In this section, we analyze the advantages and disadvantages of adaptive sex ratio variation in sea lamprey populations under changing resource availability, based on a comprehensive modeling of their life cycle and supported by a population-level competition mechanism through simulation experiments.
3.1. Two-Species Competition Mechanism
This part will focus on creating a model for population competition, integrating the lampreys’ life cycle traits and gender determination processes.
Population expansion occurs in three distinct phases according to the life cycle
The life cycle of lampreys is a complex process divided into fourmain stages, each with its own unique physiological and ecological characteristics. These fourstages include the Hatching Stage, Larval Stage, Metamorphosis Stage, Adult Stage. The four-phase life cycle change is shown in
Figure 3 below.
The Hatching Stage (HS) is the first stage of the life cycle of lampreys. Lamprey eggs are diminutive yet yield a substantial amount. On each occasion, their egg production ranges from 80,000 to 100,000. Typically, eggs are found clinging to sandy and gravelly surfaces. Soon after emerging, they transform into fry and progress to the larval phase.
The Larval Stage (LS) can last up to seven years or more, which is the longest stage of the lampreys’ life cycle [
12]. During this period, larvae live on the bottom of freshwater streams, usually buried in silt or sand, forming U-shaped or slanted burrows.
The Metamorphosis Stage (MS) is relatively short and marks a rapid transition from larva to adult. At the end of this stage, individuals complete sex differentiation, which may be influenced by environmental conditions such as food availability.
The Adult Stage (AS) is the final stage of the lampreys’ life cycle and is shorter in duration relative to the larval stage. Mature lampreys evolve into predators, consuming tiny fish, invertebrates, and various animals. At maturity, lampreys return to freshwater streams to lay eggs and die shortly thereafter, completing their life cycle [
12,
13].
During the modeling process, the metamorphosis stage was ignored because it was in a transitional phase and had a relatively short duration. In conjunction with the stage characterization of sea lampreys, we divided the population growth model into three stages. The relationship between number changes in the three phases is shown in
Figure 4 below. After the larva’s sex is determined, it transforms into an adult. The adult stage of lampreys is divided into male and female.
Consequently, the number of the lamprey population is calculated as
where
,
,
, and
are the number in the hatching stage, the number in the larval stage, the number of male adults, and the number of female adults, respectively.
To build a model reflecting the number of the sea lamprey population, we need to define the pattern of change and the interrelationship of numbers in each stage. The incubation phase (N1) encompasses the transformation of fertilized eggs into larvae. The success rate of hatching can be affected by multiple elements, such as temperature, the intensity of predation, and the prevailing environmental conditions. Larval stage (N2) refers to the phase post-hatching where larvae undergo growth until their transformation into adults. Number growth during this period can be affected by factors such as food availability, mortality, and growth rate. Adult stage (N3 for males and N4 for females): larvae pass through the metamorphosis stage to become adults, with the sexes differentiating at the end of the metamorphosis stage. The number of adults (males and females) is affected by factors such as the success of metamorphosis, the proportion of sex differentiation, and the mortality rate of adults. Sex differentiation can be expressed as the proportion of females
and the proportion of males
in the adult population, while we have
From this, we used a set of differential equations with time as the independent variable to describe the number of lampreys at each stage. We adopted Assumption 3 (overlook the impact of disease spread and movement within and outside populations on the size of the population) and Assumption 5 (individuals at an identical developmental phase in a population are considered equal here).
where
is the hatching success rate,
are the maturation rate from fertilized eggs to juveniles, the maturation rate from juveniles to adults, and the mating rate of adults, and
are the mortality rates of lamprey eggs, juveniles, and adult males and females, respectively. Here,
is the lower number of males and females, which determines the likelihood of mating. This setting ensures that the number of mating events is limited by the sex ratio balance.
Relationship between sex ratio and food availability under sex-determined mechanisms
Lampreys’ sex determination mechanism is environmentally delicate and, in contrast to numerous other species, does not rely exclusively on genetic elements. Specifically, the sex of the lampreys is not fixed at fertilization but is influenced by specific conditions in the environment, especially food availability. This mechanism of sex determination is characterized by the following:
Association between growth rate and sex differentiation: During the larval stage of lampreys, the growth rate of an individual affects its eventual sex. This means that sex determination is directly related to the environmental conditions encountered by the individual during the initial phase of development.
Role of food availability: Food availability is considered a key factor in determining the growth rate of larvae. Larvae in environments abundant in food exhibit quicker growth, typically leading to a greater female ratio. Conversely, in environments with limited food availability, larvae exhibit slower growth and a comparatively larger male population.
Variability in sex ratio: Since sex is determined based on environmental conditions, this leads to significant variability in the sex ratio of lampreys under different environmental conditions. For example, when food availability is low, the proportion of males can reach about 78% of the population, whereas in environments abundant in food, the proportion of males may drop to about 56% [
1].
Therefore, we first constructed a sex ratio function for lampreys.
Equations (8) and (9) describe the relationship between sex ratio and food availability per unit of larva.
is the proportion of males in the total adult population.
denotes the average amount of food available per larva, where
is the total amount of food available and
is the number of lampreys in the larval stage. We adopted Assumption 1 (food supply is the main factor influencing the growth rate of sea eel larvae).
is a decreasing function reflecting the ecological trend that greater food availability leads to a lower male proportion and a higher female proportion. The specific form of the sex ratio function is as follows:
The effect of food quantity on the sex ratio is regulated by parameters and . This function may reflect the tendency for the proportion of males to decrease and the proportion of females to increase as food availability increases.
Since food availability is dynamic, we developed an equation for food availability over time
where
represents the natural rate of growth of food and
is the coefficient on the rate of food consumption. When studying the advantages and disadvantages of sex ratio variation in lampreys and analyzing whether changing the sex ratio is beneficial to coping with changes in external conditions, we should compare the differences between the two situations with adaptive sex mechanisms and without sex mechanisms. A two-species competition mechanism was subsequently introduced [
14], in which a population of sea lampreys with adaptive sex ratios competed with a population of sea lampreys with fixed sex ratios to more visually examine changes in sex ratios, reproductive success, and population size in the face of being competed against by conspecifics. To integrate the gender composition into the model of population competition, the initial value of this function,
, is utilized to ascertain the population’s initial gender makeup through a gender adaptation process:
The initial gender composition of the adaptive mechanism population is
where
is the total amount of food available at the initial time,
is the number of female lampreys in the larval stage at the initial time,
is the number of male lampreys in the larval stage at the initial time, and
is the the number of lampreys in the larval stage at the initial time.
Dominant population model based on competitive mechanisms
The basic mathematical model of the competition between two species is constructed below.
Let two kinds of groups
, in the same environment rely on the same limited resources to survive. Set the moment
when the two species’ total numbers were
and
, the growth of the population is subject to their laws, the natural growth rate of
and
, respectively, when the other side of the extinction of the number of survival is, respectively,
and
. In this context,
is the
referenced in Equation (12).
Set the initial moment when both populations are small. If the consumption of resources by the individuals of the second population is
times the consumption of the individuals of the first population, then the growth rate of the first species group is
Similarly, if individuals of the first population consume
times more resources than individuals of the second population, the growth rate of the second species group is
The growth rate of each population (
and
) reflects the natural ability to grow in the absence of competitive pressures. This rate depends on the reproduction rate, mortality rate, and viability of individuals. Here, we have employed basic Assumption 4 (limited environmental carrying capacity).
and
denote the maximum population size that each population can reach in the absence of competition, i.e., the carrying capacity of the environment.
and
denote the resource consumption between two populations. This ratio determines the amount of competitive pressure one population exerts on the other. Thus, the total number of two populations
and
satisfy the following set of differential equations
Introducing
where
are competition coefficients that characterize the strength of interactions between populations. These coefficients reflect the degree of influence of one population on the growth of another. Substituting Equation (17) into Equation (16) yields
Once initial values are allocated to the populations, the equation system becomes the mathematical representation of the two-species competitive system.
3.2. Results Analysis
In this subsection, we will perform experimental simulations using the previously developed two-population competition model and present and analyze the experimental results. In the analysis results, we employed the two hypotheses mentioned earlier:
Assumption 2: Variations in the sex ratio have a direct impact on the rate of reproduction and the size of the population.
Assumption 6: Once established, the sexes remain unchanged.
Before comparing these two competing populations, we first give the algorithm for solving the differential equations and define two new metrics for comparison, including environmental resource utilization and reproductive success.
The following Algorithm 1 is designed to address the differential equations in the intricate growth model introduced above.
| Algorithm 1. Fourth-order Runge-Kutta method solves ODE |
| INPUT: the initial conditions: , , …, iteration step range |
| OUTPUT: changes over time results |
| 1: | for in do |
| 2: | |
| 3: | while do |
| 4: |
|
| 5: |
|
| 6: |
|
| 7: | similar … |
| 8: | # Update |
| 9: |
|
| 10: |
|
| 11: | end while |
| 12: | return and determine convergence and error |
| 13: | end for |
Figure 5 displays the outcomes of applying the aforementioned algorithm to model variations in lampreys’ number and sex ratio over time, considering different conditions of food availability. The outcomes under conditions of food adequacy are depicted in
Figure 5a, while
Figure 5b presents the findings in scenarios of food scarcity.
Comparing
Figure 5a,b, we can see that in the first five years, the male ratio of the food-sufficient group was lower, while the male ratio of the food-deficient group was higher, which is consistent with the theory that “males have a lower energy threshold for growth”. As time went on, the number of food-deficient groups dropped sharply and approached extinction. When contrasted, the proportion of males in the group lacking food was found to be lower. This indicates that the male ratio is affected by multiple factors and has an opposite trend over time.
Over time, as depicted in
Figure 5a,b, it becomes evident that the male ratio tends to stabilize at a specific value. Now, we will precisely analyze whether such a balance point exists. If it does exist, then:
From Equations (6) and (7), we can obtain:
that is,
From Equation (11), we can obtain:
since
is one-to-one corresponding to
, when
remains constant, by analyzing the above equation, we can conclude that the fluctuation of
depends on
:
When , is greater than .
When , is less than .
When , is equal to .
The proportion of hatchery and juvenile lampreys was large and similar, with the sum of the two exceeding 80%, and the ratio of males to females gradually decreased from an initial 3:1 to nearly 1:1, as depicted in
Figure 6.
In summary, the complete population growth model of lampreys takes into account its life cycle stage pattern and sex determination mechanism and synthesizes the complexity and ecological dynamics of biological processes. Modifying the parameters of the model enables the forecasting of how various environmental scenarios and management strategies impact the lampreys’ population composition and number. It can be seen that food sufficiency leads to an increase in the number of sea lampreys and eventually stabilization, and the male ratio increases and stabilizes, while food scarcity leads to a decrease in the number of sea lampreys and an increase in the male-to-female ratio followed by a decrease and a gradual tendency toward 1:1.
Furthermore, we set up two types of lampreys with identical conditions except for the trait of whether the sex ratio changed over time. First, we introduce two comparative indicators, including environmental resource utilization and reproductive success.
Environmental Resource Utilization : the ratio of a population’s number to its environmental holding capacity . We adopted Assumption 4.
Reproductive success (): the product of hatching success and mating success in Equation (4).
The expression is as follows:
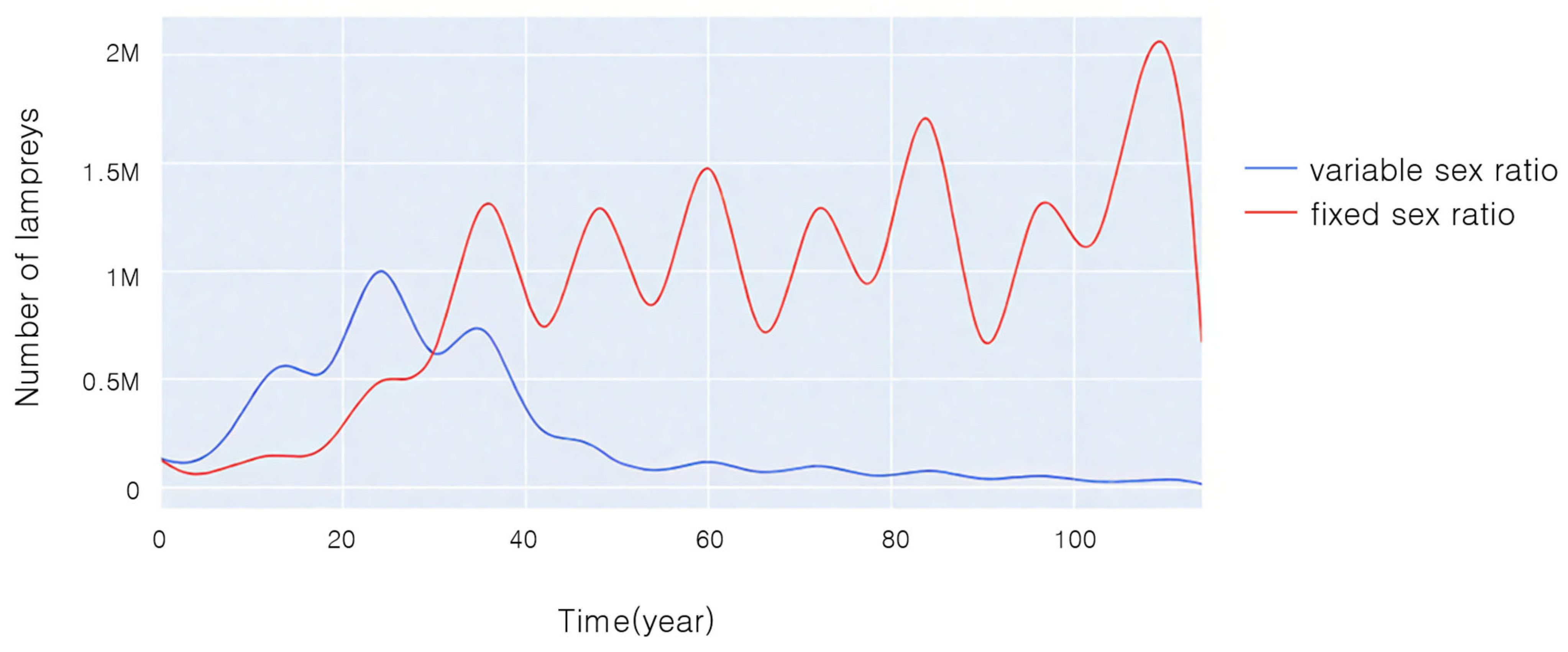
Figure 7 illustrates the variations in population sizes of two species over time under experimental conditions with the same initial sex ratio and initial age composition when sea lampreys with sex ratios that can change in response to the environment compete with sea lampreys with unchanging sex ratios.
From
Figure 7, it can be seen that in the first 30 years, the population size of both lampreys tended to become larger, and the population size of sea lampreys with a variable sex ratio was significantly larger than that of those with a non-variable sex ratio. Thirty years later, the population size of sea lampreys with a non-variable sex ratio exceeded that of those with a variable sex ratio, and gradually tended to be in dynamic equilibrium, whereas the population size of lampreys with a variable sex ratio gradually became smaller and smaller, approaching nearly extinct status.
Figure 8 compares the differences of the following indices between the two populations:
Stability of population sex ratio: We set the initial male ratio of both lampreys to 0.7 and found that the male ratio of lampreys with an unchanged sex ratio gradually returned to normal, i.e., 0.5, while the male ratio of lampreys with a changeable sex ratio in the external environment was maintained at 0.7, with a tendency to increase.
Stability of age composition structure: For the juvenile ratio of both lampreys, we set the initial juvenile ratio of both at 0.96 and found that the juvenile ratio of lampreys with no change in sex ratio stayed at 0.96.
Reproductive success: Our finding indicates that the reproductive success rate of lampreys with an unchanged sex ratio gradually decreased, and the decrease was large, while the reproductive success rate of lampreys with a changeable sex ratio was maintained at about 0.7.
Environmental resource utilization: Regarding the resource utilization of the two species, it was found that the resource utilization of lampreys, which did not change their sex ratio, showed a decreasing trend and remained low for the first 20 years. On the other hand, the resource utilization of lampreys that can change their sex ratio following the external environment showed a significant upward trend, dynamically increasing from 10% to 70%.
From this, we can further conclude that the advantages of a variable sex ratio in lamprey populations include increased adaptability to environmental changes, the optimization of resource use, and reproductive success by adjusting the sex ratio.
We summarize the advantages and disadvantages of variable sex ratios in lamprey populations, and overall, if a population with variable sex ratios is allowed to compete with non-variable populations, it is not advantageous.
The advantages include increased resilience to environmental change by adjusting sex ratios to optimize environmental resource utilization and reproductive success. Within environments abundant in material resources, an increased number of females might promote swift population increase and proliferation. Such a mechanism may help populations to remain stable when environmental conditions change.
The disadvantage of this sex mechanism is that in resource-constrained and competitive environments, the population structure (composed of age composition and sex ratio) is unstable, and excessive bias toward one sex may lead to a reduction in genetic diversity. Over time, this could impact the population’s survival and reproductive capabilities.
We speculate that fluctuations in sex ratios may have an impact on other species in the ecosystem, altering the existing balance of the food chain, which will be discussed in subsequent sections. The impacts on ecosystems are complex and multifaceted, and further research is needed to gain a deeper understanding of the long-term impacts of sex ratio fluctuations on ecological balance and other species.




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}