
An Efficient Rice Virus-Induced Gene Silencing System Mediated by Wheat Dwarf Virus



Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Construction of the Vector
2.3. WDV Infection Method
2.4. Infection of Magnaporthe Oryzae
2.5. Data Analysis
3. Results
3.1. Detection and Phenotype Identification of Transgenic Rice Plants Transformed Using the VIGS Gene Silencing System
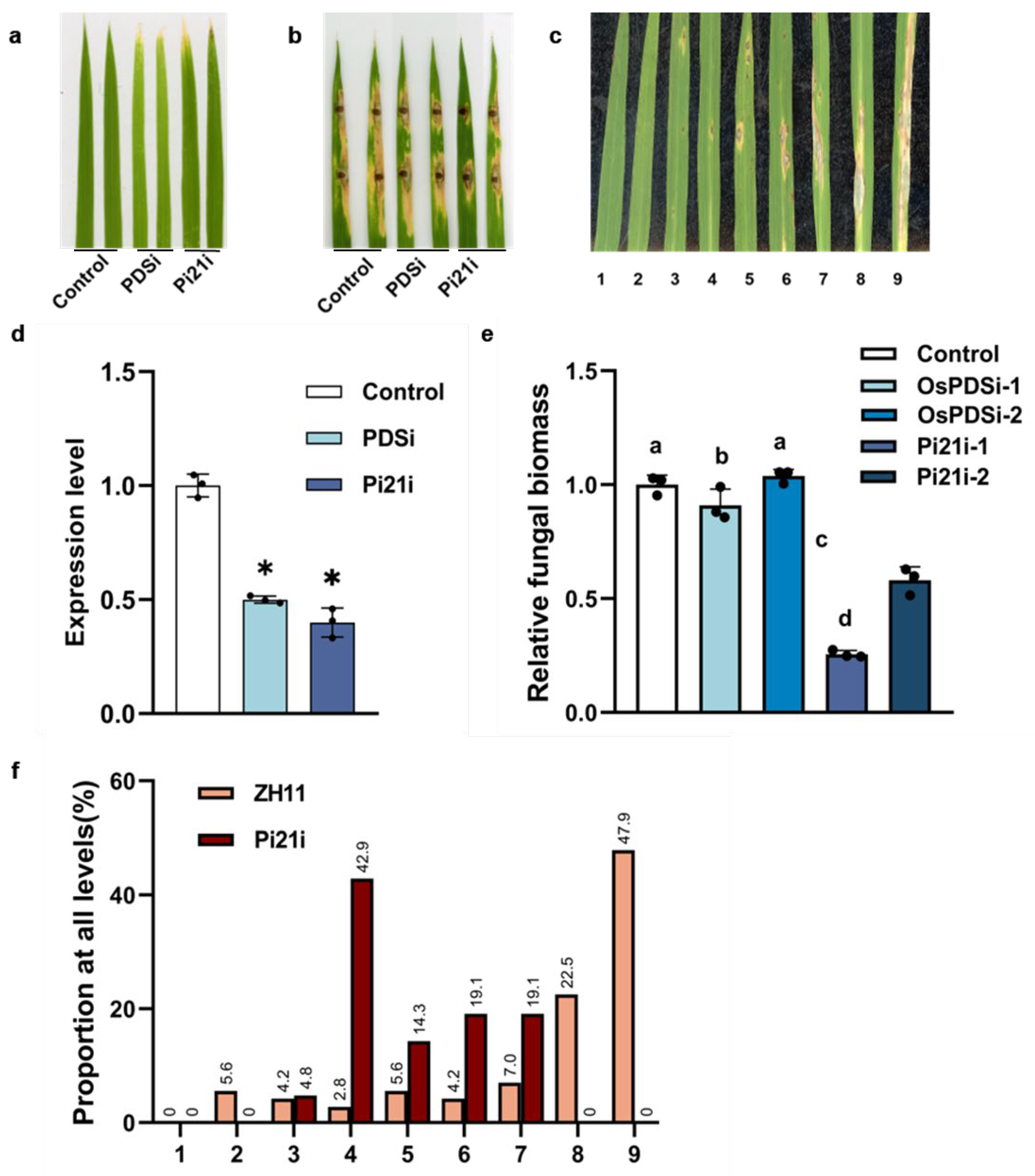
3.2. Silence OsPi21 by the WDV-VIGS System to Enhances Rice Blast Resistance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zulfiqar, S.; Farooq, M.A.; Zhao, T.; Wang, P.; Tabusam, J.; Wang, Y.; Xuan, S.; Zhao, J.; Chen, X.; Shen, S.; et al. Virus-Induced Gene Silencing (VIGS): A Powerful Tool for Crop Improvement and Its Advancement towards Epigenetics. Int. J. Mol. Sci. 2023, 24, 5608. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Yellina, A.L.; Orashakova, S.; Becker, A. Virus-Induced Gene Silencing (VIGS) in Plants: An Overview of Target Species and the Virus-Derived Vector Systems. Methods Mol. Biol. 2013, 975, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jagram, N.; Dasgupta, I. Principles and Practice of Virus Induced Gene Silencing for Functional Genomics in Plants. Virus Genes 2023, 59, 173–187. [Google Scholar] [CrossRef]
- Rössner, C.; Lotz, D.; Becker, A. VIGS Goes Viral: How VIGS Transforms Our Understanding of Plant Science. Annu. Rev. Plant Biol. 2022, 73, 703–728. [Google Scholar] [CrossRef]
- Wege, S.; Scholz, A.; Gleissberg, S.; Becker, A. Highly Efficient Virus-Induced Gene Silencing (VIGS) in California Poppy (Eschscholzia californica): An Evaluation of VIGS as a Strategy to Obtain Functional Data from Non-Model Plants. Ann. Bot. 2007, 100, 641–649. [Google Scholar] [CrossRef]
- Abt, I.; Souquet, M.; Angot, G.; Mabon, R.; Dallot, S.; Thébaud, G.; Jacquot, E. Functional Transcomplementation between Wheat Dwarf Virus Strains in Wheat and Barley. Viruses 2019, 12, 34. [Google Scholar] [CrossRef]
- Rahman, J.; Baldwin, I.T.; Gase, K. California TRV-Based VIGS Vectors Mediate Gene Silencing at Elevated Temperatures but with Greater Growth Stunting. BMC Plant Biol. 2021, 21, 553. [Google Scholar] [CrossRef]
- McCaghey, M.; Shao, D.; Kurcezewski, J.; Lindstrom, A.; Ranjan, A.; Whitham, S.A.; Conley, S.P.; Williams, B.; Smith, D.L.; Kabbage, M. Host-Induced Gene Silencing of a Sclerotinia sclerotiorum oxaloacetate acetylhydrolase Using Bean Pod Mottle Virus as a Vehicle Reduces Disease on Soybean. Front. Plant Sci. 2021, 12, 677631. [Google Scholar] [CrossRef]
- Li, T.; Hu, J.; Sun, Y.; Li, B.; Zhang, D.; Li, W.; Liu, J.; Li, D.; Gao, C.; Zhang, Y.; et al. Highly Efficient Heritable Genome Editing in Wheat Using an RNA Virus and Bypassing Tissue Culture. Mol. Plant 2021, 14, 1787–1798. [Google Scholar] [CrossRef]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Blast Resistance in Rice: A Review of Conventional Breeding to Molecular Approaches. Mol. Biol. Rep. 2013, 40, 2369–2388. [Google Scholar] [CrossRef]
- Pandey, P.; Senthil-Kumar, M.; Mysore, K.S. Advances in Plant Gene Silencing Methods. Methods Mol. Biol. 2015, 1287, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Pacak, A.; Geisler, K.; Jørgensen, B.; Barciszewska-Pacak, M.; Nilsson, L.; Nielsen, T.H.; Johansen, E.; Grønlund, M.; Jakobsen, I.; Albrechtsen, M. Investigations of Barley Stripe Mosaic Virus as a Gene Silencing Vector in Barley Roots and in Brachypodium distachyon and Oat. Plant Methods 2010, 6, 26. [Google Scholar] [CrossRef]
- Bennypaul, H.S.; Mutti, J.S.; Rustgi, S.; Kumar, N.; Okubara, P.A.; Gill, K.S. Virus-Induced Gene Silencing (VIGS) of Genes Expressed in Root, Leaf, and Meiotic Tissues of Wheat. Funct. Integr. Genom. 2012, 12, 143–156. [Google Scholar] [CrossRef]
- Ding, X.S.; Schneider, W.L.; Chaluvadi, S.R.; Mian, M.A.R.; Nelson, R.S. Characterization of a Brome mosaic virus Strain and Its Use as a Vector for Gene Silencing in Monocotyledonous Hosts. Mol. Plant-Microbe Interact. 2006, 19, 1229–1239. [Google Scholar] [CrossRef]
- Liou, M.-R.; Huang, Y.-W.; Hu, C.-C.; Lin, N.-S.; Hsu, Y.-H. A Dual Gene-Silencing Vector System for Monocot and Dicot Plants. Plant Biotechnol. J. 2014, 12, 330–343. [Google Scholar] [CrossRef]
- Purkayastha, A.; Mathur, S.; Verma, V.; Sharma, S.; Dasgupta, I. Virus-Induced Gene Silencing in Rice Using a Vector Derived from a DNA Virus. Planta 2010, 232, 1531–1540. [Google Scholar] [CrossRef]
- Kumar, G.; Kumari, K.; Dasgupta, I. RTBV-Based VIGS Vector for Functional Genomics in Rice: Methodology, Advances, Challenges, and Future Implications. Methods Mol. Biol. 2022, 2408, 117–131. [Google Scholar] [CrossRef]
- Cejnar, P.; Ohnoutkova, L.; Ripl, J.; Kundu, J.K. Wheat dwarf virus Infectious Clones Allow to Infect Wheat and Triticum monococcum Plants. Plant Prot. Sci. 2019, 55, 81–89. [Google Scholar] [CrossRef]
- Baltes, N.J.; Gil-Humanes, J.; Cermak, T.; Atkins, P.A.; Voytas, D.F. DNA Replicons for Plant Genome Engineering. Plant Cell 2014, 26, 151–163. [Google Scholar] [CrossRef]
- Wei, S.; Liu, L.; Chen, G.; Yang, H.; Huang, L.; Gong, G.; Luo, P.; Zhang, M. Molecular Evolution and Phylogeographic Analysis of Wheat Dwarf Virus. Front. Microbiol. 2024, 15, 1314526. [Google Scholar] [CrossRef]
- Tian, Y.; Zhong, D.; Li, X.; Shen, R.; Han, H.; Dai, Y.; Yao, Q.; Zhang, X.; Deng, Q.; Cao, X.; et al. High-Throughput Genome Editing in Rice with a Virus-Based Surrogate System. J. Integr. Plant Biol. 2023, 65, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Cejnar, P.; Ohnoutkova, L.; Ripl, J.; Vlcko, T.; Kundu, J.K. Two Mutations in the Truncated Rep Gene RBR Domain Delayed the Wheat dwarf virus Infection in Transgenic Barley Plants. J. Integr. Agric. 2018, 17, 2492–2500. [Google Scholar] [CrossRef]
- Ashkani, S.; Rafii, M.Y.; Shabanimofrad, M.; Miah, G.; Sahebi, M.; Azizi, P.; Tanweer, F.A.; Akhtar, M.S.; Nasehi, A. Molecular Breeding Strategy and Challenges Towards Improvement of Blast Disease Resistance in Rice Crop. Front. Plant Sci. 2015, 6, 886. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Xie, J.; Chen, Y.; Shen, Z.; Shi, H.; Naqvi, N.I.; Qian, Q.; Liang, Y.; Kou, Y. Warm Temperature Compromises JA-Regulated Basal Resistance to Enhance Magnaporthe oryzae Infection in Rice. Mol. Plant 2022, 15, 723–739. [Google Scholar] [CrossRef]
- Fukuoka, S.; Saka, N.; Koga, H.; Ono, K.; Shimizu, T.; Ebana, K.; Hayashi, N.; Takahashi, A.; Hirochika, H.; Okuno, K.; et al. Loss of Function of a Proline-Containing Protein Confers Durable Disease Resistance in Rice. Science 2009, 325, 998–1001. [Google Scholar] [CrossRef]
- Liu, M.; Wang, F.; He, B.; Hu, J.; Dai, Y.; Chen, W.; Yi, M.; Zhang, H.; Ye, Y.; Cui, Z.; et al. Targeting Magnaporthe oryzae Effector MoErs1 and Host Papain-like Protease OsRD21 Interaction to Combat Rice Blast. Nat. Plants 2024, 10, 618–632. [Google Scholar] [CrossRef]
- Fukuoka, S.; Okuno, K. QTL Analysis and Mapping of Pi21, a Recessive Gene for Field Resistance to Rice Blast in Japanese Upland Rice. Theor. Appl. Genet. 2001, 103, 185–190. [Google Scholar] [CrossRef]
- Pandey, P.; Mysore, K.S.; Senthil-Kumar, M. Recent Advances in Plant Gene Silencing Methods. Methods Mol. Biol. 2022, 2408, 1–22. [Google Scholar] [CrossRef]
- Dommes, A.B.; Gross, T.; Herbert, D.B.; Kivivirta, K.I.; Becker, A. Virus-Induced Gene Silencing: Empowering Genetics in Non-Model Organisms. J. Exp. Bot. 2019, 70, 757–770. [Google Scholar] [CrossRef]
- Bruun-Rasmussen, M.; Madsen, C.T.; Jessing, S.; Albrechtsen, M. Stability of Barley Stripe Mosaic Virus-Induced Gene Silencing in Barley. Mol. Plant-Microbe Interact. 2007, 20, 1323–1331. [Google Scholar] [CrossRef]
- Lacomme, C.; Hrubikova, K.; Hein, I. Enhancement of Virus-Induced Gene Silencing through Viral-Based Production of Inverted-Repeats. Plant J. Cell Mol. Biol. 2003, 34, 543–553. [Google Scholar] [CrossRef]
- Zaidi, S.S.-E.-A.; Vasudevan, K.; Lentz, E.M.; Vanderschuren, H. Virus-Induced Gene Silencing (VIGS) in Cassava Using Geminivirus Agroclones. Methods Mol. Biol. 2020, 2172, 51–64. [Google Scholar] [CrossRef]
- Zhou, T.; Dong, L.; Jiang, T.; Fan, Z. Silencing Specific Genes in Plants Using Virus-Induced Gene Silencing (VIGS) Vectors. Methods Mol. Biol. 2022, 2400, 149–161. [Google Scholar] [CrossRef]
- Liu, N.; Xie, K.; Jia, Q.; Zhao, J.; Chen, T.; Li, H.; Wei, X.; Diao, X.; Hong, Y.; Liu, Y. Foxtail Mosaic Virus-Induced Gene Silencing in Monocot Plants. Plant Physiol. 2016, 171, 1801–1807. [Google Scholar] [CrossRef]
- Gil-Humanes, J.; Wang, Y.; Liang, Z.; Shan, Q.; Ozuna, C.V.; Sánchez-León, S.; Baltes, N.J.; Starker, C.; Barro, F.; Gao, C.; et al. High-Efficiency Gene Targeting in Hexaploid Wheat Using DNA Replicons and CRISPR/Cas9. Plant J. Cell Mol. Biol. 2017, 89, 1251–1262. [Google Scholar] [CrossRef]
- Wang, M.; Lu, Y.; Botella, J.R.; Mao, Y.; Hua, K.; Zhu, J.-K. Gene Targeting by Homology-Directed Repair in Rice Using a Geminivirus-Based CRISPR/Cas9 System. Mol. Plant 2017, 10, 1007–1010. [Google Scholar] [CrossRef]
- Huang, S.; Jia, A.; Song, W.; Hessler, G.; Meng, Y.; Sun, Y.; Xu, L.; Laessle, H.; Jirschitzka, J.; Ma, S.; et al. Identification and Receptor Mechanism of TIR-Catalyzed Small Molecules in Plant Immunity. Plant Sci. 2022, 377, eabq3297. [Google Scholar] [CrossRef]
- Köklü, G.; Ramsell, J.N.E.; Kvarnheden, A. The Complete Genome Sequence for a Turkish Isolate of Wheat dwarf virus (WDV) from Barley Confirms the Presence of Two Distinct WDV Strains. Virus Genes 2007, 34, 359–366. [Google Scholar] [CrossRef]
- Kant, R.; Dasgupta, I. Phenotyping of VIGS-Mediated Gene Silencing in Rice Using a Vector Derived from a DNA Virus. Plant Cell Rep. 2017, 36, 1159–1170. [Google Scholar] [CrossRef]
- Ahlquist, P. Parallels among Positive-Strand RNA Viruses, Reverse-Transcribing Viruses and Double-Stranded RNA Viruses. Nat. Rev. Microbiol. 2006, 4, 371–382. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Primers Name | Primers Sequence |
|---|---|
| WF | TGGTGTTACTTCTGCAGGAGCTCGGCAGTGACTCCGTCTCTG |
| WR | CCTCGCCCTTGCTCACCATGGATCCCAGCTATGACCATGATTACG |
| WDV-F | TCAACTACTCGTTCGCCTCC |
| WDV-R | GCTTGGCTTGCTTACACTCTG |
| WDV-OsPDSi-F | CGACACGCGTACTAGTAGTGAAATC |
| WDV-OsPDSi-R | CCTAGAGCACCGAGCCTCCGAC |
| WDV-OsPi21i-F | CGCGACGCGTACTAGTATGGGTATATTG |
| WDV-OsPi21i-R | CCTCCTGATCTTGGCATC |
| WDV-Rep-F | TACGCCTTGGATTCAC |
| WDV-Rep-R | TACGACCTGGAGTTG |
| qOsPi21-F | GGTATATTGGTCATCTTGG |
| qOsPi21-R | ATCACCCTGTTGTTCTTC |
| qOsPDS-F | TGCCTGTCATCTATGAAC |
| qOsPDS-R | CCTAGAGCACCGAGCCTCCGAC |
| UBQ5-F | AGACCTACACCAAGCCCAAGAAGAT |
| UBQ5-R | CCAGCACCGCACTCAGCATTAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, X.; Yu, L.; Yan, Y.; Zhu, S.; Huang, W.; Zhang, X.; Dang, C.; Xue, D. An Efficient Rice Virus-Induced Gene Silencing System Mediated by Wheat Dwarf Virus. Appl. Sci. 2025, 15, 5818. https://doi.org/10.3390/app15115818
Zhang Y, Zhang X, Yu L, Yan Y, Zhu S, Huang W, Zhang X, Dang C, Xue D. An Efficient Rice Virus-Induced Gene Silencing System Mediated by Wheat Dwarf Virus. Applied Sciences. 2025; 15(11):5818. https://doi.org/10.3390/app15115818
Chicago/Turabian StyleZhang, Yaqian, Xiaowan Zhang, Lu Yu, Yijie Yan, Senzhen Zhu, Wanting Huang, Xian Zhang, Cong Dang, and Dawei Xue. 2025. "An Efficient Rice Virus-Induced Gene Silencing System Mediated by Wheat Dwarf Virus" Applied Sciences 15, no. 11: 5818. https://doi.org/10.3390/app15115818
APA StyleZhang, Y., Zhang, X., Yu, L., Yan, Y., Zhu, S., Huang, W., Zhang, X., Dang, C., & Xue, D. (2025). An Efficient Rice Virus-Induced Gene Silencing System Mediated by Wheat Dwarf Virus. Applied Sciences, 15(11), 5818. https://doi.org/10.3390/app15115818