
The Cortical “Upper Motoneuron” in Health and Disease

{kind=link}
Abstract
1. A Critique of the Term ‘Upper Motoneuron’
2. Origins of the Corticospinal Tract Projection (CSP)
3. The Wide Spectrum of Primate Corticospinal Fibres
4. Corticospinal Projections and Terminations
5. Somatotopy in the Corticospinal Tract
6. The Origin of the Cortico-Motoneuronal Projections
7. Organisation of the CM System as an Exemplar of the UMN
8. What Does the CM System Contribute to Skilled Movement?
9. The UMN and Amyotrophic Lateral Sclerosis
- A general concept on the function of the CM system is that it may subserve complex adaptive motor behaviours (including the examples of skilled tool use, but also vocalisation and advanced forms of locomotion), which obviously characterise much of the human motor repertoire [58,60]. It is striking that it is these same highly evolved behaviours that are affected in the early forms of the disease, as commented on over 130 years ago by Hughlings Jackson in his Croonian Lecture [61].
- The split-hand syndrome, with dramatic differences in the strength of thenar vs. hypothenar intrinsic hand muscles is now recognised as a key feature in some forms of ALS [56,62]. This pattern of weakness appears to reflect loss of monosynaptic corticospinal inputs to the respective motoneuron pools that were first shown in the non-human primate, where stimulation of motor cortex was shown to evoke significantly larger CM EPSPs in thenar vs. hypothenar motoneurons [63]. For the intrinsic thumb muscles, it may be that the CM input represents such a large component of the excitatory input that when this input is compromised by ALS, the result is both a weakness and a poverty of movement. In the case of the split-hand syndrome, there is now good evidence that the pattern of weakness in ALS patients reflects changes in the corticospinal influence over these muscles, with clear changes in the short latency MEP [62]. This MEP is known to be dominated by direct CM action but may include other complex actions set up by the TMS pulse. Since investigation of the LMN in these patients suggests that peripheral changes do not make a significant contribution to the development of the split-hand phenomenon [64], this reinforces the importance of CM pathology in ALS.The accumulating evidence for cortical involvement in the early stages of ALS [18,58] is consistent with what we know about the degenerative processes affecting motor cortex pyramidal neurons with long corticofugal projections, including Betz cells. Recent work has particularly highlighted the loss of dendritic spines and altered synaptic inputs to the apical dendrites of Betz cells in post-mortem brains of patients with different forms of ALS, but not in other neurodegenerative diseases [65]. Similar changes have been seen in layer V pyramidal neurons in the cortex of TDP-43 mouse models of ALS [66,67]. It is further suggested that transmission of the pathogen pTDP-43 by the long axons of motor cortex corticofugal neurons results, in turn, in the degeneration of neurons in their subcortical targets. In particular, direct CM projections could provide a monosynaptic route for pTDP-43 transmission from motor cortex, resulting in degeneration of α-motoneurons in the brainstem and spinal cord [18].
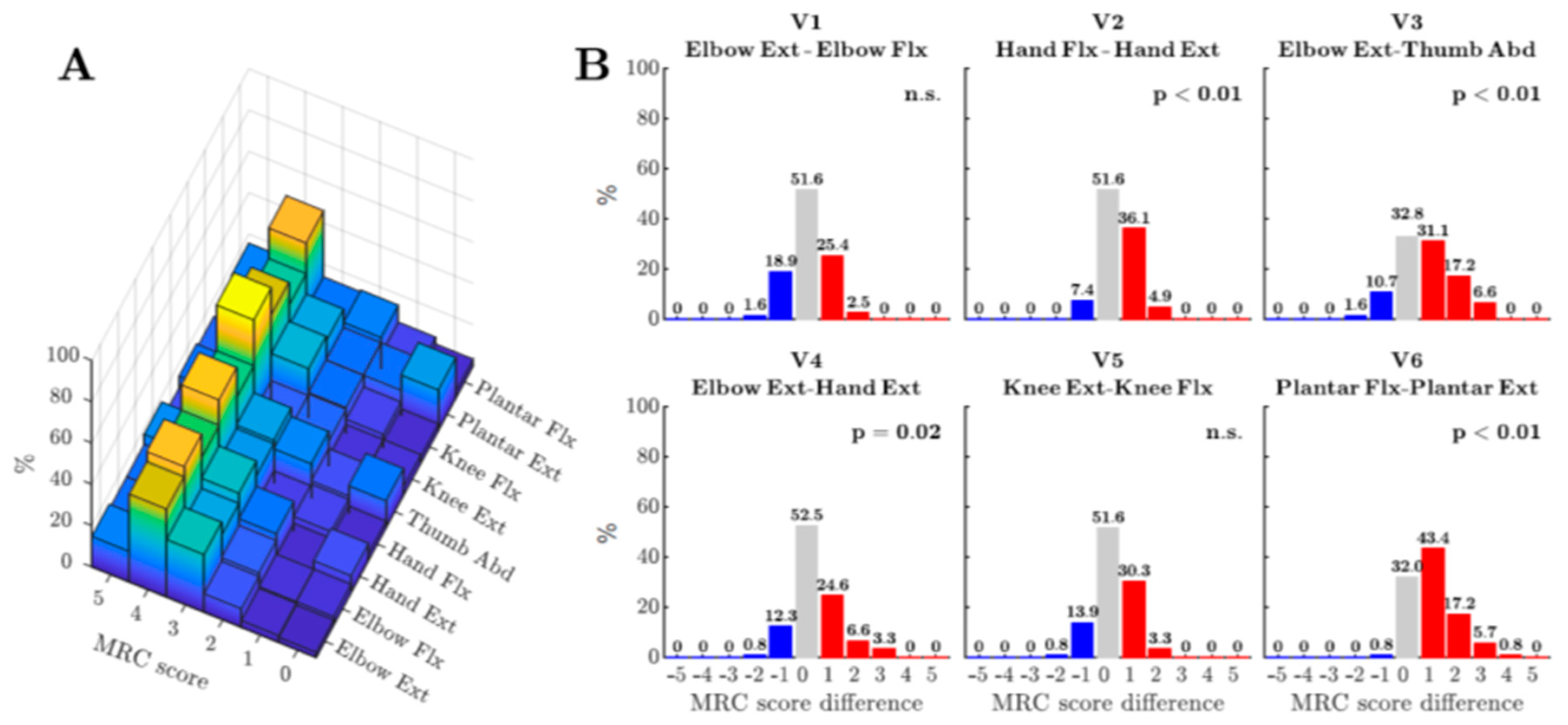
- Other recent studies have highlighted significant differences in the pattern of weakness in ALS patients which are consistent with the known connectivity of the CSP in humans and non-human primates [68,69]. An example is shown in Figure 1 from a recent study by Ludolph et al. [68], which included both retrospective and prospective studies of a large cohort of ALS patients whose muscle strength was assessed in different pairs of both upper and lower limb muscles, using the MRC scale. The study looked for any statistical sign of asymmetry, in each patient, between pairs of muscle groups, with one group known to receive strong CM connections and the other known to receive weaker connections. The pairs tested included wrist extensors vs. flexors, elbow flexors vs. extensors and, in the lower limb, knee flexors vs. extensors and plantar extensors vs. flexors. In the upper limb, strength was compared for a muscle group known to receive one of the strongest CM projections (thumb abductors) vs. one of the weakest (elbow extensors). The results showed a greater relative weakness for the muscle groups with normally receive strong CM projections, consistent with loss of CM influence during ALS [70]. These findings need following up, in a manner similar to that followed for the split-hand syndrome, to investigate the relative contribution of UMN and LMN to the weakness [59].
- A neurodegenerative process selectively targeting, in its early phase, pyramidal neurons with long corticofugal axons, including CM projections, may result in an UMN syndrome that is different from that caused, for example, by acute stroke or spinal injury to the UMN and its axon. Kinnear Wilson [71] famously questioned the suggestion that ALS could be considered as a disease of the “pyramidal system” because although ALS patients exhibited “awkwardness of finger movements”, they only suffered from slight weakness and there was no spasticity. However, all the recent studies discussed above actually point to a consistent pattern of weakness in the hands and radial digits of ALS patients. Moreover, since selective damage to the CST is well known to result in poverty of hand and finger movement without spasticity [72,73,74], there is no need to invoke higher order motor deficits to explain the symptoms observed in early ALS, although such deficits may well play a part in later stages of the disease.
- Progress in the understanding and treatment of ALS is dependent upon animal models. ALS appears to be a uniquely human disease and this has certainly made finding a good animal model difficult [18]. The involvement of the fast conducting CST and particularly of the CM system in ALS poses the question of whether a macaque model, whose motor system exhibits these same features, would allow additional insights to the progress already gained by the focus on rat and mouse studies [66,67,75], which do not share these features. Indeed, because in the rodent, corticospinal projections from sensorimotor cortex avoid the ventral horn and have limited direct effects on motor control [2,3], the layer V pyramidal neurons giving rise to these projections do not strictly qualify as ‘upper motor neurons’ in the sense that I have discussed in this short review. There are of course many features of motor cortex pyramidal neurons which are common to mouse, rat, macaque, and human [65,66,67,75], but we should also be aware that there are some striking differences, for example, in the shape and duration of their action potentials [12] and in some of the fast K+ ion channels involved in repolarisation [76]. These features would undoubtedly impact on the signals transmitted by the corticospinal tract and therefore on the different species-specific functions of this system.
10. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phillips, C.G.; Landau, W.M. Upper and lower motor neuron: The little old synecdoche that works. Neurology 1990, 40, 884–886. [Google Scholar] [CrossRef]
- Lemon, R.N. Descending pathways in motor control. Annu. Rev. Neurosci. 2008, 31, 195–218. [Google Scholar] [CrossRef]
- Lemon, R. Recent advances in our understanding of the primate corticospinal system. F1000Research 2019, 8, 274. [Google Scholar] [CrossRef]
- Baker, S.N. The primate reticulospinal tract, hand function and functional recovery. J. Physiol. 2011, 589, 5603–5612. [Google Scholar] [CrossRef]
- Dum, R.P.; Strick, P.L. The origin of corticospinal projections from the premotor areas in the frontal lobe. J. Neurosci. 1991, 11, 667–689. [Google Scholar] [CrossRef]
- Glickstein, M. Neuroscience: A Historical Introduction; MIT Press: Cambridge, MA, USA, 2014; p. 418. ISBN 0262026805. [Google Scholar]
- Kuypers, H.G.J.M. Anatomy of the Descending Pathways. In Handbook of Physiology the Nervous System II; The American Physiological Society: Rockville, MD, USA, 1981; pp. 597–666. [Google Scholar]
- Leenen, L.; Meek, J.; Posthuma, P.; Nieuwenhuys, R. A detailed morphometrical analysis of the pyramidal tract of the rat. Brain Res. 1985, 359, 65–80. [Google Scholar] [CrossRef]
- Firmin, L.; Field, P.; Maier, M.A.; Kraskov, A.; Kirkwood, P.A.; Nakajima, K.; Lemon, R.N.; Glickstein, M. Axon diameters and conduction velocities in the macaque pyramidal tract. J. Neurophysiol. 2014, 112, 1229–1240. [Google Scholar] [CrossRef]
- Innocenti, G.M.; Caminiti, R.; Rouiller, E.M.; Knott, G.; Dyrby, T.B.; Descoteaux, M.; Thiran, J.-P. Diversity of cortico-descending projections: Histological and diffusion MRI characterization in the monkey. Cereb. Cortex 2018, 29, 788–801. [Google Scholar] [CrossRef]
- Witham, C.L.; Fisher, K.M.; Edgley, S.A.; Baker, S.N. Corticospinal Inputs to Primate Motoneurons Innervating the Forelimb from Two Divisions of Primary Motor Cortex and Area 3a. J. Neurosci. 2016, 36, 2605–2616. [Google Scholar] [CrossRef]
- Vigneswaran, G.; Kraskov, A.; Lemon, R.N. Large Identified Pyramidal Cells in Macaque Motor and Premotor Cortex Exhibit “Thin Spikes”: Implications for Cell Type Classification. J. Neurosci. 2011, 31, 14235–14242. [Google Scholar] [CrossRef]
- Venkadesan, M.; Valero-Cuevas, F.J. Neural Control of Motion-to-Force Transitions with the Fingertip. J. Neurosci. 2008, 28, 1366–1373. [Google Scholar] [CrossRef]
- Kraskov, A.; Baker, S.; Soteropoulos, D.; Kirkwood, P.; Lemon, R. The Corticospinal Discrepancy: Where are all the Slow Pyramidal Tract Neurons? Cereb. Cortex 2019, 29, 3977–3981. [Google Scholar] [CrossRef]
- Kraskov, A.; Soteropoulos, D.S.; Glover, I.S.; Lemon, R.N.; Baker, S.N. Slowly-Conducting Pyramidal Tract Neurons in Macaque and Rat. Cereb. Cortex 2019, 30, 3403–3418. [Google Scholar] [CrossRef]
- Quencer, R.M.; Bunge, R.P.; Egnor, M.; Green, B.A.; Puckett, W.; Naidich, T.P.; Post, M.J.D.; Norenberg, M. Acute traumatic central cord syndrome: MRI-pathological correlations. Neuroradiology 1992, 34, 85–94. [Google Scholar] [CrossRef]
- Blight, A.R. Morphometric analysis of a model of spinal cord injury in guinea pigs, with behavioral evidence of delayed secondary pathology. J. Neurol. Sci. 1991, 103, 156–171. [Google Scholar] [CrossRef]
- Braak, H.; Brettschneider, J.; Ludolph, A.C.; Lee, V.M.; Trojanowski, J.Q.; Del Tredici, K. Amyotrophic lateral sclerosis—a model of corticofugal axonal spread. Nat. Rev. Neurol. 2013, 9, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Eisen, A.; Stewart, H.G.; Andersen, P.M. Preserved slow conducting corticomotoneuronal projections in amyotrophic lateral sclerosis with autosomal recessive D90A CuZn-superoxide dismutase mutation. Brain 2000, 123, 1505–1515. [Google Scholar] [CrossRef]
- Maier, M.; Armand, J.; Kirkwood, P.; Yang, H.-W.; Davis, J.; Lemon, R. Differences in the Corticospinal Projection from Primary Motor Cortex and Supplementary Motor Area to Macaque Upper Limb Motoneurons: An Anatomical and Electrophysiological Study. Cereb. Cortex 2002, 12, 281–296. [Google Scholar] [CrossRef]
- Morecraft, R.J.; Ge, J.; Stilwell-Morecraft, K.S.; McNeal, D.W.; Pizzimenti, M.A.; Darling, W.G. Terminal distribution of the corticospinal projection from the hand/arm region of the primary motor cortex to the cervical enlargement in rhesus monkey. J. Comp. Neurol. 2013, 521, 4205–4235. [Google Scholar] [CrossRef]
- Morecraft, R.J.; Ge, J.; Stilwell-Morecraft, K.S.; Rotella, D.L.; Pizzimenti, M.A.; Darling, W.G. Terminal organization of the corticospinal projection from the lateral premotor cortex to the cervical enlargement (C5–T1) in rhesus monkey. J. Comp. Neurol. 2019, 527, 2761–2789. [Google Scholar] [CrossRef] [PubMed]
- Ralston, D.D.; Ralston, H.J. The terminations of corticospinal tract axons in the macaque monkey. J. Comp. Neurol. 1985, 242, 325–337. [Google Scholar] [CrossRef]
- Fisher, K.M.; Lilak, A.; Garner, J.; Darian-Smith, C. Extensive somatosensory and motor corticospinal sprouting occurs following a central dorsal column lesion in monkeys. J. Comp. Neurol. 2018, 526, 2373–2387. [Google Scholar] [CrossRef]
- Lacroix, S.; Havton, L.A.; McKay, H.; Yang, H.; Brant, A.; Roberts, J.; Tuszynski, M.H. Bilateral corticospinal projections arise from each motor cortex in the macaque monkey: A quantitative study. J. Comp. Neurol. 2004, 473, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Morecraft, R.J.; McNeal, D.W.; Dvanajscak, Z.; Ge, J.; Schneider, P.; Stilwell-Morecraft, K.S. Localization of arm representation in the cerebral peduncle of the non-human primate. J. Comp. Neurol. 2007, 504, 149–167. [Google Scholar] [CrossRef]
- Foerster, O. Symptomatologie der Erkrankungen des Ruckenmarks. In Handbook of Neurology; Bumke, O., Foerster, O., Eds.; Springer: Berlin, Germany, 1936; Volume 5, pp. 80–83. [Google Scholar]
- Levi, A.D.; Tator, C.H.; Bunge, R.P. Clinical Syndromes Associated with Disproportionate Weakness of the Upper versus the Lower Extremities after Cervical Spinal Cord Injury. Neurosurgery 1996, 38, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Morecraft, R.J.; Stilwell-Morecraft, K.S.; Ge, J.; Kraskov, A.; Lemon, R.N. Lack of somatotopy among corticospinal tract fibers passing through the primate craniovertebral junction and cervical spinal cord: Pathoanatomic substrate of central cord syn-drome and cruciate paralysis. J. Neurosurg. (in press).
- Rathelot, J.-A.; Strick, P.L. Subdivisions of primary motor cortex based on cortico-motoneuronal cells. Proc. Natl. Acad. Sci. USA 2009, 106, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Geyer, S.; Ledberg, A.; Schleicher, A.; Kinomura, S.; Schormann, T.; Bürgel, U.; Klingberg, T.; Larsson, J.; Zilles, K.; Roland, P.E. Two different areas within the primary motor cortex of man. Nat. Cell Biol. 1996, 382, 805–807. [Google Scholar] [CrossRef] [PubMed]
- Rathelot, J.-A.; Strick, P.L. Muscle representation in the macaque motor cortex: An anatomical perspective. Proc. Natl. Acad. Sci. USA 2006, 103, 8257–8262. [Google Scholar] [CrossRef]
- Porter, R.; Lemon, R. Corticospinal Function and Voluntary Movement; Oxford University Press: Oxford, UK, 1993; ISBN 9780198523758. [Google Scholar]
- Schoen, J. Comparative Aspects of the Descending Fibre Systems in the Spinal Cord. Prog. Brain Res. 1964, 11, 203–222. [Google Scholar]
- Iwatsubo, T.; Kuzuhara, S.; Kanemitsu, A.; Shimada, H.; Toyokura, Y. Corticofugal projections to the motor nuclei of the brainstem and spinal cord in humans. Neurology 1990, 40, 309. [Google Scholar] [CrossRef]
- Rothwell, J.C.; Thompson, P.D.; Day, B.L.; Boyd, S.; Marsden, C.D. Stimulation of the human motor cortex through the scalp. Exp. Physiol. 1991, 76, 159–200. [Google Scholar] [CrossRef] [PubMed]
- De Noordhout, A.M.; Rapisarda, G.; Bogacz, D.; Gérard, P.; De Pasqua, V.; Pennisi, G.; Delwaide, P.J. Corticomotoneuronal synaptic connections in normal man: An electrophysiological study. Brain 1999, 122, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Palmer, E.; Ashby, P. Corticospinal projections to upper limb motoneurones in humans. J. Physiol. 1992, 448, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, B.; Ashby, P. Corticospinal projections to upper and lower limb spinal motoneurons in man. Electroencephalogr. Clin. Neurophysiol. 1990, 76, 509–519. [Google Scholar] [CrossRef]
- Colebatch, J.G.; Rothwell, J.C.; Day, B.L.; Thompson, P.D.; Marsden, C.D. Cortical outflow to proximal arm muscles in man. Brain 1990, 113, 1843–1856. [Google Scholar] [CrossRef]
- Fornia, L.; Ferpozzi, V.; Montagna, M.; Rossi, M.; Riva, M.; Pessina, F.; Boneschi, F.M.; Borroni, P.; Lemon, R.N.; Bello, L.; et al. Functional Characterization of the Left Ventrolateral Premotor Cortex in Humans: A Direct Electrophysiological Approach. Cereb. Cortex 2016, 28, 167–183. [Google Scholar] [CrossRef]
- Brouwer, B.; Ashby, P. Corticospinal projections to lower limb motoneurons in man. Exp. Brain Res. 1992, 89, 649–654. [Google Scholar] [CrossRef]
- Bawa, P.; Chalmers, G.R.; Stewart, H.; Eisen, A.A. Responses of ankle extensor and flexor motoneurons to transcranial magnetic stimulation. J. Neurophysiol. 2002, 88, 124–132. [Google Scholar] [CrossRef]
- Fetz, E.E.; Cheney, P.D. Postspike facilitation of forelimb muscle activity by primate corticomotoneuronal cells. J. Neurophysiol. 1980, 44, 751–772. [Google Scholar] [CrossRef]
- Buys, E.J.; Lemon, R.N.; Mantel, G.W.; Muir, R.B. Selective facilitation of different hand muscles by single corticospinal neurones in the conscious monkey. J. Physiol. 1986, 381, 529–549. [Google Scholar] [CrossRef] [PubMed]
- McKiernan, B.J.; Marcario, J.K.; Karrer, J.H.; Cheney, P.D. Corticomotoneuronal Postspike Effects in Shoulder, Elbow, Wrist, Digit, and Intrinsic Hand Muscles During a Reach and Prehension Task. J. Neurophysiol. 1998, 80, 1961–1980. [Google Scholar] [CrossRef] [PubMed]
- Muir, R.; Lemon, R. Corticospinal neurons with a special role in precision grip. Brain Res. 1983, 261, 312–316. [Google Scholar] [CrossRef]
- Griffin, D.M.; Hudson, H.M.; Belhaj-Saïf, A.; McKiernan, B.J.; Cheney, P.D. Do Corticomotoneuronal Cells Predict Target Muscle EMG Activity? J. Neurophysiol. 2008, 99, 1169–1986. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Griffin, D.M.; Hoffman, D.S.; Strick, P.L. Corticomotoneuronal cells are “functionally tuned”. Science 2015, 350, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Fetz, E.E.; Finocchio, D.V. Operant Conditioning of Specific Patterns of Neural and Muscular Activity. Science 1971, 174, 431–435. [Google Scholar] [CrossRef]
- Fetz, E.E.; Finocchio, D.V. Correlations between activity of motor cortex cells and arm muscles during operantly conditioned response patterns. Exp. Brain Res. 1975, 23, 217–240. [Google Scholar] [CrossRef]
- Schieber, M.H. Dissociating motor cortex from the motor. J. Physiol. 2011, 589, 5613–5624. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.G. Evolution of the corticospinal tract in primates with special reference to the hand. In Proceedings of the Third International Congress of Primatology, Zurich, Switzerland, 2–5 August 1970; Karger Publishers: Basel, Switzerland, 1971; Volume 2, pp. 2–23. [Google Scholar]
- Bortoff, G.A.; Strick, P.L. Corticospinal terminations in two new-world primates: Further evidence that corticomotoneuronal connections provide part of the neural substrate for manual dexterity. J. Neurosci. 1993, 13, 5105–5118. [Google Scholar] [CrossRef] [PubMed]
- Quallo, M.M.; Kraskov, A.; Lemon, R.N. The Activity of Primary Motor Cortex Corticospinal Neurons during Tool Use by Macaque Monkeys. J. Neurosci. 2012, 32, 17351–17364. [Google Scholar] [CrossRef] [PubMed]
- Eisen, A. The Dying Forward Hypothesis of ALS: Tracing Its History. Brain Sci. 2021, 11, 300. [Google Scholar] [CrossRef]
- Eisen, A.; Kim, S.; Pant, B. Amyotrophic lateral sclerosis (ALS): A phylogenetic disease of the corticomotoneuron? Muscle Nerve 1992, 15, 219–224. [Google Scholar] [CrossRef]
- Eisen, A.; Braak, H.; Del Tredici, K.; Lemon, R.; Ludolph, A.C.; Kiernan, M.C. Cortical influences drive amyotrophic lateral sclerosis. J. Neurol. Neurosurg. Psychiatry 2017, 88, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.D.; Eisen, A. ALS Split Phenotypes—To what extent do they exist? Clin. Neurophysiol. 2020, 131, 847–849. [Google Scholar] [CrossRef]
- Eisen, A.; Turner, M.R.; Lemon, R. Tools and talk: An evolutionary perspective on the functional deficits associated with amyotrophic lateral sclerosis. Muscle Nerve 2013, 49, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.H. Croonian Lectures on the Evolution and Dissolution of the Nervous System. Br. Med. J. 1884, 1, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Eisen, A.; Stewart, H.; Hirota, N. The split hand in ALS has a cortical basis. J. Neurol. Sci. 2000, 180, 66–70. [Google Scholar] [CrossRef]
- Clough, J.F.M.; Kernell, D.; Phillips, C.G. The distribution of monosynaptic excitation from the pyramidal tract and from primary spindle afferents to motoneurones of the baboon’s hand and forearm. J. Physiol. 1968, 198, 145–166. [Google Scholar] [CrossRef]
- Menon, P.; Kiernan, M.C.; Vucic, S. ALS pathophysiology: Insights from the split-hand phenomenon. Clin. Neurophysiol. 2014, 125, 186–193. [Google Scholar] [CrossRef]
- Genç, B.; Jara, J.H.; Lagrimas, A.K.B.; Pytel, P.; Roos, R.P.; Mesulam, M.M.; Geula, C.; Bigio, E.H.; Özdinler, P.H. Apical dendrite degeneration, a novel cellular pathology for Betz cells in ALS. Sci. Rep. 2017, 7, srep41765. [Google Scholar] [CrossRef]
- Fogarty, M.J.; Klenowski, P.M.; Lee, J.D.; Drieberg-Thompson, J.R.; Bartlett, S.E.; Ngo, S.T.; Hilliard, M.A.; Bellingham, M.C.; Noakes, P.G. Cortical synaptic and dendritic spine abnormalities in a presymptomatic TDP-43 model of amyotrophic lateral sclerosis. Sci. Rep. 2016, 6, 37968. [Google Scholar] [CrossRef] [PubMed]
- Gautam, M.; Jara, J.H.; Kocak, N.; Rylaarsdam, L.E.; Kim, K.D.; Bigio, E.H.; Özdinler, P.H. Mitochondria, ER, and nuclear membrane defects reveal early mechanisms for upper motor neuron vulnerability with respect to TDP-43 pathology. Acta Neuropathol. 2019, 137, 47–69. [Google Scholar] [CrossRef]
- Ludolph, A.C.; Emilian, S.; Dreyhaupt, J.; Rosenbohm, A.; Kraskov, A.; Lemon, R.N.; Del Tredici, K.; Braak, H. Pattern of paresis in ALS is consistent with the physiology of the corticomotoneuronal projections to different muscle groups. J. Neurol. Neurosurg. Psychiatry 2020, 91, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, R.; Martin, S.; Ellis, C.; Burman, R.; Sreedharan, J.; Shaw, C.; Leigh, P.N.; Turner, M.R.; Al-Chalabi, A. Relative preservation of triceps over biceps strength in upper limb-onset ALS: The ‘split elbow’. J. Neurol. Neurosurg. Psychiatry 2019, 90, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.R. ALS—Dying forward, backward or outward? Nat. Rev. Neurol. 2014, 10, 660. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.A.K. Amyotrophic lateral sclerosis. In Neurology; Edward Arnold: London, UK, 1940; pp. 1007–1032. [Google Scholar]
- Lawrence, D.G.; Kuypers, H.G.J.M. The functional organization of the motor system in the monkey. Brain 1968, 91, 1–14. [Google Scholar] [CrossRef]
- Lemon, R.N.; Landau, W.; Tutssel, D.; Lawrence, D.G. Lawrence and Kuypers (1968a, b) revisited: Copies of the original filmed material from their classic papers in Brain. Brain 2012, 135, 2290–2295. [Google Scholar] [CrossRef] [PubMed]
- Tower, S.S. Pyramidal lesion in the monkey. Brain 1940, 63, 36–90. [Google Scholar] [CrossRef]
- Genc, B.; Gozutok, O.; Ozdinler, P.H. Complexity of generating mouse models to study the upper motor neurons: Let us shift focus from mice to neurons. Int. J. Mol. Sci. 2019, 20, 3848. [Google Scholar] [CrossRef] [PubMed]
- Soares, D.; Goldrick, I.; Lemon, R.N.; Kraskov, A.; Greensmith, L.; Kalmar, B. Expression of Kv3.1b potassium channel is widespread in macaque motor cortex pyramidal cells: A histological comparison between rat and macaque. J. Comp. Neurol. 2017, 525, 2164–2174. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemon, R.N. The Cortical “Upper Motoneuron” in Health and Disease. Brain Sci. 2021, 11, 619. https://doi.org/10.3390/brainsci11050619
Lemon RN. The Cortical “Upper Motoneuron” in Health and Disease. Brain Sciences. 2021; 11(5):619. https://doi.org/10.3390/brainsci11050619
Chicago/Turabian StyleLemon, Roger N. 2021. "The Cortical “Upper Motoneuron” in Health and Disease" Brain Sciences 11, no. 5: 619. https://doi.org/10.3390/brainsci11050619
APA StyleLemon, R. N. (2021). The Cortical “Upper Motoneuron” in Health and Disease. Brain Sciences, 11(5), 619. https://doi.org/10.3390/brainsci11050619