, 40, 45–51. [Google Scholar] [CrossRef]
- Baldassarre, A.; Lewis, C.; Committeri, G.; Snyder, A.Z.; Romani, G.L.; Corbetta, M. Individual variability in functional connectivity predicts performance of a perceptual task. Proc. Natl. Acad. Sci. USA 2012, 109, 3516–3521. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.D.; Finn, E.S.; Scheinost, D.; Papademetris, X.; Shen, X.; Constable, R.; Chun, M. A neuromarker of sustained attention from whole-brain functional connectivity. Nat. Neurosci. 2016, 19, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Salice, S.; Esposito, R.; Ciavardelli, D.; Pizzi, S.D.; Di Bastiano, R.; Tartaro, A. Combined 3 Tesla MRI Biomarkers Improve the Differentiation between Benign vs Malignant Single Ring Enhancing Brain Masses. PLoS ONE 2016, 11, e0159047. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Yoshihara, Y.; Hashimoto, R.; Yahata, N.; Ichikawa, N.; Sakai, Y.; Yamada, T.; Matsukawa, N.; Okada, G.; Tanaka, S.C.; et al. A prediction model of working memory across health and psychiatric disease using whole-brain functional connectivity. eLife 2018, 7, e38844. [Google Scholar] [CrossRef] [PubMed]
- Chiarelli, A.M.; Bianco, F.; Perpetuini, D.; Bucciarelli, V.; Filippini, C.; Cardone, D.; Zappasodi, F.; Gallina, S.; Merla, A. Data-driven assessment of cardiovascular ageing through multisite photoplethysmography and electrocardiography. Med Eng. Phys. 2019, 73, 39–50. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guidotti, R.; Del Gratta, C.; Perrucci, M.G.; Romani, G.L.; Raffone, A. Neuroplasticity within and between Functional Brain Networks in Mental Training Based on Long-Term Meditation. Brain Sci. 2021, 11, 1086. https://doi.org/10.3390/brainsci11081086
Guidotti R, Del Gratta C, Perrucci MG, Romani GL, Raffone A. Neuroplasticity within and between Functional Brain Networks in Mental Training Based on Long-Term Meditation. Brain Sciences. 2021; 11(8):1086. https://doi.org/10.3390/brainsci11081086
Chicago/Turabian StyleGuidotti, Roberto, Cosimo Del Gratta, Mauro Gianni Perrucci, Gian Luca Romani, and Antonino Raffone. 2021. "Neuroplasticity within and between Functional Brain Networks in Mental Training Based on Long-Term Meditation" Brain Sciences 11, no. 8: 1086. https://doi.org/10.3390/brainsci11081086
APA StyleGuidotti, R., Del Gratta, C., Perrucci, M. G., Romani, G. L., & Raffone, A. (2021). Neuroplasticity within and between Functional Brain Networks in Mental Training Based on Long-Term Meditation. Brain Sciences, 11(8), 1086. https://doi.org/10.3390/brainsci11081086