

Self-Induced Crystalline Morphology at the Mineral–Fluid Interface: Silica–Carbonate Biomorphs of Alkaline Earth Metals as a Case Study

 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Composition of Obsidian
2.2. Synthesis of Biomorphs
2.2.1. Synthesis of Control Biomorphs
2.2.2. Synthesis of Biomorphs on Obsidian
2.3. Characterization of the Biomorphs
- (a)
- Scanning electron microscopy (SEM)
- (b)
- Raman spectroscopy
- (c)
- Fourier transform infrared spectroscopy (FTIR)
- (d)
- Powder X-Ray diffraction (PXRD)
3. Results and Discussion
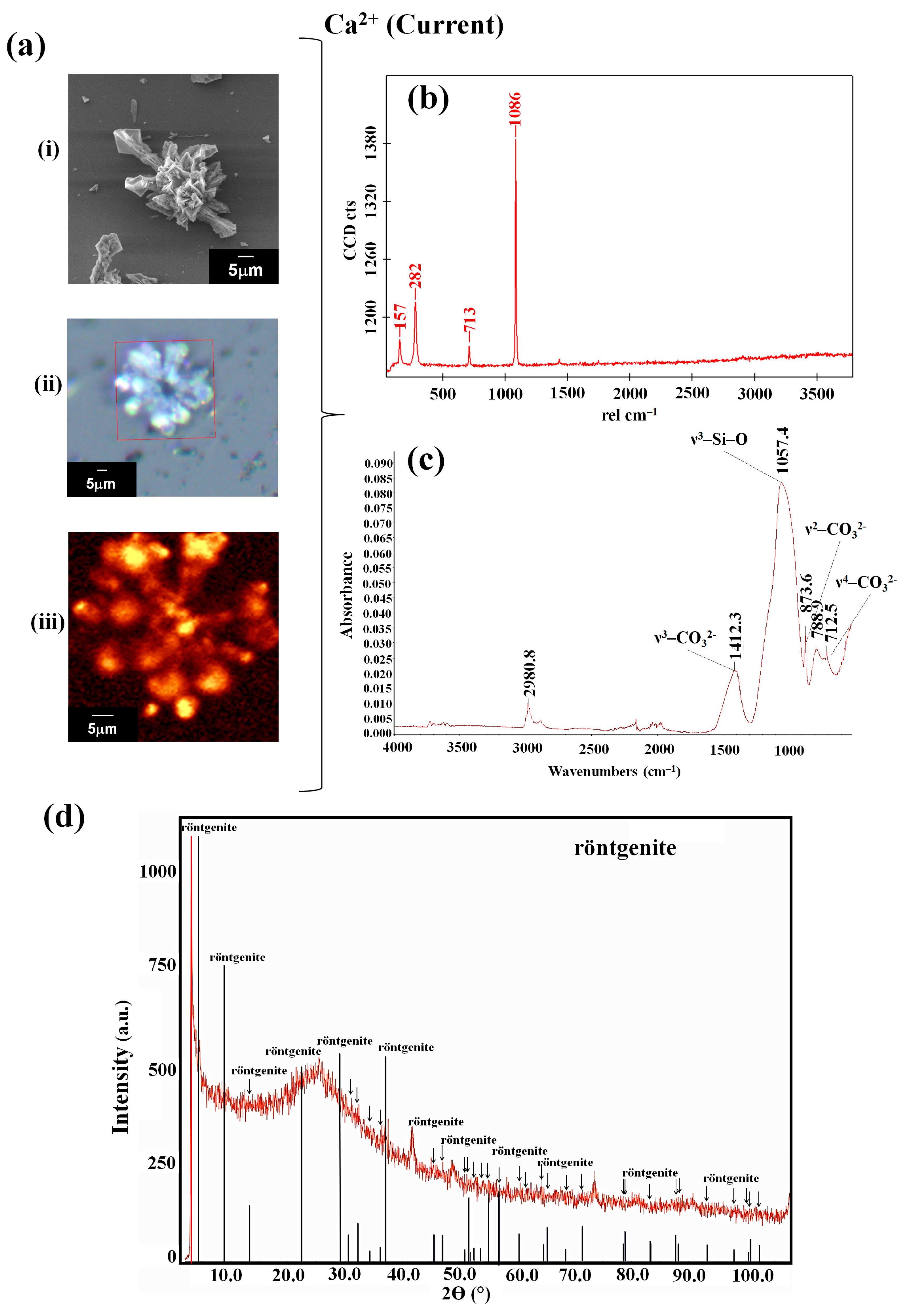
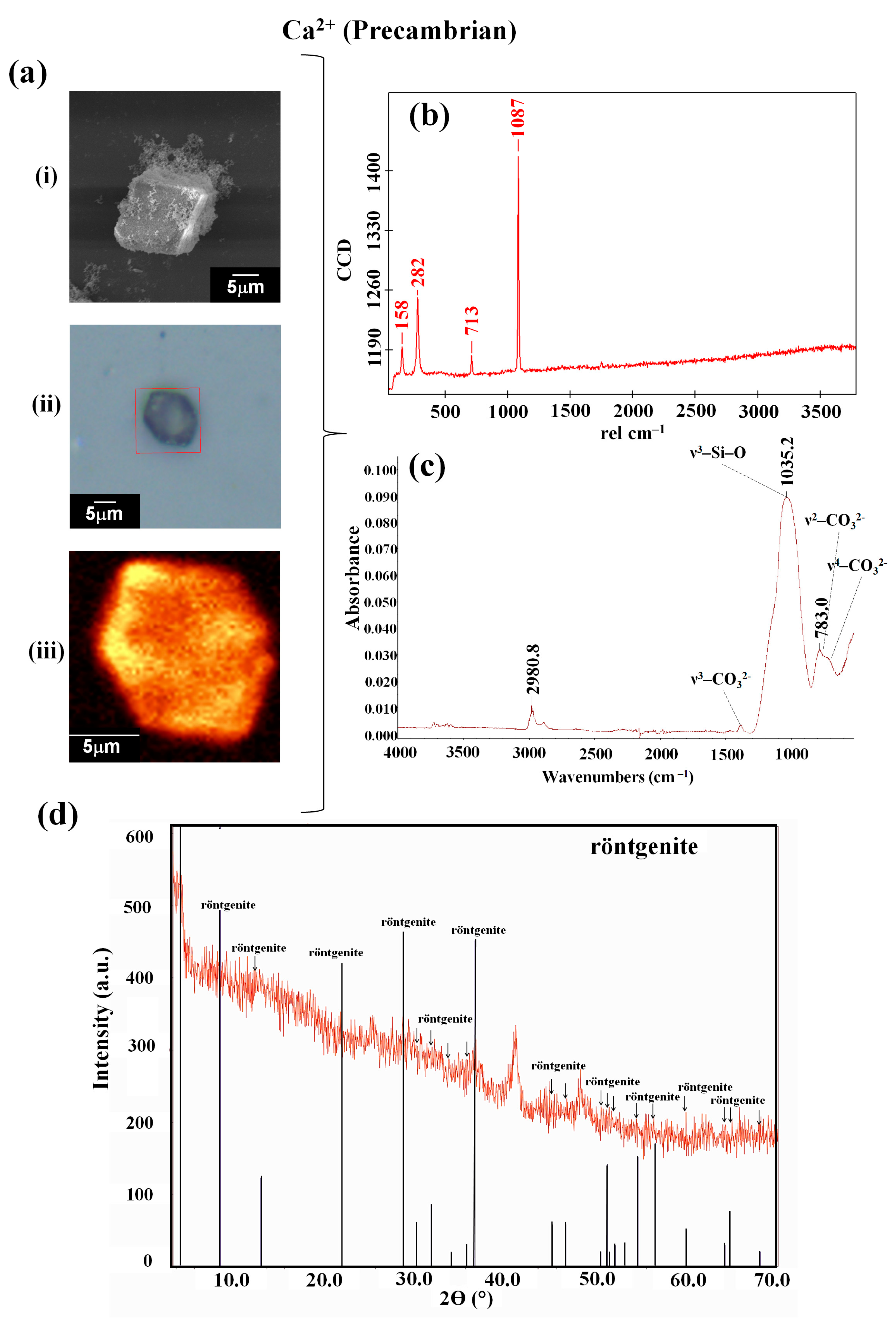
3.1. The Interaction of Obsidian with the Reaction Mixture Favored the Synthesis of Biomorphs with a Crystalline Phase of the Röntgenite Polymorph
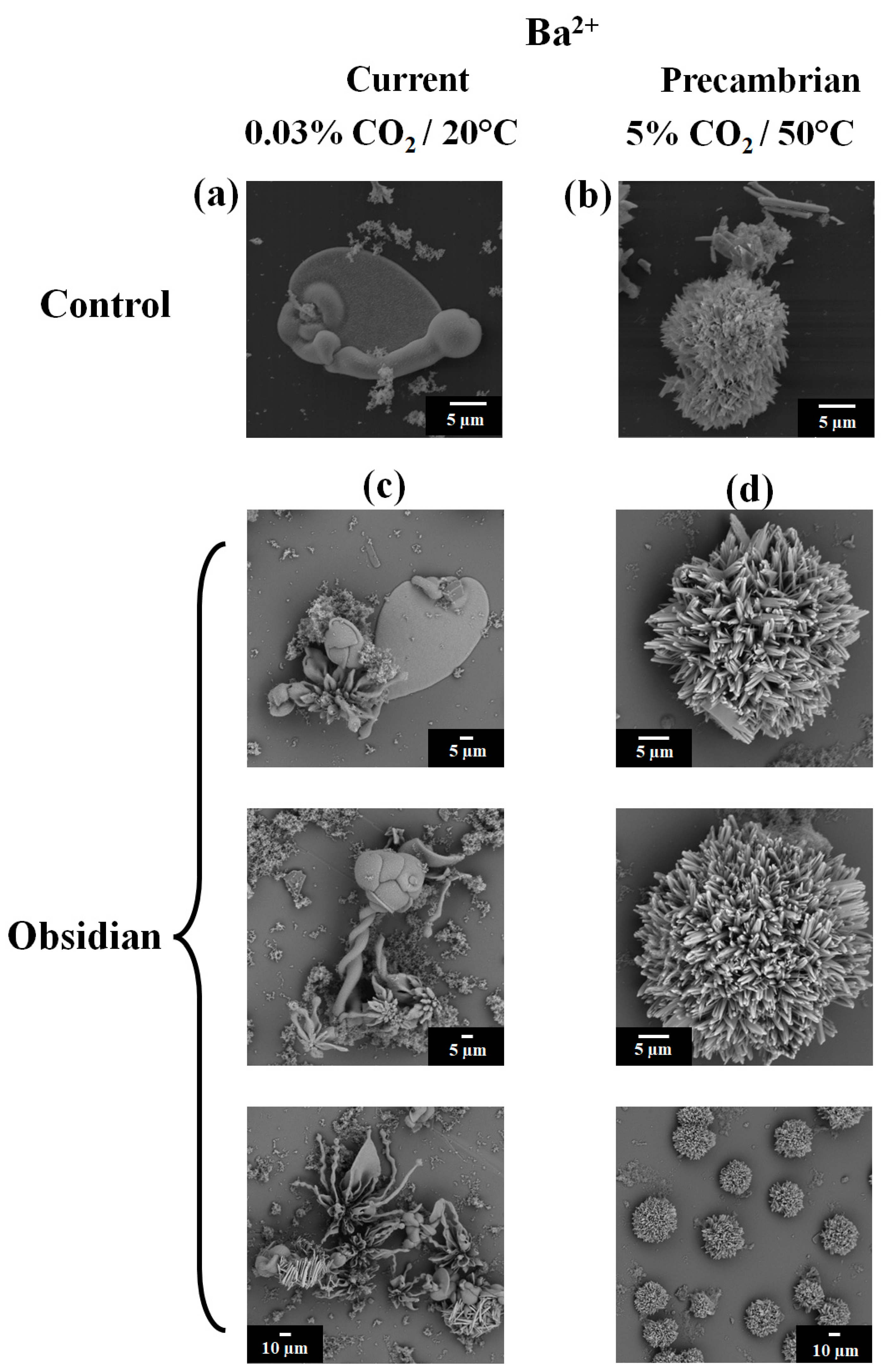
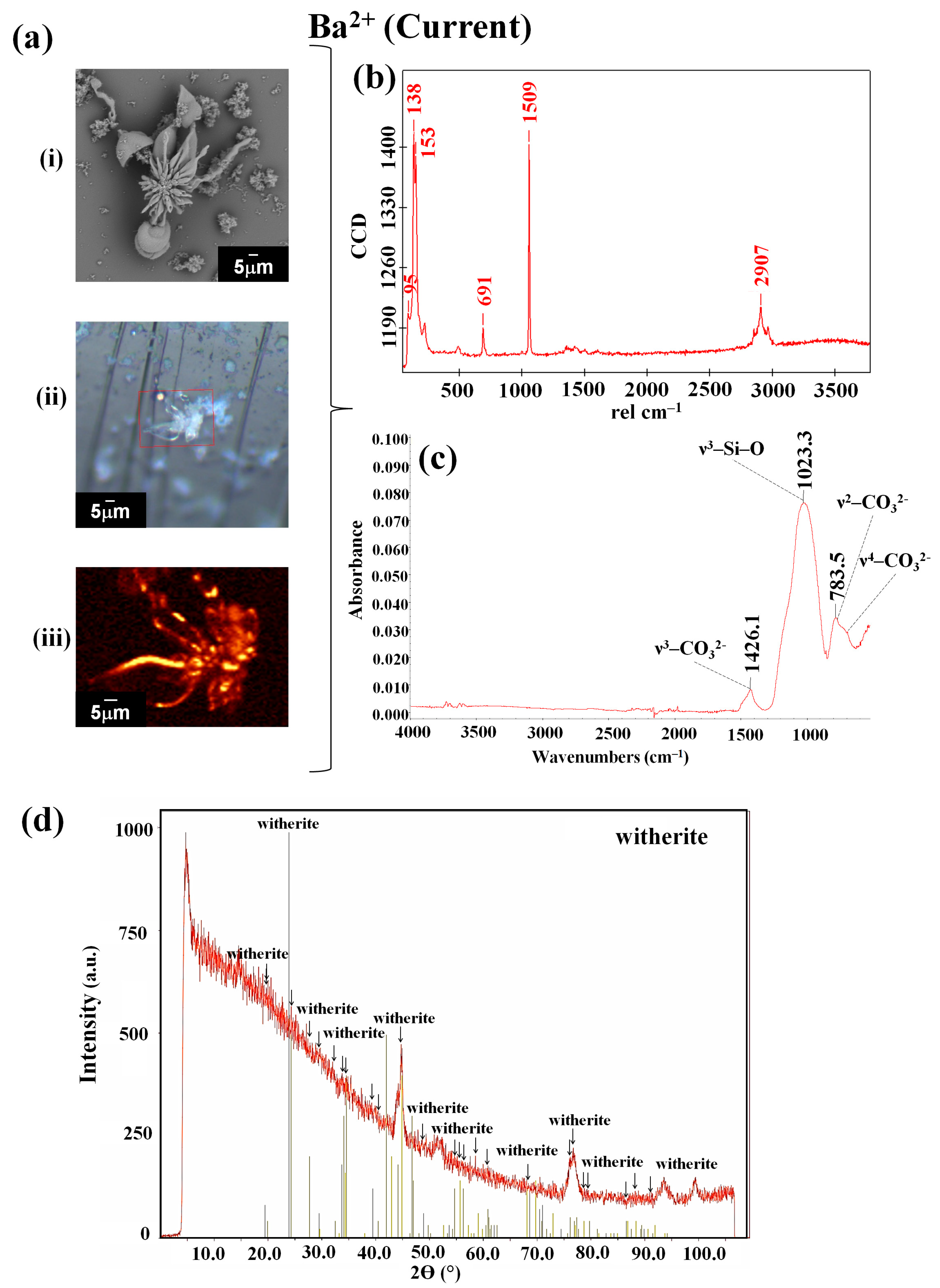
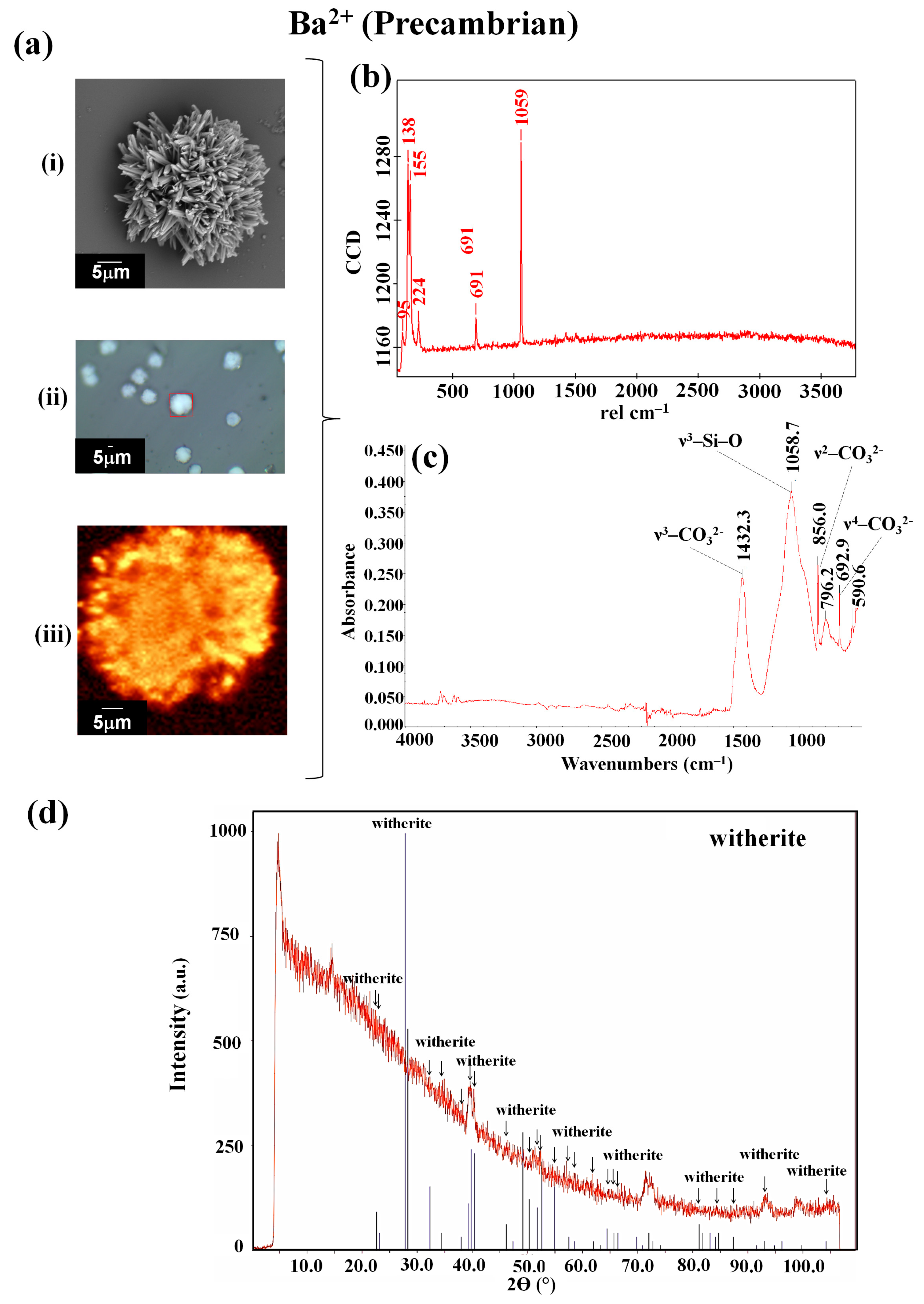
3.2. Barium Silico-Carbonate Biomorphs Are Possibly the First Inorganic Structure That Formed Parts of Various Forms of Life in the Precambrian
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hazen, R.M.; Papineau, D.; Bleeker, W.; Downs, R.T.; Ferry, J.M.; McCoy, T.J.; Sverjensky, D.A.; Yang, H. Mineral evolution. Am. Miner. 2008, 93, 1693–1720. [Google Scholar] [CrossRef]
- Ringwood, A.E. Chemical evolution of the terrestrial planets. Geochim. Cosmochim. Acta 1966, 30, 41–104. [Google Scholar] [CrossRef]
- Ringwood, A.E. On the chemical evolution and densities of the planets. Geochim. Cosmochim. Acta 1959, 15, 257–283. [Google Scholar] [CrossRef]
- Scott, E.R.D.; Krot, A.N. Chondrites and Their Components. In Treatise on Geochemistry; Elsevier Ebooks: Amsterdam, The Netherlands, 2003; pp. 1–72. [Google Scholar]
- Skinner, B.J. Earth resources. Proc. Natl. Acad. Sci. USA 1979, 76, 4212–4217. [Google Scholar] [CrossRef]
- Monger, H.C.; Kelly, E.F. Silica Minerals; Soil Science Society of America Book Series; Wiley: Hoboken, NJ, USA, 2018; pp. 611–636. [Google Scholar]
- Huang, P.M. Feldspars, Olivines, Pyroxenes, and Amphiboles; Soil Science Society of America Book Series; Wiley: Hoboken, NJ, USA, 2018; pp. 975–1050. [Google Scholar]
- Drees, L.R.; Wilding, L.P.; Smeck, N.E.; Senkayi, A.L. Silica in Soils: Quartz and Disordered Silica Polymorphs; Soil Science Society of America Book Series; Wiley: Hoboken, NJ, USA, 2018; pp. 913–974. [Google Scholar]
- Ericson, J.E.; Makishima, A.; Mackenzie, J.D.; Berger, R. Chemical and physical properties of obsidian: A naturally occuring glass. J. Non-Cryst. Solids 2018, 17, 129–142. [Google Scholar] [CrossRef]
- Glascock, M.D. Obsidian Provenance Research in the Americas. Acc. Chem. Res. 2002, 35, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Fuller, R.E. The mode of origin of the color of certain varicolored obsidians. J. Geol. 1927, 35, 570–573. [Google Scholar] [CrossRef]
- Cann, J.; Renfrew, C. The Characterization of Obsidian and its Application to the Mediterranean Region. Proc. Phys. Soc. 1964, 30, 111–133. [Google Scholar] [CrossRef]
- Carballo, D.M.; Carballo, J.; Neff, H. Formative and Classic Period Obsidian Procurement in Central Mexico: A Compositional Study Using Laser Ablation-Inductively Coupled Plasma-Mass Spectrometry. Lat. Am. Antiq. 2007, 18, 27–43. [Google Scholar] [CrossRef]
- Charlton, T.H.; Grove, D.C.; Hopke, P.K. El Paredón Mexico, Obsidian Source and Early Formative Exchange. Science 1978, 201, 807–809. [Google Scholar] [CrossRef]
- Hazen, R.M.; Sverjensky, D.A. Mineral Surfaces, Geochemical Complexities, and the Origins of Life. Cold Spring Harb. Perspect. Biol. 2010, 2, a002162. [Google Scholar] [CrossRef]
- Mary, A.; Mary, A. Observations sur la morphogénèse en Plasmologie. Memoires Soc. Sci. “Antonio Alzate” 1909, 29, 242–250. [Google Scholar]
- Lillie, R.S.; Johnston, E.N. Precipitating-structures simulating organic growth. II. A Contribution to the Physico-Chemical Analysis of Growth and Heredity. Biol. Bull. 1919, 36, 225–272. [Google Scholar] [CrossRef]
- Ober, P.; Kolbinger, S.H.; Backus, E.H.G.; Bonn, M. Ion-Specific Interactions at a Mineral-Water Interface Revealed by Surface-Sensitive Spectroscopy under Flow Conditions. J. Phys. Chem. C 2023, 127, 13005–13010. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M. The equation of life in the Universe: Biomorphs as reminiscence of the first forms of life. Prog. Cryst. Growth Charact. Mater. 2024, 70, 100624. [Google Scholar] [CrossRef]
- González-Ramírez, L.A.; Moreno, A.; Ng, J.D.; García-Ruiz, J.M.; González-Ramírez, L.A.; Moreno, A.; Ng, J.D.; García-Ruiz, J.M. Investigations on the role of iron (III) and silica-iron (III) for the DNA protection against highly intense UV radiation: Tracking the connection of prebiotic chemistry to biology. Astrobiology 2023, 23, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Cuéllar-Cruz, M. New insights on the origin of life: The role of silico-carbonates of Ba (II) to preserve DNA against highly intense UV radiation. ACS Omega 2023, 8, 29585–29594. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Islas, S.R.; González, G.; Moreno, A. Influence of nucleic acids on the synthesis of crystalline Ca (II), Ba (II), and Sr (II) silica-carbonate biomorphs: Implications for the chemical origin of life on primitive Earth. Cryst. Growth Des. 2019, 19, 4667–4682. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Moreno, A. The role of calcium and strontium as the most dominant elements during combinations of different alkaline Earth metals in the synthesis of crystalline silica-carbonate biomorphs. Crystals 2019, 9, 381. [Google Scholar] [CrossRef]
- Bittarello, E.; Aquilano, D. Self-assembled nanocrystals of barium carbonate in biomineral-like structures. Eur. J. Mineral. 2017, 19, 345–351. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Moreno, A. Synthesis of crystalline silica-carbonate biomorphs of Ba (II) under the presence of RNA and positively- and negatively charged ITO electrodes: Obtainment of graphite via bio-reduction of CO2 and its implications to the chemical origin of life on primitive Earth. ACS Omega 2020, 5, 5460–5469. [Google Scholar] [CrossRef]
- Islas, S.R.; Cuéllar-Cruz, M. Silica-carbonate of Ba(II) and Fe2+/Fe3+ Complex as Study Models to Understand Prebiotic Chemistry. ACS Omega 2021, 6, 35629–35640. [Google Scholar] [CrossRef]
- Garcia-Ruiz, J.M.; Melero-Garcia, E.; Hyde, S.T. Morphogenesis of Self-Assembled Nanocrystalline Materials of Barium Carbonate and Silica. Science 2009, 323, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Eggins, S.M.; Woodhead, J.D.; Kinsley, L.P.J.; Mortimer, G.E.; Sylvester, P.; McCulloch, M.T.; Hergt, J.M.; Handler, M.R. A Simple Method for the Precise Determination of >=40 Trace Elements in Geological Samples by ICP-MS Using Enriched Isotope Internal Standardization. Chem. Geol. 1997, 134, 311–326. [Google Scholar] [CrossRef]
- Chaussidon, R.F.; M. A palaeotemperature curve for the Precambrian oceans based on silicon isotopes in cherts. Nature 2006, 443, 969–972. [Google Scholar] [CrossRef]
- Kasting, J.F.; Howard, M.T. Atmospheric composition and climate on the early Earth. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1733–1741; discussion 1741–1742. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Ramírez-Cardona, M.; Moreno, A. Influence of different types of clay minerals on the shape and form of silica-carbonates (Biomorphs) of Ca(II), Ba(II), and Sr(II). ACS Earth Space Chem. 2022, 6, 3054–3065. [Google Scholar] [CrossRef]
- Lilli, M.A.; Nikolaidis, Ν.P.; Karatzas, G.P.; Kalogerakis, N. Identifying the controlling mechanism of geogenic origin chromium release in soils. J. Hazard. Mater. 2019, 366, 169–176. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Schneider, D.K.; Stojanoff, V.; Islas, S.R.; Sánchez-Puig, N.; Arreguín-Espinosa, R.; Delgado, J.M.S.; Moreno, A. Formation of Crystalline Silica–Carbonate Biomorphs of Alkaline Earth Metals (Ca, Ba, Sr) from Ambient to Low Temperatures: Chemical Implications during the Primitive Earth’s Life. Cryst. Growth Des. 2019, 20, 1186–1195. [Google Scholar] [CrossRef]
- Sánchez-Puig, N.; Cuéllar-Cruz, M.; Islas, S.R.; Tapia-Vieyra, J.V.; Arreguín-Espinosa, R.A.; Moreno, A. The influence of silicateins on the shape and crystalline habit of silica carbonate biomorphs of alkaline Earth metals (Ca, Ba, Sr). Crystals 2021, 11, 438. [Google Scholar] [CrossRef]
- Opel, J.; Wimmer, F.P.; Kellermeier, M.; Colfen, H. Functionalisation of silica-carbonate biomorphs. Nanoscale Horiz. 2016, 1, 144–149. [Google Scholar] [CrossRef]
- Voinescu, A.E.; Touraud, D.; Lecker, A.; Pfitzner, A.; Kunz, W.; Ninham, B.W. Mineralization of CaCO3 in the presence of egg White lysozyme. Langmuir 2007, 23, 12269–12274. [Google Scholar] [CrossRef]
- Frampton, M.B.; Zelisko, P.M. Organosilicon Biotechnology. Silicon 2009, 1, 147–163. [Google Scholar] [CrossRef]
- Noorduin, W.L.; Grinthal, A.; Mahadevan, L.; Aizenberg, J. Rationally Designed Complex, Hierarchical Microarchitectures. Science 2013, 340, 832–837. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Zhang, J.; Wang, L.; Lu, Z.; Wang, F.; Liu, Z.; Zeng, H. Spontaneous hierarchical surface engineering of minerals through coupled dissolution-precipitation chemistry. Aggregate 2023, 5, e452. [Google Scholar] [CrossRef]
- Buzgar, N.; Apopei, A.I. The Raman study of certain carbonates. Geol. Tomul L 2009, 55, 97–112. [Google Scholar]
- Kasatkin, A.V.; Škoda, R.; Nestola, F.; Kuznetsov, A.M.; Belogub, E.V.; Agakhanov, A.A. Röentgenite-(Ce) and other REE fluorcarbonates from vein no. 35, Vishnevye Mountains, southern Urals. Mineralogija 2019, 5, 10–22. [Google Scholar] [CrossRef]
- Lin, C.C.; Liu, L.G. High-pressure Raman spectroscopic study of post-aragonite phase transition in witherite (BaCO3). Eur. J. Miner. 1997, 9, 785–792. [Google Scholar] [CrossRef]
- Morse, J.W.; Mackenzie, F.T. Chapter 2 Interactions Between Carbonate Minerals and Solutions. Dev. Sedimentol. 1990, 48, 39–86. [Google Scholar] [CrossRef]
- Hunger, J.; Schaefer, J.; Ober, P.; Seki, T.; Wang, Y.; Prädel, L.; Nagata, Y.; Bonn, M.; Jan Bonthuis, D.J.; Backus, E.H.G. Nature of cations critically affects water at the negatively charged silica interface. J. Am. Chem. Soc. 2022, 144, 19726–19738. [Google Scholar] [CrossRef]
- Hazen, R.M. Genesis: Rocks, Minerals, and the Geochemical Origin of Life. Elements 2005, 1, 135–137. [Google Scholar] [CrossRef]
- Benetoli, L.O.B.; De Souza, C.M.D.; Da Silva, K.L.; De Souza, I.G.; De Santana, H.; Paesano, A.; Da Costa, A.C.S.; Zaia, C.T.B.V.; Zaia, D.A.M. Amino Acid Interaction with and Adsorption on Clays: FT-IR and Mössbauer Spectroscopy and X-ray Diffractometry Investigations. Orig. Life Evol. Biosph. 2007, 37, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Beltrami, G.; Martucci, A.; Pasti, L.; Chenet, T.; Ardit, M.; Gigli, L.; Cescon, M.; Suard, E. L-Lysine Amino Acid Adsorption on Zeolite L: A Combined Synchrotron, X-Ray and Neutron Diffraction Study. ChemistryOpen 2020, 9, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Villafanñe-Barajas, S.A.; Baú, J.P.T.; Colin-Garcia, M.; Negron-Mendoza, A.; Heredia-Barbero, A.; Pi-Puig, T.; Zaia, D.A.M. Salinity Effects on the Adsorption of Nucleic Acid Compounds on Na-Montmorillonite: A Prebiotic Chemistry Experiment. Orig. Life Evol. Biosph. 2018, 48, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Pitsch, S.; Eschenmoser, A.; Gedulin, B.; Hui, S.; Arrhenius, G. Mineral Induced Formation of Sugar Phosphates. Orig. Life Evol. Biosph. 1995, 25, 297–334. [Google Scholar] [CrossRef]
- Hartmann, M. Ordered Mesoporous Materials for Bioadsorption and Biocatalysis. Chem. Mater. 2005, 17, 4577–4593. [Google Scholar] [CrossRef]
- Hashizume, H. Adsorption of Nucleic Acid Bases, Ribose, and Phosphate by some Clay minerals. Life 2015, 5, 637–650. [Google Scholar] [CrossRef]
- Lin, R.; Wang, Y.; Li, X.; Liu, Y.; Zhao, Y. pH-Dependent Adsorption of Peptides on Montmorillonite for Resisting UV Irradiation. Life 2020, 10, 45. [Google Scholar] [CrossRef]
- Gillams, R.J.; Jia, T.Z. Mineral Surface-Templated Self-Assembling Systems: Case Studies from Nanoscience and Surface Science towards Origins of Life Research. Life 2018, 8, 10. [Google Scholar] [CrossRef]
- Erastova, V.; Degiacomi, M.T.; Fraser, D.G.; Greenwell, H.C. Mineral Surface Chemistry Control for Origin of Prebiotic Peptides. Nat. Commun. 2017, 8, 2033. [Google Scholar] [CrossRef]
- Kloprogge, J.T.; Hartman, H. Clays and the Origin of Life: The Experiments. Life 2022, 12, 259. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Trejo, D.; Villanueva-Barragan, P.S.; Zamudio-Ramirez, R.; Cervantes-de la Cruz, K.E.; Mejia-Luna, I.; Chacon-Baca, E.; Negron-Mendoza, A.; Ramos-Bernal, S.; Heredia-Barbero, A. Histidine Self-assembly and Stability on Mineral Surfaces as a Model of Prebiotic Chemical Evolution: An Experimental and Computational Approach. Orig. Life Evol. Biosph. 2021, 51, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Aguilar, C.D.; Cuéllar-Cruz, M. The formation of crystalline minerals and their role in the origin of life on Earth. Prog. Cryst. Growth Charact. Mater. 2022, 68, 100558. [Google Scholar] [CrossRef]
- Rickard, D.; Luther, G.W. Chemistry of Iron Sulfides. Chem. Rev. 2007, 107, 514–562. [Google Scholar] [CrossRef] [PubMed]
- Poulton, S.W.; Canfield, D.E. Ferruginous Conditions: A Dominant Feature of the Ocean through Earth’s History. Elements 2011, 7, 107–112. [Google Scholar] [CrossRef]
- Rasmussen, B. Filamentous microfossils in a 3,235-million-year-old volcanogenic massive sulphide deposit. Nature 2000, 405, 676–679. [Google Scholar] [CrossRef]
- Martin, W.; Baross, J.A.; Kelley, D.S.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef]
- Hazen, R.M.; Filley, T.R.; Goodfriend, G.A. Selective adsorption of L- and D-amino acids on calcite: Implications for biochemical homochirality. Proc. Natl. Acad. Sci. USA 2001, 98, 5487–5490. [Google Scholar] [CrossRef]
- Mendoza-Torres, E.; Cruz-Cataneda, J.; Negron-Mendoza, A.; Heredia, A. Computer and Experimental Simulation of Alloxazine Synthesis from Gamma Irradiation of Amino Acids on Iceland Spar: A Prebiotic Chemistry Perspective. J. Mol. Evol. 2020, 88, 284–291. [Google Scholar] [CrossRef]
- Schrenk, M.O.; Brazelton, W.J.; Lang, S.Q. Serpentinization, carbon, and deep life. Rev. Mineral. Geochem. 2013, 75, 575–606. [Google Scholar] [CrossRef]
- Sojo, V.; Herschy, B.; Whicher, A.; Camprubi, E.; Lane, N. The origin of life in alkaline hydrothermal vents. Astrobiology 2016, 16, 181–197. [Google Scholar] [CrossRef]
- Russell, M.J.; Hall, A.J.; Martin, W. Serpentinization as a source of energy at the origin of life. Geobiology 2010, 8, 355–371. [Google Scholar] [CrossRef]
- Russell, M.J.; Ponce, A. Six ‘Must-Have’ Minerals for Life´s Emergence: Olivine, Pyrrhotite, Bridgmanite, Serpentine, Fougerite and Mackinawite. Life 2020, 10, 291. [Google Scholar] [CrossRef] [PubMed]
- Swadling, J.B.; Coveney, P.V.; Greenwell, H.C. Stability of free and mineral-protected nucleic acids: Implications for the RNA world. Geochim. Cosmochim. Acta 2012, 83, 360–378. [Google Scholar] [CrossRef]
- Hartman, H. Photosynthesis and the Origin of Life. Orig. Life Evol. Biosph. 1998, 28, 515–521. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M. The equation of the origin of life in the Universe (Part II): The combination of chemical elements does not determine the emergence of life on Earth. Prog. Cryst. Growth Charact. Mater. 2024, 70, 100625. [Google Scholar] [CrossRef]
- Templeton, W.L.; Brown, V.M. The Relationship between the concentrations of calcium, strontium and strontium-90 in wild brown trout, Salmo-Trutta L. and the concentrations of the stable elements in some waters of the United-Kingdom, and the implications in radiological health studies. Air Water Pollut. 1964, 8, 49–75. [Google Scholar]
- Madejón, P. Barium. In Heavy Metals in Soils; Alloway, B., Ed.; Environmental Pollution; Springer: Dordrecht, The Netherlands, 2013; pp. 507–514. [Google Scholar]
- Lowenstam, H.A. Minerals formed by organisms. Science 1981, 211, 1126–1131. [Google Scholar] [CrossRef]
- Lowenstam, H.A.; Margulis, L. Evolutionary prerequisites for early phanerozoic calcareous skeletons. Biosystems 1980, 12, 27–41. [Google Scholar] [CrossRef]
- Ruiz-Arellano, R.; Moreno, A. Obtainment of Spherical-Shaped Calcite Crystals Induced by Intramineral Proteins Isolated from Eggshells of Ostrich and Emu. Cryst. Growth Des. 2014, 14, 5137–5143. [Google Scholar] [CrossRef]
- Ruiz-Arellano, R.; Medrano, F.; Moreno, A.; Romero, A. Structure of struthiocalcin-1, an intramineral protein from Struthio camelus eggshell, in two crystal forms. Acta Crystallogr. Sect. D-Biol. Crystallogr. 2015, 71, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Grajeda, J.P.; Moreno, A.; Romero, A. Crystal Structure of Ovocleidin-17, a Major Protein of the Calcified Gallus gallus Eggshell: Implications in the calcite mineral growth pattern. J. Biol. Chem. 2004, 279, 40876–40881. [Google Scholar] [CrossRef] [PubMed]
- Cölfen, H. A crystal-clear view. Nat. Mater. 2010, 9, 960–961. [Google Scholar] [CrossRef]
- Khan, S.R. Histological aspects of the “fixed particle” model of stone formation: Animal studies. Urolithiasis 2016, 45, 75–87. [Google Scholar] [CrossRef]
- Mann, K. The calcified eggshell matrix proteome of a songbird, the zebra finch (Taeniopygia guttata). Proteome Sci. 2015, 13, 29. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Mann, M. The proteome of the calcified layer organic matrix of turkey (Meleagris gallopavo) eggshell. Proteome Sci. 2013, 11, 40. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M. Synthesis of inorganic and organic crystals mediated by proteins in different biological organisms. A mechanism of biomineralization conserved throughout evolution in all living species. Prog. Cryst. Growth Charact. Mater. 2017, 63, 94–103. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ion | Sample | Type of Synthesis | Raman (cm−1) | FTIR (cm−1) | Composition/Reference |
|---|---|---|---|---|---|
| Ca2+ | Control | Current | 160, 285, 714, 1087 | 712.1, 871.1, 1393.9 | calcite [40] |
| Precambrian | 162, 287, 717, 1089 | 713.1, 871.2, 1406.2 | calcite [40] | ||
| Obsidian | Current | 157, 282, 713, 1086 | 712.5, 788.9, 873.6, 1057.4, 1412.3, 2980.8 | rontgënite (DRX) [41] | |
| Precambrian | 158, 282, 713, 1087 | 783.0, 1035.2, 1450.0, 2980.8 | rontgënite (DRX) [41] | ||
| Current | 98, 141, 157, 226, 692, 1060 | 692.6, 768.5, 855.5, 1059.0, 1416.9 | witherite [42] | ||
| Precambrian | 96, 144, 162, 225, 694, 1063 | 692.4, 855.2, 1077.3, 1417.2 | witherite [42] | ||
| Ba2+ | Current | 95, 138, 153, 691, 1059, 2907 | 783.5, 1023.3, 1425.1 | witherite [42] | |
| Precambrian | 95, 138, 155, 224, 691, 1059 | 590.6, 692.9, 796.2, 856.0, 1058.7, 1432.3 | witherite [42] |
| d(hkl) | Cell Parameters | ||||
|---|---|---|---|---|---|
| Sample | d(003)/Å | d(1,1,15)/Å | c/Å | a = b/Å | V/Å3 |
| Ca_CO2 | 22.867 | 2.8148 | 68.601 | 7.143 c | 3031 |
| Ca_CO2-T | 23.299 | 2.8149 | 69.897 | 7.064 c | 3020 |
| [41] a | 23.094(10) b | — | 69.283(30) | 7.049(1) c | 2981(1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuéllar-Cruz, M.; Zúñiga-Estrada, E.A.; Zúñiga-Estrada, M.A.; Islas, S.R.; Moreno, A. Self-Induced Crystalline Morphology at the Mineral–Fluid Interface: Silica–Carbonate Biomorphs of Alkaline Earth Metals as a Case Study. Appl. Sci. 2025, 15, 4593. https://doi.org/10.3390/app15094593
Cuéllar-Cruz M, Zúñiga-Estrada EA, Zúñiga-Estrada MA, Islas SR, Moreno A. Self-Induced Crystalline Morphology at the Mineral–Fluid Interface: Silica–Carbonate Biomorphs of Alkaline Earth Metals as a Case Study. Applied Sciences. 2025; 15(9):4593. https://doi.org/10.3390/app15094593
Chicago/Turabian StyleCuéllar-Cruz, Mayra, Erick Alfredo Zúñiga-Estrada, Marcelino Antonio Zúñiga-Estrada, Selene R. Islas, and Abel Moreno. 2025. "Self-Induced Crystalline Morphology at the Mineral–Fluid Interface: Silica–Carbonate Biomorphs of Alkaline Earth Metals as a Case Study" Applied Sciences 15, no. 9: 4593. https://doi.org/10.3390/app15094593
APA StyleCuéllar-Cruz, M., Zúñiga-Estrada, E. A., Zúñiga-Estrada, M. A., Islas, S. R., & Moreno, A. (2025). Self-Induced Crystalline Morphology at the Mineral–Fluid Interface: Silica–Carbonate Biomorphs of Alkaline Earth Metals as a Case Study. Applied Sciences, 15(9), 4593. https://doi.org/10.3390/app15094593