
A Bridge between the Breath and the Brain: Synchronization of Respiration, a Pupillometric Marker of the Locus Coeruleus, and an EEG Marker of Attentional Control State

 ,
,
Abstract
1. Introduction
2. Methods
2.1. Participants
2.2. Data Recording
2.3. Data Preparation
2.4. Data Analysis
3. Results
3.1. Phase Synchronization Analysis
3.2. Multivariate Granger-Causality Analysis
4. Discussion
5. Study Limitations
6. Conclusions
7. Future Research Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adrian, E. The electrical activity of the mammalian olfactory bulb. Electroencephalogr. Clin. Neurophysiol. 1950, 2, 377–388. [Google Scholar] [CrossRef]
- Alnaes, D.; Sneve, M.H.; Espeseth, T.; Endestad, T.; Van De Pavert, S.H.P.; Laeng, B. Pupil size signals mental effort deployed during multiple object tracking and predicts brain activity in the dorsal attention network and the locus coeruleus. J. Vis. 2014, 14, 1. [Google Scholar] [CrossRef]
- Angelidis, A.; Hagenaars, M.; van Son, D.; van der Does, W.; Putman, P. Do not look away! Spontaneous frontal EEG theta/beta ratio as a marker for cognitive control over attention to mild and high threat. Biol. Psychol. 2018, 135, 8–17. [Google Scholar] [CrossRef]
- Muktibhodananda, S. Hatha Yoga Pradapika, 3rd ed.; Bihar School of Yoga Publishers: Bihar, India, 2013. [Google Scholar]
- Sai, V. Ananda Sutra; Sri Vasantha Sai Books and Publications Trust: Andhra Pradesh, India, 2010. [Google Scholar]
- Satchidananda, S.S. The Yoga Sutras of Patanjali: Reprint Edition; Integral Yoga Publications: Buckingham, VA, USA, 2012. [Google Scholar]
- Biskamp, J.; Bartos, M.; Sauer, J.-F. Organization of prefrontal network activity by respiration-related oscillations. Sci. Rep. 2017, 7, srep45508. [Google Scholar] [CrossRef]
- Hobson, J.A. Respiration and EEG Synchronization in the Frog. Nature 1967, 213, 988–989. [Google Scholar] [CrossRef]
- Melnychuk, M.C.; Dockree, P.M.; O’Connell, R.G.; Murphy, P.R.; Balsters, J.H.; Robertson, I.H. Coupling of respiration and attention via the locus coeruleus: Effects of meditation and pranayama. Psychophysiology 2018, 55, e13091. [Google Scholar] [CrossRef] [PubMed]
- Stankovski, T.; Petkoski, S.; Raeder, J.; Smith, A.F.; McClintock, P.; Stefanovska, A. Alterations in the coupling functions between cortical and cardio-respiratory oscillations due to anaesthesia with propofol and sevoflurane. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150186. [Google Scholar] [CrossRef] [PubMed]
- Tort, A.B.; Brankačk, J.; Draguhn, A. Respiration-Entrained Brain Rhythms Are Global but Often Overlooked. Trends Neurosci. 2018, 41, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Yackle, K.; Schwarz, L.A.; Kam, K.; Sorokin, J.M.; Huguenard, J.R.; Feldman, J.L.; Luo, L.; Krasnow, M.A. Breathing control center neurons that promote arousal in mice. Science 2017, 355, 1411–1415. [Google Scholar] [CrossRef]
- Zelano, C.; Jiang, H.; Zhou, G.; Arora, N.; Schuele, S.; Rosenow, J.; Gottfried, J.A. Nasal Respiration Entrains Human Limbic Oscillations and Modulates Cognitive Function. J. Neurosci. 2016, 36, 12448–12467. [Google Scholar] [CrossRef]
- Gellhorn, E.; Kraines, S.H. The influence of hyperpnea and of variations in the o2-and co2-tension of the inspired air on word-associations. Science 1936, 83, 266–267. [Google Scholar] [CrossRef]
- Lehmann, A. Revue Philosophique de la France Et de l’Etranger. 1893. Available online: https://gallica.bnf.fr/ark:/12148/cb34349223n/date (accessed on 31 August 2021).
- Macdougall, R.; Munsterberg, H. Studies from the Harvard Psychological Laboratory (IV): The physical characteristics of attention. Psychol. Rev. 1896, 3, 158–180. [Google Scholar] [CrossRef][Green Version]
- Porges, S.W.; Raskin, D.C. Respiratory and heart rate components of attention. J. Exp. Psychol. 1969, 81, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.W. The Effect of Certain Stimuli upon the Attention Wave. Am. J. Psychol. 1901, 12, 335. [Google Scholar] [CrossRef]
- Winkler, C. Attention and respiration. K. Ned. Akad. Van Wet. Proc. Ser. B Phys. Sci. 1898, 1, 1898–1899. [Google Scholar]
- Christoff, K.; Gordon, A.M.; Smallwood, J.; Smith, R.; Schooler, J.W. Experience sampling during fMRI reveals default network and executive system contributions to mind wandering. Proc. Natl. Acad. Sci. USA 2009, 106, 8719–8724. [Google Scholar] [CrossRef] [PubMed]
- Mason, M.F.; Norton, M.I.; Van Horn, J.D.; Wegner, D.M.; Grafton, S.T.; Macrae, C.N. Wandering Minds: The Default Network and Stimulus-Independent Thought. Science 2007, 315, 393–395. [Google Scholar] [CrossRef]
- Barron, E.; Riby, L.M.; Greer, J.; Smallwood, J. Absorbed in thought: The effect of mind wandering on the processing of relevant and irrelevant events. Psychol. Sci. 2011, 22, 596–601. [Google Scholar] [CrossRef]
- Smallwood, J.; Beach, E.; Schooler, J.W.; Handy, T.C. Going AWOL in the brain: Mind wandering reduces cortical analysis of external events. J. Cogn. Neurosci. 2008, 20, 458–469. [Google Scholar] [CrossRef]
- Arnau, S.; Löffler, C.; Rummel, J.; Hagemann, D.; Wascher, E.; Schubert, A. Inter-trial alpha power indicates mind wandering. Psychophysiology 2020, 57, e13581. [Google Scholar] [CrossRef]
- Baldwin, C.L.; Roberts, D.M.; Barragan, D.; Lee, J.D.; Lerner, N.; Higgins, J.S. Detecting and Quantifying Mind Wandering during Simulated Driving. Front. Hum. Neurosci. 2017, 11, 406. [Google Scholar] [CrossRef]
- Compton, R.J.; Gearinger, D.; Wild, H. The wandering mind oscillates: EEG alpha power is enhanced during moments of mind-wandering. Cogn. Affect. Behav. Neurosci. 2019, 19, 1184–1191. [Google Scholar] [CrossRef]
- Braboszcz, C.; Delorme, A. Lost in thoughts: Neural markers of low alertness during mind wandering. NeuroImage 2010, 54, 3040–3047. [Google Scholar] [CrossRef]
- Van Son, D.; De Blasio, F.M.; Fogarty, J.S.; Angelidis, A.; Barry, R.J.; Putman, P. Frontal EEG theta/beta ratio during mind wandering episodes. Biol. Psychol. 2018, 140, 19–27. [Google Scholar] [CrossRef]
- Angelidis, A.; Van der Does, W.; Schakel, L.; Putman, P. Frontal EEG theta/beta ratio as an electrophysiological marker for attentional control and its test-retest reliability. Biol. Psychol. 2016, 121, 49–52. [Google Scholar] [CrossRef]
- Putman, P.; Verkuil, B.; Arias-Garcia, E.; Pantazi, I.; Van Schie, C. EEG theta/beta ratio as a potential biomarker for attentional control and resilience against deleterious effects of stress on attention. Cogn. Affect. Behav. Neurosci. 2013, 14, 782–791. [Google Scholar] [CrossRef]
- Schutter, D.J.; Van Honk, J. Electrophysiological ratio markers for the balance between reward and punishment. Cogn. Brain Res. 2005, 24, 685–690. [Google Scholar] [CrossRef]
- Barry, R.J.; Clarke, A.R.; Johnstone, S.; McCarthy, R.; Selikowitz, M. Electroencephalogram θ/β Ratio and Arousal in Attention-Deficit/Hyperactivity Disorder: Evidence of Independent Processes. Biol. Psychiatry 2009, 66, 398–401. [Google Scholar] [CrossRef] [PubMed]
- Barry, R.J.; Clarke, A.R.; McCarthy, R.; Selikowitz, M.; A Rushby, J.; Ploskova, E. EEG differences in children as a function of resting-state arousal level. Clin. Neurophysiol. 2004, 115, 402–408. [Google Scholar] [CrossRef]
- Barry, R.J.; Clarke, A.R.; Johnstone, S.J. A review of electrophysiology in attention deficit/hyperactivity disorder: I. Qualitative and quantitative electroencephalography. Clin. Neurophysiol. 2003, 114, 171–183. [Google Scholar] [CrossRef]
- Clarke, A.R.; Barry, R.J.; Dupuy, F.E.; McCarthy, R.; Selikowitz, M.; Johnstone, S.J. Excess beta activity in the EEG of children with attention-deficit/hyperactivity disorder: A disorder of arousal? Int. J. Psychophysiol. 2013, 89, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.R.; Barry, R.J.; Karamacoska, D.; Johnstone, S.J. The EEG theta/beta ratio: A marker of arousal or cognitive processing capacity? Appl. Psychophysiol. Biofeedback 2019, 44, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Lubar, J.F. Discourse on the development of EEG diagnostics and biofeedback for attention-deficit/hyperactivity disorders. Appl. Psychophysiol. Biofeedback 1991, 16, 201–225. [Google Scholar] [CrossRef] [PubMed]
- Mann, C.A.; Lubar, J.F.; Zimmerman, A.W.; Miller, C.A.; Muenchen, R.A. Quantitative analysis of EEG in boys with attention-deficit-hyperactivity disorder: Controlled study with clinical implications. Pediatr. Neurol. 1992, 8, 30–36. [Google Scholar] [CrossRef]
- Clayton, E.; Rajkowski, J.; Cohen, J.D.; Aston-Jones, G. Phasic Activation of Monkey Locus Ceruleus Neurons by Simple Decisions in a Forced-Choice Task. J. Neurosci. 2004, 24, 9914–9920. [Google Scholar] [CrossRef] [PubMed]
- Derryberry, D.; Reed, M.A. Anxiety-related attentional biases and their regulation by attentional control. J. Abnorm. Psychol. 2002, 111, 225–236. [Google Scholar] [CrossRef]
- Putman, P.; van Peer, J.; Maimari, I.; van der Werff, S. EEG theta/beta ratio in relation to fear-modulated response-inhibition, attentional control, and affective traits. Biol. Psychol. 2010, 83, 73–78. [Google Scholar] [CrossRef]
- Massar, S.; Rossi, V.; Schutter, D.; Kenemans, J. Baseline EEG theta/beta ratio and punishment sensitivity as biomarkers for feedback-related negativity (FRN) and risk-taking. Clin. Neurophysiol. 2012, 123, 1958–1965. [Google Scholar] [CrossRef]
- Massar, S.A.; Kenemans, J.L.; Schutter, D.J. Resting-state EEG theta activity and risk learning: Sensitivity to reward or punishment? Int. J. Psychophysiol. 2014, 91, 172–177. [Google Scholar] [CrossRef]
- Schutter, D.J.L.G.; Knyazev, G.G. Cross-frequency coupling of brain oscillations in studying motivation and emotion. Motiv. Emot. 2011, 36, 46–54. [Google Scholar] [CrossRef]
- Schutter, I.; Kenemans, J.L.; Schutter, D.J.L.G. Resting-state theta/beta EEG ratio is associated with reward- and punishment-related reversal learning. Cogn. Affect. Behav. Neurosci. 2017, 17, 754–763. [Google Scholar] [CrossRef]
- Van Son, D.; de Rover, M.; De Blasio, F.M.; van der Does, W.; Barry, R.J.; Putman, P. Electroencephalography theta/beta ratio covaries with mind wandering and functional connectivity in the executive control network. Ann. N. Y. Acad. Sci. 2019, 1452, 52. [Google Scholar] [CrossRef] [PubMed]
- Hasenkamp, W.; Wilson-Mendenhall, C.D.; Duncan, E.; Barsalou, L.W. Mind wandering and attention during focused meditation: A fine-grained temporal analysis of fluctuating cognitive states. NeuroImage 2011, 59, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Fontanini, A.; Bower, J.M. Slow-waves in the olfactory system: An olfactory perspective on cortical rhythms. Trends Neurosci. 2006, 29, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Heck, D.H.; McAfee, S.; Liu, Y.; Babajani-Feremi, A.; Rezaie, R.; Freeman, W.J.; Wheless, J.W.; Papanicolaou, A.C.; Ruszinkó, M.; Sokolov, Y.; et al. Breathing as a Fundamental Rhythm of Brain Function. Front. Neural Circuits 2017, 10, 115. [Google Scholar] [CrossRef]
- Ito, J.; Roy, S.K.; Liu, Y.; Cao, Y.; Fletcher, M.L.; Lu, L.; Boughter, J.D.; Grün, S.; Heck, D.H. Whisker barrel cortex delta oscillations and gamma power in the awake mouse are linked to respiration. Nat. Commun. 2014, 5, 3572. [Google Scholar] [CrossRef]
- Moberly, A.H.; Schreck, M.; Bhattarai, J.P.; Zweifel, L.S.; Luo, W.; Ma, M. Olfactory inputs modulate respiration-related rhythmic activity in the prefrontal cortex and freezing behavior. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Jerath, R.; Edry, J.W.; Barnes, V.A.; Jerath, V. Physiology of long pranayamic breathing: Neural respiratory elements may provide a mechanism that explains how slow deep breathing shifts the autonomic nervous system. Med. Hypotheses 2006, 67, 566–571. [Google Scholar] [CrossRef]
- Farb, N.A.S.; Segal, Z.V.; Anderson, A.K. Attentional Modulation of Primary Interoceptive and Exteroceptive Cortices. Cereb. Cortex 2012, 23, 114–126. [Google Scholar] [CrossRef]
- Fuxe, K.; Hökfelt, T.; Ungerstedt, U. Morphological and Functional Aspects of Central Monoamine Neurons. Int. Rev. Neurobiol. 1970, 13, 93–126. [Google Scholar] [CrossRef]
- Loughlin, S.; Foote, S.; Bloom, F. Efferent projections of nucleus locus coeruleus: Topographic organization of cells of origin demonstrated by three-dimensional reconstruction. Neuroscience 1986, 18, 291–306. [Google Scholar] [CrossRef]
- Carter, M.; Yizhar, O.; Chikahisa, S.; Nguyen, H.; Adamantidis, A.; Nishino, S.; Deisseroth, K.; De Lecea, L. Tuning arousal with optogenetic modulation of locus coeruleus neurons. Nat. Neurosci. 2010, 13, 1526–1533. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Cohen, J.D. An integrative theory of locus coeruleus-norepinephrine function: Adaptive Gain and Optimal Performance. Annu. Rev. Neurosci. 2005, 28, 403–450. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Rajkowski, J.; Kubiak, P. Conditioned responses of monkey locus coeruleus neurons anticipate acquisition of discriminative behavior in a vigilance task. Neuroscience 1997, 80, 697–715. [Google Scholar] [CrossRef]
- Bouret, S.; Sara, S.J. Reward expectation, orientation of attention and locus coeruleus-medial frontal cortex interplay during learning. Eur. J. Neurosci. 2004, 20, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Rajkowski, J.; Majczyński, H.; Clayton, E.; Aston-Jones, G. Activation of Monkey Locus Coeruleus Neurons Varies With Difficulty and Performance in a Target Detection Task. J. Neurophysiol. 2004, 92, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Usher, M.; Cohen, J.D.; Servan-Schreiber, D.; Rajkowski, J.; Aston-Jones, G. The Role of Locus Coeruleus in the Regulation of Cognitive Performance. Science 1999, 283, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Aston-Jones, G.; Rajkowski, J.; Lu, W.; Zhu, Y.; Cohen, J.D.; Morecraft, R.J. Prominent projections from the orbital prefrontal cortex to the locus coeruleus in monkey. J. Neurosci. 2002, 28, 86–89. [Google Scholar]
- Chandler, D.J.; Gao, W.-J.; Waterhouse, B.D. Heterogeneous organization of the locus coeruleus projections to prefrontal and motor cortices. Proc. Natl. Acad. Sci. USA 2014, 111, 6816–6821. [Google Scholar] [CrossRef]
- Rajkowski, J.; Lu, W.; Zhu, Y.; Cohen, J.; Aston-Jones, G. Prominent projections from the anterior cingulate cortex to the locus coeruleus in Rhesus monkey. Soc. Neurosci. Abstr. 2000, 26, 2230. [Google Scholar]
- Zhu, Y.; Iba, M.; Rajkowski, J.; Aston-Jones, G. Projection from the orbitofrontal cortex to the locus coeruleus in monkeys revealed by anterograde tracing. Soc. Neurosci. Abstr. 2004, 30, 211–213. [Google Scholar]
- Aston-Jones, G.; Rajkowski, J.; Cohen, J. Role of locus coeruleus in attention and behavioral flexibility. Biol. Psychiatry 1999, 46, 1309–1320. [Google Scholar] [CrossRef]
- Ivanova, S.; Rajkowski, J.; Silakov, V.; Watanabe, T.; Aston-Jones, G. Local chemomanipulations of locus coeruleus (LC) activity in monkeys alter cortical event-related potentials (ERPs) and task performance. Soc. Neurosci. Abstr. 1997, 23, 1587. [Google Scholar]
- Biancardi, V.; Bicego, K.; Almeida, M.C.; Gargaglioni, L.H. Locus coeruleus noradrenergic neurons and CO2 drive to breathing. Pflügers Arch.-Eur. J. Physiol. 2007, 455, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Filosa, J.A.; Dean, J.B.; Putnam, R.W. Role of intracellular and extracellular pH in the chemosensitive response of rat locus coeruleus neurones. J. Physiol. 2002, 541, 493–509. [Google Scholar] [CrossRef] [PubMed]
- Gargaglioni, L.H.; Hartzler, L.K.; Putnam, R.W. The locus coeruleus and central chemosensitivity. Respir. Physiol. Neurobiol. 2010, 173, 264–273. [Google Scholar] [CrossRef]
- Noronha-De-Souza, C.R.; Bícego, K.C.; Michel, G.; Glass, M.L.; Branco, L.G.S.; Gargaglioni, L.H. Locus coeruleus is a central chemoreceptive site in toads. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R997–R1006. [Google Scholar] [CrossRef]
- Oyamada, Y.; Ballantyne, D.; Mückenhoff, K.; Scheid, P. Respiration-modulated membrane potential and chemosensitivity of locus coeruleus neurones in thein vitrobrainstem-spinal cord of the neonatal rat. J. Physiol. 1998, 513, 381–398. [Google Scholar] [CrossRef]
- Pineda, J.; Aghajanian, G. Carbon dioxide regulates the tonic activity of locus coeruleus neurons by modulating a proton- and polyamine-sensitive inward rectifier potassium current. Neuroscience 1997, 77, 723–743. [Google Scholar] [CrossRef]
- Band, D.M.; Cameron, I.R.; Semple, S.J. Oscillations in arterial pH with breathing in the cat. J. Appl. Physiol. 1969, 26, 261–267. [Google Scholar] [CrossRef]
- Band, D.M.; Wolff, C.B.; Ward, J.; Cochrane, G.M.; Prior, J. Respiratory oscillations in arterial carbon dioxide tension as a control signal in exercise. Nature 1980, 283, 84–85. [Google Scholar] [CrossRef] [PubMed]
- Honda, Y.; Ueda, M. Fluctuations of arterial PH associated with the respiratory cycle in dogs. Jpn. J. Physiol. 1961, 11, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Li, Y.; Kalwani, R.M.; Gold, J.I. Relationships between Pupil Diameter and Neuronal Activity in the Locus Coeruleus, Colliculi, and Cingulate Cortex. Neuron 2015, 89, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rodenkirch, C.; Moskowitz, N.; Schriver, B.; Wang, Q. Dynamic Lateralization of Pupil Dilation Evoked by Locus Coeruleus Activation Results from Sympathetic, Not Parasympathetic, Contributions. Cell Rep. 2017, 20, 3099–3112. [Google Scholar] [CrossRef] [PubMed]
- Reimer, J.; McGinley, M.J.; Liu, Y.; Rodenkirch, C.; Wang, Q.; A McCormick, D.; Tolias, A.S. Pupil fluctuations track rapid changes in adrenergic and cholinergic activity in cortex. Nat. Commun. 2016, 7, 13289. [Google Scholar] [CrossRef]
- Unsworth, N.; Robison, M.K. Pupillary correlates of lapses of sustained attention. Cogn. Affect. Behav. Neurosci. 2016, 16, 601–615. [Google Scholar] [CrossRef]
- Varazzani, C.; San-Galli, A.; Gilardeau, S.; Bouret, S. Noradrenaline and Dopamine Neurons in the Reward/Effort Trade-Off: A Direct Electrophysiological Comparison in Behaving Monkeys. J. Neurosci. 2015, 35, 7866–7877. [Google Scholar] [CrossRef]
- Lachaux, J.P.; Rodriguez, E.; Martinerie, J.; Varela, F.J. Measuring phase synchrony in brain signals. Hum. Brain Mapp. 1999, 8, 194–208. [Google Scholar] [CrossRef]
- Barnett, L.; Seth, A.K. The MVGC multivariate Granger causality toolbox: A new approach to Granger-causal inference. J. Neurosci. Methods 2014, 223, 50–68. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Berkovits, I.; Hancock, G.R.; Nevitt, J. Bootstrap Resampling Approaches for Repeated Measure Designs: Relative Robustness to Sphericity and Normality Violations. Educ. Psychol. Meas. 2000, 60, 877–892. [Google Scholar] [CrossRef]
- Wilson, D.J. The harmonic mean p-value for combining dependent tests. Proc. Natl. Acad. Sci. USA 2019, 116, 1195–1200. [Google Scholar] [CrossRef]
- Field, A. Discovering Statistics Using IBM SPSS Statistics; Sage: Los Angeles, CA, USA, 2013. [Google Scholar]
- Lehmann, A. Uber die Beziehung swischen Athmung und Aufmerksamkeit. Phil. Stud. 1894, 9, 66–91. [Google Scholar]
- Lange, N. Beitrige zur Theorie der Aufmerksamkeit und der activen Apperception. Phil. Stud. 1888, 4, 390–422. [Google Scholar]
- Urbantschitsch, V. Ueber eine Eigenthiimlichkeit der Schallempfindungen geringster Intensittit, Centralbl. f. d. med. Wiss. 1875. Available online: https://dejure.org/dienste/vernetzung/rechtsprechung?Gericht=BVerfG&Datum=15.01.1958&Aktenzeichen=1%20BvR%20400%2F51 (accessed on 31 August 2021).
- Langner, R.; Eickhoff, S.B. Sustaining attention to simple tasks: A meta-analytic review of the neural mechanisms of vigilant attention. Psychol. Bull. 2013, 139, 870–900. [Google Scholar] [CrossRef] [PubMed]
- Robertson, I.H.; Ridgeway, V.; Greenfield, E.; Parr, A. Motor recovery after stroke depends on intact sustained attention: A 2-year follow-up study. Neuropsychology 1997, 11, 290–295. [Google Scholar] [CrossRef]
- Stadler, M.; Erke, H. Über einige periodische Vorgänge in der Figuralwahrnehmung. Vis. Res. 1968, 8, 1081–1092. [Google Scholar] [CrossRef]
- Sripada, C.S. An Exploration/Exploitation Trade-off between Mind-Wandering and Goal-Directed Thinking. Oxf. Handb. Spontaneous Thought Mind-Wander. Creat. Dreaming 2018, 34, 23–34. [Google Scholar] [CrossRef]
- Foxe, J.J.; Snyder, A.C. The Role of Alpha-Band Brain Oscillations as a Sensory Suppression Mechanism during Selective Attention. Front. Psychol. 2011, 2, 154. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W.; Sauseng, P.; Hanslmayr, S. EEG alpha oscillations: The inhibition–timing hypothesis. Brain Res. Rev. 2007, 53, 63–88. [Google Scholar] [CrossRef] [PubMed]
- Mathewson, K.E.; Lleras, A.; Beck, D.M.; Fabiani, M.; Ro, T.; Gratton, G. Pulsed Out of Awareness: EEG Alpha Oscillations Represent a Pulsed-Inhibition of Ongoing Cortical Processing. Front. Psychol. 2011, 2, 99. [Google Scholar] [CrossRef]
- Pfurtscheller, G. Induced Oscillations in the Alpha Band: Functional Meaning. Epilepsia 2003, 44, 2–8. [Google Scholar] [CrossRef]
- Uusberg, A.; Uibo, H.; Kreegipuu, K.; Allik, J. EEG alpha and cortical inhibition in affective attention. Int. J. Psychophysiol. 2013, 89, 26–36. [Google Scholar] [CrossRef]
- Canteroa, J.L.; Atienzaa, M.; Salasa, R.M. Alpha EEG coherence in different brain states: An electrophysiological index of the arousal level in human subjects. Neurosci. Lett. 1999, 271, 170. [Google Scholar] [CrossRef]
- Mikutta, C.; Altorfer, A.; Strik, W.; Koenig, T. Emotions, arousal, and frontal alpha rhythm asymmetry during Beethoven’s 5th symphony. Brain Topogr. 2012, 25, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Eysenck, M. Attention and Arousal: Cognition and Performance; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Yerkes, R.M.; Dodson, J.D. The relation of strength of stimulus to rapidity of habit-formation. J. Comp. Neurol. Psychol. 1908, 18, 459–482. [Google Scholar] [CrossRef]
- Garey, J.; Goodwillie, A.; Frohlich, J.; Morgan, M.; Gustafsson, J.A.; Smithies, O.; Korach, K.; Ogawa, S.; Pfaff, D.W. Genetic contributions to generalized arousal of brain and behavior. Proc. Natl. Acad. Sci. USA 2003, 100, 11019–11022. [Google Scholar] [CrossRef] [PubMed]
- Hebb, D.O. Drives and the c.n.s. (conceptual nervous system)*. Psychol. Rev. 1966, 62, 67–84. [Google Scholar] [CrossRef]
- Pfaff, D.W. Brain Arousal and Information Theory: Neural and Genetic Mechanisms; Harvard University Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Bradley, M.M.; Miccoli, L.; Escrig, M.A.; Lang, P.J. The pupil as a measure of emotional arousal and autonomic activation. Psychophysiology 2008, 45, 602–607. [Google Scholar] [CrossRef]
- Bradshaw, J.L. Pupil Size as a Measure of Arousal during Information Processing. Nat. Cell Biol. 1967, 216, 515–516. [Google Scholar] [CrossRef]
- Hess, E.H.; Polt, J.M. Pupil Size as Related to Interest Value of Visual Stimuli. Science 1960, 132, 349–350. [Google Scholar] [CrossRef]
- Huijbers, W.; Pennartz, C.; Beldzik, E.; Domagalik, A.; Vinck, M.; Hofman, W.F.; Cabeza, R.; Daselaar, S.M. Respiration phase-locks to fast stimulus presentations: Implications for the interpretation of posterior midline “deactivations”. Hum. Brain Mapp. 2014, 35, 4932–4943. [Google Scholar] [CrossRef]
- Glover, G.H.; Li, T.Q.; Ress, D. Image-based method for retrospective correction of physiological motion effects in fMRI: RETROICOR. Magn. Reson. Med. 2000, 44, 162–167. [Google Scholar] [CrossRef]
- Birn, R.M. The role of physiological noise in resting-state functional connectivity. NeuroImage 2012, 62, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Zotev, V.; Phillips, R.; Bodurka, J. Correlated slow fluctuations in respiration, EEG, and BOLD fMRI. NeuroImage 2013, 79, 81–93. [Google Scholar] [CrossRef]
- Lorenz, E.N. Deterministic nonperiodic flow. J. Atmos. Sci. 1963, 20, 130–141. [Google Scholar] [CrossRef]
- Kox, M.; van Eijk, L.T.; Zwaag, J.; Wildenberg, J.V.D.; Sweep, F.; van der Hoeven, J.G.; Pickkers, P. Voluntary activation of the sympathetic nervous system and attenuation of the innate immune response in humans. Proc. Natl. Acad. Sci. USA 2014, 111, 7379–7384. [Google Scholar] [CrossRef] [PubMed]
- Muzik, O.; Reilly, K.T.; Diwadkar, V.A. “Brain over body”—A study on the willful regulation of autonomic function during cold exposure. NeuroImage 2018, 172, 632–641. [Google Scholar] [CrossRef]
- Berwian, I.M.; Wenzel, J.G.; Collins, A.G.E.; Seifritz, E.; Stephan, K.E.; Walter, H.; Huys, Q.J.M. Computational Mechanisms of Effort and Reward Decisions in Patients With Depression and Their Association With Relapse After Antidepressant Discontinuation. JAMA Psychiatry 2020, 77, 513–522. [Google Scholar] [CrossRef]
- Grueschow, M.; Stenz, N.; Thörn, H.; Ehlert, U.; Breckwoldt, J.; Maeder, M.B.; Exadaktylos, A.K.; Bingisser, R.; Ruff, C.C.; Kleim, B. Real-world stress resilience is associated with the responsivity of the locus coeruleus. Nat. Commun. 2021, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kaldewaij, R.; Koch, S.B.J.; Hashemi, M.M.; Zhang, W.; Klumpers, F.; Roelofs, K. Anterior prefrontal brain activity during emotion control predicts resilience to post-traumatic stress symptoms. Nat. Hum. Behav. 2021, 5, 1055–1064. [Google Scholar] [CrossRef]
- Kurniawan, I.T.; Grueschow, M.; Ruff, C.C. Anticipatory Energization Revealed by Pupil and Brain Activity Guides Human Effort-Based Decision Making. J. Neurosci. 2021, 41, 6328–6342. [Google Scholar] [CrossRef] [PubMed]
- Maier, S.U.; Grueschow, M. Pupil dilation predicts individual self-regulation success across domains. Sci. Rep. 2021, 11, 1–18. [Google Scholar] [CrossRef]
- Marzo, A.; Bai, J.; Otani, S. Neuroplasticity regulation by noradrenaline in mammalian brain. Curr. Neuropharmacol. 2009, 7, 286–295. [Google Scholar] [CrossRef]
- Chen, M.J.; Nguyen, T.V.; Pike, C.J.; Russo-Neustadt, A.A. Norepinephrine induces BDNF and activates the PI-3K and MAPK cascades in embryonic hippocampal neurons. Cell. Signal. 2007, 19, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Ivy, A.S.; Rodriguez, F.G.; Garcia, C.; Chen, M.J.; Russo-Neustadt, A.A. Noradrenergic and serotonergic blockade inhibits BDNF mRNA activation following exercise and antidepressant. Pharmacol. Biochem. Behav. 2003, 75, 81–88. [Google Scholar] [CrossRef]
- Russo-Neustadt, A.A.; Alejandre, H.; Garcia, C.; Ivy, A.S.; Chen, M.J. Hippocampal brain-derived neurotrophic factor expression following treatment with reboxetine, citalopram, and physical exercise. Neuropsychopharmacology 2004, 29, 2189–2199. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
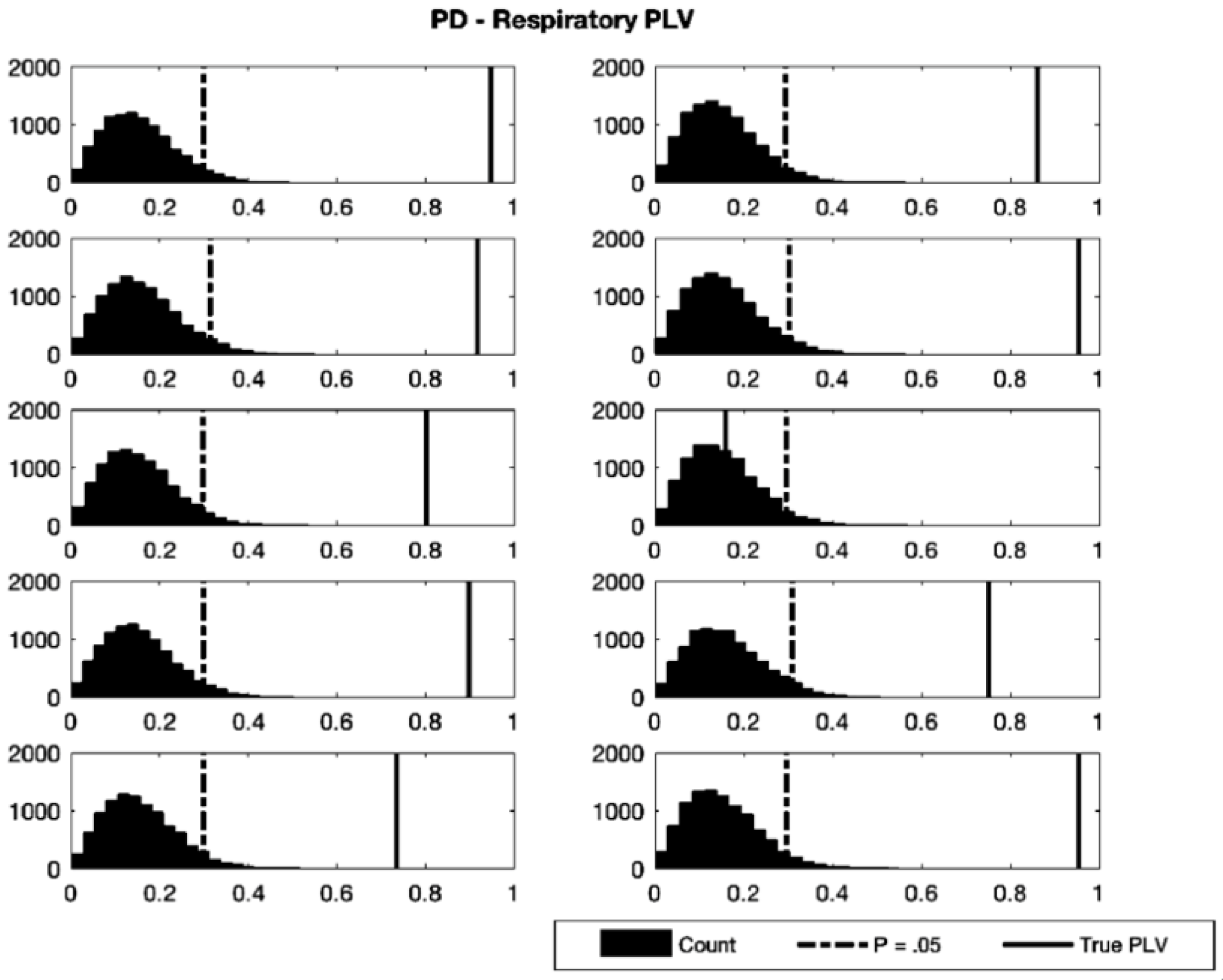
| Participant | Test PLV | Critical PLV | p-Value |
|---|---|---|---|
| 1 | 0.500 | 0.294 | 0.0001 |
| 2 | 0.568 | 0.295 | <0.0001 |
| 3 | 0.523 | 0.296 | 0.0002 |
| 4 | 0.644 | 0.300 | <0.0001 |
| 5 | 0.239 | 0.297 | 0.147 |
| 6 | 0.400 | 0.297 | 0.005 |
| 7 | 0.291 | 0.295 | 0.046 |
| 8 | 0.588 | 0.304 | <0.0001 |
| 9 | 0.390 | 0.302 | 0.005 |
| 10 | 0.103 | 0.295 | 0.036 |
| 11 | 0.079 | 0.296 | 0.371 |
| 0.0003 |
| Participant | Test PLV | Critical PLV | p-Value |
|---|---|---|---|
| 1 | 0.948 | 0.296 | <0.0001 |
| 2 | 0.862 | 0.295 | <0.0001 |
| 3 | 0.917 | 0.305 | <0.0001 |
| 4 | 0.955 | 0.302 | <0.0001 |
| 5 | 0.803 | 0.299 | <0.0001 |
| 6 | 0.160 | 0.297 | 0.414 |
| 7 | 0.899 | 0.298 | <0.0001 |
| 8 | 0.753 | 0.306 | <0.0001 |
| 9 | 0.736 | 0.303 | <0.0001 |
| 10 | 0.955 | 0.297 | <0.0001 |
| < 0.0001 |
| Model Order | A.C. Lags | Epochs | Mean MCD | Harmonic Mean (p-Value) | |
|---|---|---|---|---|---|
| P1 | 17 | 7052 | 1 | 0.0002 | 0.49 |
| P2 | 63 | 20,000 | 1 | 0.0090 | 0.00020 |
| P3 | 50 | 10,000 | 41 | 0.0023 | 0.00015 |
| P4 | 15 | 3144 | 20 | 0.0061 | 0.00012 |
| P5 | 17 | 10,000 | 1 | 0.0480 | 0.00005 |
| P6 | 12 | 10,000 | 1 | 0.0093 | 0.00020 |
| P7 | 40 | 10,000 | 1 | 0.0010 | 0.00055 |
| P8 | 40 | 10,000 | 1 | 0.0017 | 0.00020 |
| P9 | 60 | 20,000 | 1 | 0.0090 | 0.00030 |
| P10 | 40 | 10,000 | 1 | 0.0109 | 0.00028 |
| Grand Mean | 0.0097 | 0.00017 |
| Resp Pupil | Resp Fz | Pupil Resp | Pupil Fz | Fz Resp | Fz Pupil | |
|---|---|---|---|---|---|---|
| P1 | 0.00021 (0.61) | 0.00027 (0.32) | 0.00025 (0.43) | 0.00023 (0.51) | 0.00018 (0.74) | 0.00022 (0.54) |
| P2 | 0.0017 (<0.0001) * | 0.0016 (<0.0001) * | 0.048 (<0.0001) * | 0.0009 (0.27) | 0.0011 (0.036) | 0.0007 (0.79) |
| P3 | 0.0009 (0.23) | 0.0018 (<0.0001) * | 0.006 (<0.0001) * | 0.0009 (0.28) | 0.0024 (<0.0001) * | 0.0016 (0.0001) * |
| P4 | 0.0005 (0.01) * | 0.0065 (<0.0001) * | 0.0019 (<0.0001) * | 0.0245 (<0.0001) * | 0.0022 (<0.0001) * | 0.0010 (<0.0001) * |
| P5 | 0.0005 (0.001) * | 0.0002 (0.41) | <0.0001 (>0.99) | 0.0003 (0.08) | 0.2870 (<0.00001) * | 0.0005 (<0.0001) * |
| P6 | 0.0003 (0.03) | 0.0112 (<0.0001) * | 0.0062 (<0.0001) * | 0.0003 (0.03) | 0.0373 (<0.0001) * | 0.0004 (0.004) * |
| P7 | 0.0005 (0.95) | 0.0013 (0.001) * | 0.0006 (0.69) | 0.0007 (0.45) | 0.002 (<0.0001) * | 0.001 (0.08) |
| P8 | 0.0005 (0.44) | 0.0017 (<0.0001) * | 0.0015 (<0.0001) * | 0.0005 (0.33) | 0.0059 (<0.0001) * | 0.0005 (0.42) |
| P9 | 0.0030 (<0.0001) * | 0.0008 (0.098) | 0.0483 (<0.0001) * | 0.0002 (>0.99) | 0.0007 (0.24) | 0.0009 (0.01) * |
| P10 | 0.0011 (<0.001) * | 0.0028 (<0.0001) * | 0.0008 (0.06) | 0.0005 (0.66) | 0.0590 (<0.0001) * | 0.0010 (0.002) * |
| Mean MCD | 0.00106 | 0.002817 | 0.011365 | 0.002903 | 0.039778 | 0.000782 |
| Fisher’s χ2 (p-value) | 86.3 (<0.0001) * | 133.05 (<0.0001) * | 118.60 (<0.0001) * | 41.64 (0.0031) * | 143.65 (<0.0001) * | 96.43 (<0.0001) * |
| Resp | Fz | Pupil | |
|---|---|---|---|
| P1 | 2.0001 (0.99) | 2.0040 (0.60) | 2.0040 (0.18) |
| P2 | 1.9979 (0.77) | 2.0017 (0.82) | 1.9797 (<0.01) |
| P3 | 2.0037 (0.65) | 2.0064 (0.43) | 1.9965 (0.66) |
| P4 | 1.9830 (0.05) | 1.9244 (<0.01) | 2.0004 (0.97) |
| P5 | 1.9778 (<0.01) | 1.9999 (0.98) | 2.0015 (0.82) |
| P6 | 1.9881 (0.10) | 1.9997 (0.96) | 1.9306 (<0.01) |
| P7 | 2.0197 (0.0213) | 2.0028 (0.75) | 2.0017 (0.85) |
| P8 | 1.9961 (0.57) | 2.0007 (0.92) | 1.9120 (< 0.01) |
| P9 | 2.0017 (0.80) | 2.0005 (0.95) | 1.7753 * (<0.01) |
| P10 | 2.0128 (0.09) | 1.9996 (0.96) | 1.9997 (0.96) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melnychuk, M.C.; Robertson, I.H.; Plini, E.R.G.; Dockree, P.M. A Bridge between the Breath and the Brain: Synchronization of Respiration, a Pupillometric Marker of the Locus Coeruleus, and an EEG Marker of Attentional Control State. Brain Sci. 2021, 11, 1324. https://doi.org/10.3390/brainsci11101324
Melnychuk MC, Robertson IH, Plini ERG, Dockree PM. A Bridge between the Breath and the Brain: Synchronization of Respiration, a Pupillometric Marker of the Locus Coeruleus, and an EEG Marker of Attentional Control State. Brain Sciences. 2021; 11(10):1324. https://doi.org/10.3390/brainsci11101324
Chicago/Turabian StyleMelnychuk, Michael Christopher, Ian H. Robertson, Emanuele R. G. Plini, and Paul M. Dockree. 2021. "A Bridge between the Breath and the Brain: Synchronization of Respiration, a Pupillometric Marker of the Locus Coeruleus, and an EEG Marker of Attentional Control State" Brain Sciences 11, no. 10: 1324. https://doi.org/10.3390/brainsci11101324
APA StyleMelnychuk, M. C., Robertson, I. H., Plini, E. R. G., & Dockree, P. M. (2021). A Bridge between the Breath and the Brain: Synchronization of Respiration, a Pupillometric Marker of the Locus Coeruleus, and an EEG Marker of Attentional Control State. Brain Sciences, 11(10), 1324. https://doi.org/10.3390/brainsci11101324