
Diversity of Soil-Borne Fungi Isolated from Places Frequently Visited by People in the City of Wrocław (Poland)



Abstract
1. Introduction
2. Materials and Methods
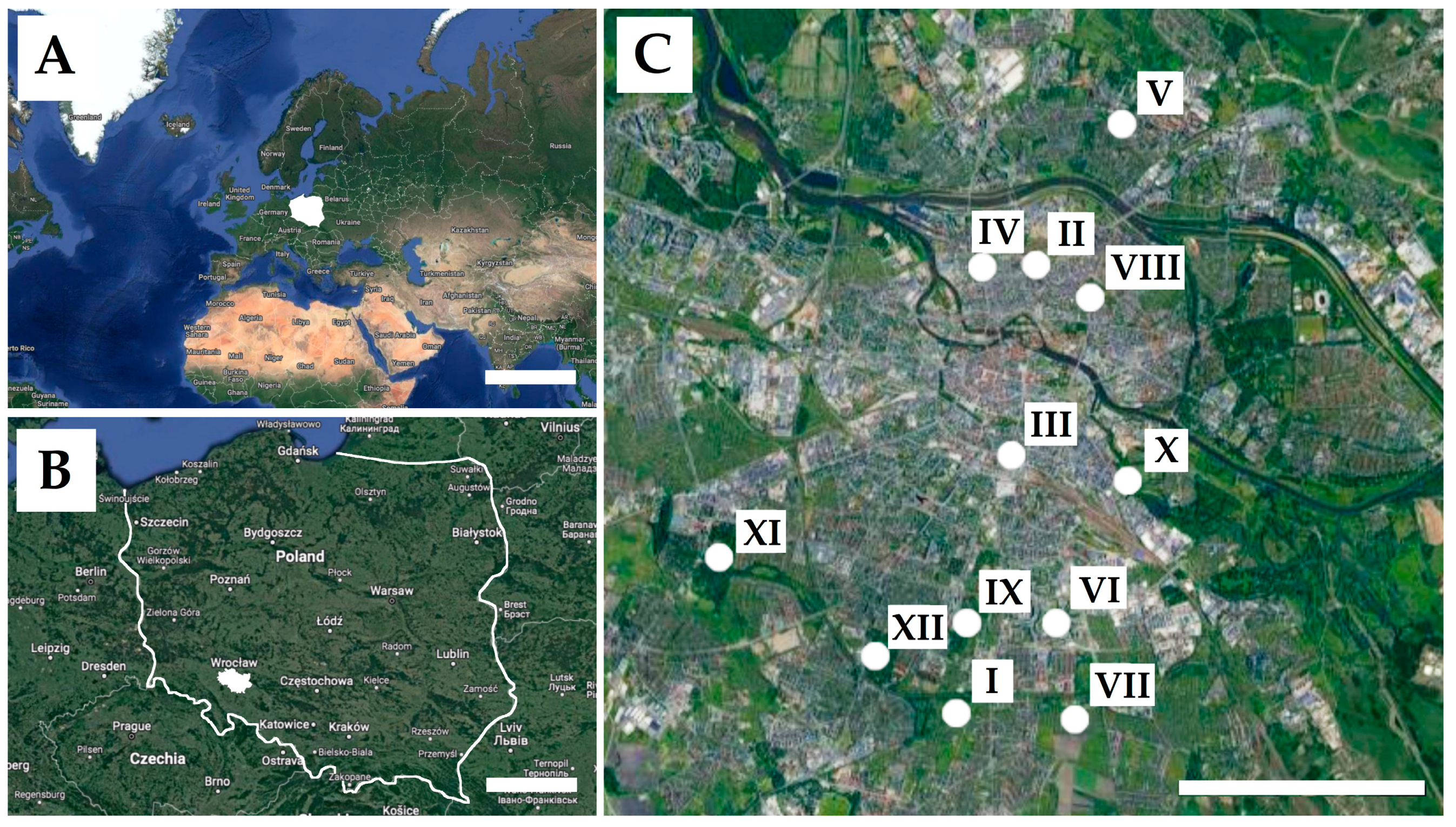
2.1. Study Area
2.2. Sampling
2.3. Culture
2.4. Fungal Identification
2.5. Data Analyses
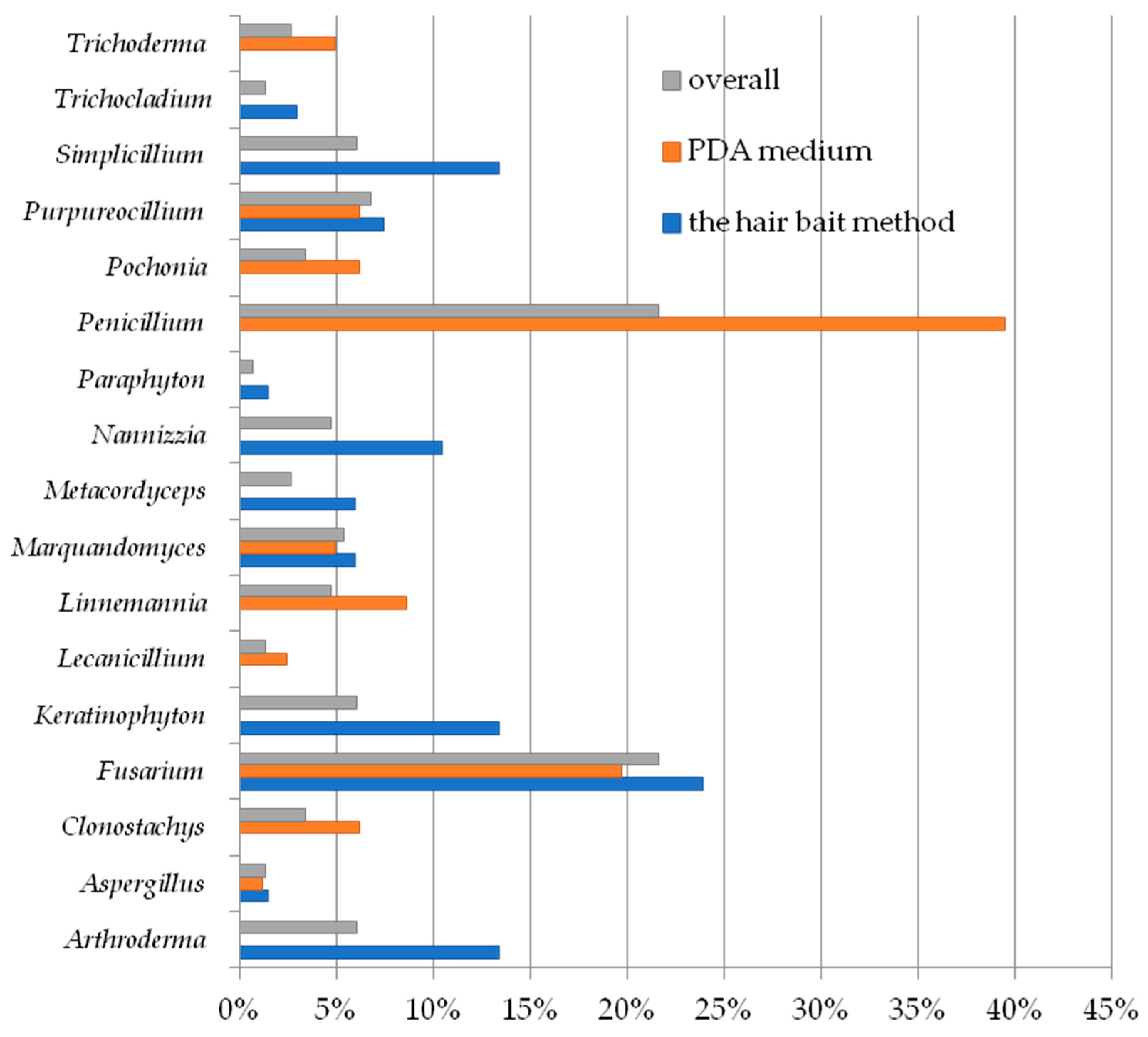
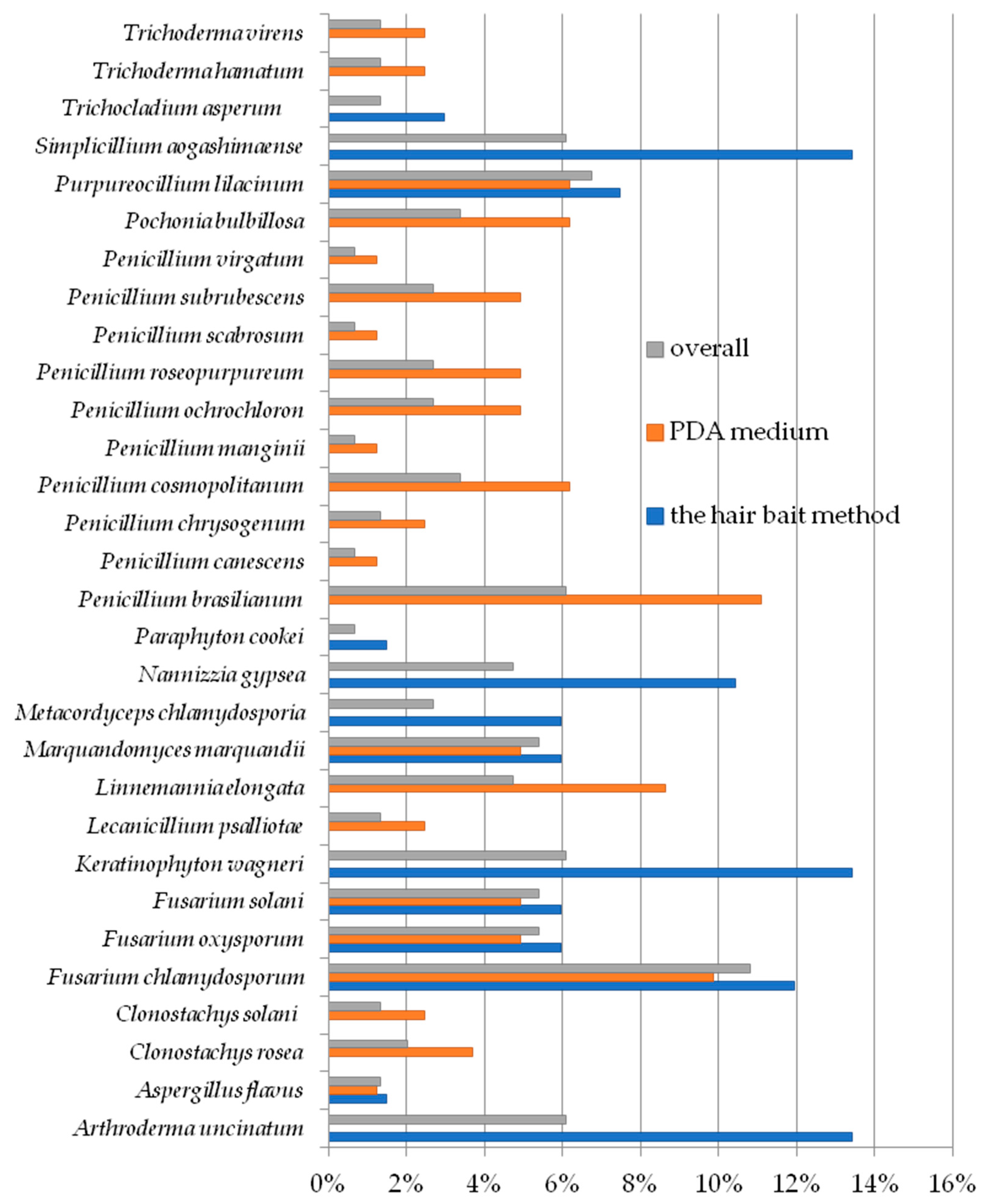
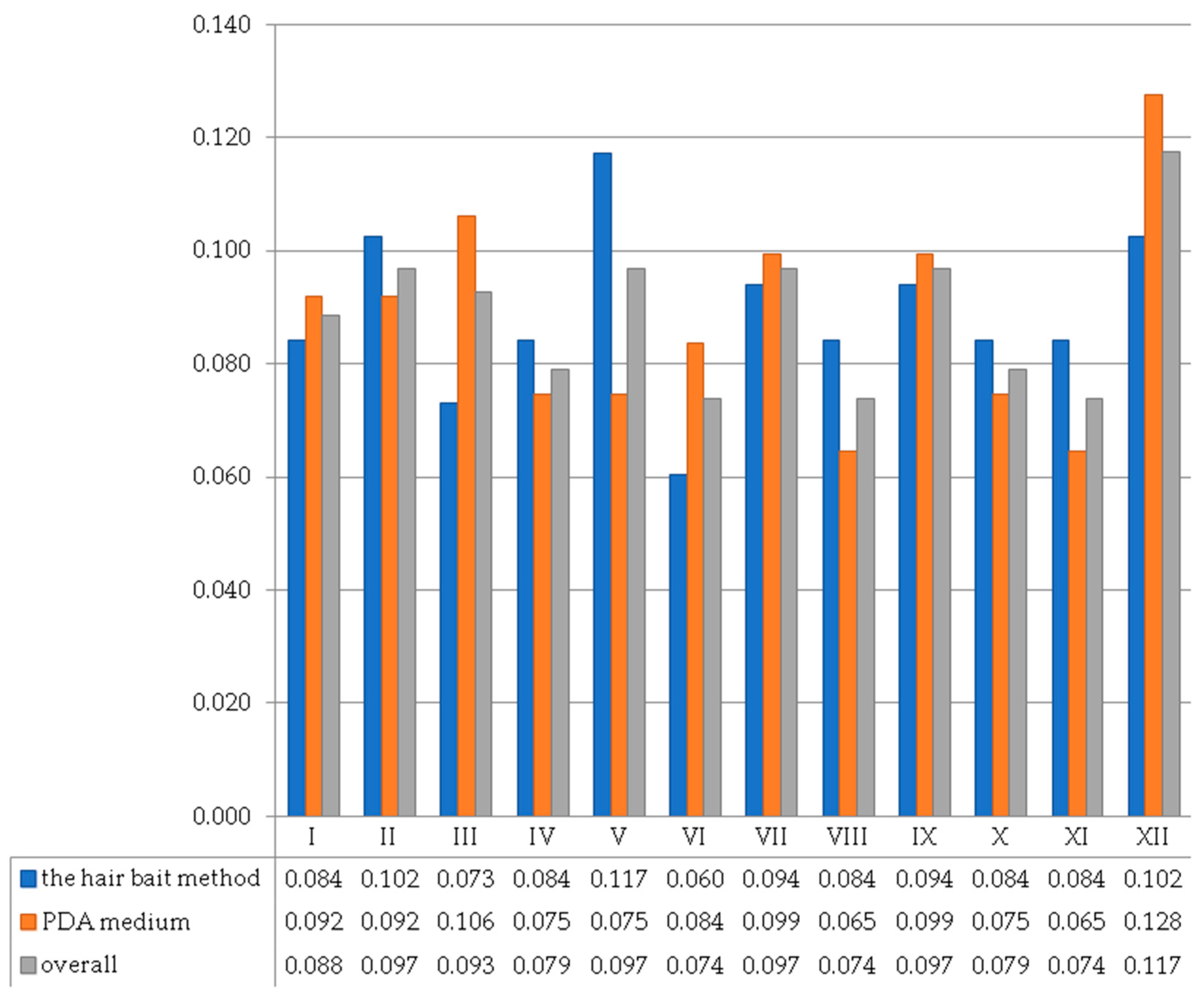
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal Biodiversity and Their Role in Soil Health. Front. Microbiol. 2018, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- Frąc, M.; Jezierska-Tys, S.; Takashi, Y. Occurrence, detection, and molecular and metabolic characterization of heat-resistant fungi in soils and plants and their risk to human health. Adv. Agron. 2015, 132, 161–204. [Google Scholar]
- Kuzikova, I.L.; Medvedeva, N.G. Long-Chain Alkylphenol Biodegradation Potential of Soil Ascomycota. Dokl. Biol. Sci. 2023, 511, 228–234. [Google Scholar] [CrossRef] [PubMed]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrell, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Collaet, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Žifčáková, L.; Vetrovský, T.; Howe, A.; Baldrian, P. Microbial activity in forest soil reflects the changes in ecosystem properties between summer and winter. Environ. Microbiol. 2016, 18, 288–301. [Google Scholar] [CrossRef]
- Baldrian, P. Interactions of heavy metals with white-rot fungi. Enzym. Microb. Technol. 2003, 32, 78–91. [Google Scholar] [CrossRef]
- Gu, L.; Coulombe, P. Keratin function in skin epithelia: A broadening palette with surprising shades. Curr. Opin. Cell Biol. 2007, 19, 13–23. [Google Scholar] [CrossRef]
- Chu, P.G.; Weiss, L.M. Keratin expression in human tissues and neoplasms. Histopathology 2002, 40, 403–439. [Google Scholar] [CrossRef]
- Nnolim, N.; Udenigwe, C.; Okoh, A.; Nwodo, U. Microbial Keratinase: Next Generation Green Catalyst and Prospective Applications. Front. Microbiol. 2020, 11, 580164. [Google Scholar] [CrossRef]
- Ulfig, K. The occurrence of keratinolytic fungi in waste and waste-contaminated habitats. In Biology of Dermatophytes and Other Keratinophilic Fungi; Kushwaha, R.K.S., Guarro, J., Eds.; Revista Iberoamericana de Micología: Bilbao, Spain, 2000; Volume 17, pp. 44–50. [Google Scholar]
- Jain, N.; Sharma, M. Influence of Temperature and Culture Conditions on the Survival of Keratinophilic and Dermatophytic Fungi. Braz. Arch. Biol. Technol. 2022, 65, e22210337. [Google Scholar] [CrossRef]
- Begum, J.; Mir, N.A.; Lingaraju, M.C.; Buyamayum, B.; Dev, K. Recent advances in the diagnosis of dermatophytosis. J. Basic Microbiol. 2020, 60, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Newbound, M.; Mccarthy, M.A.; Lebel, T. Fungi and the urban environment: A review. Landsc. Urban Plan. 2010, 96, 138–145. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Shao, Q.Y.; Li, X.; Chen, W.H.; Liang, J.D.; Han, Y.F.; Huang, J.Z.; Liang, Z.Q. Culturable Fungi from Urban Soils in China I: Description of 10 New Taxa. Microbiol. Spectr. 2021, 9, e0086721. [Google Scholar] [CrossRef]
- Al-Defiery, M.E.; Al-Shaam, T.J.B.; Al-Husaniy, L.R. Fungal diversity in some soils of Hillah city. AIP Conf. Proc. 2023, 2977, 040016. [Google Scholar] [CrossRef]
- Lykov, I.N.; Pavlova, O.P.; Rudova, S.A. Sanitary and hygienic aspects of urban environment pollution by dog feces. In Proceedings of the IV International Scientific Conference: AGRITECH-IV-2020: Agribusiness, Environmental Engineering and Biotechnologies, Krasnoyarsk, Russia, 18–20 November 2020; IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 677, p. 052033. [Google Scholar] [CrossRef]
- McGuire, K.L.; Payne, S.G.; Palmer, M.I.; Gillikin, C.M.; Keefe, D.; Kim, S.J.; Gedallovich, S.M.; Discenza, J.; Rangamannar, R.; Koshner, J.A.; et al. Digging the New York City Skyline: Soil fungal communities in green roofs and city parks. PLoS ONE 2013, 8, e58020. [Google Scholar] [CrossRef] [PubMed]
- Abrego, N.; Crosier, B.; Somervuo, P.; Ivanova, N.; Abrahamyan, A.; Abdi, A.; Hämäläinen, K.; Junninen, K.; Maunula, M.; Purhonen, J.; et al. Fungal communities decline with urbanization-more in air than in soil. ISME J. 2020, 14, 2806–2815. [Google Scholar] [CrossRef] [PubMed]
- Szymanowski, M.; Kryza, M. GIS-based techniques for urban heatisland spatialization. Clim. Res. 2009, 38, 171–187. [Google Scholar] [CrossRef]
- Wrocław Climate (Poland). Data and graphs for weather & climate in Wrocław. 2023. Available online: https://en.climate-data.org/europe/poland/lower-silesian-voivodeship/wroc%C5%82aw-4531/ (accessed on 30 December 2023).
- Ogórek, R.; Piecuch, A.; Višňovská, Z.; Cal, M.; Niedźwiecka, K. First report on the occurence of dermatophytes of Microsporum cookei clade and close affinities to Paraphyton cookei in the Harmanecká Cave (Veľká Fatra Mts., Slovakia). Diversity 2019, 11, 191. [Google Scholar] [CrossRef]
- Vanbreuseghem, R. Technique biologique pour l’isolement des dermatophytes du sol [Biological technique for isolating dermatophytes from soil]. Ann. Soc. Belg. Med. Trop. 1952, 32, 173–178. [Google Scholar]
- Dukik, K.; de Hoog, G.S.; Stielow, J.B.; Freeke, J.; van den Ende, B.G.; Vicente, V.A.; Menken, S.B.J.; Ahmed, S.A. Molecular and Phenotypic Characterization of Nannizzia (Arthrodermataceae). Mycopathologia 2020, 185, 9–35. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Samson, R.A.; Stolk, A.C. A new species of Penicillium, P. scabrosum. Persoonia 1990, 14, 177–182. [Google Scholar]
- Hafizi, R.; Salleh, B.; Latiffah, Z. Morphological and molecular characterization of Fusarium solani and F. oxysporum associated with crown disease of oil palm. Braz. J. Microbiol. 2014, 44, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Frisvad, J.C.; Samson, R.A. Taxonomy of Penicillium section Citrina. Stud. Mycol. 2011, 70, 53–138. [Google Scholar] [CrossRef] [PubMed]
- Kwasna, H.; Nirenberg, H.I. Delimitation of Penicillium virgatum sp. nov. and P. daleae on the basis of morphological and molecular characters. Mycol. Res. 2005, 109, 974–982. [Google Scholar] [CrossRef] [PubMed]
- Labuda, R.; Bernreiter, A.; Hochenauer, D.; Kubátová, A.; Kandemir, H.; Schüller, C. Molecular systematics of Keratinophyton: The inclusion of species formerly referred to Chrysosporium and description of four new species. IMA Fungus 2012, 12, 17. [Google Scholar] [CrossRef]
- Luangsa-Ard, J.J.; Houbraken, J.; van Doorn, T.; Hong, S.B.; Borman, A.M.; Hywel-Jones, N.L.; Samson, R.A. Purpureocillium, a new genus for the medically important Paecilomyces lilacinus. FEMS Microbiol. Lett. 2011, 321, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, S.; Houbraken, J.; Samson, R.A.; Frisvad, J.C.; Christensen, M.; Tuthill, D.E.; Koutaniemi, S.; Hatakka, A.; Lankinen, P. A new Penicillium species efficiently producing inulinase. Antonie Van Leeuwenhoek 2013, 103, 1343–1357. [Google Scholar] [CrossRef] [PubMed]
- Mongkolsamrit, S.; Khonsanit, A.; Thanakitpipattana, D.; Tasanathai, K.; Noisripoom, W.; Lamlertthon, S.; Himaman, W.; Houbraken, J.; Samson, R.A.; Luangsa-Ard, J. Revisiting Metarhizium and the description of new species from Thailand. Stud. Mycol. 2020, 95, 171–251. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, K.; Kaifuchi, S.; Masuma, R.; Omura, S. Five new species of Simplicillium from soil in Tokyo, Japan. Mycoscience 2013, 54, 42–53. [Google Scholar] [CrossRef]
- Schroers, H.J. A monograph of Bionectria (Ascomycota, Hypocreales, Bionectriaceae) and its Clonostachys anamorphs. Stud. Mycol. 2001, 46, 1–214. [Google Scholar]
- Schroers, H.-J.; Samuels, G.J.; Seifert, K.A.; Gams, W. Classification of the mycoparasite Gliocladium roseum in Clonostachys as C. rosea, its relationship to Bionectria ochroleuca, and notes on other Gliocladium-like fungi. Mycologia 1999, 91, 365–385. [Google Scholar] [CrossRef]
- Siddiquee, S. Morphology-Based Characterization of Trichoderma Species. In Practical Handbook of the Biology and Molecular Diversity of Trichoderma Species from Tropical Regions. Fungal Biology; Springer: Cham, Switzerland, 2017; pp. 41–73. [Google Scholar] [CrossRef]
- Summerell, B.A.; Salleh, B.; Leslie, J.F. A Utilitarian Approach to Fusarium Identification. Plant Dis. 2003, 87, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Thom, C. Cultural Studies of Species of Penicillium; Wentworth Press: Sydney, Australia, 1910; Volume 118, pp. 1–107. [Google Scholar]
- Vandepol, N.; Liber, J.; Desirò, A.; Na, H.; Kennedy, M.; Barry, K.; Grigoriev, I.V.; Miller, A.N.; O’Donnell, K.; Stajich, J.E.; et al. Resolving the Mortierellaceae phylogeny through synthesis of multi-gene phylogenetics and phylogenomics. Fungal Divers. 2020, 104, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Yang, F.Y.; Meijer, M.; Kraak, B.; Sun, B.D.; Jiang, Y.L.; Wu, Y.M.; Bai, F.Y.; Seifert, K.A.; Crous, P.W.; et al. Redefining Humicola sensu stricto and related genera in the Chaetomiaceae. Stud. Mycol. 2019, 93, 65–153. [Google Scholar] [CrossRef]
- Zare, R.; Gams, W.; Evans, H.C. A revision of Verticillium sect. Prostrata. V. The genus Pochonia, with notes on Rotiferophthora. Nova Hedwig. 2001, 73, 51–86. [Google Scholar]
- Zare, R.; Gams, W. A revision of Verticillium section Prostrata. IV. The genera Lecanicillium and Simplicillium. Nova Hedwig. 2001, 73, 1–50. [Google Scholar] [CrossRef]
- Piecuch, A.; Ogórek, R. Quantitative and qualitative assessment of mycological air pollution in a dormitory bathroom with high humidity and fungal stains on the ceiling. Case Study. Pol. J. Environ. Stud. 2021, 30, 1955–1960. [Google Scholar] [CrossRef]
- Krzyściak., P.; Skóra, M.; Macura, A.B. Atlas of Human Pathogenic Fungi; MedPharm Polska: Wrocław, Poland, 2010. [Google Scholar]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar] [CrossRef]
- Ogórek, R.; Dyląg, M.; Kozak, B. Dark stains on rock surfaces in Driny Cave (Little Carpathian Mountains, Slovakia). Extremophiles 2016, 20, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wei, X.L.; Zhang, Y.Q.; Liu, H.-Y.; Yu, L.-Y. Diversity and distribution of lichen-associated fungi in the Ny-Ålesund Region (Svalbard, High Arctic) as revealed by pyrosequencing. Sci. Rep. 2015, 14, 14850. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Fedor, P. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’ Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef]
- Shannon, C.E.; Wiener, W. The Mathematical Theory of Communication; University Illinois Press: Urbana, IL, USA, 1963; p. 360. [Google Scholar]
- Workneh, F.; Van Bruggen, A.H.C. Microbial density, composition, and diversity in organically and conventionally managed rhizosphere soil in relation to suppression of corky root of tomatoes. Appl. Soil Ecol. 1994, 1, 219–230. [Google Scholar] [CrossRef]
- Srinivasan, R.; Prabhu, G.; Prasad, M.; Mishra, M.; Chaudhary, M.; Srivastava, R. Penicillium. In Beneficial Microbes in Agro-Ecology, 1st ed.; Amaresan, N., Kumar, M.S., Annapurna, K., Kumar, K., Sankaranarayanan, A., Eds.; Elservier: Amsderdam, The Netherlands, 2020; pp. 651–667. [Google Scholar] [CrossRef]
- Vrabl, P.; Siewert, B.; Winkler, J.; Schöbel, H.; Schinagl, C.W.; Knabl, L.; Orth-Höller, D.; Fiala, J.; Meijer, M.S.; Bonnet, S.; et al. Xanthoepocin, a photolabile antibiotic of Penicillium ochrochloron CBS 123823 with high activity against multiresistant gram-positive bacteria. Microb. Cell Factories 2022, 21, 1. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, H.; Sang, Z.; Qiu, K.; Wei, S.; Duan, F.; Zou, Z.; Tan, H. Two new secondary metabolites isolated from the fungus Penicillium virgatum T49-A. Fitoterapia 2023, 168, 105513. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Gong, Y.; Shi, Z.; Qi, C.; Chen, C.; Tong, Q.; Liu, J.; Wang, J.; Zhu, H.; Zhang, Y. Multioxidized aromatic polyketides produced by a soil-derived fungus Penicillium canescens. Phytochemistry 2022, 193, 113012. [Google Scholar] [CrossRef]
- Bazioli, J.M.; Amaral, L.D.S.; Fill, T.P.; Rodrigues-Filho, E. Insights into Penicillium brasilianum Secondary Metabolism and Its Biotechnological Potential. Molecules 2017, 22, 858. [Google Scholar] [CrossRef]
- Licorish, K.; Novey, H.S.; Kozak, P.; Fairshter, R.D.; Wilson, A.F. Role of Alternaria and Penicillium spores in the pathogenesis of asthma. J. Allergy Clin. Immunol. 1985, 76, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.J.; Straus, D.C. The roles of Penicillium and Aspergillus in sick building syndrome. Adv. Appl. Microbiol. 2004, 55, 215–238. [Google Scholar] [CrossRef] [PubMed]
- Sang, M.K.; Han, G.D.; Oh, J.Y.; Chun, S.C.; Kim, K.D. Penicillium brasilianum as a novel pathogen of onion (Allium cepa L.) and other fungi predominant on market onion in Korea. Crop Prot. 2014, 65, 138–142. [Google Scholar] [CrossRef]
- Kaplun, O.; Kekatos, P.; Creed, M.; Psevdos, G. Penicillium brasilianum Fungal Infection of Thumb Nail in a Patient Living with HIV-1. Infect. Dis. Clin. Pract. 2019, 27, e11. [Google Scholar] [CrossRef]
- Barcus, A.L.; Burdette, S.D.; Herchline, T.E. Intestinal invasion and disseminated disease associated with Penicillium chrysogenum. Ann. Clin. Microbiol. Antimicrob. 2005, 4, 21. [Google Scholar] [CrossRef] [PubMed]
- Shokouhi, S.; Tehrani, S.; Hemmatian, M. Mixed Pulmonary Infection with Penicillium notatum and Pneumocystis jiroveci in a Patient with Acute Myeloid Leukemia. Tanaffos 2016, 15, 53–56. [Google Scholar] [PubMed]
- Avilés-Robles, M.; Gómez-Ponce, C.; Reséndiz-Sánchez, J.; Rodríguez-Tovar, A.V.; Ceballos-Bocanegra, A.; Martínez-Rivera, Á. Disseminated penicilliosis due to Penicillium chrysogenum in a pediatric patient with Henoch–Schönlein syndrome. Int. J. Infect. Dis. 2016, 51, 78–80. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.V.M.; Corrêa-Moreira, D.; Mendes, T.V.; da Costa, G.L.; Vieira, R.M.; Buchele, C.M.N.; Lins, R.S.; Ferreira, A.B.T.B.C.; Veira, D.B.; Pedroso, R.S.A.; et al. First report of fungal meningoencephalitis by Penicillium chrysogenum in Brazil. Int. Infect. Dis. 2023, 126, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The Current Status of Its Application in Agriculture for the Biocontrol of Fungal Phytopathogens and Stimulation of Plant Growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Xie, W.; Zhang, J.; Hu, Q. Biodiversity of Entomopathogenic Fungi in the Soils of South China. Microorganisms 2019, 7, 311. [Google Scholar] [CrossRef] [PubMed]
- Razinger, J.; Lutz, M.; Schroers, H.J.; Urek, G.; Grunder, J. Evaluation of insect associated and plant growth promoting fungi in the control of cabbage root flies. J. Econ. Entomol. 2014, 107, 1348–1354. [Google Scholar] [CrossRef]
- Iqbal, M.; Dubey, M.; McEwan, K.; Menzel, U.; Franko, M.A.; Viketoft, M.; Jensen, D.F.; Karlsson, M. Evaluation of Clonostachys rosea for Control of Plant-Parasitic Nematodes in Soil and in Roots of Carrot and Wheat. Phytopathology 2018, 108, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, H.; Kandula, D.R.W.; Hampton, J.G.; Stewart, A.; Leung, D.W.M.; Edwards, Y.; Smith, C. Urease producing microorganisms under dairy pasture management in soils across New Zealand. Geoderma Reg. 2017, 11, 78–85. [Google Scholar] [CrossRef]
- Harm, G.F.S.; Papanicolaou, A.; Cuddy, W.S.; Park, R.F.; Moffitt, M.C. Draft Genome Sequence of the Fungus Lecanicillium psalliotae Strain HWLR35, Isolated from a Wheat Leaf Infected with Leaf Rust (Caused by Puccinia triticina). Genome Announc. 2018, 6, e01442-17. [Google Scholar] [CrossRef]
- Li, F.; Chen, L.; Redmile-Gordon, M.; Zhang, J.; Zhang, C.; Ning, Q.; Li, W. Mortierella elongata’s roles in organic agriculture and crop growth promotion in a mineral soil. Land Degrad. Dev. 2018, 29, 1642–1651. [Google Scholar] [CrossRef]
- Vandepol, N.; Liber, J.; Yocca, A.; Matlock, J.; Edger, P.; Bonito, G. Linnemannia elongata (Mortierellaceae) stimulates Arabidopsis thaliana aerial growth and responses to auxin, ethylene, and reactive oxygen species. PLoS ONE 2022, 17, e0261908. [Google Scholar] [CrossRef]
- Verekar, S.A.; Deshmukh, S.K. Keratinophilic fungi distribution, pathogenicity and biotechnological potentials. In Developments in Fungal Biology and Applied Mycology; Satyanarayana, T., Deshmukh, S., Johri, B., Eds.; Springer: Singapore, 2017; pp. 75–97. [Google Scholar]
- Călin, M.; Constantinescu-Aruxandei, D.; Alexandrescu, E.; Răut, I.; Doni, M.B.; Arsene, M.L.; Oancea, F.; Jecu, L.; Lazăr, V. Degradation of keratin substrates by keratinolytic fungi. Electron. J. Biotechnol. 2017, 28, 101–112. [Google Scholar] [CrossRef]
- Ali-Shtayeh, M.S.; Jamous, R.M. Keratinophilic fungi and related dermatophytes in polluted soil and water habitats. In Biology of Dermatophytes and other Keratinophilic Fungi; Kushwaha, R.K.S., Guarro, J., Eds.; Revista Iberoamericana de Micología: Bilbao, Spain, 2000; Volume 17, pp. 51–59. [Google Scholar]
- Ali-Shtayeh, M.S.; Arda, H.M. Isolation of keratinophilic fungi from floor dust in Arab elementary and preparatory schools in the West Bank of Jordan. Mycopathologia 1989, 106, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Moskaluk, A.E.; VandeWoude, S. Current topics in Dermatophyte classification and clinical diagnosis. Pathogens 2022, 11, 957. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Dukik, K.; Monod, M.; Packeu, A.; Stubbe, D.; Hendrickx, M.; Kupsch, C.; Stielow, J.B.; Freeke, J.; Göker, M.; et al. Toward a Novel Multilocus Phylogenetic Taxonomy for the Dermatophytes. Mycopathologia 2017, 182, 5–31. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, I.; Summerbell, R.C. The dermatophytes. Clin. Microbiol. Rev. 1995, 8, 240–259. [Google Scholar] [CrossRef]
- Shtayeh, M.S.A.; Arda, H.M. Incidence of dermatophytosis in Jordan with special reference to tinea capitis. Mycopathologia 1985, 92, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Cortez, A.C.A.; de Souza, J.V.B.; Sadahiro, A.; de Oliveira, J.A.A. Frequency and aetiology of dermatophytosis in children age 12 and under in the state of Amazonas, Brazil. Rev. Iberoam. Micol. 2012, 29, 223–226. [Google Scholar] [CrossRef]
- Mantovani, A. The role of animals in the epidemiology of the mycoses. Mycopathologia 1978, 65, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Bohacz, J.; Korniłłowicz-Kowalska, T. Species diversity of keratinophilic fungi in various soil types. Cent. Eur. J. Biol. 2012, 7, 259–266. [Google Scholar] [CrossRef]
- Mohanty, S.S.; Prakash, S. Comparative efficacy and pathogenicity of keratinophilic soil fungi against Culex quinquefasciatus larvae. Indian J. Microbiol. 2010, 50, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Ogórek, R.; Suchodolski, J.; Piecuch, A.; Przywara, K.; Višňovská, Z. Keratinophilic and Keratinolytic Fungi in Cave Ecosystems: A Culture-Based Study of Brestovská Cave and Demänovská Ľadová and Slobody Caves (Slovakia). Appl. Sci. 2022, 12, 1455. [Google Scholar] [CrossRef]
- Ginter, G. Ecology, epidemiology and clinical symptomatology of Microsporum gypseum infections. Mycoses 1989, 32, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Dolenc-Voljč, M.; Gasparič, J. Human infections with Microsporum gypseum complex (Nannizzia gypsea) in Slovenia. Mycopathologia 2017, 182, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Soankasina, A.H.; Rakotozandrindrainy, N.; Andrianteloasy, S.; Zafindraibe, N.J.; Rasamoelina, T.; Rafalimanana, C.; Cornet, M.; Razanakolona, L.R.; Rasamindrakotroka, A.; Andrianarivelo, M.R. Dermatophyte infection caused by Nannizzia gypsea: A rare case report from Madagascar. Med. Mycol. Case Rep. 2018, 20, 7–9. [Google Scholar] [CrossRef]
- Cruciani, D.; Papini, M.; Broccatelli, S.; Agnetti, F.; Spina, S.; Natalini, Y.; Crotti, S. Presumptive zoonotic kerion by Nannizzia gypsea: Case report. Front. Vet. Sci. 2021, 8, 718766. [Google Scholar] [CrossRef] [PubMed]
- Spiewak, R.; Szostak, W. Zoophilic and geophilic dermatophytoses among farmers and non-farmers in Eastern Poland. Ann. Agric. Environ. Med. 2000, 7, 125–129. [Google Scholar] [PubMed]
- Muir, D.B.; Pritchard, R.C.; Gregory, J.D. Dermatophytes identified at the Australian National Reference Laboratory in medical mycology 1966–1982. Pathology 1984, 16, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.Y.; Yao, Y.F.; Zhu, Y.F.; Zhang, Y.M.; Zhou, P.; Jin, Y.Q.; Zhang, B. Fungal spectrum identified by a new slide culture and in vitro drug susceptibility using Etest in fungal keratitis. Curr. Eye Res. 2005, 30, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Castro Lopez, N.; Casas, C.; Sopo, L.; Rojas, A.; Del Portillo, P.; Cepero de Garcia, M.C.; Restrepo, S. Fusarium species detected in onychomycosis in Colombia. Mycoses 2009, 52, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Segal, B.H.; Walsh, T.J.; Liu, J.M.; Wilson, J.D.; Kwon-Chung, K.J. Invasive infection with Fusarium chlamydosporum in a patient with aplastic anemia. J. Clin. Microbiol. 1998, 36, 1772–1776. [Google Scholar] [CrossRef] [PubMed]
- Parihar, T.J.; Sofi, M.Y.; Rasool, R.S.; Khursheed, S.; Bhat, Z.A.; Hussain, K.; Dhekale, B.; Zargar, S.M.; Hakak, A.S.; Shah, M.D.; et al. Fusarium chlamydosporum, causing wilt disease of chili (Capsicum annum L.) and brinjal (Solanum melongena L.) in Northern Himalayas: A first report. Sci. Rep. 2022, 12, 20392. [Google Scholar] [CrossRef]
- Liu, F.; Cai, L. Morphological and molecular characterization of a novel species of Simplicillium from China. Cryptogam. Mycol. 2012, 33, 137–144. [Google Scholar] [CrossRef]
- Chen, R.S.; Huang, C.C.; Li, J.C.; Tsay, J.G. First report of Simplicillium lanosoniveum causing brown spot on Salvinia auriculata and S. molesta in Taiwan. Plant Dis. 2008, 92, 1589. [Google Scholar] [CrossRef]
- Chen, W.H.; Han, Y.F.; Liang, J.D.; Liang, Z.Q. Taxonomic and phylogenetic characterizations reveal four new species of Simplicillium (Cordycipitaceae, Hypocreales) from Guizhou, China. Sci. Rep. 2021, 11, 15300. [Google Scholar] [CrossRef]
- Teasdale, S.E.; Caradus, J.R.; Johnson, L.J. Fungal endophyte diversity from tropical forage grass Brachiaria. Plant Ecol. Divers. 2018, 11, 611–624. [Google Scholar] [CrossRef]
- Zhu, M.; Duan, X.; Cai, P.; Li, Y.F.; Qiu, Z. Deciphering the genome of Simplicillium aogashimaense to understand its mechanisms against the wheat powdery mildew fungus Blumeria graminis f. sp. tritici. Phytopathol. Res. 2022, 4, 16. [Google Scholar] [CrossRef]
- Mazurkiewicz-Zapałowicz, K.; Wróbel, J.; Janowicz, K. Influence of Selected Soil Saprotrophes on Gas Exchange, Growth and Yield of Solanum Tuberosum. Acta Physiol. Plant 2004, 26, 157–164. [Google Scholar] [CrossRef]
- Semenov, A.M.; Batomunkueva, B.P.; Nizovtseva, D.V.; Panikov, N.S. Method of determination of cellulase activity in soils and in microbial cultures, and its calibration. J. Microbiol. Methods 1996, 24, 259–267. [Google Scholar] [CrossRef]
- Hopsu-Havu, V.K.; Sonck, C.E.; Tunnela, E. Production of Elastase by Pathogenic and Non-Pathogenic Fungi. Mycoses 2009, 15, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Ali-Shtayeh, M.S. Keratinophilic fungi isolated from children’s sandpits in the Nablus area, West Bank of Jordan. Mycopathologia 1988, 103, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Javoreková, S.; Labuda, R.; Maková, J.; Novák, J.; Medo, J.; Majerčíková, K. Keratinophilic fungi isolated from soils of long-term fold-grazed, degraded pastures in national parks of Slovakia. Mycopathologia 2012, 174, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Cavello, I.A.; Hours, R.A.; Cavalitto, S.F. Bioprocessing of “hair waste” by Paecilomyces lilacinus as a source of a bleach-stable, alkaline, and thermostable keratinase with potential application as a laundry detergent additive: Characterization and wash performance analysis. Biotechnol. Res. Int. 2012, 2012, 369308. [Google Scholar] [CrossRef] [PubMed]
- Kotwal, S.; Sumbali, G. Preferential utilization and colonization of keratin baits by different myco-keratinophiles. Springerplus 2016, 5, 1–6. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, H.S.; Adhikari, M.; Yadav, D.R.; Kim, S.W.; Um, Y.H.; Lee, H.B.; Lee, Y.S. First report of Metacordyceps chlamydosporia (Cordyceps chlamydosporia) isolated from soil in Korea. Korean J. Mycol. 2016, 44, 48–50. [Google Scholar] [CrossRef]
- Scott, J.A.; Untereiner, W.A. Determination of keratin degradation by fungi using keratin azure. Med. Mycol. 2004, 42, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Gradišar, H.; Friedrich, J.; Krizaj, I.; Jerala, R. Similarities and specificities of fungal keratinolytic proteases: Comparison of keratinases of Paecilomyces marquandii and Doratomyces microsporus to some known proteases. Appl. Environ. Microbiol. 2005, 71, 3420–3426. [Google Scholar] [CrossRef] [PubMed]
- Hamm, P.S.; Mueller, R.C.; Kuske, C.R.; Porras-Alfaro, A. Keratinophilic fungi: Specialized fungal communities in a desert ecosystem identified using cultured-based and Illumina sequencing approaches. Microbiol. Res. 2020, 239, 126530. [Google Scholar] [CrossRef]
- Friedrich, J.; Gradišar, H.; Mandin, D.; Chaumont, J.P. Screening fungi for synthesis of keratinolytic enzymes. Lett. Appl. Microbiol. 1999, 28, 127–130. [Google Scholar] [CrossRef]
- Sun, Z.B.; Li, S.D.; Ren, Q.; Xu, J.L.; Lu, X.; Sun, M.H. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Kuzikova, I.L.; Medvedeva, N.G. Opportunistic Fungi as Contaminants of Human Environment and Their Potential Pathogenicity. Ekol. Cheloveka (Hum. Ecol.) 2021, 3, 4–14. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Sites | Name of Location | Latitude (N) | Longitude (E) | Type of Usage | Predominant Vegetation | Accessibility for Animals |
|---|---|---|---|---|---|---|
| I | “Gaj” Allotment Garden | 51°07′75″ | 17°05′52″ | Allotment garden | Useful plants | Occasionally |
| II | Słowiańska Hill | 51°12′48″ | 17°04′35″ | Other green areas | Deciduous trees | Yes |
| III | Dąbrowskiego Street | 51°10′11″ | 17°04′19″ | Other green areas | Grass and individual deciduous trees | Occasionally |
| IV | Staszic Park | 51°12′42″ | 17°03′07″ | Park | Deciduous trees | Yes |
| V | Bełza Square | 51°14′19″ | 17°05′45″ | Other green areas | Deciduous trees | Yes |
| VI | Słoneczny Park | 51°08′06″ | 17°04′41″ | Park | Deciduous trees | Yes |
| VII | Allotment “Pod Topolami” | 51°06′98″ | 17°02′85″ | Allotment garden | Useful plants | Occasionally |
| VIII | Tołpa Park | 51°12′05″ | 17°05′39″ | Park | Deciduous trees | Yes |
| IX | Skowroni Park | 51°08′04″ | 17°02′74″ | Park | Deciduous trees | Yes |
| X | Na Niskich Łąkach Park | 51°09′88″ | 17°.05′79″ | Park | Deciduous trees | Yes |
| XI | Grabiszyński Park | 51°08′79″ | 16°97′88″ | Park | Deciduous trees | Yes |
| XII | Południowy Park | 51°07′28″ | 17°01′45″ | Park | Deciduous trees | Yes |
| Fungi Obtained in the Study | Identity with Sequence from GenBank | |||||||
|---|---|---|---|---|---|---|---|---|
| Isolate Number | Identified Species | Phylum | Family | Isolation Method | GenBank Accession No. | The Sequence Length (bp) | Identity, % | Accession |
| UWR_390 | Arthroderma uncinatum | Ascomycota | Arthrodermataceae | V | PP034006 | 575 | 100 | KP132800.1 |
| UWR_391 | Aspergillus flavus | Aspergillaceae | V, P | PP034007 | 511 | 100 | OK663515.1 | |
| UWR_392 | Clonostachys rosea | Bionectriaceae | P | PP034008 | 358 | 100 | MT447494.1 | |
| UWR_393 | Clonostachys solani | Bionectriaceae | P | PP034009 | 393 | 100 | MH860460.1 | |
| UWR_394 | Fusarium chlamydosporum | Nectriaceae | V, P | PP034010 | 483 | 100 | MN737771.1 | |
| UWR_395 | Fusarium oxysporum | Nectriaceae | V, P | PP034011 | 415 | 100 | OQ155213.1 | |
| UWR_396 | V, P | PP034012 | 389 | 99.49 | MT557539.1 | |||
| UWR_397 | Fusarium solani | Nectriaceae | V, P | PP034013 | 435 | 100 | KT876722.1 | |
| UWR_398 | V, P | PP034014 | 426 | 100 | MT560378.1 | |||
| UWR_399 | V, P | PP034015 | 361 | 100 | MT557344.1 | |||
| UWR_400 | Keratinophyton wagneri | Onygenaceae | V | PP034016 | 542 | 100 | MT903275.1 | |
| UWR_401 | Lecanicillium psalliotae | Cordycipitaceae | P | PP034017 | 450 | 100 | AB360366.1 | |
| UWR_402 | Linnemannia elongata | Mucoromycota | Mortierellaceae | P | PP034018 | 556 | 100 | MH860032.1 |
| UWR_403 | Marquandomyces marquandii | Ascomycota | Clavicipitaceae | V, P | PP034019 | 377 | 100 | MH063648.1 |
| UWR_404 | Metacordyceps chlamydosporia | Clavicipitaceae | V | PP034020 | 491 | 100 | AB378544.1 | |
| UWR_405 | Nannizzia gypsea | Arthrodermataceae | V | PP034021 | 585 | 100 | MH858847.1 | |
| UWR_406 | Paraphyton cookei | Arthrodermataceae | V | PP034022 | 583 | 100 | OW984928.1 | |
| UWR_407 | Penicillium brasilianum | Aspergillaceae | P | PP034023 | 442 | 100 | MK543248.1 | |
| UWR_408 | Penicillium canescens | Aspergillaceae | P | PP034024 | 493 | 100 | KY684281.1 | |
| UWR_409 | Penicillium chrysogenum | Aspergillaceae | P | PP034025 | 512 | 100 | MT312739.1 | |
| UWR_410 | Penicillium cosmopolitanum | Aspergillaceae | P | PP034026 | 444 | 100 | MT573489.1 | |
| UWR_411 | Penicillium manginii | Aspergillaceae | P | PP034027 | 465 | 100 | MK390495.1 | |
| UWR_412 | Penicillium ochrochloron | Aspergillaceae | P | PP034028 | 512 | 100 | MK304092.1 | |
| UWR_413 | Penicillium roseopurpureum | Aspergillaceae | P | PP034029 | 506 | 100 | MK179263.1 | |
| UWR_414 | Aspergillaceae | P | PP034030 | 455 | 100 | MN206951.1 | ||
| UWR_415 | Penicillium scabrosum | Aspergillaceae | P | PP034031 | 501 | 100 | MT995062.1 | |
| UWR_416 | Penicillium subrubescens | Aspergillaceae | P | PP034032 | 500 | 100 | OW984373.1 | |
| UWR_417 | Penicillium virgatum | Aspergillaceae | P | PP034033 | 421 | 100 | KY989162.1 | |
| UWR_418 | Pochonia bulbillosa | Clavicipitaceae | P | PP034034 | 440 | 99.77 | AB709835.1 | |
| UWR_419 | Purpureocillium lilacinum | Ophiocordycipitaceae | V, P | PP034035 | 520 | 100 | MT254824.1 | |
| UWR_420 | V, P | PP034036 | 459 | 100 | MT279298.1 | |||
| UWR_421 | V, P | PP034037 | 535 | 100 | MT420635.1 | |||
| UWR_422 | V, P | PP034038 | 396 | 100 | MT446187.1 | |||
| UWR_423 | Simplicillium aogashimaense | Cordycipitaceae | V | PP034039 | 491 | 100 | MK579180.1 | |
| UWR_424 | Trichocladium asperum | Chaetomiaceae | V | PP034040 | 488 | 100 | MN643061.1 | |
| UWR_425 | Trichoderma hamatum | Hypocreaceae | P | PP034041 | 487 | 100 | MF383138.1 | |
| UWR_426 | Trichoderma virens | Hypocreaceae | P | PP034042 | 509 | 100 | MK459332.1 | |
| Fungi Species | Fungi Isolate | Study Sites | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | ||
| Arthroderma uncinatum | UWR_390 | + | + | − | + | + | − | + | + | + | + | − | + |
| Aspergillus flavus | UWR_391 | − | − | − | − | − | − | − | − | + | − | − | − |
| Fusarium chlamydosporum | UWR_394 | + | + | − | − | + | + | + | + | − | − | + | + |
| Fusarium oxysporum | UWR_395 | − | − | − | − | + | − | − | − | + | + | − | − |
| UWR_396 | − | − | − | − | − | − | − | + | − | + | − | − | |
| Fusarium solani | UWR_397 | − | − | − | − | − | − | − | − | − | − | − | + |
| UWR_398 | − | + | − | − | − | − | − | − | − | − | − | − | |
| UWR_399 | − | + | + | − | − | − | + | − | − | − | − | − | |
| Keratinophyton wagneri | UWR_400 | − | + | + | + | + | − | + | + | + | + | − | + |
| Marquandomyces marquandii | UWR_403 | − | + | − | − | + | − | + | − | − | − | − | + |
| Metacordyceps chlamydosporia | UWR_404 | − | − | − | − | + | − | − | − | − | + | + | + |
| Nannizzia gypsea | UWR_405 | + | + | − | + | + | + | − | − | − | − | + | + |
| Paraphyton cookei | UWR_406 | − | − | − | − | − | − | − | − | − | − | + | − |
| Purpureocillium lilacinum | UWR_419 | + | − | − | − | − | − | − | − | − | − | − | − |
| UWR_420 | + | − | + | − | + | − | − | − | − | − | − | − | |
| UWR_421 | + | + | − | + | − | − | − | − | − | − | − | − | |
| UWR_422 | − | + | − | − | − | − | − | − | − | − | − | − | |
| Simplicillium aogashimaense | UWR_423 | + | − | + | + | + | + | + | + | + | − | + | − |
| Trichocladium asperum | UWR_424 | − | − | − | − | − | − | − | − | + | + | − | − |
| Fungi Species | Fungi Isolate | Study Sites | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | ||
| Aspergillus flavus | UWR_391 | − | − | − | − | − | − | − | − | + | − | − | − |
| Clonostachys rosea | UWR_392 | − | − | − | − | − | − | + | − | − | + | − | + |
| Clonostachys solani | UWR_393 | − | − | − | − | − | − | − | − | − | − | + | + |
| Fusarium chlamydosporum | UWR_394 | + | + | − | − | + | + | + | + | − | − | + | + |
| Fusarium oxysporum | UWR_395 | − | − | − | − | + | − | − | − | + | + | − | − |
| UWR_396 | − | − | − | − | − | − | − | + | − | + | − | − | |
| Fusarium solani | UWR_397 | − | − | − | − | − | − | − | − | − | − | − | + |
| UWR_398 | − | + | − | − | − | − | − | − | − | − | − | − | |
| UWR_399 | − | + | + | − | − | − | + | − | − | − | − | − | |
| Lecanicillium psalliotae | UWR_401 | − | − | − | + | − | − | − | − | − | − | − | + |
| Linnemannia elongata | UWR_402 | + | + | + | − | − | − | − | + | + | + | − | + |
| Marquandomyces marquandii | UWR_403 | − | + | − | − | + | − | + | − | − | − | − | + |
| Penicillium brasilianum | UWR_407 | + | + | + | − | + | + | + | − | + | + | − | + |
| Penicillium canescens | UWR_408 | − | − | + | − | − | − | − | − | − | − | − | − |
| Penicillium chrysogenum | UWR_409 | − | − | − | − | − | − | + | − | + | − | − | − |
| Penicillium cosmopolitanum | UWR_410 | + | + | − | + | − | + | − | − | + | − | − | − |
| Penicillium manginii | UWR_411 | − | − | − | − | − | − | − | − | − | − | − | + |
| Penicillium ochrochloron | UWR_412 | + | − | − | + | − | − | + | − | − | − | + | − |
| Penicillium roseopurpureum | UWR_413 | + | − | − | − | − | − | − | − | − | + | − | − |
| UWR_414 | + | − | + | − | − | − | − | − | − | + | − | + | |
| Penicillium scabrosum | UWR_415 | − | − | − | − | − | + | − | − | − | − | − | − |
| Penicillium subrubescen | UWR_416 | − | − | + | − | − | + | + | + | − | − | − | − |
| Penicillium virgatum | UWR_417 | − | − | − | − | − | − | − | − | − | − | − | + |
| Pochonia bulbillosa | UWR_418 | − | − | + | − | − | + | − | − | + | − | + | + |
| Purpureocillium lilacinum | UWR_419 | + | − | − | − | − | − | − | − | − | − | − | − |
| UWR_420 | + | − | + | − | + | − | − | − | − | − | − | − | |
| UWR_421 | + | + | − | + | − | − | − | − | − | − | − | − | |
| UWR_422 | − | + | − | − | − | − | − | − | − | − | − | − | |
| Trichoderma hamatum | UWR_425 | − | − | − | − | − | − | − | − | + | − | − | + |
| Trichoderma virens | UWR_426 | − | − | + | + | − | − | − | − | − | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spychała, K.; Kłosińska, K.; Salwińska, W.; Ogórek, R. Diversity of Soil-Borne Fungi Isolated from Places Frequently Visited by People in the City of Wrocław (Poland). Appl. Sci. 2024, 14, 2782. https://doi.org/10.3390/app14072782
Spychała K, Kłosińska K, Salwińska W, Ogórek R. Diversity of Soil-Borne Fungi Isolated from Places Frequently Visited by People in the City of Wrocław (Poland). Applied Sciences. 2024; 14(7):2782. https://doi.org/10.3390/app14072782
Chicago/Turabian StyleSpychała, Klaudyna, Katarzyna Kłosińska, Weronika Salwińska, and Rafał Ogórek. 2024. "Diversity of Soil-Borne Fungi Isolated from Places Frequently Visited by People in the City of Wrocław (Poland)" Applied Sciences 14, no. 7: 2782. https://doi.org/10.3390/app14072782
APA StyleSpychała, K., Kłosińska, K., Salwińska, W., & Ogórek, R. (2024). Diversity of Soil-Borne Fungi Isolated from Places Frequently Visited by People in the City of Wrocław (Poland). Applied Sciences, 14(7), 2782. https://doi.org/10.3390/app14072782