Where Sounds Occur Matters: Context Effects Influence Processing of Salient Vocalisations


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Visual Stimuli
2.3. Auditory Stimuli
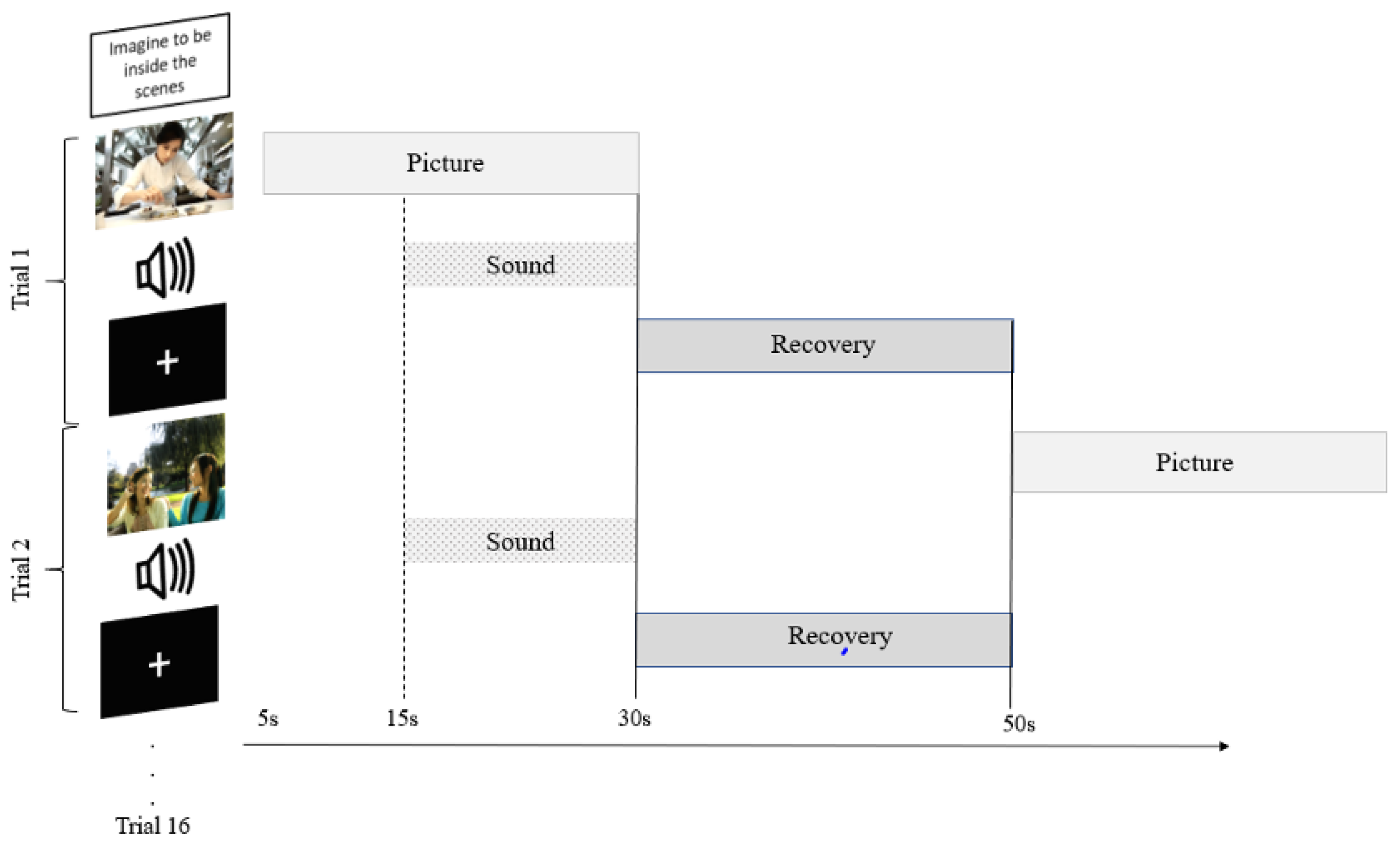
2.4. Experimental Design
2.5. Procedure
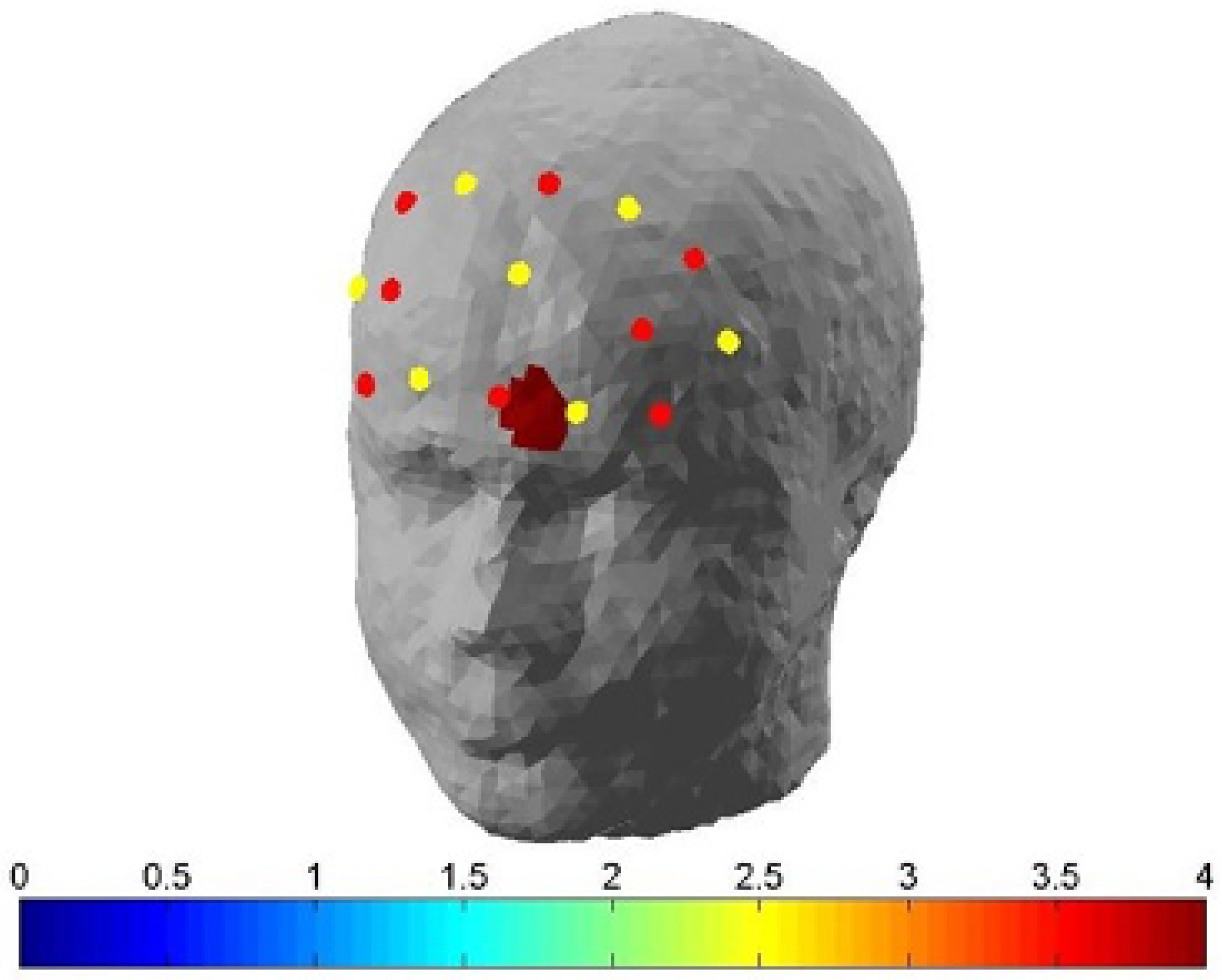
2.6. Functional Near-Infrared Spectroscopy (fNIRS) Data Acquisition
2.7. fNIRS Pre-Processing and Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BA | Brodmann Area |
| CH | Channel |
| DE | Domestic Environment |
| DeoxyHB | Deoxyhemoglobin |
| DLPFC | Dorso Lateral Pre Frontal Cortex |
| fNIRS | functiona Near-InfraRed Spectroscopy |
| GLM | General Linear Model |
| HRF | Haemodynamic Response Function |
| OE | Outdoor Environment |
| OxyHB | Oxygenated Haemoglobin |
| PFC | Prefrontal Cortex |
| AC | Adult Cry |
| CS | Control Sounds |
| IC | Infant Cry |
| IL | Infant Laugh |
References
- Patel, S.; Scherer, K.R.; Björkner, E.; Sundberg, J. Mapping emotions into acoustic space: The role of voice production. Biol. Psychol. 2011, 87, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, J.; Patel, S.; Bjorkner, E.; Scherer, K.R. Interdependencies among Voice Source Parameters in Emotional Speech. IEEE Trans. Affect. Comput. 2011, 2, 162–174. [Google Scholar] [CrossRef]
- Sauter, D.A.; Eisner, F.; Calder, A.J.; Scott, S.K. Perceptual cues in nonverbal vocal expressions of emotion. Q. J. Exp. Psychol. 2010, 63, 2251–2272. [Google Scholar] [CrossRef] [PubMed]
- Sauter, D.A.; Eisner, F.; Ekman, P.; Scott, S.K. Emotional vocalisations are recognized across cultures regardless of the valence of distractors. Psychol. Sci. 2015, 26, 354–356. [Google Scholar] [CrossRef]
- Lima, C.F.; Anikin, A.; Monteiro, A.C.; Scott, S.K.; Castro, S.L. Automaticity in the recognition of nonverbal emotional vocalisations. Emotion 2018, 19, 219. [Google Scholar] [CrossRef]
- Maren, S.; Phan, K.L.; Liberzon, I. The contextual brain: Implications for fear conditioning, extinction and psychopathology. Nat. Rev. Neurosci. 2013, 14, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Ibañez, A.; Manes, F. Contextual social cognition and the behavioral variant of frontotemporal dementia. Neurology 2012, 78, 1354–1362. [Google Scholar] [CrossRef] [PubMed]
- Barbas, H.; Zikopoulos, B.; Timbie, C. Sensory pathways and emotional context for action in primate prefrontal cortex. Biol. Psychiatry 2011, 69, 1133–1139. [Google Scholar] [CrossRef]
- Lang, S.; Kroll, A.; Lipinski, S.J.; Wessa, M.; Ridder, S.; Christmann, C.; Schad, L.R.; Flor, H. Context conditioning and extinction in humans: Differential contribution of the hippocampus, amygdala and prefrontal cortex. Eur. J. Neurosci. 2009, 29, 823–832. [Google Scholar] [CrossRef]
- Herry, C.; Bach, D.R.; Esposito, F.; Di Salle, F.; Perrig, W.J.; Scheffler, K.; Lüthi, A.; Seifritz, E. Processing of temporal unpredictability in human and animal amygdala. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 5958–5966. [Google Scholar] [CrossRef]
- Shackman, A.J.; Maxwell, J.S.; McMenamin, B.W.; Greischar, L.L.; Davidson, R.J. Stress Potentiates Early and Attenuates Late Stages of Visual Processing. Neuroscience 2011, 31, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Bachorowski, J.A.; Owren, M.J. Sounds of emotion: Production and perception of affect-related vocal acoustics. Ann. N. Y. Acad. Sci. 2003, 1000, 244–265. [Google Scholar] [CrossRef]
- Cheney, D.L.; Seyfarth, R.M. Cognitive reappraisal of emotion: A meta-analysis of human neuroimaging studies. Proc. Natl. Acad. Sci. USA 2018, 115, 1974–1979. [Google Scholar] [CrossRef] [PubMed]
- Crivelli, C.; Fridlund, A.J. Facial displays are tools for social influence. Trends Cogn. Sci. 2018, 22, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Rigo, P.; De Pisapia, N.; Bornstein, M.H.; Esposito, G.; Venuti, P. Brain processes in women and men in response to emotive sounds. Soc. Neurosci. 2017, 12, 150–162. [Google Scholar] [CrossRef][Green Version]
- Rigo, P.; Esposito, G.; Bornstein, M.H.; De Pisapia, N.; Manzardo, C.; Venuti, P. Brain processes in mothers and nulliparous women in response to cry in different situational contexts: A default mode network study. Parent. Sci. Pract. 2019, 19, 69–85. [Google Scholar] [CrossRef]
- Atias, D.; Liraz, S.; Eidinger, A.; Dror, I.; Maymon, Y.; Aviezer, H. Loud and unclear: Intense real-life vocalisations during affective situations are perceptually ambiguous and contextually malleable. J. Exp. Psychol. Gen. 2019, 148, 1842–1848. [Google Scholar] [CrossRef]
- Baker, J.M.; Liu, N.; Cui, X.; Vrticka, P.; Saggar, M.; Hosseini, S.M.H.; Reiss, A.L. Sex differences in neural and behavioral signatures of cooperation revealed by fNIRS hyperscanning. Sci. Rep. 2016, 6, 26492. [Google Scholar] [CrossRef]
- Yovel, G.; Belin, P. A unified coding strategy for processing faces and voices. Trends Cogn. Sci. 2013, 17, 263–271. [Google Scholar] [CrossRef]
- Ethofer, T.; Anders, S.; Wiethoff, S.; Erb, M.; Herbert, C.; Saur, R.; Grodd, W.; Wildgruber, D. Effects of prosodic emotional intensity on activation of associative auditory cortex. Neuroreport 2006, 17, 249–253. [Google Scholar] [CrossRef]
- Friederici, A.D. The cortical language circuit: From auditory perception to sentence comprehension. Trends Cogn. Sci. 2012, 16, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Rauschecker, J.P.; Scott, S.K. Maps and streams in the auditory cortex: Nonhuman primates illuminate human speech processing. Nat. Neurosci. 2009, 12, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Frühholz, S.; Grandjean, D. Processing of emotional vocalisations in bilateral inferior frontal cortex. Neurosci. Biobehav. Rev. 2013, 37, 2847–2855. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, A.; Kotz, S.A. Beyond the right hemisphere: Brain mechanisms mediating vocal emotional processing. Trends Cogn. Sci. 2006, 10, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Buhle, J.T.; Silvers, J.A.; Wager, T.D.; Lopez, R.; Onyemekwu, C.; Kober, H.; Weber, J.; Ochsner, K.N. Cognitive reappraisal of emotion: A meta-analysis of human neuroimaging studies. Cereb. Cortex 2014, 24, 2981–2990. [Google Scholar] [CrossRef]
- Jung, Y.; Larsen, B.; Walther, D.B. Modality-Independent Coding of Scene Categories in Prefrontal Cortex. J. Neurosci. Off. J. Soc. Neurosci. 2018, 38, 5969–5981. [Google Scholar] [CrossRef]
- Azhari, A.; Truzzi, A.; Rigo, P.; Bornstein, M.H.; Esposito, G. Putting salient vocalisations in context: Adults’ physiological arousal to emotive cues in domestic and external environments. Physiol. Behav. 2018, 196, 25–32. [Google Scholar] [CrossRef]
- Davies, C.D.; Craske, M.G. Psychophysiological responses to unpredictable threat: Effects of cue and temporal unpredictability. Emotion 2015, 15, 195–200. [Google Scholar] [CrossRef]
- Kveraga, K.; Ghuman, A.S.; Bar, M. Top-down predictions in the cognitive brain. Brain Cogn. 2007, 65, 145–168. [Google Scholar] [CrossRef]
- Lorberbaum, J.P.; Newman, J.D.; Horwitz, A.R.; Dubno, J.R.; Lydiard, R.B.; Hamner, M.B.; Bohning, D.E.; George, M.S. A potential role for thalamocingulate circuitry in human maternal behavior. Biol. Psychiatry 2002, 51, 431–445. [Google Scholar] [CrossRef]
- Swain, J.E.; Lorberbaum, J.P. Imaging the Human Parental Brain. Neurobiol. Parent. Brain 2008, 83–100. [Google Scholar]
- Azhari, A.; Leck, W.Q.; Gabrieli, G.; Bizzego, A.; Rigo, P.; Setoh, P.; Bornstein, M.H.; Esposito, G. Parenting Stress Undermines Mother-Child Brain-to-Brain Synchrony: A Hyperscanning Study. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Azhari, A.; Lim, M.; Bizzego, A.; Gabrieli, G.; Bornstein, M.H.; Esposito, G. Physical Presence of Spouse Enhances Brain-to-Brain Synchrony in Co-Parenting Couples. Sci. Rep. 2020, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Christoff, K.; Prabhakaran, V.; Dorfman, J.; Zhao, Z.; Kroger, J.K.; Holyoak, K.J.; Gabrieli, J. Rostrolateral prefrontal cortex involvement in relational integration during reasoning. NeuroImage 2001, 14, 1136–1149. [Google Scholar] [CrossRef] [PubMed]
- Bunge, S.A.; Helskog, E.H.; Wendelken, C. Left, but not right, rostrolateral prefrontal cortex meets a stringent test of the relational integration hypothesis. NeuroImage 2009, 46, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.P.; Barros, C.; Pedrosa, J. Salience in a social landscape: Electrophysiological effects of task-irrelevant and infrequent vocal change. Soc. Cogn. Affect. Neurosci. 2016, 11, 127–139. [Google Scholar] [CrossRef]
- Jessen, S.; Kotz, S.A. On the role of crossmodal prediction in audiovisual emotion perception. Front. Hum. Neurosci. 2013, 7, 369. [Google Scholar] [CrossRef]
- Chennu, S.; Noreika, V.; Gueorguiev, D.; Blenkmann, A.; Kochen, S.; Ibáñez, A.; Owen, A.M.; Bekinschtein, T.A. Expectation and attention in hierarchical auditory prediction. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 11194–11205. [Google Scholar] [CrossRef]
- Fenske, M.J.; Raymond, J.E. Affective Influences of Selective Attention. Curr. Dir. Psychol. Sci. 2006, 15, 312–316. [Google Scholar] [CrossRef]
- Sander, D.; Grandjean, D.; Pourtois, G.; Schwartz, S.; Seghier, M.L.; Scherer, K.R.; Vuilleumier, P. Emotion and attention interactions in social cognition: Brain regions involved in processing anger prosody. NeuroImage 2005, 28, 848–858. [Google Scholar] [CrossRef]
- Brück, C.; Kreifelts, B.; Wildgruber, D. Emotional voices in context: A neurobiological model of multimodal affective information processing. Phys. Life Rev. 2011, 8, 383–403. [Google Scholar] [CrossRef] [PubMed]
- Thayer, J.F.; Lane, R.D. Claude Bernard and the heart–brain connection: Further elaboration of a model of neurovisceral integration. Neurosci. Biobehav. Rev. 2009, 33, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, B.H.; Thayer, J.F.; Laberg, J.C.; Wormnes, B.; Raadal, M.; Skaret, E.; Kvale, G.; Berg, E. Attentional and physiological characteristics of patients with dental anxiety. J. Anxiety Disord. 2003, 17, 75–87. [Google Scholar] [CrossRef]
- Hansen, A.L.; Johnsen, B.H.; Thayer, J.F. Vagal influence on working memory and attention. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2003, 48, 263–274. [Google Scholar] [CrossRef]
- Hedger, N.; Adams, W.J.; Garner, M. Autonomic arousal and attentional orienting to visual threat are predicted by awareness. J. Exp. Psychol. Hum. Percept. Perform. 2015, 41, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, A.; Zysset, S.; Kotz, S.A.; von Cramon, D.Y. Gender differences in the activation of inferior frontal cortex during emotional speech perception. NeuroImage 2004, 12, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, A.; Striano, T.; Friederici, A.D. Sex differences in the preattentive processing of vocal emotional expressions. Neuroreport 2005, 16, 635–639. [Google Scholar] [CrossRef]
- Schirmer, A.; Simpson, E.; Escoffier, N. Listen up! Processing of intensity change differs for vocal and nonvocal sounds. Brain Res. 2007, 1176, 103–112. [Google Scholar] [CrossRef]
- Schirmer, A.; Kotz, S.A. ERP Evidence for a Sex-Specific Stroop Effect in Emotional Speech. J. Cogn. Neurosci. 2003, 15, 1135–1148. [Google Scholar] [CrossRef]
- Stevens, J.S.; Hamann, S. Sex differences in brain activation to emotional stimuli: A meta-analysis of neuroimaging studies. Neuropsychologia 2012, 50, 1156–1161. [Google Scholar] [CrossRef]
- Thompson, A.E.; Voyer, D. Sex differences in the ability to recognise non-verbal displays of emotion: A meta-analysis. Cogn. Emot. 2014, 28, 1164–1195. [Google Scholar] [CrossRef] [PubMed]
- Economist Intelligence Unit. Safe Cities Index 2019; The Economist Intelligence Unit Limited: London, UK, 2019. [Google Scholar]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azhari, A.; Rigo, P.; Bornstein, M.H.; Esposito, G. Where Sounds Occur Matters: Context Effects Influence Processing of Salient Vocalisations. Brain Sci. 2020, 10, 429. https://doi.org/10.3390/brainsci10070429
Azhari A, Rigo P, Bornstein MH, Esposito G. Where Sounds Occur Matters: Context Effects Influence Processing of Salient Vocalisations. Brain Sciences. 2020; 10(7):429. https://doi.org/10.3390/brainsci10070429
Chicago/Turabian StyleAzhari, Atiqah, Paola Rigo, Marc H. Bornstein, and Gianluca Esposito. 2020. "Where Sounds Occur Matters: Context Effects Influence Processing of Salient Vocalisations" Brain Sciences 10, no. 7: 429. https://doi.org/10.3390/brainsci10070429
APA StyleAzhari, A., Rigo, P., Bornstein, M. H., & Esposito, G. (2020). Where Sounds Occur Matters: Context Effects Influence Processing of Salient Vocalisations. Brain Sciences, 10(7), 429. https://doi.org/10.3390/brainsci10070429