
New Approach for Untangling the Role of Uncommon Calcium-Binding Proteins in the Central Nervous System


Abstract
1. About Ca2+ Briefly
2. Calcium in Neurons
3. The Role of Ca2+ Binding
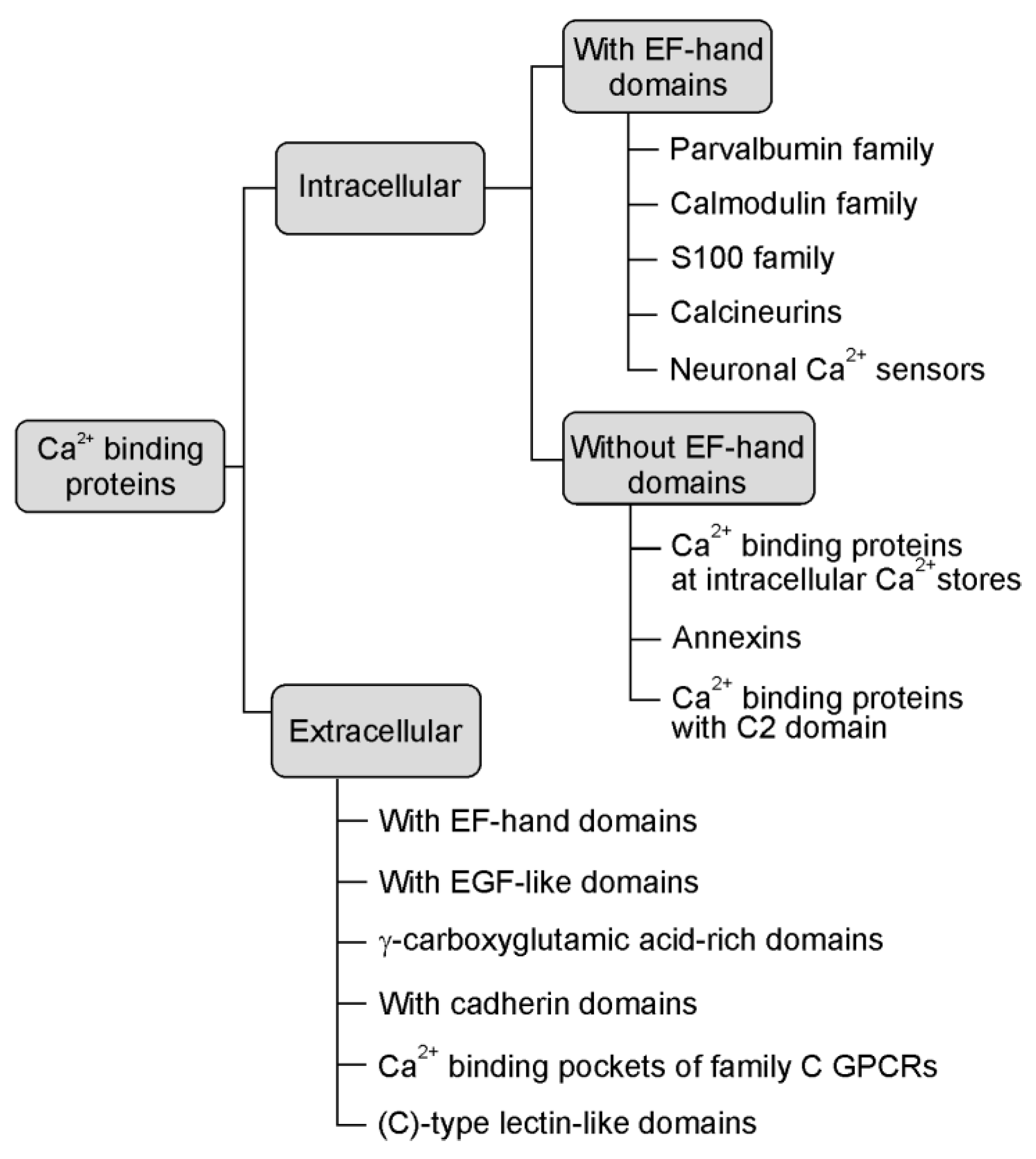
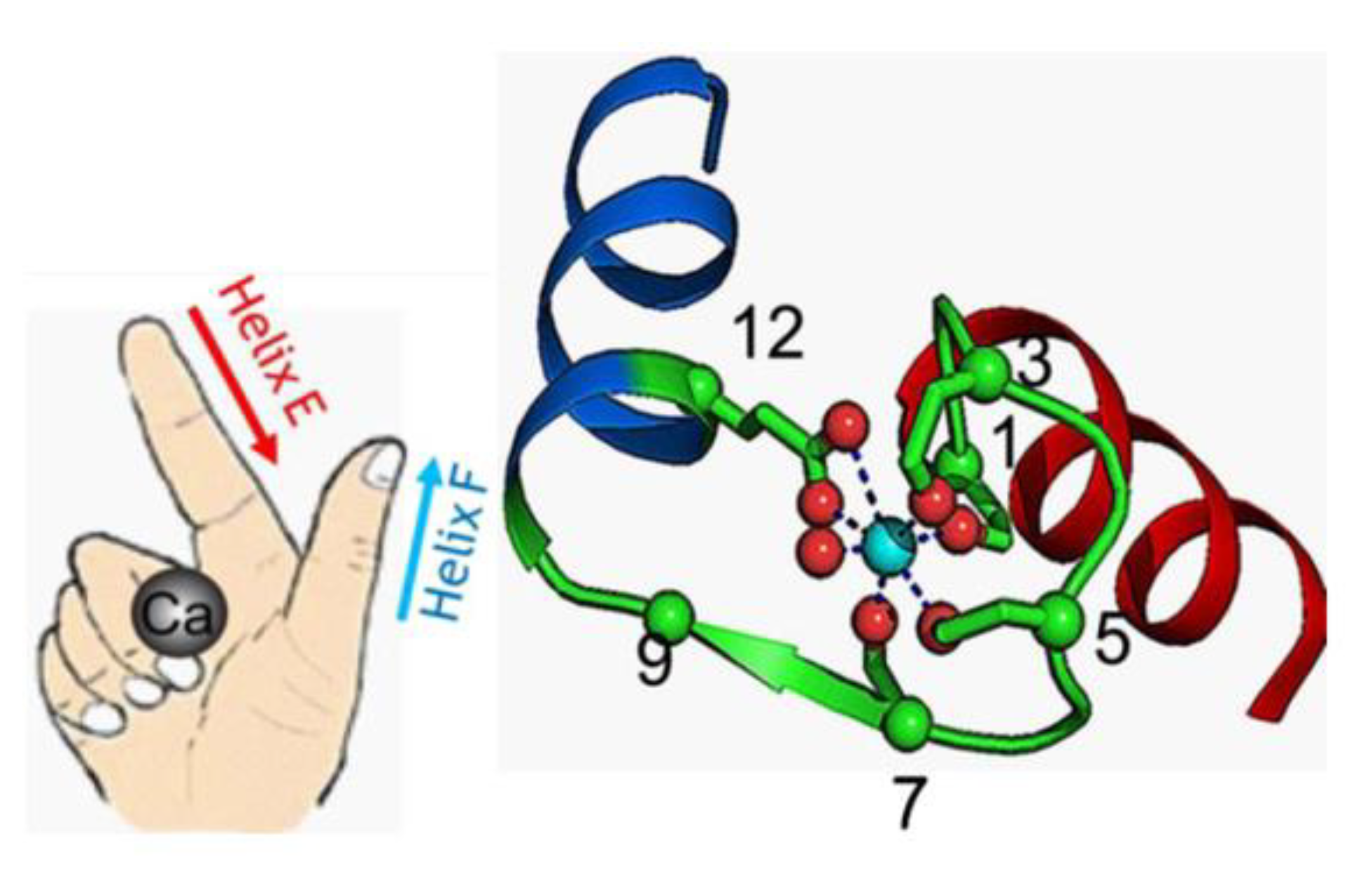
4. The EF-Hand Calcium-Binding Proteins
5. The Regional Arrangement of the EF-Hand CaBPs Throughout the Central Nervous System (CNS)
6. The “Classical” CaBPs of the CNS
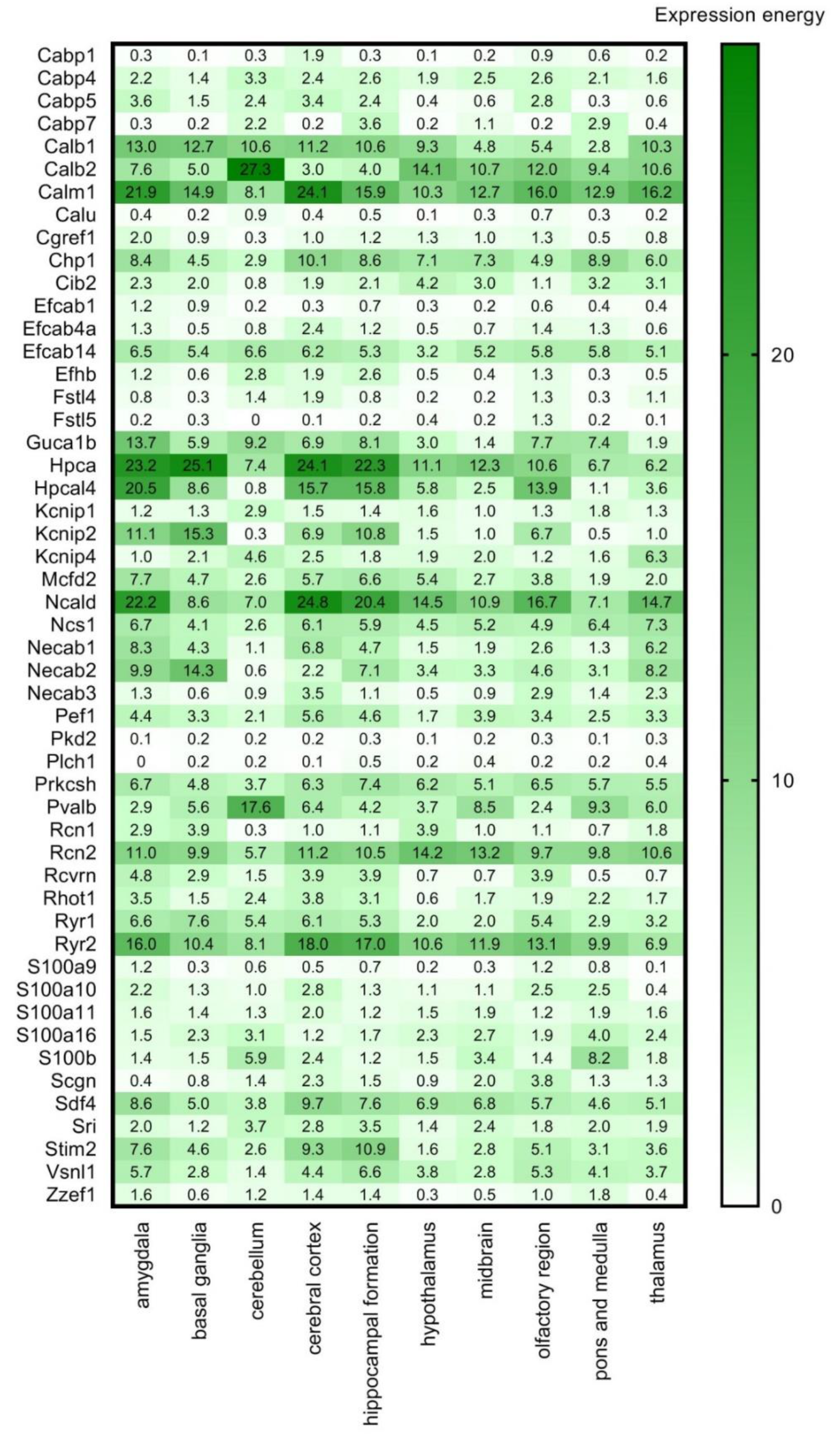
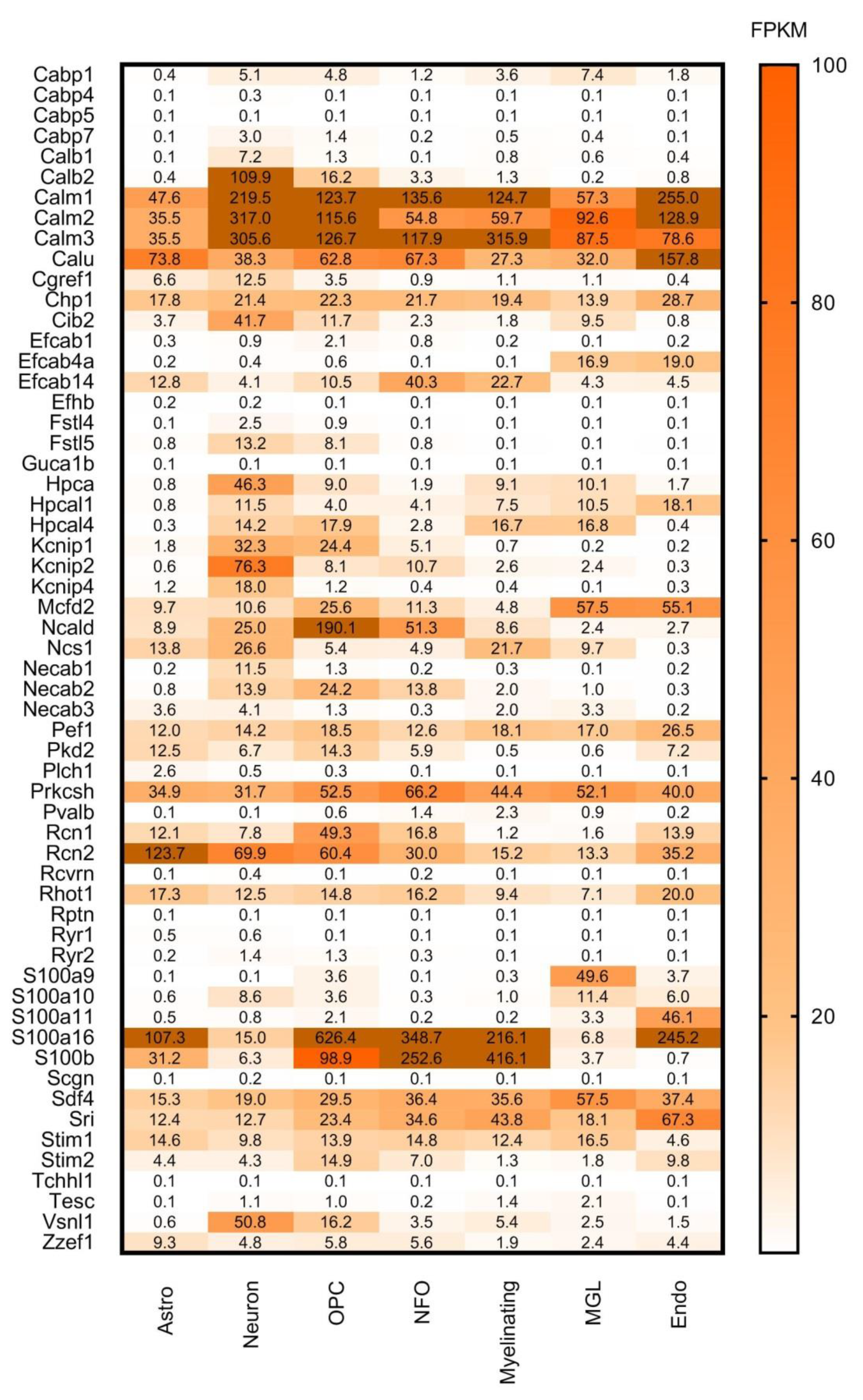
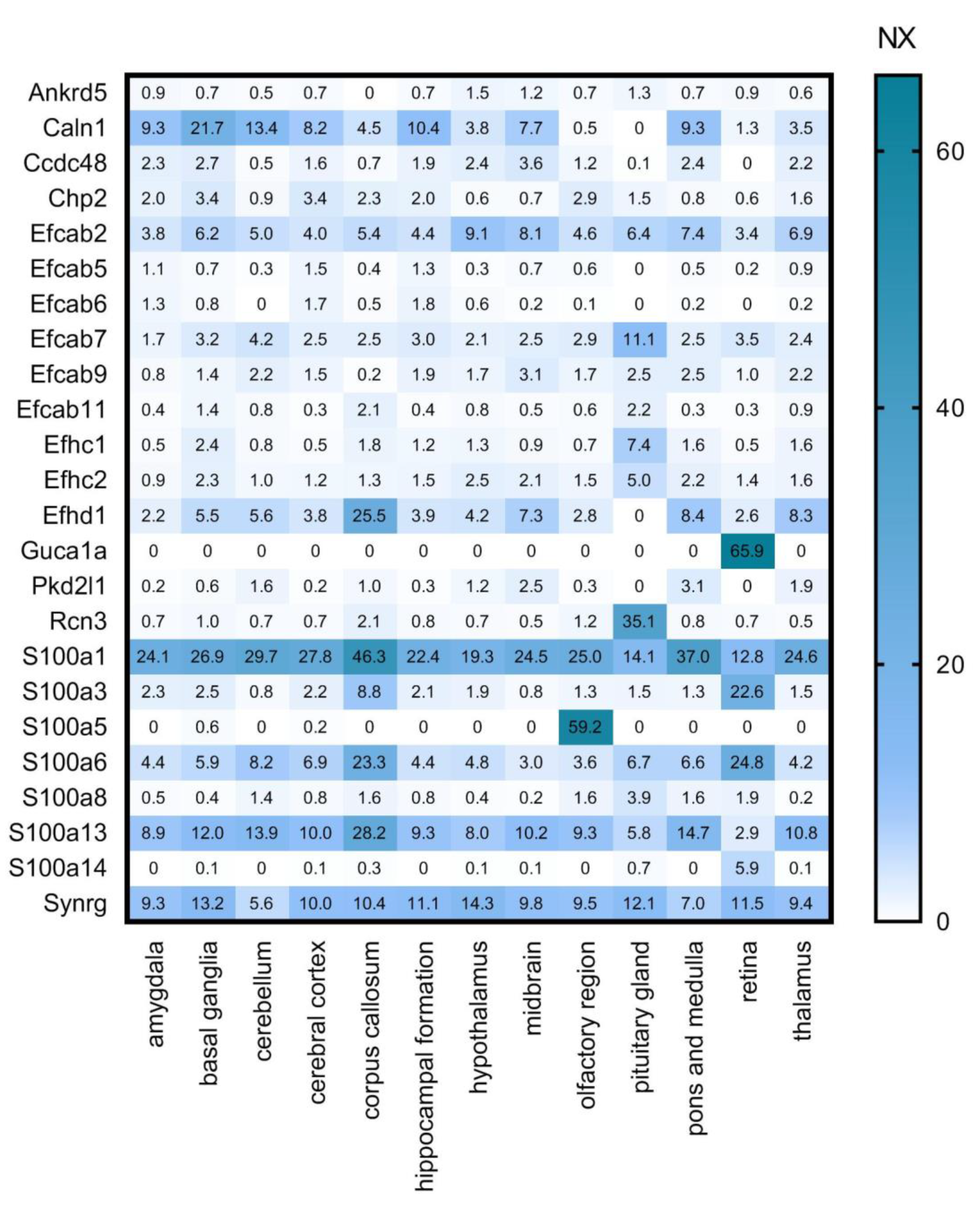
7. Search for New CaBPs. Detailed Analysis of RNA-Seq and ISH Databases
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blaustein, M.P. Calcium transport and buffering in neurons. Trends Neurosci. 1988, 11, 438–443. [Google Scholar] [CrossRef]
- Kasai, H. Comparative biology of Ca2+-dependent exocytosis: Implications of kinetic diversity for secretory function. Trends Neurosci. 1999, 22, 88–93. [Google Scholar] [CrossRef]
- Kandel, E.R.; Schwartz, J.H.; Jessell, T.M.; Siegelbaum, S.A.; Hudspeth, A.J. Principles of Neural Science, 5th ed.; Health Professions Division; McGraw-Hill: New York, NY, USA, 2013. [Google Scholar]
- Augustine, G.J.; Santamaria, F.; Tanaka, K. Local Calcium Signaling in Neurons. Neuron 2003, 40, 331–346. [Google Scholar] [CrossRef]
- Lodish, H. Molecular Cell Biology, 8th ed.; W.H. Freeman: New York, NY, USA, 2016. [Google Scholar]
- Gleichmann, M.; Mattson, M.P. Neuronal Calcium Homeostasis and Dysregulation. Antioxid. Redox Signal. 2011, 14, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef]
- Bastianelli, E. Distribution of calcium-binding proteins in the cerebellum. Cerebellum 2003, 2, 242–262. [Google Scholar] [CrossRef] [PubMed]
- Elíes, J.; Yáñez, M.; Pereira, T.M.C.; Gil-Longo, J.; MacDougall, D.A.; Campos-Toimil, M. An Update to Calcium Binding Proteins. Adv. Exp. Med. Biol. 2019, 1131, 183–213. [Google Scholar]
- Xu, J.-H.; Tang, F.-R. Voltage-Dependent Calcium Channels, Calcium Binding Proteins, and Their Interaction in the Pathological Process of Epilepsy. Int. J. Mol. Sci. 2018, 19, 2735. [Google Scholar] [CrossRef] [PubMed]
- Bagur, R.; Hajnóczky, G. Intracellular Ca2+ Sensing: Its Role in Calcium Homeostasis and Signaling. Mol. Cell 2017, 66, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Yáñez, M.; Gil-Longo, J.; Campos-Toimil, M. Calcium Binding Proteins. In Advances in Experimental Medicine and Biology; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2012; Volume 740, pp. 461–482. [Google Scholar]
- Tang, S.; Deng, X.; Jiang, J.; Kirberger, M.; Yang, J.J. Design of Calcium-Binding Proteins to Sense Calcium. Molecules 2020, 25, 2148. [Google Scholar] [CrossRef] [PubMed]
- Schwaller, B. The continuing disappearance of “pure” Ca2+ buffers. Cell. Mol. Life Sci. 2009, 66, 275–300. [Google Scholar] [CrossRef]
- Lewit-Bentley, A.; Réty, S. EF-hand calcium-binding proteins. Curr. Opin. Struct. Biol. 2000, 10, 637–643. [Google Scholar] [CrossRef]
- Zhou, Y.; Frey, T.K.; Yang, J.J. Viral calciomics: Interplays between Ca2+ and virus. Cell Calcium 2009, 46, 1–17. [Google Scholar] [CrossRef]
- Gifford, J.L.; Walsh, M.P.; Vogel, H.J. Structures and metal-ion-binding properties of the Ca2+-binding helix–loop–helix EF-hand motifs. Biochem. J. 2007, 405, 199–221. [Google Scholar] [CrossRef] [PubMed]
- Andressen, C.; Blümcke, I.; Celio, M.R. Calcium-binding proteins: Selective markers of nerve cells. Cell Tissue Rev. Artic. 1993, 271, 181–208. [Google Scholar] [CrossRef]
- McMahon, S.M.; Jackson, M.B. An Inconvenient Truth: Calcium Sensors Are Calcium Buffers. Trends Neurosci. 2018, 41, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Kreutz, M.R.; Naranjo, J.R.; Koch, K.; Schwaller, B. The Neuronal Functions of EF-hand Ca(2+)-binding Proteins. Front. Mol. Neurosci. 2012, 5, 92. [Google Scholar] [CrossRef]
- Gattoni, G.; Bernocchi, G. Calcium-Binding Proteins in the Nervous System during Hibernation: Neuroprotective Strategies in Hypometabolic Conditions? Int. J. Mol. Sci. 2019, 20, 2364. [Google Scholar] [CrossRef] [PubMed]
- Schwaller, B. Emerging Functions of the “Ca2+ Buffers” Parvalbumin, Calbindin D-28k and Calretinin in the Brain. In Handbook of Neurochemistry and Molecular Neurobiology; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2007; pp. 197–221. [Google Scholar]
- Alpár, A.; Attems, J.; Mulder, J.; Hökfelt, T.; Harkany, T. The renaissance of Ca2+-binding proteins in the nervous system: Secretagogin takes center stage. Cell. Signal. 2012, 24, 378–387. [Google Scholar] [CrossRef]
- Kántor, O.; Mezey, S.; Adeghate, J.; Naumann, A.; Nitschke, R.; Énzsöly, A.; Szabó, A.; Lukáts, Á.; Németh, J.; Somogyvári, Z.; et al. Calcium buffer proteins are specific markers of human retinal neurons. Cell Tissue Res. 2016, 365, 29–50. [Google Scholar] [CrossRef]
- Mulder, J.; Zilberter, M.; Spence, L.; Tortoriello, G.; Uhlén, M.; Yanagawa, Y.; Aujard, F.; Hökfelt, T.; Harkany, T. Secretagogin is a Ca2+-binding protein specifying subpopulations of telencephalic neurons. Proc. Natl. Acad. Sci. USA 2009, 106, 22492–22497. [Google Scholar] [CrossRef]
- Fairless, R.; Williams, S.K.; Diem, R. Calcium-Binding Proteins as Determinants of Central Nervous System Neuronal Vulnerability to Disease. Int. J. Mol. Sci. 2019, 20, 2146. [Google Scholar] [CrossRef]
- Miczán, V.; Kelemen, K.; Glavinics, J.R.; László, Z.I.; Barti, B.; Kenesei, K.; Kisfali, M.; Katona, I. NECAB1 and NECAB2 are Prevalent Calcium-Binding Proteins of CB1/CCK-Positive GABAergic Interneurons. Cereb. Cortex 2021, 31, 1786–1806. [Google Scholar] [CrossRef] [PubMed]
- Klausberger, T.; Somogyi, P. Neuronal Diversity and Temporal Dynamics: The Unity of Hippocampal Circuit Operations. Science 2008, 321, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Pelkey, K.A.; Chittajallu, R.; Craig, M.T.; Tricoire, L.; Wester, J.C.; McBain, C.J. Hippocampal GABAergic inhibitory interneurons. Physiol. Rev. 2017, 97, 1619–1747. [Google Scholar] [CrossRef] [PubMed]
- Girard, F.; Venail, J.; Schwaller, B.; Celio, M. The EF-hand Ca2+-binding protein super-family: A genome-wide analysis of gene expression patterns in the adult mouse brain. Neuroscience 2015, 294, 116–155. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a knowledge-based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Sjöstedt, E.; Zhong, W.; Fagerberg, L.; Karlsson, M.; Mitsios, N.; Adori, C.; Oksvold, P.; Edfors, F.; Limiszewska, A.; Hikmet, F.; et al. An atlas of the protein-coding genes in the human, pig, and mouse brain. Science 2020, 367, eaay5947. [Google Scholar] [CrossRef] [PubMed]
- Ezaldivar, A.; Krichmar, J.L. Allen Brain Atlas-Driven Visualizations: A web-based gene expression energy visualization tool. Front. Aging Neurosci. 2014, 8, 51. [Google Scholar]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-Sequencing Transcriptome and Splicing Database of Glia, Neurons, and Vascular Cells of the Cerebral Cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef]
- Haeseleer, F.; Imanishi, Y.; Maeda, T.; E Possin, D.; Maeda, A.; Lee, A.; Rieke, F.; Palczewski, K. Essential role of Ca2+-binding protein 4, a Cav1.4 channel regulator, in photoreceptor synaptic function. Nat. Neurosci. 2004, 7, 1079–1087. [Google Scholar] [CrossRef]
- Haeseleer, F.; Sokal, I.; Verlinde, C.L.M.J.; Erdjument-Bromage, H.; Tempst, P.; Pronin, A.N.; Benovic, J.L.; Fariss, R.N.; Palczewski, K. Five Members of a Novel Ca2+-binding Protein (CABP) Subfamily with Similarity to Calmodulin. J. Biol. Chem. 2000, 275, 1247–1260. [Google Scholar] [CrossRef]
- Haverkamp, S.; Ghosh, K.K.; Hirano, A.A.; Wässle, H. Immunocytochemical description of five bipolar cell types of the mouse retina. J. Comp. Neurol. 2002, 455, 463–476. [Google Scholar] [CrossRef]
- Ghosh, K.K.; Bujan, S.; Haverkamp, S.; Feigenspan, A.; Wässle, H. Types of bipolar cells in the mouse retina. J. Comp. Neurol. 2003, 469, 70–82. [Google Scholar] [CrossRef]
- Makino, C.L.; Peshenko, I.V.; Wen, X.-H.; Olshevskaya, E.V.; Barrett, R.; Dizhoor, A.M. A Role for GCAP2 in Regulating the Photoresponse. J. Biol. Chem. 2008, 283, 29135–29143. [Google Scholar] [CrossRef] [PubMed]
- Puthussery, T.; Gayet-Primo, J.; Taylor, W.R. Localization of the calcium-binding protein secretagogin in cone bipolar cells of the mammalian retina. J. Comp. Neurol. 2009, 518, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Ames, J.B.; Ishima, R.; Tanaka, T.; Gordon, J.I.; Stryer, L.; Ikura, M. Molecular mechanics of calcium–myristoyl switches. Nat. Cell Biol. 1997, 389, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Borsatto, A.; Marino, V.; Abrusci, G.; Lattanzi, G.; Dell’Orco, D. Effects of Membrane and Biological Target on the Structural and Allosteric Properties of Recoverin: A Computational Approach. Int. J. Mol. Sci. 2019, 20, 5009. [Google Scholar] [CrossRef]
- Chen, Y.; Palczewski, K. Systems Pharmacology Links GPCRs with Retinal Degenerative Disorders. Annu. Rev. Pharm. Toxicol. 2016, 56, 273–298. [Google Scholar] [CrossRef]
- Xiong, H.; Kovacs, I.; Zhang, Z. Differential distribution of KChIPs mRNAs in adult mouse brain. Mol. Brain Res. 2004, 128, 103–111. [Google Scholar] [CrossRef]
- Shibata, R.; Misonou, H.; Campomanes, C.R.; Anderson, A.E.; Schrader, L.A.; Doliveira, L.C.; Carroll, K.I.; Sweatt, J.D.; Rhodes, K.J.; Trimmer, J.S. A Fundamental Role for KChIPs in Determining the Molecular Properties and Trafficking of Kv4.2 Potassium Channels. J. Biol. Chem. 2003, 278, 36445–36454. [Google Scholar] [CrossRef] [PubMed]
- Misonou, H.; Trimmer, J.S.; Misono, H. Determinants of Voltage-Gated Potassium Channel Surface Expression and Localization in Mammalian Neurons. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 125–145. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, K.J.; Carroll, K.I.; Sung, M.A.; Doliveira, L.C.; Monaghan, M.M.; Burke, S.L.; Strassle, B.W.; Buchwalder, L.; Menegola, M.; Cao, J.; et al. KChIPs and Kv4 Subunits as Integral Components of A-Type Potassium Channels in Mammalian Brain. J. Neurosci. 2004, 24, 7903–7915. [Google Scholar] [CrossRef] [PubMed]
- Dabrowska, J.; Rainnie, D.G. Expression and distribution of Kv4 potassium channel subunits and potassium channel inter-acting proteins in subpopulations of interneurons in the basolateral amygdala. Neuroscience 2010, 171, 721–733. [Google Scholar] [CrossRef]
- Norris, A.J.; Foeger, N.C.; Nerbonne, J.M. Interdependent Roles for Accessory KChIP2, KChIP3, and KChIP4 Subunits in the Generation of Kv4-Encoded IA Channels in Cortical Pyramidal Neurons. J. Neurosci. 2010, 30, 13644–13655. [Google Scholar] [CrossRef]
- Néant, I.; Haiech, J.; Kilhoffer, M.-C.; Aulestia, F.J.; Moreau, M.; Leclerc, C. Ca2+-Dependent Transcriptional Repressors KCNIP and Regulation of Prognosis Genes in Glioblastoma. Front. Mol. Neurosci. 2018, 11, 472. [Google Scholar] [CrossRef]
- Takamatsu, K.; Noguchi, T. Hippocalcin: A calcium-binding protein of the EF-hand superfamily dominantly expressed in the hippocampus. Neurosci. Res. 1993, 17, 291–295. [Google Scholar] [CrossRef]
- Saitoh, S.; Takamatsu, K.; Kobayashi, M.; Noguchi, T. Distribution of hippocalcin mRNA and immunoreactivity in rat brain. Neurosci. Lett. 1993, 157, 107–110. [Google Scholar] [CrossRef]
- Palmer, C.L.; Lim, W.; Hastie, P.G.; Toward, M.; Korolchuk, V.I.; Burbidge, S.A.; Banting, G.; Collingridge, G.L.; Isaac, J.T.; Henley, J.M. Hippocalcin Functions as a Calcium Sensor in Hippocampal LTD. Neuron 2005, 47, 487–494. [Google Scholar] [CrossRef]
- Tzingounis, A.V.; Kobayashi, M.; Takamatsu, K.; Nicoll, R.A. Hippocalcin Gates the Calcium Activation of the Slow Afterhyperpolarization in Hippocampal Pyramidal Cells. Neuron 2007, 53, 487–493. [Google Scholar] [CrossRef]
- Braunewell, K.-H.; Gundelfinger, E.D. Intracellular neuronal calcium sensor proteins: A family of EF-hand calcium-binding proteins in search of a function. Cell Tissue Res. 1999, 295, 1–12. [Google Scholar] [CrossRef]
- Zhao, C.; Braunewell, K.-H. Expression of the neuronal calcium sensor visinin-like protein-1 in the rat hippocampus. Neuroscience 2008, 153, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- Braunewell, K.H. The visinin-like proteins VILIP-1 and VILIP-3 in Alzheimer’s disease—Old wine in new bottles. Front. Mol. Neurosci. 2012, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Liebl, M.P.; Kaya, A.M.; Tenzer, S.; Mittenzwei, R.; Koziollek-Drechsler, I.; Schild, H.; Moosmann, B.; Behl, C.; Clement, A.M. Dimerization of visinin-like protein 1 is regulated by oxidative stress and calcium and is a pathological hallmark of amyotrophic lateral sclerosis. Free. Radic. Biol. Med. 2014, 72, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-W.; Chang, L.-C.; Tseng, G.C.; Kirkwood, C.M.; Sibille, E.L.; Sweet, R.A. VSNL1 Co-Expression Networks in Aging Include Calcium Signaling, Synaptic Plasticity, and Alzheimer’s Disease Pathways. Front. Psychiatry 2015, 6, 30. [Google Scholar] [CrossRef]
- Recabarren-Leiva, D.; Alarcón, M. New insights into the gene expression associated to amyotrophic lateral sclerosis. Life Sci. 2018, 193, 110–123. [Google Scholar] [CrossRef]
- Palecek, J.; Lips, M.B.; Keller, B.U. Calcium dynamics and buffering in motoneurones of the mouse spinal cord. J. Physiol. 1999, 520, 485–502. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Complete Name (Alternative Name) | Gene Ontology/CNS Function(s) |
|---|---|---|
| Cabp1 | Ca2+-binding protein 1, (Caldendrin) | Modulator of Ca2+ channel activity; Ca2+ sensor; fine-tuning of CaV.1/2 |
| Cabp4 | Ca2+ -binding protein 4 | Modulator of Ca2+ channel activity |
| Cabp5 | Ca2+-binding protein 5 | Modulator of Ca2+ channel activity |
| Cabp7 | Ca2+-binding protein 7, (Calneuron-2) | Modulator of Ca2+ channel activity |
| Calb1 | Calbindin D-28 K | Ca2+ sensor/buffer activity |
| Calb2 | Calretinin, (Calbindin 2) | Ca2+ sensor/buffer activity |
| Calm1 | Calmodulin 1 | Ca2+ sensor/Ca2+ signalling |
| Calm2 | Calmodulin 2 | Ca2+ sensor/Ca2+ signalling |
| Calm3 | Calmodulin 3 | Ca2+ sensor/Ca2+ signalling |
| Calu | Calumenin (Crocalbin) | Unknown |
| Cgref1 | Cell growth regulator with EF-hand domain 1 | Unknown |
| Chp1 | Calcineurin-like EF-hand protein 1, (1500003O03Rik RIKEN cDNA 1500003O03 gene) | Unknown |
| Cib2 | Ca2+- and integrin-binding family member 2 (Calmyrin 2, Kip2) | Unknown |
| Efcab1 | EF-hand Ca2+-binding domain 1 | Unknown |
| Efcab4a | EF-hand Ca2+-binding domain 4A, (Cracr2b) | Ca2+ sensor activity |
| Efcab12 | EF-hand Ca2+-binding domain 12 (BC060267) | Unknown |
| Efcab14 | EF-hand calcium binding domain 14, (4732418C07Rik, RIKEN cDNA 4732418C07 gene, Kiaa0494) | Unknown |
| Efhb | EF-hand domain family member B | Unknown |
| Fstl4 | Follistatin-like 4 (Spig1) | Unknown; negative regulator of BDNF maturation |
| Fstl5 | Follistatin-like 5 | Unknown |
| Guca1b | Guanylate cyclase activator 1B (Gcap2, Rp48) | Ca2+-sensitive guanylate cyclase activator activity, Ca2+ sensor activity |
| Hpca | Hippocalcin | Ca2+ sensor activity |
| Hpcal1 | Hippocalcin-like 1 (Vilip3, Visinin-like protein 3) | Ca2+ sensor activity |
| Hpcal4 | Hippocalcin-like 4 (Vilip2, Neural visinin-like protein 2) | Ca2+ sensor activity |
| Kcnip1 | kV channel-interacting protein 1 (Kchip1) | Ca2+ sensor activity; modulation of Kv4 activity/control of neuronal excitability |
| Kcnip2 | kV channel-interacting protein 2 (Kchip2) | Ca2+ sensor activity; modulation of Kv4 activity/control of neuronal excitability |
| Kcnip4 | kV channel-interacting protein 4 (Kchip4, Calp) | Ca2+ sensor activity; modulation of Kv4 activity/control of neuronal excitability |
| Mcfd2 | Multiple coagulation factor deficiency 2 (F5F8D, LMAN1IP, SDNSF) | Unknown; survival factor for neural stem cells |
| Ncald | Neurocalcin delta | Ca2+ sensor activity |
| Ncs1 | Neuronal Ca2+ sensor1 (frequenin homolog) | Ca2+ sensor activity; modulation of synaptic plasticity/neuronal secretion |
| Necab1 | N-terminal EF-hand Ca2+-binding protein 1 | Unknown |
| Necab2 | N-terminal EF-hand Ca2+-binding protein 2 | Unknown |
| Necab3 | N-terminal EF-hand Ca2+-binding protein 3 | Unknown |
| Pef1 | Penta-EF-hand domain containing 1 (Peflin) | Unknown |
| Pkd2 | Polycystic kidney disease 2 (PKD4, PC2, TRPP2) | Ca2+ channel activity |
| Plch1 | Phospholipase C eta 1, (Kiaa1069) | Phosphoinositide phospholipase C activity; Ca2+ sensor activity |
| Prkcsh | Protein kinase C substrate 80 K-H, (Glucosidase 2 subunit beta) | Ca2+ sensor activity |
| Pvalb | Parvalbumin (Parvalbumin alpha) | Ca2+ sensor/buffer activity |
| Rcn1 | Reticulocalbin 1 (Rcal) | Unknown |
| Rcn2 | Reticulocalbin 2 | Unknown |
| Rcvrn | Recoverin | Ca2+ buffer activity in phototransduction; Ca2+ sensor activity |
| Rhot1 | Ras homolog gene family, member T1 (Miro1) | GTPase activity; Ca2+ sensor activity/mitochondrial trafficking in neurons |
| Rptn | Repetin | Unknown |
| Ryr1 | Ryanodine receptor 1 | Ryanodine-sensitive Ca2+-release channel activity |
| Ryr2 | Ryanodine receptor 2, cardiac | Ryanodine-sensitive Ca2+-release channel activity |
| S100a9 | S100 Ca2+-binding protein A9 (Calgranulin B) | Ca2+ buffer activity; neuro-inflammatory process |
| S100a10 | S100 Ca2+-binding protein A10 (Calpactin) | Ca2+ buffer activity; serotonergic signalling |
| S100a11 | S100 Ca2+-binding protein A11 (Calgizzarin) | Ca2+ buffer activity |
| S100a16 | S100 Ca2++-binding protein A16 (Protein S100F) | Ca2+ buffer activity |
| S100b | S100 protein, beta polypeptide, neural | Ca2+ buffer activity; biological marker of brain damage |
| Scgn | Secretagogin | Ca2+ sensor/buffer activity |
| Sdf4 | Stromal cell-derived factor 4 (Cab45) | Unknown |
| Sri | Sorcin | Intracellular Ca2+ transport; modulator of ryanodine-sensitive Ca2+-release channel |
| Stim1 | Stromal interaction molecule 1 | Ca2+ sensor activity; Ca2+ signalling/storage/release |
| Stim2 | Stromal interaction molecule 2 (Kiaa1482) | Ca2+ sensor activity |
| Tchhl1 | Trichohyalin-like 1 (S100a17) | Unknown |
| Tesc | Tescalcin (Calcineurin-like protein 3) | Unknown |
| Vsnl1 | Visinin-like 1 (Vilip-1, Hippocalcin-like protein 3) | Ca2+ sensor activity; regulator of receptors (P2X, glycine, nicotinic acetylcholine) |
| Zzef1 | Zinc finger, ZZ-type with EF-hand domain 1, (Kiaa0399) | Unknown |
| Gene | Complete Name (Alternative Name) | Gene Ontology/CNS Function(s) |
|---|---|---|
| Ankrd5 | Ankyrin repeat domain 5 (Ankef1) | Unknown |
| Caln1 | Calneuron 1 | Ca2+-buffer/sensor activity |
| Ccdc48 | Coiled-coil domain containing 48 (Efcc1, EF-hand and coiled-coil domain containing 1) | Unknown |
| Chp2 | Calcineurin B homologous protein 2 (2010110p09rik, RIKEN cDNA 2010110P09 gene) | Unknown |
| Efcab2 | EF-hand Ca2+-binding domain 2 | Unknown |
| Efcab5 | EF-hand Ca2+-binding domain 5 | Unknown |
| Efcab6 | EF-hand Ca2+-binding domain 6 | Unknown |
| Efcab7 | EF-hand Ca2+-binding domain 7 | Unknown |
| Efcab9 | EF-hand Ca2+-binding domain 9 | Unknown |
| Efcab4b | EF-hand Ca2+-binding domain 4B (CRACR2A) | Unknown |
| Efcab10 | EF-hand Ca2+-binding domain 10 | Unknown |
| Efcab11 | EF-hand Ca2+-binding domain 11 (Egfem1, EGF-like and EMI domain containing 1) | Unknown |
| Efhc1 | EF-hand domain (C-terminal) containing 1 | Unknown |
| Efhc2 | EF-hand domain (C-terminal) containing 2 | Unknown |
| Efhd1 | EF-hand domain containing 1 | Unknown |
| Guca1a | Guanylate cyclase activator 1a | Ca2+-sensitive guanylate cyclase activator activity, Ca2+ sensor activity |
| Pkd2l1 | Polycystic kidney disease 2-like 1 | Cation channel activity |
| Rcn3 | Reticulocalbin 3 | Unknown |
| S100a1 | S100 Ca2+-binding protein A1 | Ca2+-buffer activity |
| S100a3 | S100 Ca2+-binding protein A3 | Ca2+-buffer activity |
| S100a4 | S100 Ca2+-binding protein A4 | Ca2+-buffer activity |
| S100a5 | S100 Ca2+-binding protein A5 | Ca2+-buffer activity |
| S100a6 | S100 Ca2+-binding protein A6 (Calcyclin) | Ca2+-buffer activity |
| S100a7a | 100 Ca2+-binding protein A7A | Ca2+-buffer activity |
| S100a8 | S100 Ca2+-binding protein A8 (Calgranulin A) | Ca2+-buffer activity |
| S100a13 | S100 Ca2+-binding protein A13 | Ca2+-buffer activity |
| S100a14 | S100 Ca2+-binding protein A14 | Ca2+-buffer activity |
| Synrg | Synergin, gamma | Unknown |
| 1700109H08Rik | RIKEN cDNA 1700109H08 gene | Unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelemen, K.; Szilágyi, T. New Approach for Untangling the Role of Uncommon Calcium-Binding Proteins in the Central Nervous System. Brain Sci. 2021, 11, 634. https://doi.org/10.3390/brainsci11050634
Kelemen K, Szilágyi T. New Approach for Untangling the Role of Uncommon Calcium-Binding Proteins in the Central Nervous System. Brain Sciences. 2021; 11(5):634. https://doi.org/10.3390/brainsci11050634
Chicago/Turabian StyleKelemen, Krisztina, and Tibor Szilágyi. 2021. "New Approach for Untangling the Role of Uncommon Calcium-Binding Proteins in the Central Nervous System" Brain Sciences 11, no. 5: 634. https://doi.org/10.3390/brainsci11050634
APA StyleKelemen, K., & Szilágyi, T. (2021). New Approach for Untangling the Role of Uncommon Calcium-Binding Proteins in the Central Nervous System. Brain Sciences, 11(5), 634. https://doi.org/10.3390/brainsci11050634