1. Introduction
Many fundamental principles of mammalian brain development were derived from studies of neocortical development. Early during neocortical brain development, neuroepithelial cells in the ventricular zone divide symmetrically to expand the precursor population [
1]. Then, neuroepithelial cells transform into radial glia cells, the neural stem cells of the developing brain, which divide asymmetrically to generate differentiated progeny [
2]. Radial glia cells first generate neurons and then undergo a fate switch to generate glia [
3,
4]. This neurogenic-to-gliogenic switch in neocortical radial glia cells is coordinated at the population level such that neurogenesis occurs from embryonic day 12 (E12) in mice until late embryonic development, and gliogenesis occurs from late embryonic development into early postnatal development. Once radial glia cells complete gliogenesis, they transform into astrocytes and lose their neural stem cell properties [
4]. As a result, neurogenesis and gliogenesis occur as sequential and largely nonoverlapping processes during neocortical development [
5]. The timing and mechanisms regulating the neurogenic-to-gliogenic switch in the neocortex is well-studied [
5]. However, it remains unclear whether principles of neocortical development directly apply to non-neocortical regions of the cerebral cortex, such as the archicortex or hippocampus, of which development has been studied less extensively.
Thymidine analog methods were used in developmental biology research for decades to assay for proliferation and to determine the timing of cytogenesis across development [
6,
7,
8,
9,
10,
11]. Thymidine analogs incorporate into the DNA sequence during the S-phase of the cell cycle and can be used to assay proliferation at different time points after administration. Once incorporated into the DNA, the thymidine analog content gets diluted over subsequent cell divisions. It was estimated that 2–4 cell divisions dilute the thymidine analog content to undetectable levels [
10,
11]. However, if a cell is undergoing its final division before differentiating when it incorporates a thymidine analog, then it retains the thymidine analog label, which birth-dates the cell. Original birth-dating studies used a radioactive thymidine analog (
3H-thymidine) to label dividing cells, which could be detected by autoradiography [
11]. The second generation of thymidine analogs included 5′-bromo-2′-deoxyuridine (BrdU), 5′-iodo-2′-deoxyuridine (IdU) and 5′-chloro-2′deoxyuridine (CldU), which can be detected by antibody immunohistochemistry [
7,
8,
9]. However, antibody detection of BrdU, IdU and CldU requires a caustic acid antigen retrieval step, which ruins the epitopes of many other proteins, making colocalization of cell type-specific markers challenging. The third generation of thymidine analogs, 5-ethynyl-2′-deoxyuridine (EdU), is widely used today and was modified to be compatible with Click chemistry, which utilizes a biochemical reaction to detect the EdU labeling and does not require a caustic acid antigen retrieval step [
6].
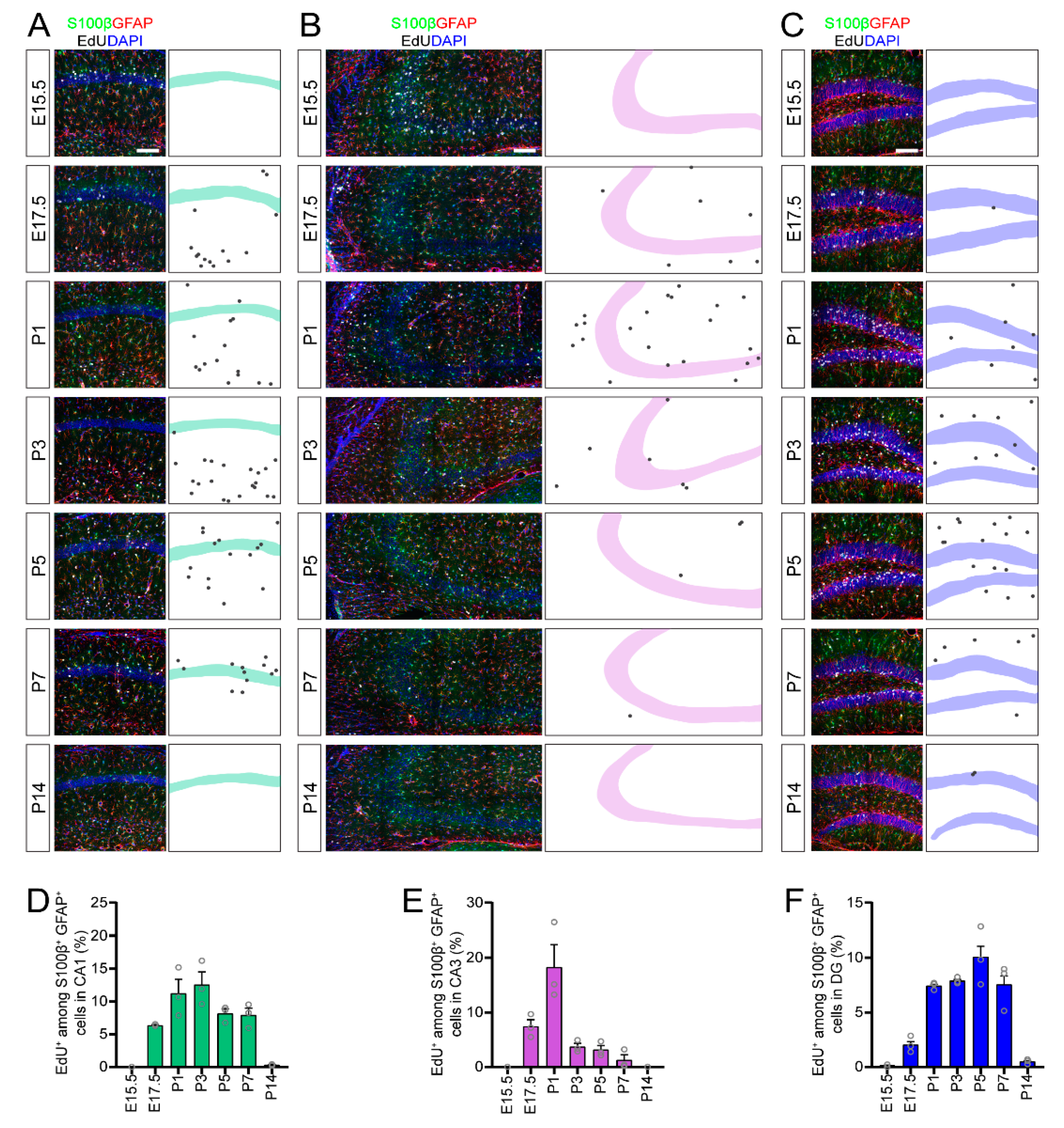
The hippocampus can be subdivided into three regions, the Cornu Ammonis (CA) 1, the CA3 and the dentate gyrus (DG). Seminal studies by Bayer [
12,
13,
14], Angevine [
15] and colleagues used
3H-thymidine autoradiography to study the timing of neurogenesis in the developing hippocampus of rat and mouse, respectively. Collectively, these studies suggested that neurons in the CA1 and CA3 regions are generated embryonically until birth and that neurons in the DG have an extended generation time which continues postnatally. In fact, the DG maintains neural stem cells beyond development, which sustain neurogenesis throughout adulthood [
16]. Multiple studies suggested that astrocyte generation in the hippocampus occurs during early postnatal development in rodents [
17,
18,
19]. However, birth-dating techniques have never been used to directly determine the dynamics of astrocyte generation in the hippocampus and its subregions. Clonal lineage tracing studies showed that, during development, a single neural stem cell in the CA1 [
20] or DG [
21] germinal zone could generate both neurons and astrocytes for the corresponding region. However, the extended development of the DG raises the question of how neurogenesis and gliogenesis are coordinated within DG.
Here, we used thymidine-analog EdU birth-dating to investigate the timing and coordination of neurogenesis and astrogenesis in different subregions in the developing mouse hippocampus. Our results indicated that neurogenesis and astrogenesis in the CA1 and CA3 regions occur sequentially as nonoverlapping processes, similar to the neocortex, while astrogenesis in the DG completely overlaps with continuous neurogenesis in this subregion. In addition, we uncovered differential timelines of peak astrogenesis in each of the hippocampal subregions. Collectively, our results suggested that coordination of neurogenesis and gliogenesis in the DG does not mirror neocortical development and likely follows a distinct set of developmental principles.
2. Materials and Methods
2.1. Animals
All animal procedures used in this study were performed in accordance with protocols approved by the Institutional Animal Care and Use Committee of the University of Pennsylvania Perelman School of Medicine. The title of the protocol was “Molecular and cellular mechanisms regulating neural stem cells and neurogenesis”. The approval data was 2 May 2017. All mice used in this study were maintained on a C57BL/6 background. Animals were housed in a 14 h light/10 h dark cycle with food and water ad libitum. Both male and female mice were used for all experiments.
2.2. EdU Injection
EdU was administered as previously described [
21]. A stock solution of 10 mg/mL EdU (Sigma, St. Louis, MO, USA, Cat. #900584) was prepared in normal saline solution (0.9%). EdU (50 mg/kg) was injected 2 times at a 6 h interval intraperitoneally into the pregnant female for embryonic time points (E12.5, E15.5, E17.5) or the pups for postnatal time points (P1, P3, P5, P7, P14), then brains were all analyzed at P30. For embryonic time points, the timed pregnancy was determined by identifying a vaginal plug (E0.5), and then EdU was administered intraperitoneally to the pregnant females on the target embryonic day. Samples from each time point came from different litters, because EdU injections into pregnant females affect the entire litter.
2.3. Tissue Processing, Click-iT EdU Labeling and Immunohistology
Animals were transcardially perfused with ice-cold Dulbecco’s phosphate-buffered saline (DPBS), followed by ice-cold 4% paraformaldehyde (PFA). Brains were fixed overnight in 4% PFA at 4 °C, and then cryoprotected in 30% sucrose solution overnight at 4 °C. Coronal brain sections (45 µm) were sliced using a sliding microtome (Leica, SM2010R). Every sixth section of each brain was collected into 1 of 6 wells in a 24-well plate containing anti-freeze solution (300 g sucrose, 300 mL ethylene glycol, 500 mL 0.1M PBS), and sections were stored at −20 °C.
Brain sections were stained as floating sections. Click-iT EdU labeling was performed according to manufacturer’s guidelines (Click-iT EdU Alexa Fluor 647 Imaging Kit, ThermoFisher Scientific, Cat. #C10340) prior to immunohistology. Brains were then washed in Tris-buffered saline (TBS) with 0.05% TritonX-100 and incubated in primary antibody solution (3.33% donkey serum and 0.05% TritonX-100 in TBS) overnight at 4 °C. Primary antibodies and dilutions used in this study included Rabbit anti-Prox1 (Abcam, Cat. #ab37128, 1:500 dilution), Mouse anti-neuronal nuclear protein (NeuN) Alexa Fluor 488 conjugated (Millipore, Cat. #MAB377X, 1:500 dilution), Rabbit anti-S100β (Agilent, Cat. #Z0311, 1:500 dilution) and Mouse anti-glial fibrillary acidic protein (GFAP) (Millipore, Cat. #MAB360, 1:1000 dilution). Brain sections were washed in tris-buffered saline (TBS) with 0.05% TritonX-100 and then incubated in secondary antibody solution (3.33% donkey serum and 0.05% TritonX-100 in TBS) and 4′,6-diamidino-2-phenylindole (DAPI) nuclear stain (Roche, 1:1000 dilution) for 1–2 h at room temperature. Alexa Fluor 488 and 555 secondary antibodies (Invitrogen, 1:250 dilution) were used. After a second set of washes, sections were mounted with 2.5% polyvinyl alcohol mounting medium with DABCO® mounting media (Sigma, St. Louis, MO, USA, Cat. #10981).
2.4. Confocal Microscopy and Image Quantification
Brain sections across the anterior–posterior axis of the hippocampus were imaged as tiled z-stacks using a Zeiss LSM 810 confocal microscope (Carl Zeiss). Each tiled z-stack encompassed the entire hippocampus of a single brain section, and 3 anatomically matched brain sections were selected at equivalent levels across the dorso-ventral axis of the hippocampus and imaged for every sample. Either 20× or 40× objectives were used for imaging. Cells were considered positive for EdU when EdU label intensity was above background levels, regardless of the staining pattern within the nucleus. Images were analyzed and quantified using ImageJ software, and all cell counting was done blind to the timing of the EdU injection to ensure unbiased quantification. Quantification of the EdU
+ cells generated at a particular developmental time point as a proportion of the total population of that cell type was performed in line with previous methodology from classic birth-dating studies [
12,
13,
14]. Neurogenesis was quantified by counting the number of NeuN
+EdU
+ cells as a proportion of the total number of NeuN
+ cells (CA1 and CA3), or by counting the number of NeuN
+ prospero homeobox 1 (Prox1)
+EdU
+ cells as a proportion of the total number of NeuN
+Prox1
+ cells (DG). Astrogenesis was quantified by counting the number of GFAP
+S100 calcium-binding protein B
+(S100β
+)EdU
+ cells as a proportion of the total number of GFAP
+S100β
+ cells in a given region or subregion. For quantification, EdU cell labeling within animals and between animals was found to be very consistent throughout the quantitative analysis, as evidenced by consistent patterns of EdU labeling between regions within the same brain and low variability within groups.
4. Discussion
Our study used an EdU pulse-chase birth-dating technique to reveal the temporal dynamics of neurogenesis and astrogenesis in the developing mouse hippocampus and uncovered regional differences in the timing and coordination of neuron and astrocyte generation during embryonic and early postnatal hippocampal development. While many fundamental developmental processes were studied extensively in the neocortex, little is known about how neurogenesis and astrogenesis occur in the developing mammalian hippocampus. Our findings provide a comprehensive resource outlining the timing of developmental neurogenesis and astrogenesis in hippocampal subregions. In addition, by comparing the timing of neurogenesis and astrogenesis in the same experiment, we could directly study the coordination of these developmental processes, unlike many previous studies which focused individually on neurogenesis or astrogenesis [
12,
13,
14,
15,
17,
18,
19]. Our findings reveal that the timing of peak astrogenesis during early postnatal development differs by specific hippocampal subregions (
Figure 6). In addition, neurogenesis and astrogenesis in the CA1 and CA3 regions occur sequentially, similar to the neocortex. In contrast, astrogenesis completely overlaps with the extended neurogenesis that occurs in the DG, such that developmental neurogenesis and astrogenesis occur simultaneously. Together, our results uncover new insights into hippocampal astrocyte generation and suggest a unique coordination of neurogenesis and astrogenesis in the DG region of the hippocampus.
By mapping a time course of cytogenesis that spans both embryonic and postnatal development, our study could not use littermate comparisons between time points. EdU cannot practically be administered to individual embryos in utero, but instead must be administered to the entire litter by EdU administration to the dam. As a result, we could not administer EdU to littermates at different developmental timepoints. This technical constraint could potentially introduce some limitations to our study. First, it is possible that litter-to-litter variation could contribute to observed differences in cytogenesis. Second, it is possible that differences in the pharmacokinetics of EdU in embryos compared to postnatal pups could also contribute to the observed differences in cytogenesis. We used multiple high doses of EdU to achieve a saturating dose, which would be less vulnerable to subtle differences in pup-rearing between litters, and EdU pharmacokinetics between the in utero versus postnatal ages. Because we were interested in the temporal dynamics of neurogenesis and astrogenesis, rather than in quantitative measures of cell genesis, we believe that the limitations of out experimental paradigm did not greatly impact the conclusions of our study.
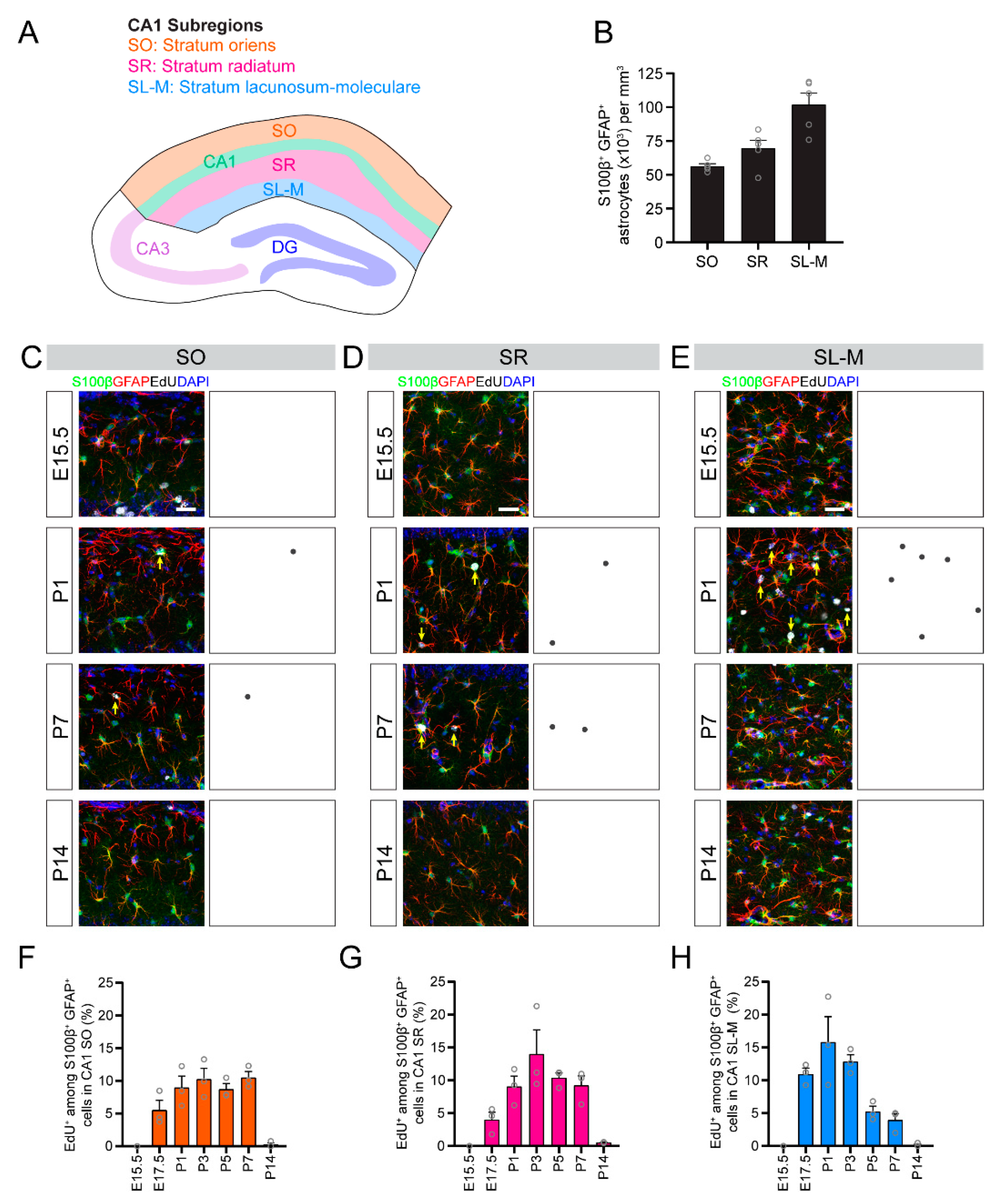
Though previous studies approximated the timing of astrogenesis in the hippocampus based on the appearance of stellate astrocytes during development [
17,
18,
19], our study used EdU incorporation and molecular markers to directly birth-date astrocytes in the hippocampus. As a result, we provided a comprehensive timeline of astrogenesis in each hippocampal subregion and revealed fundamental properties of hippocampal astrocyte generation. First, our findings suggested that astrocytes are generated in a region-specific order, such that astrogenesis first begins in the CA3 region, followed by the CA1 and DG regions (
Figure 6A). In addition, subregions within the CA1 and DG regions follow distinct temporal patterns of astrogenesis (
Figure 6B). For example, astrocytes in the stratum lacunosum-moleculare are generated earlier than astrocytes in the stratum radiatum of the CA1 region, and astrocytes in the hilus are generated earlier than astrocytes in the molecular layer of the DG region. Little is known regarding how astrocytes are generated during hippocampal development and, as a result, fundamental principles of astrogenesis in the hippocampus remain unknown. Previous work showed that spatially segregated germinal zones in the medial pallium give rise to neurons in the CA1, CA3 or DG hippocampal subregions, suggesting that NSCs in these distinct germinal zones are regionally specified [
21,
22]. However, it is unknown whether a single precursor can generate astrocytes for multiple hippocampal subregions or if astrocyte precursors are restricted to a single hippocampal subregion. In addition, though it is assumed that neural precursors that give rise to neurons in the CA1, CA3 or DG subregions also give rise to astrocytes in those regions, the source of hippocampal astrocytes was not explicitly identified. Our data suggested that there are regional differences in the timing of astrogenesis in the hippocampus, which could be a result of the source of astrocyte precursors, the timing of the neurogenic-to-astrogenic switch or the astrocytic requirements of different hippocampal subregions. Future studies should explore the basic principles of astrocyte generation in the hippocampus and how astrocyte generation is coordinated between hippocampal subregions.
By comparing neuron and astrocyte generation using the same methodology in a single experiment, our study captured the coordination of neurogenesis and astrogenesis within each hippocampal subregion. Much of what we know about the coordination of neurogenesis and astrogenesis comes from studies of neocortical development, in which neurogenesis occurs embryonically, followed sequentially by astrogenesis during early postnatal development [
3,
4]. Our data demonstrate that these fundamental properties of neocortical development apply to the CA1 and CA3 regions of the hippocampus, where we see a similar coordination and timing of neurogenesis and astrogenesis. Strikingly, development of the DG region of the hippocampus does not follow many principles of neocortical development. First, we found that neurogenesis in the DG is a gradual and protracted process. Though the onset of neurogenesis in the DG occurs during early embryonic development, similar to the neocortex and CA regions, DG neurogenesis occurs more gradually over a longer period of time that extends into postnatal life. In contrast, neurogenesis in the neocortex and CA regions occurs more rapidly and is completed by birth. Second, we found that neurogenesis and astrogenesis occur simultaneously during DG development. The majority of astrogenesis in the DG occurs during the first postnatal week, similar to that of the neocortex and CA regions of the hippocampus. However, unlike the neocortex and CA regions, in which neurogenesis and astrogenesis are temporally segregated, neurogenesis in the DG continues through and beyond the postnatal period of astrogenesis (
Figure 6A). As a result, there does not appear to be a population-wide, neurogenic-to-gliogenic switch in the DG. Throughout the stages, neurogenesis overwhelms astrogenesis, even during the peak of astrogenesis in the DG. Interestingly, active neurogenesis and astrogenesis within DG occurs concurrently with the transition of neural stem cells into quiescence [
21] (
Figure 6A). Together, these findings prompt a multitude of questions. Does a subpopulation of neural stem cells undergo a neurogenic-to-gliogenic switch to generate astrocytes, while another subpopulation continues engaging in neurogenesis postnatally? Does the neurogenic-to-gliogenic switch occur at the single cell level or do neural precursors in the DG remain multipotent during the overlap of neurogenesis and astrogenesis? What mechanisms initiate and terminate developmental astrocyte generation in the DG? The answer to some of these questions may be intertwined with the fact that the DG maintains neural stem cells beyond development, which sustain neurogenesis throughout adulthood (
Figure 6) [
16,
21]. Future research to uncover the basic properties of DG development, which seem to defy conventional principles of developmental neurobiology, would broaden our understanding of mammalian brain development.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





