
Enhanced White Matter Fiber Tracts in Advanced Jazz Improvisers



Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Behavioral Tests and MRI Scanning
2.3. DWI Data Acuisition and Image Processing
2.4. Data Analysis
Diffusion Weighted Imaging Data Analysis
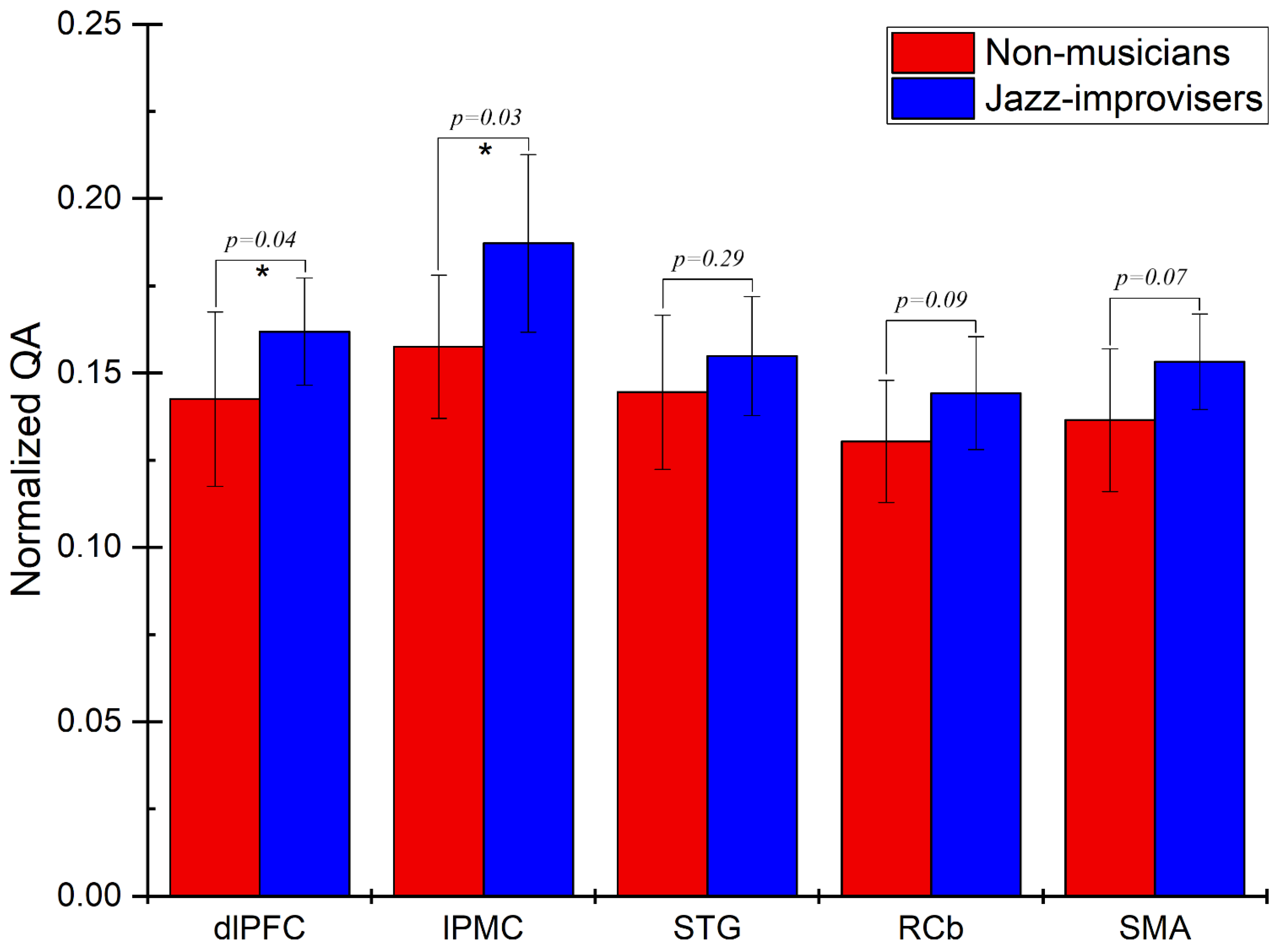
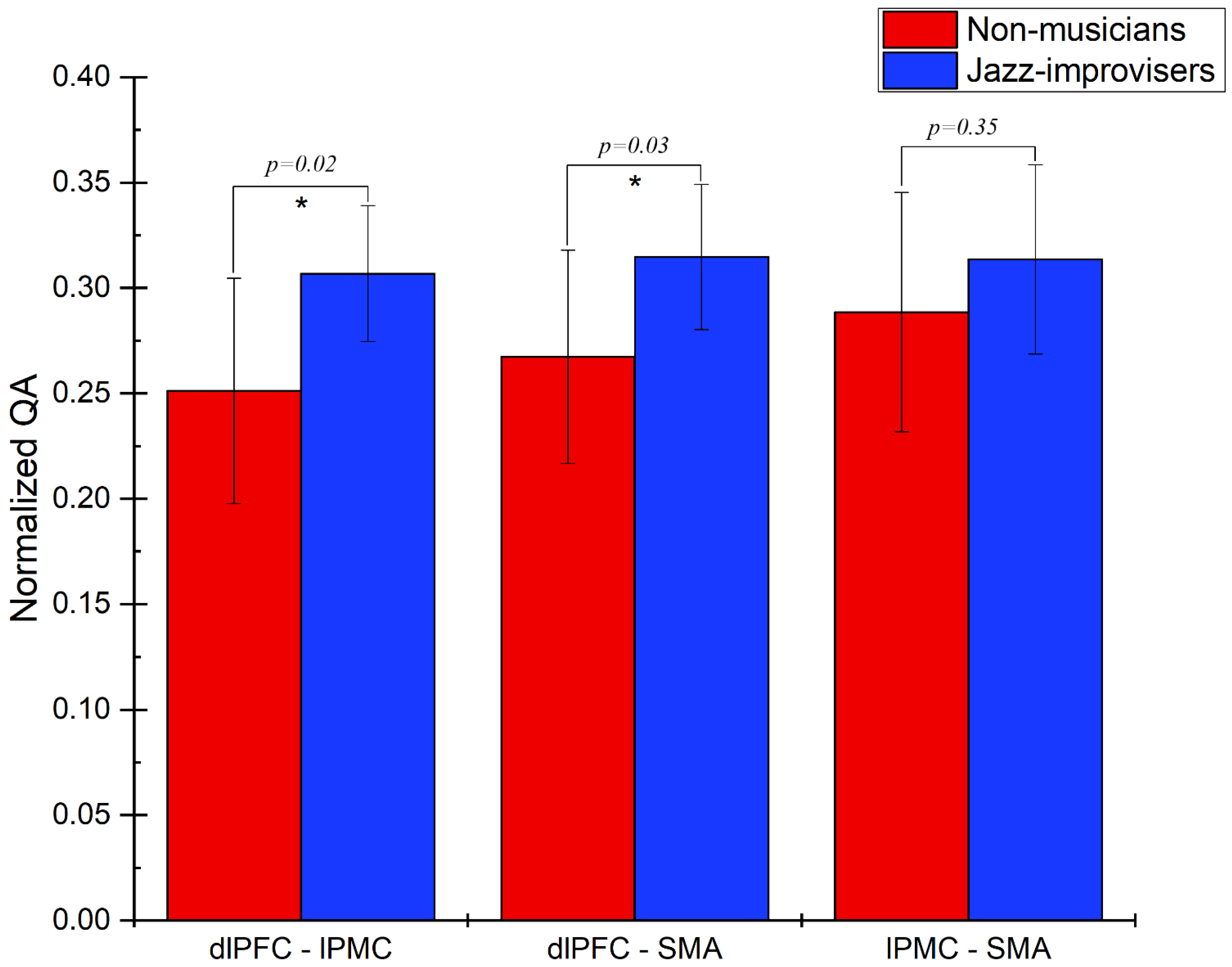
3. Results
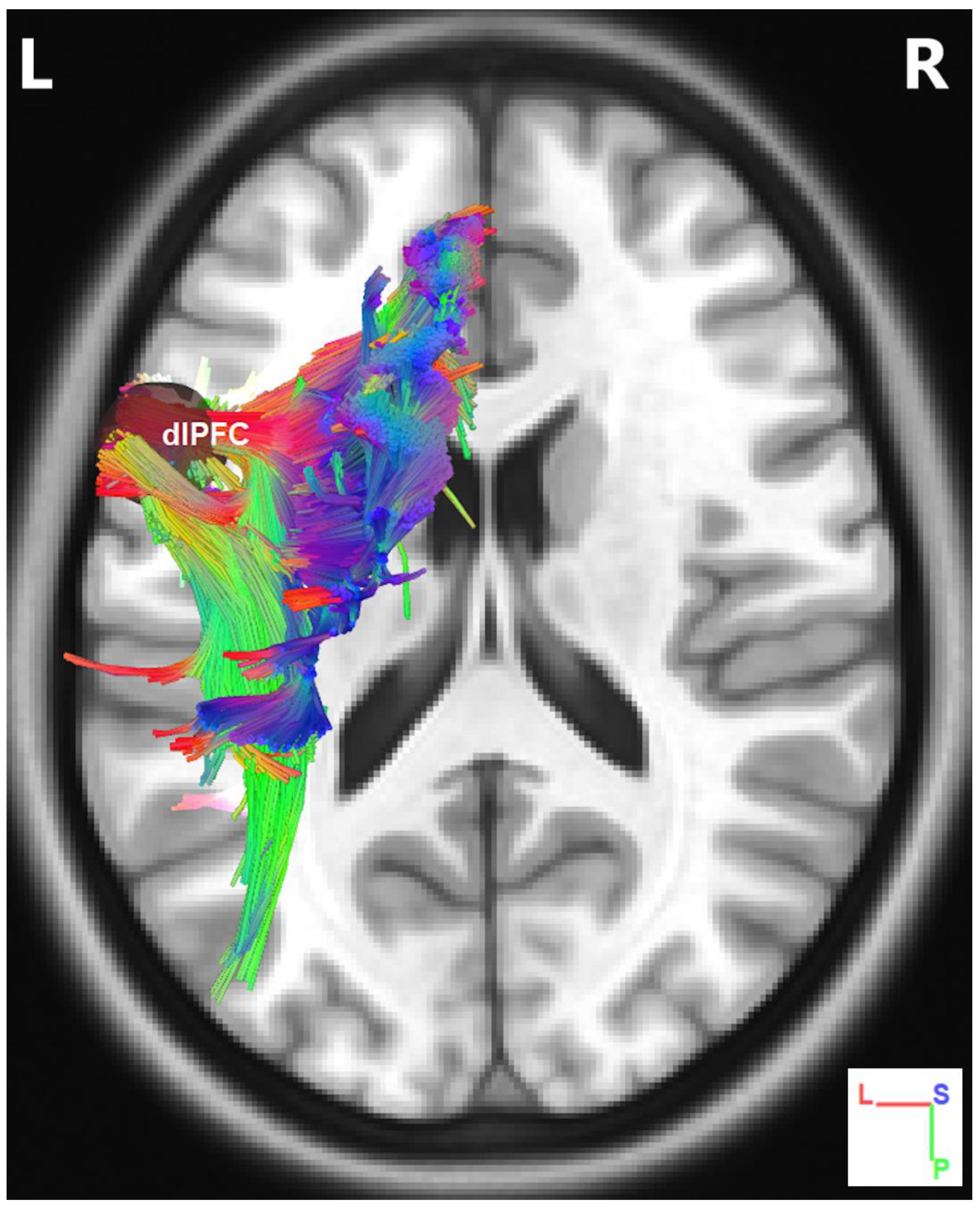
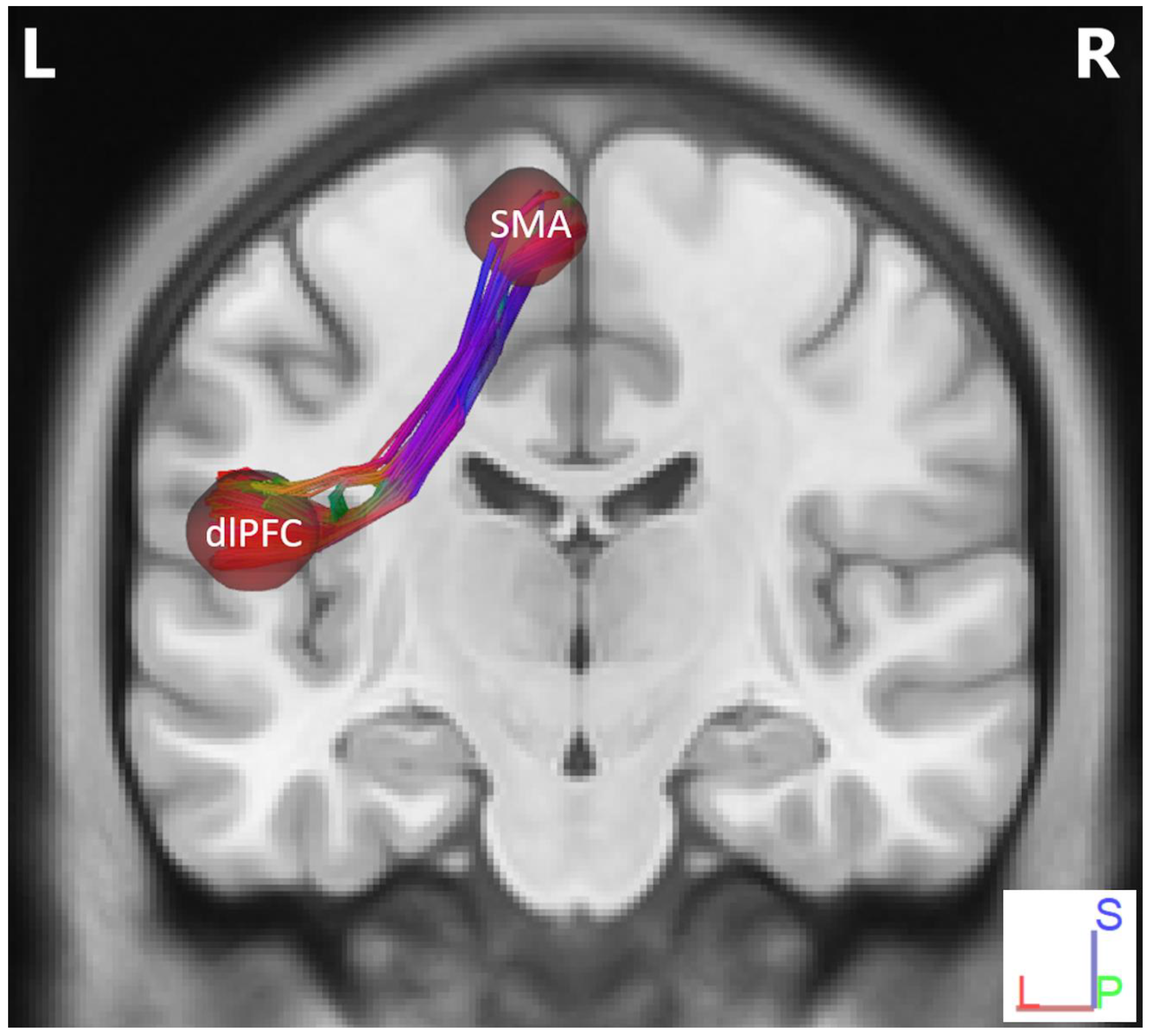
Track-Specific and Region-Based Fiber Tractography Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faber, S.E.M.; McIntosh, A.R. Towards a standard model of musical improvisation. Eur. J. Neurosci. 2020, 51, 840–849. [Google Scholar] [CrossRef]
- Beaty, R.E.; Benedek, M.; Silvia, P.J.; Schacter, D.L. Creative Cognition and Brain Network Dynamics. Trends Cogn. Sci. 2016, 20, 87–95. [Google Scholar] [CrossRef]
- Chen, Q.; Beaty, R.E.; Qiu, J. Mapping the artistic brain: Common and distinct neural activations associated with musical, drawing, and literary creativity. Hum. Brain Mapp. 2020, 41, 3403–3419. [Google Scholar] [CrossRef]
- Dhakal, K.; Norgaard, M.; Adhikari, B.M.; Yun, K.S.; Dhamala, M. Higher Node Activity with Less Functional Connectivity During Musical Improvisation. Brain Connect. 2019, 9, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Norgaard, M. Descriptions of improvisational thinking by artist-level jazz musicians. J. Res. Music Educ. 2011, 59, 109–127. [Google Scholar] [CrossRef]
- Dietrich, A. The cognitive neuroscience of creativity. Psychon. Bull. Rev. 2004, 11, 1011–1026. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.; Schaefer, R.S.; Bastin, M.E.; Roberts, N.; Overy, K. Can musical training influence brain connectivity? Evidence from diffusion tensor MRI. Brain Sci. 2014, 4, 405–427. [Google Scholar] [CrossRef] [PubMed]
- Arkin, C.; Przysinda, E.; Pfeifer, C.W.; Zeng, T.; Loui, P. Gray Matter Correlates of Creativity in Musical Improvisation. Front. Hum. Neurosci. 2019, 13, 169. [Google Scholar] [CrossRef] [PubMed]
- Zamm, A.; Schlaug, G.; Eagleman, D.M.; Loui, P. Pathways to seeing music: Enhanced structural connectivity in colored-music synesthesia. Neuroimage 2013, 74, 359–366. [Google Scholar] [CrossRef]
- Loui, P.; Li, H.C.; Schlaug, G. White matter integrity in right hemisphere predicts pitch-related grammar learning. Neuroimage 2011, 55, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Gaser, C.; Schlaug, G. Brain Structures Differ between Musicians and Non-Musicians. J. Neurosci. 2003, 23, 9240–9245. [Google Scholar] [CrossRef]
- Basser, P.J.; Mattiello, J.; Lebihan, D. Estimation of the Effective Self-Diffusion Tensor from the NMR Spin Echo. J. Magn. Reson. Ser. B 1994, 103, 247–254. [Google Scholar] [CrossRef]
- Pierpaoli, C.; Jezzard, P.; Basser, P.J.; Barnett, A.; Chiro, G.D. Diffusion tensor MR imaging of the human brain. Radiology 1996, 201, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.E.; Anderson, E.J.; Husain, M. White matter microstructure and cognitive function. Neuroscientist 2013, 19, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Scholz, J.; Tomassini, V.; Johansen-Berg, H. Individual Differences in White Matter Microstructure in the Healthy Brain. In Diffusion MRI, 2nd ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 301–316. [Google Scholar] [CrossRef]
- Yeh, F.C.; Tseng, W.Y. NTU-90: A high angular resolution brain atlas constructed by q-space diffeomorphic reconstruction. Neuroimage 2011, 58, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Wedeen, V.J.; Tseng, W.Y. Generalized q-sampling imaging. IEEE Trans. Med. Imaging 2010, 29, 1626–1635. [Google Scholar] [CrossRef]
- Yeh, F.C.; Verstynen, T.D.; Wang, Y.; Fernandez-Miranda, J.C.; Tseng, W.Y. Deterministic diffusion fiber tracking improved by quantitative anisotropy. PLoS ONE 2013, 8, e80713. [Google Scholar] [CrossRef]
- Tuch, D.S. Q-ball imaging. Magn. Reson. Med. 2004, 52, 1358–1372. [Google Scholar] [CrossRef] [PubMed]
- Fritzsche, K.H.; Laun, F.B.; Meinzer, H.-P.; Stieltjes, B. Opportunities and pitfalls in the quantification of fiber integrity: What can we gain from Q-ball imaging? NeuroImage 2010, 51, 242–251. [Google Scholar] [CrossRef]
- Yeh, F.C.; Zaydan, I.M.; Suski, V.R.; Lacomis, D.; Richardson, R.M.; Maroon, J.C.; Barrios-Martinez, J. Differential tractography as a track-based biomarker for neuronal injury. NeuroImage 2019, 202, 116131. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.-C. Shape analysis of the human association pathways. NeuroImage 2020, 223, 117329. [Google Scholar] [CrossRef]
- Yeh, F.C.; Panesar, S.; Barrios, J.; Fernandes, D.; Abhinav, K.; Meola, A.; Fernandez-Miranda, J.C. Automatic Removal of False Connections in Diffusion MRI Tractography Using Topology-Informed Pruning (TIP). Neurotherapeutics 2019, 16, 52–58. [Google Scholar] [CrossRef]
- Dhamala, M.; Rangarajan, G.; Ding, M. Analyzing information flow in brain networks with nonparametric Granger causality. Neuroimage 2008, 41, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Schmithorst, V.J.; Wilke, M. Differences in white matter architecture between musicians and non-musicians: A diffusion tensor imaging study. Neurosci. Lett. 2002, 321, 57–60. [Google Scholar] [CrossRef]
- Bengtsson, S.L.; Nagy, Z.; Skare, S.; Forsman, L.; Forssberg, H.; Ullen, F. Extensive piano practicing has regionally specific effects on white matter development. Nat. Neurosci. 2005, 8, 1148–1150. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Yang, H.; Lv, Y.T.; Zhu, C.Z.; He, Y.; Tang, H.H.; Gong, Q.Y.; Luo, Y.J.; Zang, Y.F.; Dong, Q. Gray matter density and white matter integrity in pianists’ brain: A combined structural and diffusion tensor MRI study. Neurosci. Lett. 2009, 459, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Halwani, G.F.; Loui, P.; Ruber, T.; Schlaug, G. Effects of practice and experience on the arcuate fasciculus: Comparing singers, instrumentalists, and non-musicians. Front. Psychol. 2011, 2, 156. [Google Scholar] [CrossRef]
- Imfeld, A.; Oechslin, M.S.; Meyer, M.; Loenneker, T.; Jancke, L. White matter plasticity in the corticospinal tract of musicians: A diffusion tensor imaging study. Neuroimage 2009, 46, 600–607. [Google Scholar] [CrossRef]
- Pinho, A.L.; Ullen, F.; Castelo-Branco, M.; Fransson, P.; de Manzano, O. Addressing a Paradox: Dual Strategies for Creative Performance in Introspective and Extrospective Networks. Cereb. Cortex 2016, 26, 3052–3063. [Google Scholar] [CrossRef]
- Pressing, J. Improvisation: Methods and models. In Generative Processes in Music; Sloboda, J.A., Ed.; Oxford University Press: Oxford, UK, 1987; pp. 129–178. [Google Scholar]
- De Manzano, Ö.; Ullén, F. Activation and connectivity patterns of the presupplementary and dorsal premotor areas during free improvisation of melodies and rhythms. NeuroImage 2012, 63, 272–280. [Google Scholar] [CrossRef]
- Beaty, R.E. The neuroscience of musical improvisation. Neurosci. Biobehav. Rev. 2015, 51, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Berkowitz, A.L.; Ansari, D. Generation of novel motor sequences: The neural correlates of musical improvisation. Neuroimage 2008, 41, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Koechlin, E.; Jubault, T. Broca’s Area and the Hierarchical Organization of Human Behavior. Neuron 2006, 50, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Wertz, C.J.; Chohan, M.O.; Ramey, S.J.; Flores, R.A.; Jung, R.E. White matter correlates of creative cognition in a normal cohort. Neuroimage 2020, 208, 116293. [Google Scholar] [CrossRef]
- Takeuchi, H.; Sekiguchi, A.; Taki, Y.; Yokoyama, S.; Yomogida, Y.; Komuro, N.; Yamanouchi, T.; Suzuki, S.; Kawashima, R. Training of working memory impacts structural connectivity. J. Neurosci. 2010, 30, 3297–3303. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant No. | Age (Years) | Years of Experience (Jazz Improvisation) | Primary Instrument |
|---|---|---|---|
| 01 | 31 | 24 | Piano |
| 02 | 57 | 50 | Piano |
| 03 | 41 | 31 | Saxophone |
| 04 | 43 | 34 | Piano |
| 05 | 33 | 22 | Piano |
| 06 | 20 | 6 | Guitar |
| 07 | 22 | 10 | Saxophone |
| 08 | 26 | 15 | Saxophone |
| 09 | 41 | 33 | Saxophone |
| 10 | 23 | 11 | Saxophone |
| 11 | 19 | 10 | Saxophone |
| 12 | 26 | 18 | Piano |
| 13 | 30 | 18 | Contra/Double Bass |
| 14 | 21 | 12 | Trombone |
| 15 | 28 | 14 | Drum Set |
| 16 | 23 | 14 | Saxophone |
| 17 | 23 | 12 | Saxophone |
| 18 | 42 | 33 | Trumpet |
| 19 | 38 | 28 | Saxophone |
| 20 | 22 | 11 | Trumpet |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhakal, K.; Norgaard, M.; Dhamala, M. Enhanced White Matter Fiber Tracts in Advanced Jazz Improvisers. Brain Sci. 2021, 11, 506. https://doi.org/10.3390/brainsci11040506
Dhakal K, Norgaard M, Dhamala M. Enhanced White Matter Fiber Tracts in Advanced Jazz Improvisers. Brain Sciences. 2021; 11(4):506. https://doi.org/10.3390/brainsci11040506
Chicago/Turabian StyleDhakal, Kiran, Martin Norgaard, and Mukesh Dhamala. 2021. "Enhanced White Matter Fiber Tracts in Advanced Jazz Improvisers" Brain Sciences 11, no. 4: 506. https://doi.org/10.3390/brainsci11040506
APA StyleDhakal, K., Norgaard, M., & Dhamala, M. (2021). Enhanced White Matter Fiber Tracts in Advanced Jazz Improvisers. Brain Sciences, 11(4), 506. https://doi.org/10.3390/brainsci11040506