Persistent Roseoloviruses Infection in Adult Patients with Epilepsy

 , , ,
, , , 
 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. HHV-6 and HHV-7 Serology
2.3. Nested Polymerase Chain Reaction
2.4. Restriction Endonuclease Analysis
2.5. Reverse Transcription PCR assays
2.6. Quantitative Real-Time PCR
2.7. HHV-6 Encoded Proteins Expression by Indirect Immunofluorescence
2.8. Cytokines Detection by Enzyme-Linked Immunosorbent Assay
2.9. Statistical Analysis
3. Results
3.1. Seroepidemiology
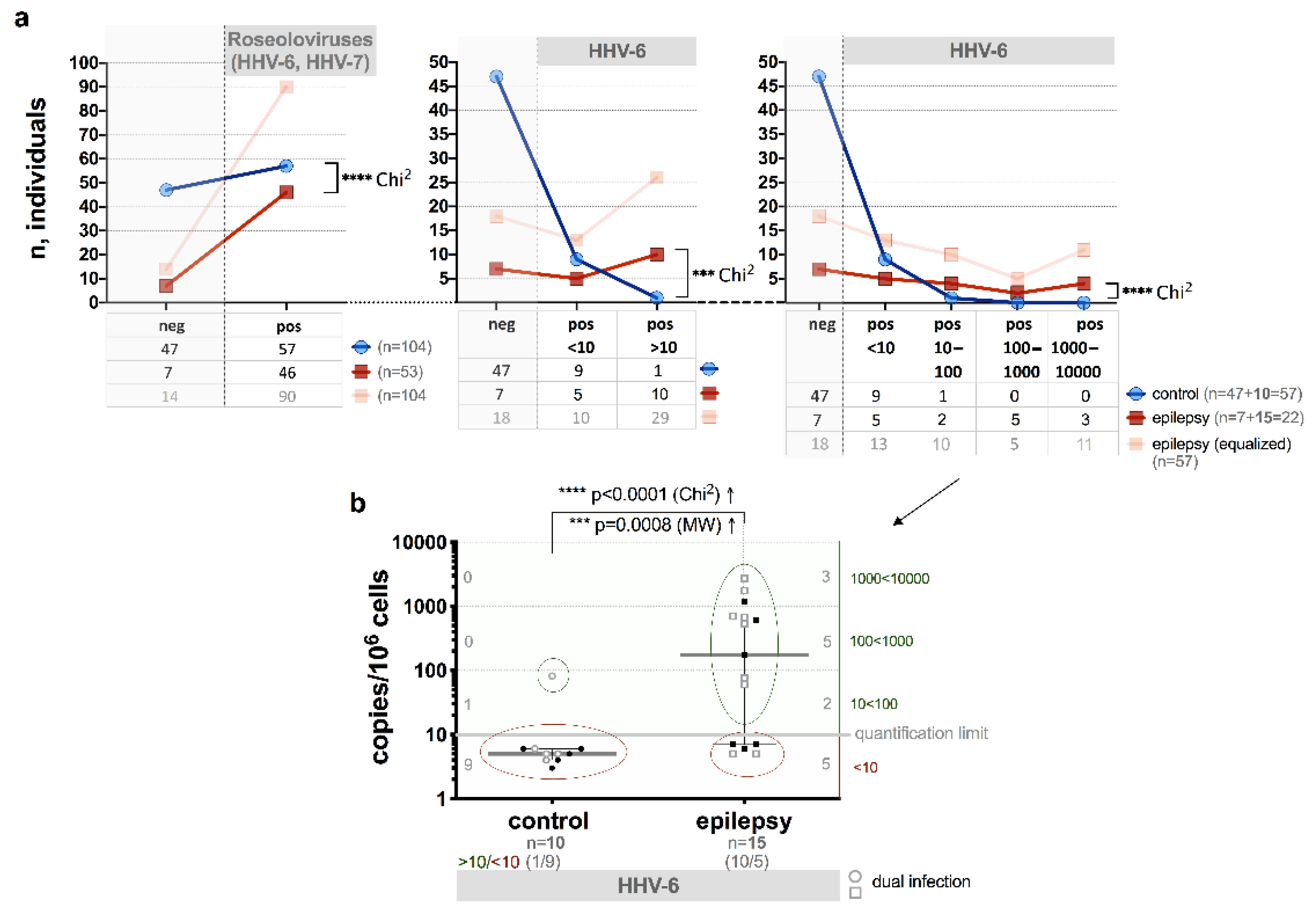
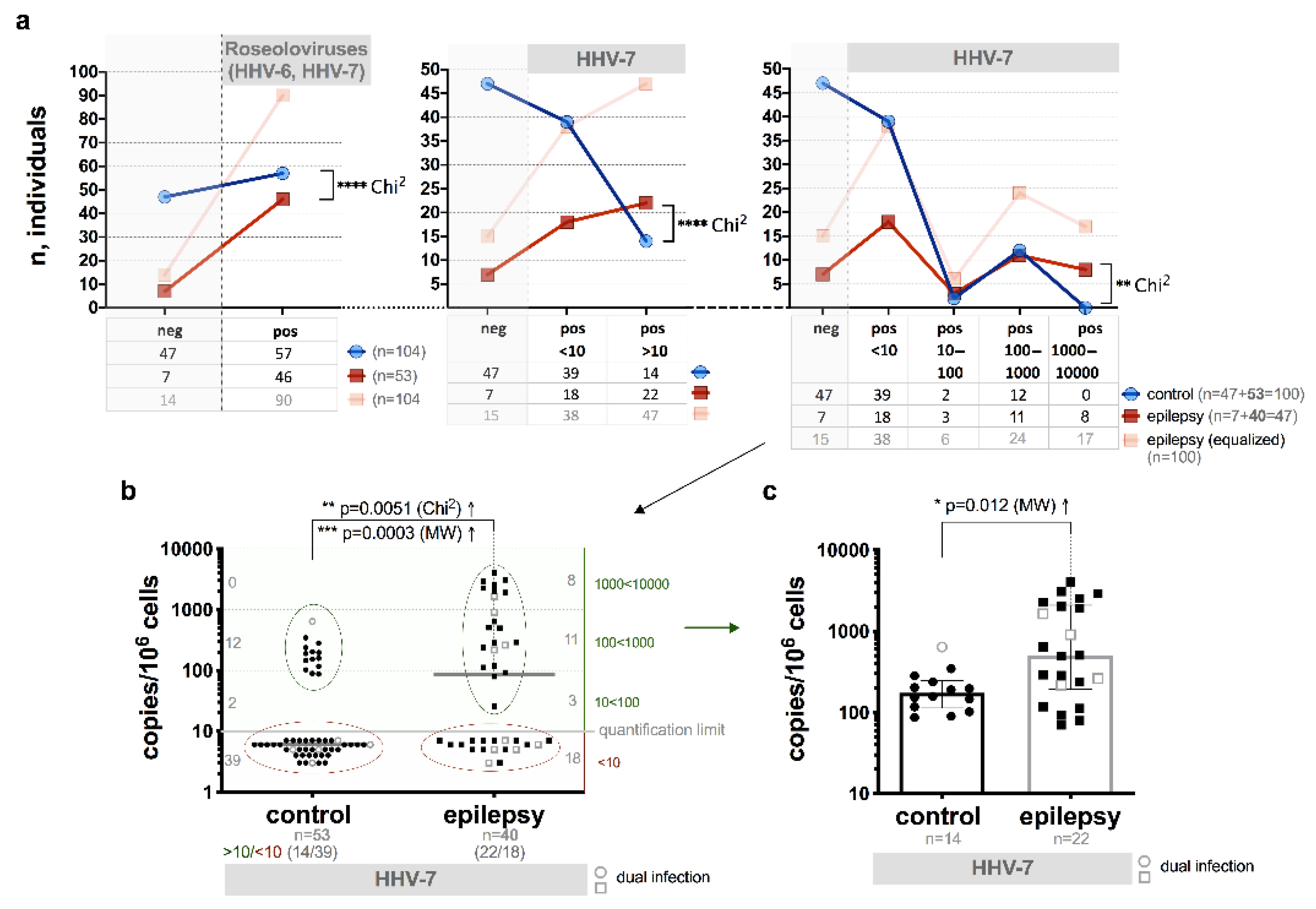
3.2. Presence of HHV-6 and/or HHV-7 Genomic Sequences
3.3. HHV-6 U89/90 Gene Expression
3.4. HHV-6 Load
3.5. HHV-6-Encoded Proteins Expression
3.6. HHV-7 U57 Gene Expression
3.7. HHV-7 Load
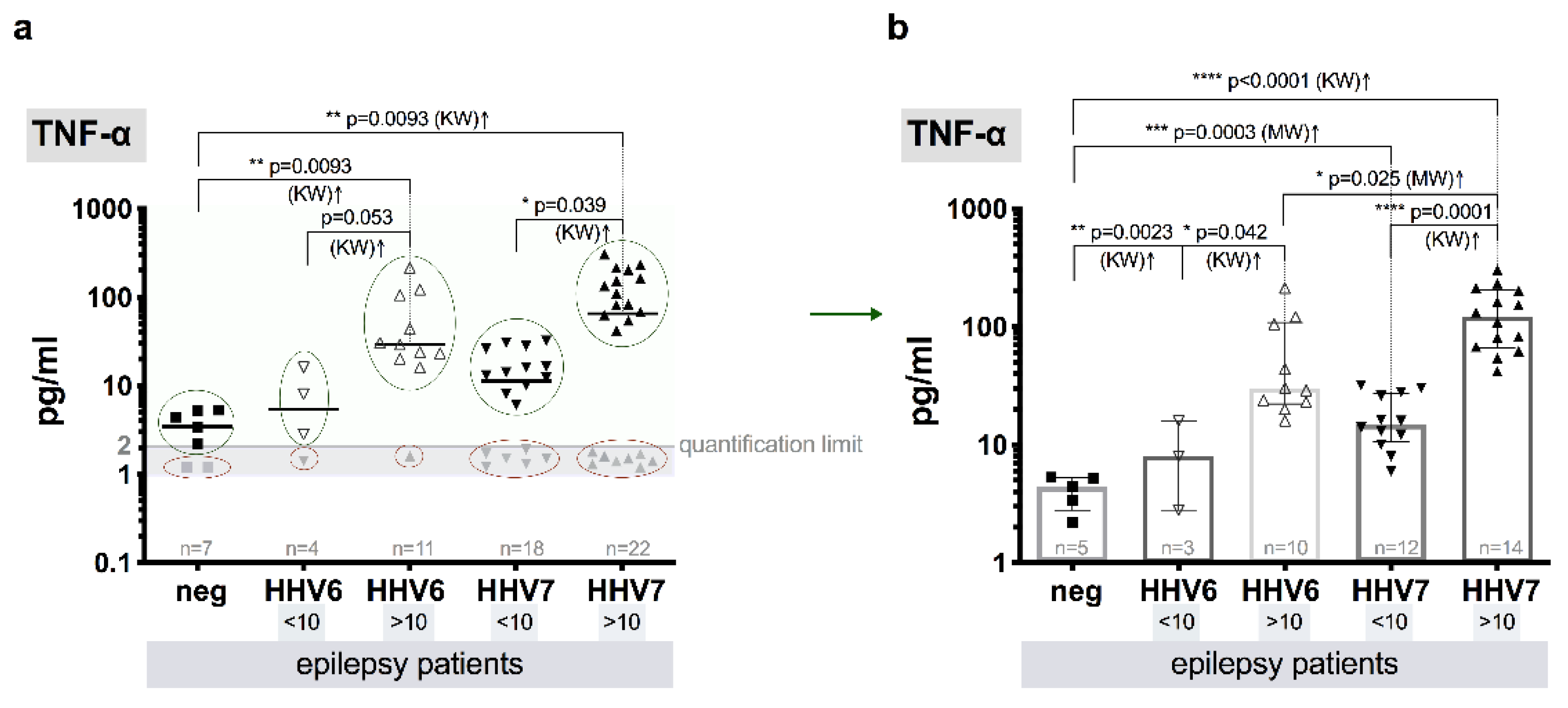
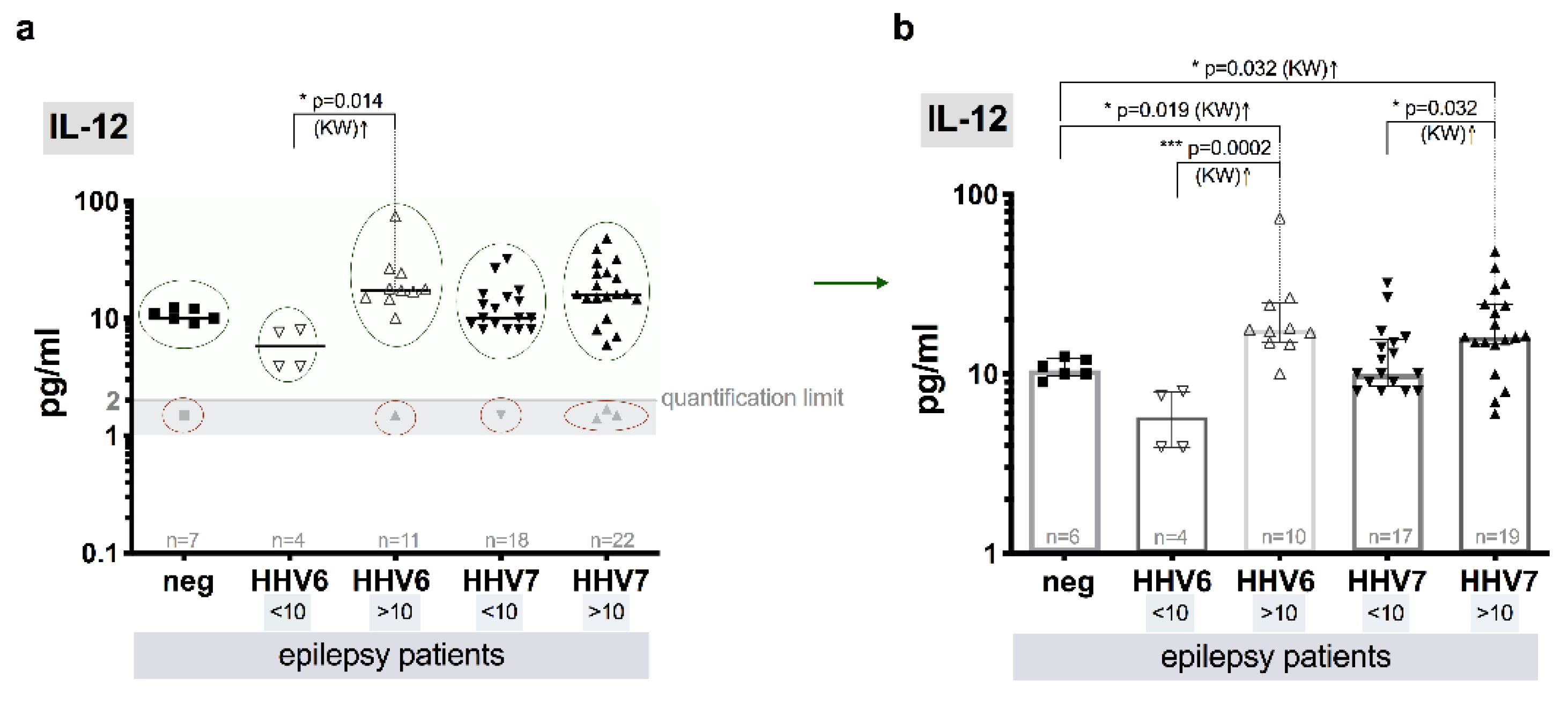
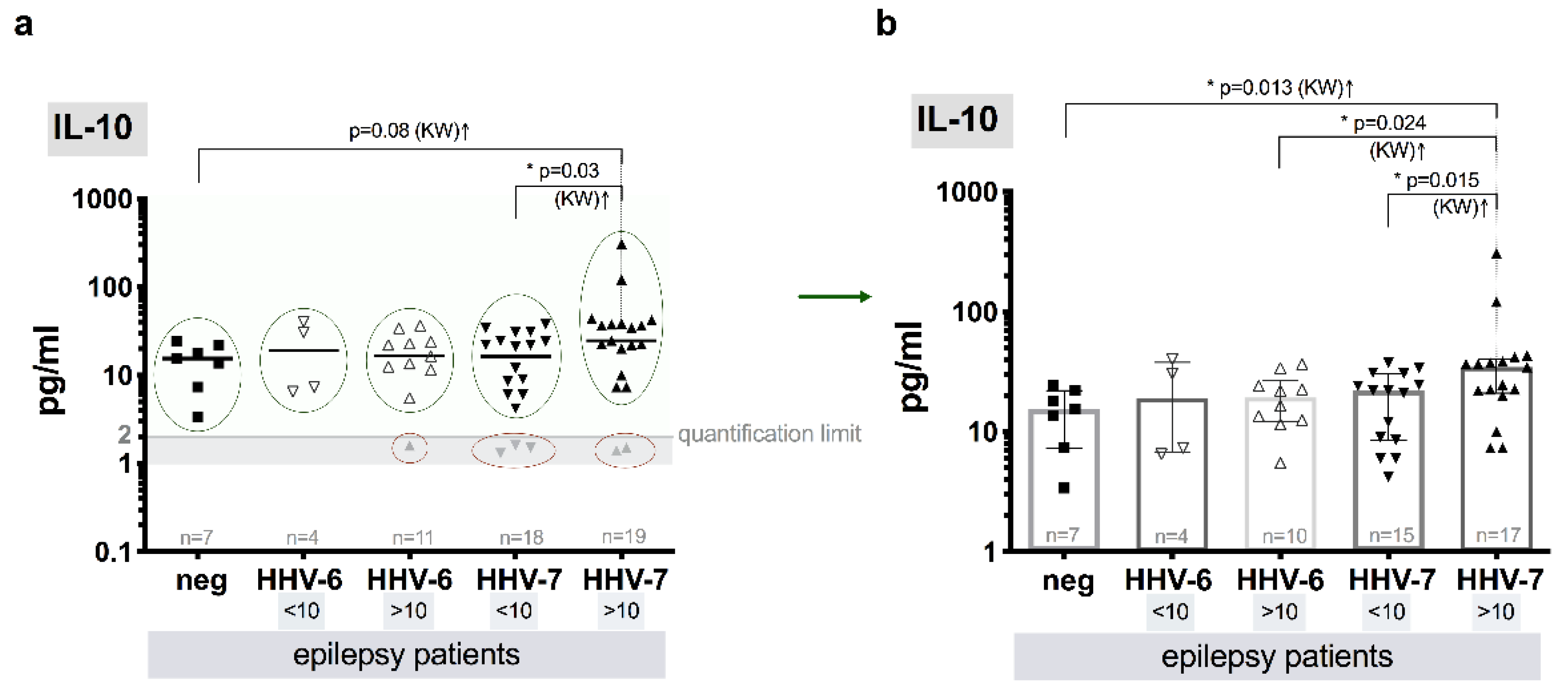
3.8. Level of Cytokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fisher, R.S.; Acevedo, C.; Arzimanoglou, A.; Bogacz, A.; Cross, J.H.; Elger, C.E.; Engel, J., Jr.; Forsgren, L.; French, J.A.; Glynn, M.; et al. ILAE official report: A practical clinical definition of epilepsy. Epilepsia 2014, 55, 475–482. [Google Scholar] [CrossRef]
- Fisher, R.S.; Boas, W.V.; Blume, W.; Elger, C.; Genton, P.; Lee, P.; Engel, J., Jr. Epileptic seizures and epilepsy: Definitions proposed by the International League Against Epilepsy (ILAE) and the International Bureau for Epilepsy (IBE). Epilepsia 2005, 46, 470–472. [Google Scholar] [CrossRef] [PubMed]
- Stafstrom, C.E.; Carmant, L. Seizures and epilepsy: An overview for neuroscientists. Cold Spring Harb. Perspect. Med. 2015, 5, a022426. [Google Scholar] [CrossRef] [PubMed]
- Kwan, P.; Brodie, M.J. Early identification of refractory epilepsy. N. Engl. J. Med. 2000, 342, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Berg, T.A.; Millichap, J.J. The 2010 revised classification of seizures and epilepsy. Continuum (Minneap Minn) 2013, 19, 571–597. [Google Scholar] [CrossRef] [PubMed]
- Engel, J., Jr.; Pitkänen, A.; Loeb, J.A.; Edward Dudek, F.; Bertram, I.I.I.E.H.; Cole, A.J.; Moshé, S.L.; Wiebe, S.; Jensen, F.E.; Mody, I.; et al. Epilepsy biomarkers. Epilepsia 2013, 54 (Suppl. 4), 61–69. [Google Scholar] [CrossRef]
- Koepp, M.J. The help of biomarkers in the prevention of epilepsy. Lancet Neurol. 2016, 15, 782–784. [Google Scholar] [CrossRef]
- Mercadé Cerdá, J.M.; López Gonzalez, F.J.; Serrano Castro, P.; Vilanova, C.; Campos Blanco, D.M.; Querol Pascual, M.R. Observational multicentre study into the use of antiepileptic drugs in Spanish neurology consultations. Neurologia 2020, 35, 115–125. [Google Scholar] [CrossRef]
- Vezzani, A.; Fujinami, R.S.; White, H.S.; Preux, P.M.; Blümcke, I.; Sander, J.W.; Löscher, W. Infections, inflammation and epilepsy. Acta Neuropathol. 2016, 131, 211–234. [Google Scholar] [CrossRef]
- Berneman, Z.N.; Ablashi, D.V.; Li, G.; Eger-Fletcher, M.; Reitz, M.S.; Hung, C.L.; Brus, I.; Komaroff, A.L.; Gallo, R.C. Human herpesvirus 7 is a T-lymphotropic virus and is related to, but significantly different from, human herpesvirus 6 and human cytomegalovirus. Proc. Natl. Acad. Sci. USA 1992, 89, 10552–10556. [Google Scholar] [CrossRef]
- Agut, H.; Bonnafous, P.; Gautheret-Dejean, A. Update on infections with human herpesviruses 6A, 6B, and 7. Med. Mal. Infect. 2017, 47, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Mori, Y.; Yamanishi, K. HHV-6A, 6B, and 7: Pathogenesis, host response, and clinical disease. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Harberts, E.; Yao, K.; Wohler, J.E.; Maric, D.; Ohayon, J.; Henkin, R.; Jacobson, S. Human herpesvirus-6 entry into the central nervous system through the olfactory pathway. Proc. Natl. Acad. Sci. USA 2011, 108, 13734–13739. [Google Scholar] [CrossRef]
- Skuja, S.; Zieda, A.; Ravina, K.; Chapenko, S.; Roga, S.; Teteris, O.; Groma, V.; Murovska, M. Structural and ultrastructural alterations in human olfactory pathways and possible associations with herpesvirus 6 infection. PLoS ONE 2017, 12, e0170071. [Google Scholar] [CrossRef] [PubMed]
- Santoro, F.; Kennedy, P.E.; Locatelli, G.; Malnati, M.S.; Berger, E.A.; Lusso, P. CD46 is a cellular receptor for human herpesvirus 6. Cell 1999, 99, 817–827. [Google Scholar] [CrossRef]
- Mazdeh, M.; Omrani, M.D.; Sayad, A.; Komaki, A.; Arsang-Jang, S.; Taheri, M.; Ghafouri-Fard, S. Expression analysis of cytokine coding genes in epileptic patients. Cytokine 2018, 110, 284–287. [Google Scholar] [CrossRef]
- Yao, K.; Crawford, J.R.; Komaroff, A.L.; Ablashi, D.V.; Jacobson, S. Review part 2: Human herpesvirus-6 in central nervous system diseases. J. Med. Virol. 2010, 82, 1669–1678. [Google Scholar] [CrossRef]
- Dagna, L.; Pritchett, J.C.; Lusso, P. Immunomodulation and immunosuppression by human herpesvirus 6A and 6B. Future Virol. 2013, 8, 273–287. [Google Scholar] [CrossRef]
- Lusso, P.; Secchiero, P.; Crowley, R.W.; Garzino-Demo, A.; Berneman, Z.N.; Gallo, R.C. CD4 is a critical component of the receptor for human herpesvirus 7: Interference with human immunodeficiency virus. Proc. Natl. Acad. Sci. USA 1994, 91, 3872–3876. [Google Scholar] [CrossRef]
- Secchiero, P.; Flamand, L.; Gibellini, D.; Falcieri, E.; Robuffo, I.; Capitani, S.; Gallo, R.C.; Zauli, G. Human Herpesvirus 7 induces CD4(+) T-cell death by two distinct mechanisms: Necrotic lysis in productively infected cells and apoptosis in uninfected or nonproductively infected cells. Blood 1997, 90, 4502–4512. [Google Scholar]
- Atedzoé, B.N.; Menezes, J.; D’Addario, M.; Xu, J.; Ongradi, J.; Ahmad, A. Modulatory effects of human herpes virus-7 on cytokine synthesis and cell proliferation in human peripheral blood mononuclear cell cultures. J. Leukoc. Biol. 1999, 66, 822–828. [Google Scholar] [CrossRef]
- Tadagaki, K.; Nakano, K.; Yamanishi, K. Human herpesvirus 7 open reading frames U12 and U51 encode functional beta-chemokine receptors. J. Virol. 2005, 79, 7068–7076. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Kondo, T.; Torigoe, S.; Okada, S.; Mukai, T.; Yamanishi, K. Human herpesvirus 7: Another causal agent for roseola (exanthem subitum). J. Pediatr. 1994, 125, 1–5. [Google Scholar] [CrossRef]
- Yamanishi, K.; Shiraki, K.; Kondo, T.; Okuno, T.; Takahashi, M.; Asano, Y.; Kurata, T. Identification of human herpesvirus-6 as a causal agent for exanthem subitum. Lancet 1988, 1, 1065–1067. [Google Scholar] [CrossRef]
- Hall, C.B.; Long, C.E.; Schnabel, K.C.; Caserta, M.T.; McIntyre, K.M.; Costanzo, M.A.; Knott, A.; Dewhurst, S.; Insel, R.A.; Epstein, L.G. Human herpesvirus-6 infection in children. A prospective study of complications and reactivation. N. Engl. J. Med. 1994, 331, 432–438. [Google Scholar] [CrossRef]
- Noguchi, T.; Yoshiura, T.; Hiwatashi, A.; Togao, O.; Yamashita, K.; Nagao, E.; Uchino, A.; Hasuo, K.; Atsumi, K.; Matsuura, T.; et al. CT and MRI findings of human herpesvirus 6-associated encephalopathy: Comparison with findings of herpes simplex virus encephalitis. AJR Am. J. Roentgenol. 2010, 194, 754–760. [Google Scholar] [CrossRef]
- Bozzola, E.; Krzysztofiak, A.; Bozzola, M.; Calcaterra, V.; Quondamcarlo, A.; Lancella, L.; Villani, A. HHV6 meningoencephalitis sequelae in previously healthy children. Infection 2012, 40, 563–566. [Google Scholar] [CrossRef]
- Hall, C.B.; Caserta, M.T.; Schnabel, K.C.; Long, C.; Epstein, L.G.; Insel, R.A.; Dewhurst, S. Persistence of human herpesvirus 6 according to site and variant: Possible greater neurotropism of variant A. Clin. Infect. Dis. 1998, 26, 132–137. [Google Scholar] [CrossRef]
- Caserta, M.T.; Hall, C.B.; Schnabel, K.; Long, C.E.; D’Heron, N. Primary human herpesvirus 7 infection: A comparison of human herpesvirus 7 and human herpesvirus 6 infections in children. J. Pediatr. 1998, 133, 386–389. [Google Scholar] [CrossRef]
- Epstein, L.G.; Shinnar, S.; Hesdorffer, D.C.; Nordli, D.R.; Hamidullah, A.; Benn, E.K.; Pellock, J.M.; Frank, L.M.; Lewis, D.V.; Moshe, S.L.; et al. Human herpesvirus 6 and 7 in febrile status epilepticus: The FEBSTAT study. Epilepsia 2012, 53, 1481–1488. [Google Scholar] [CrossRef]
- Suga, S.; Yoshikawa, T.; Nagai, T.; Asano, Y. Clinical features and virological findings in children with primary human herpesvirus 7 infection. Pediatrics 1997, 99, e4. [Google Scholar] [CrossRef]
- Torigoe, S.; Koide, W.; Yamada, M.; Miyashiro, E.; Tanaka-Taya, K.; Yamanishi, K. Human herpesvirus 7 infection associated with central nervous system manifestations. J. Pediatr. 1996, 129, 301–305. [Google Scholar] [CrossRef]
- Ward, K.N.; Kalima, P.; MacLeod, K.M.; Riordan, T. Neuroinvasion during delayed primary HHV-7 infection in an immunocompetent adult with encephalitis and flaccid paralysis. J. Med. Virol. 2002, 67, 538–541. [Google Scholar] [CrossRef] [PubMed]
- Tesini, B.L.; Epstein, L.G.; Caserta, M.T. Clinical impact of primary infection with roseoloviruses. Curr. Opin. Virol. 2014, 9, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmidt-DeMasters, B.K.; Gilden, D.H. The expanding spectrum of herpesvirus infections of the nervous system. Brain Pathol. 2001, 11, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Misra, U.K.; Tan, C.T.; Kalita, J. Viral encephalitis and epilepsy. Epilepsia 2008, 49 (Suppl. 6), 13–18. [Google Scholar] [CrossRef]
- Singhi, P. Infectious causes of seizures and epilepsy in the developing world. Dev. Med. Child Neurol. 2011, 53, 600–609. [Google Scholar] [CrossRef]
- Theodore, W.H. Epilepsy and viral infections. Epilepsy Curr. 2014, 14 (Suppl. 1), 35–42. [Google Scholar] [CrossRef]
- Chapenko, S.; Roga, S.; Skuja, S.; Rasa, S.; Cistjakovs, M.; Svirskis, S.; Zaserska, Z.; Groma, V.; Murovska, M. Detection frequency of human herpesviruses-6A, -6B, and -7 genomic sequences in central nervous system DNA samples from post-mortem individuals with unspecified encephalopathy. J. Neurovirol. 2016, 22, 488–497. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Asano, Y. Central nervous system complications in human herpesvirus-6 infection. Brain Dev. 2000, 22, 307–314. [Google Scholar] [CrossRef]
- Ongrádi, J.; Ablashi, D.V.; Yoshikawa, T.; Stercz, B.; Ogata, M. Roseolovirus-associated encephalitis in immunocompetent and immunocompromised individuals. J. Neurovirol. 2017, 23, 1–19. [Google Scholar] [CrossRef]
- Parra, M.; Alcala, A.; Amoros, C.; Baeza, A.; Galiana, A.; Tarragó, D.; García-Quesada, M.Á.; Sánchez-Hellín, V. Encephalitis associated with human herpesvirus-7 infection in an immunocompetent adult. Virol. J. 2017, 14, 97. [Google Scholar] [CrossRef] [PubMed]
- Riva, N.; Franconi, I.; Meschiari, M.; Franceschini, E.; Puzzolante, C.; Cuomo, G.; Bianchi, A.; Cavalleri, F.; Genovese, M.; Mussini, C. Acute human herpes virus 7 (HHV-7) encephalitis in an immunocompetent adult patient: A case report and review of literature. Infection 2017, 45, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Inazawa, N.; Hori, T.; Nojima, M.; Saito, M.; Igarashi, K.; Yamamoto, M.; Shimizu, N.; Yoto, Y.; Tsutsumi, H. Virus reactivations after autologous hematopoietic stem cell transplantation detected by multiplex PCR assay. J. Med. Virol. 2017, 89, 358–362. [Google Scholar] [CrossRef]
- Hall, C.B.; Caserta, M.T.; Schnabel, K.C.; McDermott, M.P.; Lofthus, G.K.; Carnahan, J.A.; Gilbert, L.M.; Dewhurst, S. Characteristics and acquisition of human herpesvirus (HHV) 7 infections in relation to infection with HHV-6. J. Infect. Dis. 2006, 193, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Donati, D.; Akhyani, N.; Fogdell–Hahn, A.; Cermelli, C.; Cassiani-Ingoni, R.; Vortmeyer, A.; Heiss, J.D.; Cogen, P.; Gaillard, W.D.; Sato, S.; et al. Detection of human herpesvirus-6 in mesial temporal lobe epilepsy surgical brain resections. Neurology 2003, 61, 1405–1411. [Google Scholar] [CrossRef]
- Esposito, L.; Drexler, J.F.; Braganza, O.; Doberentz, E.; Grote, A.; Widman, G.; Drosten, C.; Eis-Hübinger, A.M.; Schoch, S.; Elger, C.E.; et al. Large-scale analysis of viral nucleic acid spectrum in temporal lobe epilepsy biopsies. Epilepsia 2015, 56, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Fotheringham, J.; Akhyani, N.; Vortmeyer, A.; Donati, D.; Williams, E.; Oh, U.; Bishop, M.; Barrett, J.; Gea-Banacloche, J.; Jacobson, S. Detection of active human herpesvirus-6 infection in the brain: Correlation with polymerase chain reaction detection in cerebrospinal fluid. J. Infect. Dis. 2007, 195, 450–454. [Google Scholar] [CrossRef]
- Kawamura, Y.; Nakayama, A.; Kato, T.; Miura, H.; Ishihara, N.; Ihira, M.; Takahashi, Y.; Matsuda, K.; Yoshikawa, T. Pathogenic role of human herpesvirus 6B infection in mesial temporal lobe epilepsy. J. Infect. Dis. 2015, 212, 1014–1021. [Google Scholar] [CrossRef]
- Leibovitch, E.C.; Jacobson, S. Human herpesvirus 6 as a viral trigger in mesial temporal lobe epilepsy. J. Infect. Dis. 2015, 212, 1011–1013. [Google Scholar] [CrossRef]
- Niehusmann, P.; Mittelstaedt, T.; Bien, C.G.; Drexler, J.F.; Grote, A.; Schoch, S.; Becker, A.J. Presence of human herpes virus 6 DNA exclusively in temporal lobe epilepsy brain tissue of patients with history of encephalitis. Epilepsia 2010, 51, 2478–2483. [Google Scholar] [CrossRef]
- Theodore, W.H.; Epstein, L.; Gaillard, W.D.; Shinnar, S.; Wainwright, M.S.; Jacobson, S. Human herpes virus 6B: A possible role in epilepsy? Epilepsia 2008, 49, 1828–1837. [Google Scholar] [CrossRef] [PubMed]
- Wipfler, P.; Dunn, N.; Beiki, O.; Trinka, E.; Fogdell-Hahn, A. The Viral Hypothesis of Mesial Temporal Lobe Epilepsy-Is Human Herpes Virus-6 the Missing Link? A systematic review and meta-analysis. Seizure 2018, 54, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Li, J.M.; Huang, C.; Yan, B.; Wang, W.; Zhou, Q.; Sander, J.W.; Zhou, D. HHV-7 in adults with drug-resistant epilepsy: A pathological role in hippocampal sclerosis? J. Clin. Virol. 2014, 61, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Katsafanas, G.C.; Schirmer, E.C.; Wyatt, L.S.; Frenkel, N. In vitro activation of human herpesviruses 6 and 7 from latency. Proc. Natl. Acad. Sci. USA 1996, 93, 9788–9792. [Google Scholar] [CrossRef]
- Tanaka-Taya, K.; Kondo, T.; Nakagawa, N.; Inagi, R.; Miyoshi, H.; Sunagawa, T.; Okada, S.; Yamanishi, K. Reactivation of human herpesvirus 6 by infection of human herpesvirus 7. J. Med. Virol. 2000, 60, 284–289. [Google Scholar] [CrossRef]
- Ablashi, D.V.; Eastman, H.B.; Owen, C.B.; Roman, M.M.; Friedman, J.; Zabriskie, J.B.; Peterson, D.L.; Pearson, G.R.; Whitman, J.E. Frequent HHV-6 reactivation in multiple sclerosis (MS) and chronic fatigue syndrome (CFS) patients. J. Clin. Virol. 2000, 16, 179–191. [Google Scholar] [CrossRef]
- Challoner, P.B.; Smith, K.T.; Parker, J.D.; MacLeod, D.L.; Coulter, S.N.; Rose, T.M.; Schultz, E.R.; Bennett, J.L.; Garber, R.L.; Chang, M. Plaque-associated expression of human herpesvirus 6 in multiple sclerosis. Proc. Natl. Acad. Sci. USA 1995, 92, 7440–7444. [Google Scholar] [CrossRef]
- Chapenko, S.; Millers, A.; Nora, Z.; Logina, I.; Kukaine, R.; Murovska, M. Correlation between HHV-6 reactivation and multiple sclerosis disease activity. J. Med. Virol. 2003, 69, 111–117. [Google Scholar] [CrossRef]
- Leibovitch, E.C.; Jacobson, S. Evidence linking HHV-6 with multiple sclerosis: An update. Curr. Opin. Virol. 2014, 9, 127–133. [Google Scholar] [CrossRef]
- Nora-Krukle, Z.; Chapenko, S.; Logina, I.; Millers, A.; Platkajis, A.; Murovska, M. Human herpesvirus 6 and 7 reactivation and disease activity in multiple sclerosis. Medicina (Kaunas) 2011, 47, 527–531. [Google Scholar] [CrossRef]
- Opsahl, M.L.; Kennedy, P.G. Investigating the presence of human herpesvirus 7 and 8 in multiple sclerosis and normal control brain tissue. J. Neurol. Sci. 2006, 240, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Tomsone, V.; Logina, I.; Millers, A.; Chapenko, S.; Kozireva, S.; Murovska, M. Association of human herpesvirus 6 and human herpesvirus 7 with demyelinating diseases of the nervous system. J. Neurovirol. 2001, 7, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.S.; Bradley, A.S.; Bishop, K.N.; Kiani-Alikhan, S.; Ford, B. Chronic fatigue syndrome, the immune system and viral infection. Brain Behav. Immun. 2012, 26, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Chapenko, S.; Krumina, A.; Kozireva, S.; Nora, Z.; Sultanova, A.; Viksna, L.; Murovska, M. Activation of human herpesviruses 6 and 7 in patients with chronic fatigue syndrome. J. Clin. Virol. 2006, 37 (Suppl. 1), S47–S51. [Google Scholar] [CrossRef]
- Chapenko, S.; Krumina, A.; Logina, I.; Rasa, S.; Chistjakovs, M.; Sultanova, A.; Viksna, L.; Murovska, M. Association of active human herpesvirus-6, -7 and parvovirus b19 infection with clinical outcomes in patients with myalgic encephalomyelitis/chronic fatigue syndrome. Adv. Virol. 2012, 2012, 205085. [Google Scholar] [CrossRef] [PubMed]
- Rasa, S.; Nora-Krukle, Z.; Henning, N.; Eliassen, E.; Shikova, E.; Harrer, T.; Scheibenbogen, C.; Murovska, M.; Prusty, B.K. Chronic viral infections in myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS). J. Transl. Med. 2018, 16, 268. [Google Scholar] [CrossRef]
- Hogestyn, J.M.; Mock, D.J.; Mayer-Proschel, M. Contributions of neurotropic human herpesviruses herpes simplex virus 1 and human herpesvirus 6 to neurodegenerative disease pathology. Neural Regen. Res. 2018, 13, 211–221. [Google Scholar]
- Caselli, E.; D’Accolti, M.; Soffritti, I.; Zatelli, M.C.; Rossi, R.; degli Uberti, E.; Di Luca, D. HHV-6A in vitro infection of thyrocytes and T cells alters the expression of miRNA associated to autoimmune thyroiditis. Virol. J. 2017, 14, 3. [Google Scholar] [CrossRef]
- Sultanova, A.; Cistjakovs, M.; Gravelsina, S.; Chapenko, S.; Roga, S.; Cunskis, E.; Nora-Krukle, Z.; Groma, V.; Ventina, I.; Murovska, M. Association of active human herpesvirus-6 (HHV-6) infection with autoimmune thyroid gland diseases. Clin. Microbiol. Infect. 2017, 23, 50-e1–50-e5. [Google Scholar] [CrossRef]
- Secchiero, P.; Carrigan, D.R.; Asano, Y.; Benedetti, L.; Crowley, R.W.; Komaroff, A.L.; Gallo, R.C.; Lusso, P. Detection of human herpesvirus 6 in plasma of children with primary infection and immunosuppressed patients by polymerase chain reaction. J. Infect. Dis. 1995, 171, 273–280. [Google Scholar] [CrossRef]
- Pfeiffer, B.; Thomson, B.; Chandran, B. Identification and characterization of a cDNA derived from multiple splicing that encodes envelope glycoprotein gp105 of human herpesvirus 6. J. Virol. 1995, 69, 3490–3500. [Google Scholar] [CrossRef] [PubMed]
- Kozireva, S.; Užameckis, D.; Bariševs, M.; Murovska, M. Sensitivity and Reproducibility of Polymerase Chain Reaction Assays for Detection of Human Herpesviruses 6 and 7. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2009, 63, 180. [Google Scholar] [CrossRef]
- Van den Bosch, G.; Locatelli, G.; Geerts, L.; Fagà, G.; Ieven, M.; Goossens, H.; Bottiger, D.; Öberg, B.; Lusso, P.; Berneman, Z.N. Development of reverse transcriptase PCR assays for detection of active human herpesvirus 6 infection. J. Clin. Microbiol. 2001, 39, 2308–2310. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Shimizu, N.; Watanabe, K.; Saito, T.; Yoshioka, Y.; Sakane, E.; Tsunemine, H.; Akasaka, H.; Kodaka, T.; Takahashi, T. Analysis of viral infection by multiplex polymerase chain reaction assays in patients with liver dysfunction. Intern. Med. 2013, 52, 201–211. [Google Scholar] [CrossRef]
- Okuno, T.; Sao, H.; Asada, H.; Shiraki, K.; Takahashi, M.; Yamanishi, K. Analysis of a glycoprotein of human herpesvirus 6 (HHV-6) using monoclonal antibodies. Virology 1990, 176, 625–628. [Google Scholar] [CrossRef]
- Beal, S.L. Ways to fit a PK model with some data below the quantification limit. J. Pharmacokinet. Pharmacodyn. 2001, 28, 481–504. [Google Scholar] [CrossRef]
- Pantry, S.N.; Medveczky, P.G. Latency, Integration, and Reactivation of Human Herpesvirus-6. Viruses 2017, 9, 194. [Google Scholar] [CrossRef]
- Bonello, M.; Michael, B.D.; Solomon, T. Infective Causes of Epilepsy. Semin. Neurol. 2015, 35, 235–244. [Google Scholar] [CrossRef]
- Mirandola, P.; Menegazzi, P.; Merighi, S.; Ravaioli, T.; Cassai, E.; Di Luca, D. Temporal mapping of transcripts in herpesvirus 6 variants. J. Virol. 1998, 72, 3837–3844. [Google Scholar] [CrossRef]
- de Vries, E.E.; van den Munckhof, B.; Braun, K.P.; van Royen-Kerkhof, A.; de Jager, W.; Jansen, F.E. Inflammatory mediators in human epilepsy: A systematic review and meta-analysis. Neurosci. Biobehav. Rev. 2016, 63, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Vieira, É.L.; de Oliveira, G.N.; Lessa, J.M.; Gonçalves, A.P.; Oliveira, A.C.; Bauer, M.E.; Sander, J.W.; Cendes, F.; Teixeira, A.L. Peripheral leukocyte profile in people with temporal lobe epilepsy reflects the associated proinflammatory state. Brain Behav. Immun. 2016, 53, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Carlson, S.; Puttachary, S.; Sarkar, S.; Showman, L.; Putra, M.; Kanthasamy, A.G.; Thippeswamy, T. Role of the Fyn-PKCdelta signaling in SE-induced neuroinflammation and epileptogenesis in experimental models of temporal lobe epilepsy. Neurobiol. Dis. 2018, 110, 102–121. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Gao, Y.; Zhang, S.J.; Zhe, X.; Meng, F.L.; Qian, H.; Zhang, B.; Li, Y.J. Alteration of plasma cytokines in patients with active epilepsy. Acta Neurol. Scand. 2017, 135, 663–669. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Viral Genomic Sequences | Total Positive | |||||
|---|---|---|---|---|---|---|---|
| Single HHV-6 | Single HHV-7 | Co-Infection HHV-6 + HHV-7 | |||||
| WPB | Plasma | WPB | Plasma | WPB | Plasma | ||
| Epilepsy n = 53 | 6 (11.3%) | 2 b (33.3%) | 31 (58.5%) | 5 b (16.1%) | 9 * (17.0%) | 2 a,b (22.2%) | 46 ** (86.8%) |
| Control group n = 104 | 4 (3.8%) | 0 | 47 (45.2%) | 0 | 6 (5.8%) | 0 | 57 (54.8%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasa-Dzelzkaleja, S.; Gravelsina, S.; Chapenko, S.; Krukle, Z.-N.; Svirskis, S.; Suna, N.; Kashuba, E.; Karelis, G.; Murovska, M. Persistent Roseoloviruses Infection in Adult Patients with Epilepsy. Brain Sci. 2020, 10, 287. https://doi.org/10.3390/brainsci10050287
Rasa-Dzelzkaleja S, Gravelsina S, Chapenko S, Krukle Z-N, Svirskis S, Suna N, Kashuba E, Karelis G, Murovska M. Persistent Roseoloviruses Infection in Adult Patients with Epilepsy. Brain Sciences. 2020; 10(5):287. https://doi.org/10.3390/brainsci10050287
Chicago/Turabian StyleRasa-Dzelzkaleja, Santa, Sabine Gravelsina, Svetlana Chapenko, Zaiga-Nora Krukle, Simons Svirskis, Normunds Suna, Elena Kashuba, Guntis Karelis, and Modra Murovska. 2020. "Persistent Roseoloviruses Infection in Adult Patients with Epilepsy" Brain Sciences 10, no. 5: 287. https://doi.org/10.3390/brainsci10050287
APA StyleRasa-Dzelzkaleja, S., Gravelsina, S., Chapenko, S., Krukle, Z.-N., Svirskis, S., Suna, N., Kashuba, E., Karelis, G., & Murovska, M. (2020). Persistent Roseoloviruses Infection in Adult Patients with Epilepsy. Brain Sciences, 10(5), 287. https://doi.org/10.3390/brainsci10050287