
Effect of Temperature on the Nutritional Quality and Growth Parameters of Yellow Mealworm (Tenebrio molitor L.): A Preliminary Study


Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Samples
2.2. Analytical Methods
2.3. Statistical Analysis
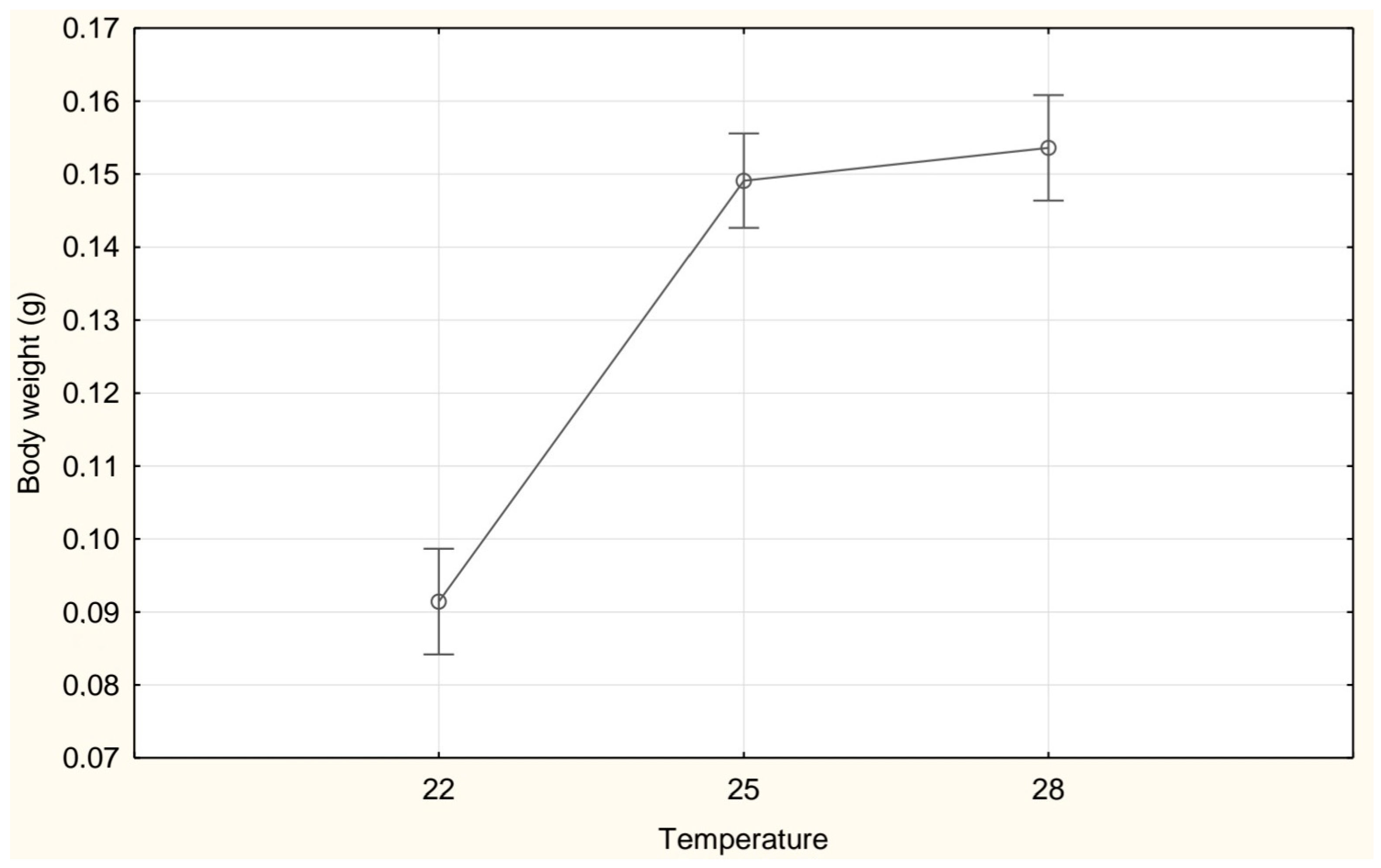
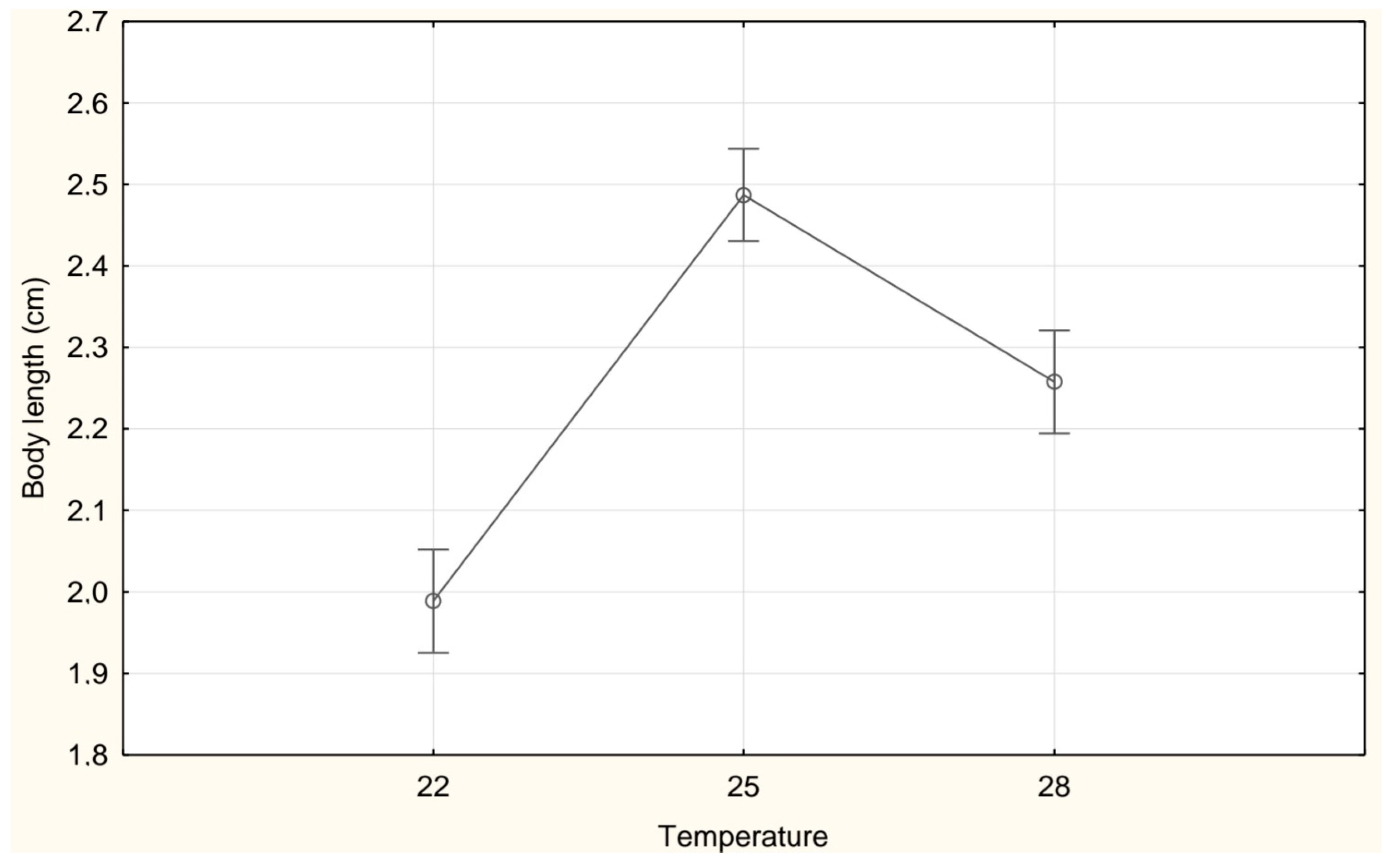
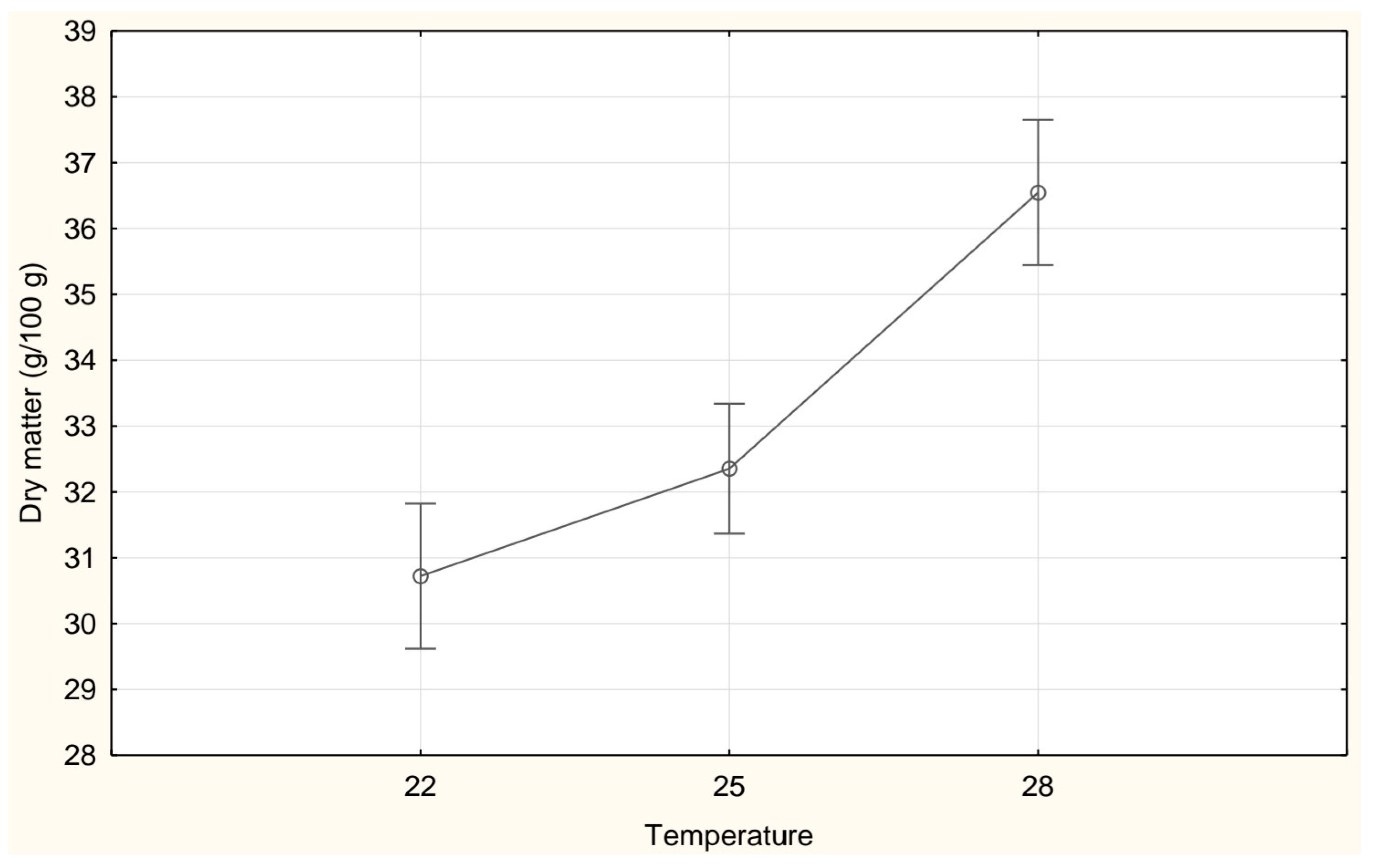
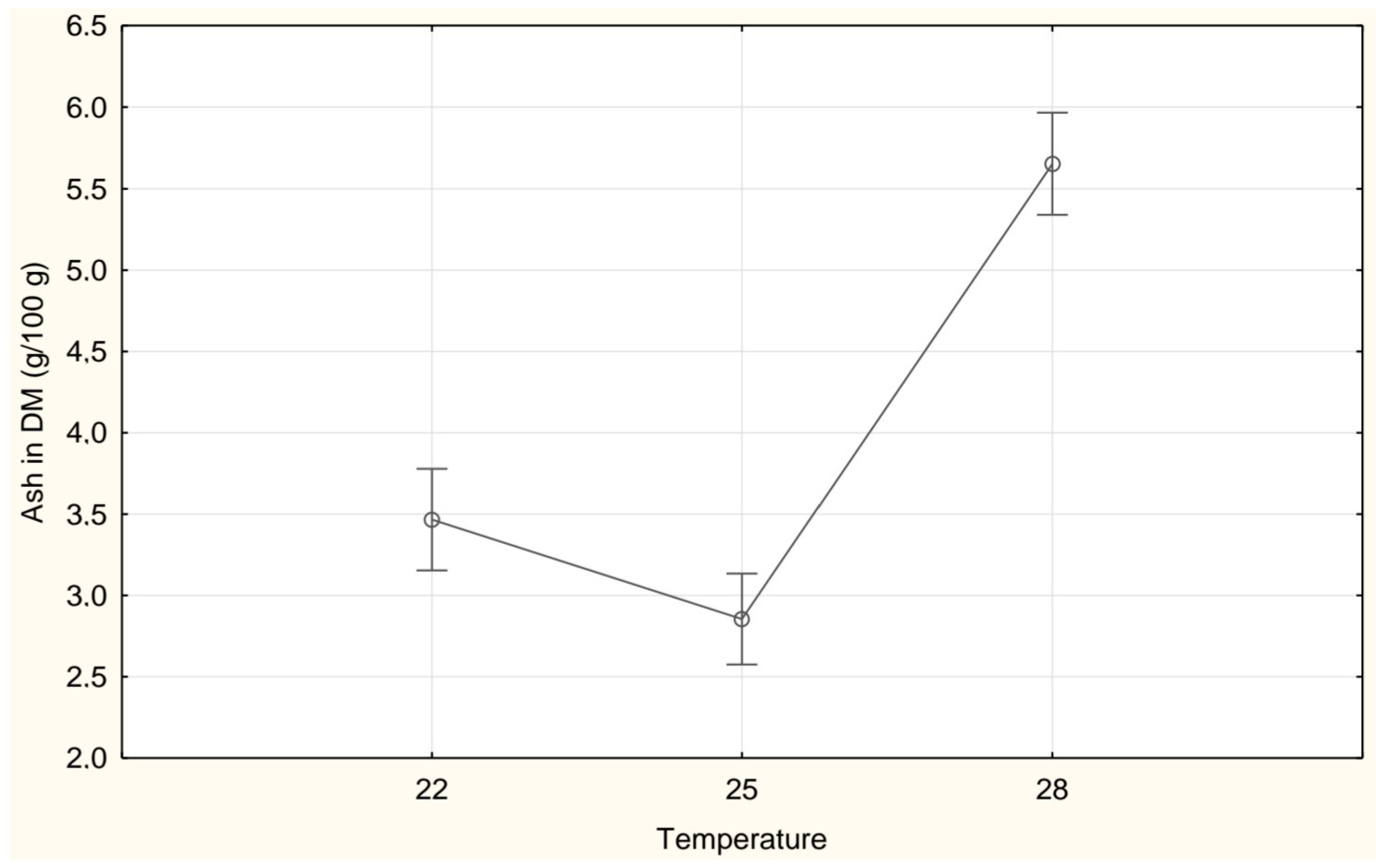
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van Huis, A.; van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Proceedings of the Nutrition Society; FAO: Rome, Italy, 2013; Volume 171, ISBN 9789251075951. [Google Scholar]
- Imathiu, S. Benefits and food safety concerns associated with consumption of edible insects. NFS J. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Kulma, M.; Kouřimská, L.; Plachý, V.; Božik, M.; Adámková, A.; Vrabec, V. Effect of sex on the nutritional value of house cricket, Acheta domestica L. Food Chem. 2019, 272, 267–272. [Google Scholar] [CrossRef] [PubMed]
- da Silva Lucas, A.J.; Menegon, O.L.; da Rocha, M.; Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chem. 2020, 311, 1266022. [Google Scholar] [CrossRef]
- Orkusz, A. Edible Insects versus Meat—Nutritional Comparison: Knowledge of Their Composition Is the Key to Good Health. Nutrients 2021, 13, 1207. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on novel foods, amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001. Off. J. Eur. Union 2015, 327, 1–22. [Google Scholar]
- Gravel, A.; Doyen, A. The use of edible insect proteins in food: Challenges and issues related to their functional properties. Innov. Food Sci. Emerg. Technol. 2020, 59, 102272. [Google Scholar] [CrossRef]
- Patel, S.; Suleria, H.; Rauf, A. Edible insects as innovative foods: Nutritional and functional assessments. Trends Food Sci. Technol. 2019, 86, 352–359. [Google Scholar] [CrossRef]
- Shafique, L.; Abdel-Latif, H.; Hassan, F.; Alagawany, M.; Naiel, M.; Dawood, M.; Yilmaz, S.; Liu, Q. The Feasibility of Using Yellow Mealworms (Tenebrio molitor): Towards a Sustainable Aquafeed Industry. Animals 2021, 11, 811. [Google Scholar] [CrossRef]
- Bjørge, J.D.; Overgaard, J.; Malte, H.; Gianotten, N.; Heckmann, L.H. Role of temperature on growth and metabolic rate in the tenebrionid beetles Alphitobius diaperinus and Tenebrio molitor. J. Insect Physiol. 2018, 107, 89–96. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, D.; Zhou, S.; Duan, H.; Guo, J.; Yan, W. Nutritional Composition, Health Benefits, and Application Value of Edible Insects: A Review. Foods 2022, 11, 3961. [Google Scholar] [CrossRef] [PubMed]
- Oibiokpa, F.; Akanya, H.; Jigam, A.; Saidu, A.; Egwim, E. Protein quality of four indigenous edible insect species in Nigeria. Food Sci. Hum. Wellness 2018, 7, 175–183. [Google Scholar] [CrossRef]
- Adámková, A.; Mlček, J.; Adámek, M.; Borkovcová, M.; Bednářová, M.; Hlobilová, V.; Knížková, I.; Juríková, T. Tenebrio molitor (Coleoptera: Tenebrionidae)—Optimization of Rearing Conditions to Obtain Desired Nutritional Values. J. Insect Sci. 2020, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- Siemanowska, E.; Kosewska, A.; Aljewicz, M.; Skibniewska, K.; Polak-Juszczak, L.; Jedras, M. Larvae of mealworm (Tenebrio molitor L.) as European novel food. Agric. Sci. 2013, 4, 287–291. [Google Scholar] [CrossRef]
- Ghosh, S.; Lee, S.; Jung, C.; Meyer-Rochow, V. Nutritional composition of five commercial edible insects in South Korea. J. Asia-Pac. Entomol. 2017, 20, 686–694. [Google Scholar] [CrossRef]
- van Huis, A. Edible insects are the future? Proc. Nutr. Soc. 2016, 75, 294–305. [Google Scholar] [CrossRef]
- Varelas, V. Food Wastes as a Potential new Source for Edible Insect Mass Production for Food and Feed: A review. Fermentation 2019, 5, 81. [Google Scholar] [CrossRef]
- Eberle, S.; Schaden, L.; Tinter, J.; Stauffer, C.; Schebeck, M. Effect of Temperature and Photoperiod on Development, Survival, and Growth Rate of Mealworms. Tenebrio molitor. Insects 2022, 13, 321. [Google Scholar] [CrossRef]
- Kulma, M.; Kouřimská, L.; Homolková, D.; Božik, M.; Plachý, V.; Vrabec, V. Effect of developmental stage on the nutritional value of edible insects. A case study with Blaberus craniifer and Zophobas morio. J. Food Compos. Anal. 2020, 92, 103507. [Google Scholar] [CrossRef]
- ISO 5983-1:2005; Animal feeding stuffs–Determination of nitrogen content and calculation of crude protein content–Part 1: Kjeldahl method. International Organization for Standardization: Geneva, Switzerland, 2005.
- ISO 1871:2009; Food and feed products–General guidelines for the determination of nitrogen by the Kjeldahl method. International Organization for Standardization: Geneva, Switzerland, 2009.
- ISO 13903:2005; Animal Feeding Stuffs—Determination of Amino Acids Content. International Organization for Standardization: Geneva, Switzerland, 2005.
- Wu, S.; Lin, H.; Li, M.; Tang, X. Determination of some important technique parameters in the course of breeding Tenebrio molitor. J. Econ. Anim. 2009, 13, 28–31. [Google Scholar]
- Ribeiro, N.; Abelho, M.; Costa, R. A Review of the Scientific Literature for Optimal Conditions for Mass Rearing Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2018, 53, 434–454. [Google Scholar] [CrossRef]
- Laštůvka, Z. Climate Change and Its Possible Influence on the Occurrence and Importance of Insect Pests. Plant Protect. Sci. 2009, 49, S53–S62. [Google Scholar] [CrossRef]
- Grau, T.; Vilcinskas, A.; Joop, G. Sustainable farming of the mealworm Tenebrio molitor for the production of food and feed. Z. Naturforschung C 2017, 72, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Rumbos, C.; Karapanagiotidis, I.; Mente, E.; Psofakis, P.; Athanassiou, C. Evaluation of various commodities for the development of the yellow mealworm. Tenebrio molitor. Sci. Rep. 2020, 10, 11224. [Google Scholar] [CrossRef] [PubMed]
- Bednářová, M.; Borkovcová, M.; Zorníková, G.; Zeman, L. Insect as a food in Czech Republic. Mendelnet 2010, 24, 674–682. [Google Scholar]
- Baek, M.; Kim, M.-A.; Kwon, Y.-S.; Hwang, J.-S.; Goo, T.-W.; Jun, M.; Yun, E.-Y. Effects of processing methods on nutritional composition and antioxidant activity of mealworm (Tenebrio molitor) larvae. Entomol. Res. 2019, 49, 284–293. [Google Scholar] [CrossRef]
- Yi, L.; Lakemond, C.; Sagis, L.; Eisner-Schadler, V.; van Huis, A.; Boekel, M. Extraction and characterisation of protein fractions from five insect species. Food Chem. 2013, 141, 3341–3348. [Google Scholar] [CrossRef]
- Bosch, G.; Zhang, S.; Oonincx, D.; Hendricks, W. Protein quality of insects as potential ingredients for dog and cat foods. J. Nutr. Sci. 2014, 3, 29. [Google Scholar] [CrossRef]
- Gkinali, A.; Matsakidou, A.; Vasileiou, E.; Paraskevopoulou, A. Potentiality of Tenebrio molitor larva-based ingredients for the food industry: A review. Trends Food Sci. Technol. 2020, 119, 495–507. [Google Scholar] [CrossRef]
- Adámková, A.; Adámek, M.; Mlček, J.; Borkovcová, M.; Bednářová, M.; Kouřimská, L.; Skácel, J.; Vítová, E. Welfare of the mealworm (Tenebrio molitor) breeding with regard to nutrition value and food safety. Potravin. Slovak J. Food Sci. 2017, 11, 460–465. [Google Scholar] [CrossRef]
- Azagoh, C.; Ducept, F.; Garcia, R.; Rakotozafy, L.; Cuvelier, M.; Keller, S.; Lewandowski, R.; Mezdour, S. Extraction and physicochemical characterization of Tenebrio molitor proteins. Food Res. J. 2016, 88, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Han, T.; Kim, Y. Mealworm (Tenebrio molitor Larvae) as an Alternative Protein Source for Monogastric Animal: A Review. Animals 2020, 10, 2068. [Google Scholar] [CrossRef] [PubMed]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Lehtovaara, V.; Roininen, H.; Valtonen, A. Optimal Temperature for Rearing the Edible Ruspolia differens (Orthoptera: Tettigoniidae). J. Econ. Entomol. 2018, 111, 6. [Google Scholar] [CrossRef]
- Finke, M. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Wu, R.A.; Ding, Q.; Yin, L.; Chi, X.; Sun, N.; He, R.; Luo, L.; Ma, H.; Li, Z. Comparison of the nutritional value of mysore thorn borer (Anoplophora chinensis) and mealworm larva (Tenebrio molitor): Amino acid, fatty acid, and element profiles. Food Chem. 2020, 323, 126818. [Google Scholar] [CrossRef] [PubMed]
- Rumpold, B.; Shlüter, O. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M.; Rybczyńska, K.; Jakubczyk, A. Selected species of edible insects as a source of nutrient composition. Food Res. J. 2015, 77, 460–466. [Google Scholar] [CrossRef]
- Velíšek, J. The Chemistry of Food, 1st ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; p. 1124. [Google Scholar]
- Costa, S.; Pedro, S.; Lourenco, H.; Batista, I.; Teixera, B.; Bandarra, N.; Murta, D.; Nunes, R.; Pires, C. Evaluation of Tenebrio molitor larvae as an alternative food source. NFS J. 2020, 21, 57–64. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | 22 °C | 25 °C | 28 °C |
|---|---|---|---|
| Weight (g) | 0.09 ± 0.03 a | 0.15 ± 0.03 b | 0.15 ± 0.04 c |
| Length (cm) | 1.99 ± 0.40 a | 2.49 ± 0.20 c | 2.26 ± 0.30 b |
| Dry matter (g/100 g FW) | 30.72 ± 1.29 a | 32.35 ± 2.70 b | 36.55 ± 0.79 c |
| Ash (g/100 g DM) | 3.47 ± 0.44 b | 2.85 ± 0.73 a | 5.65 ± 0.26 c |
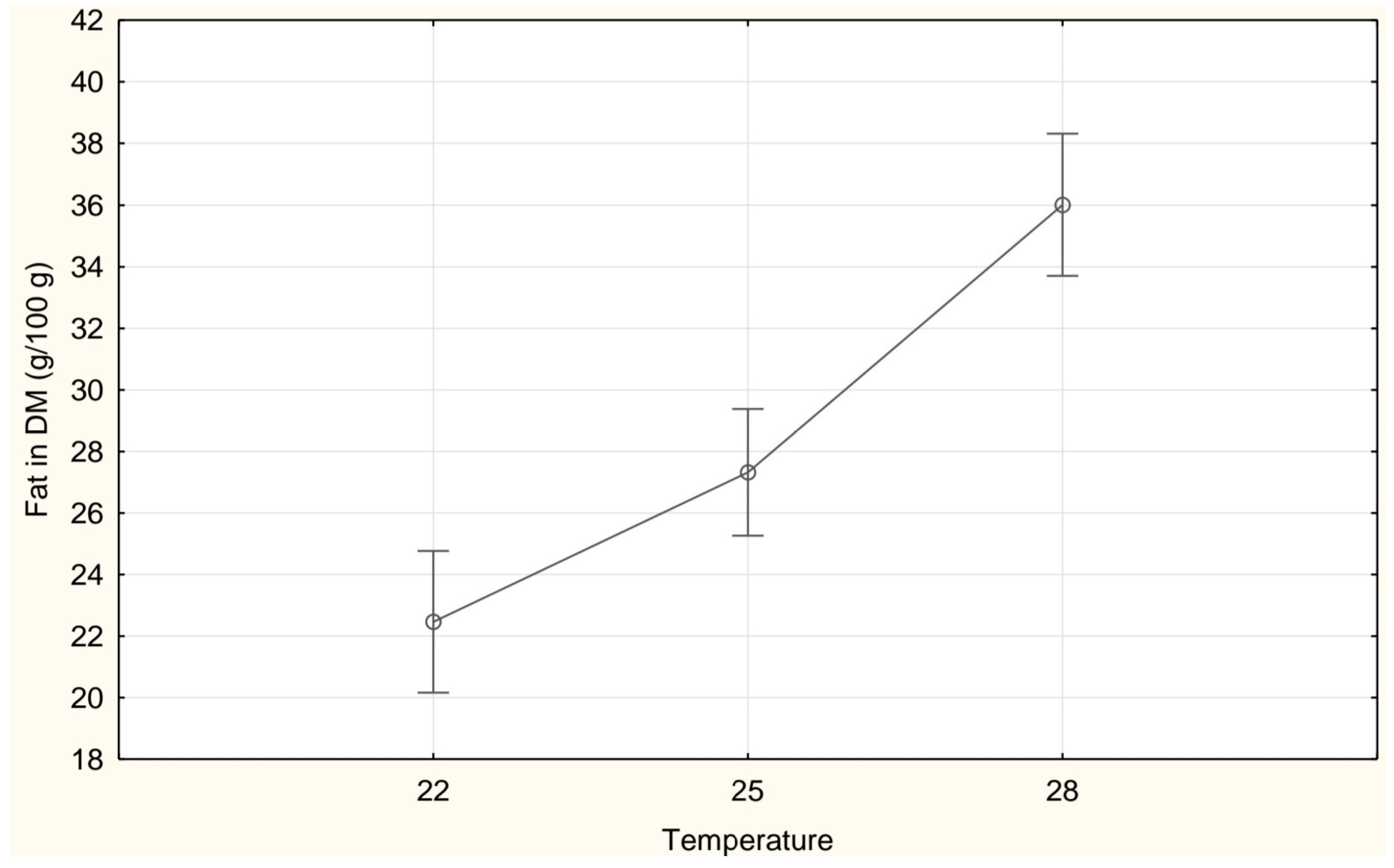
| Fat (g/100 g DM) | 22.46 ± 3.54 a | 27.32 ± 4.62 b | 36.01 ± 3.31 c |
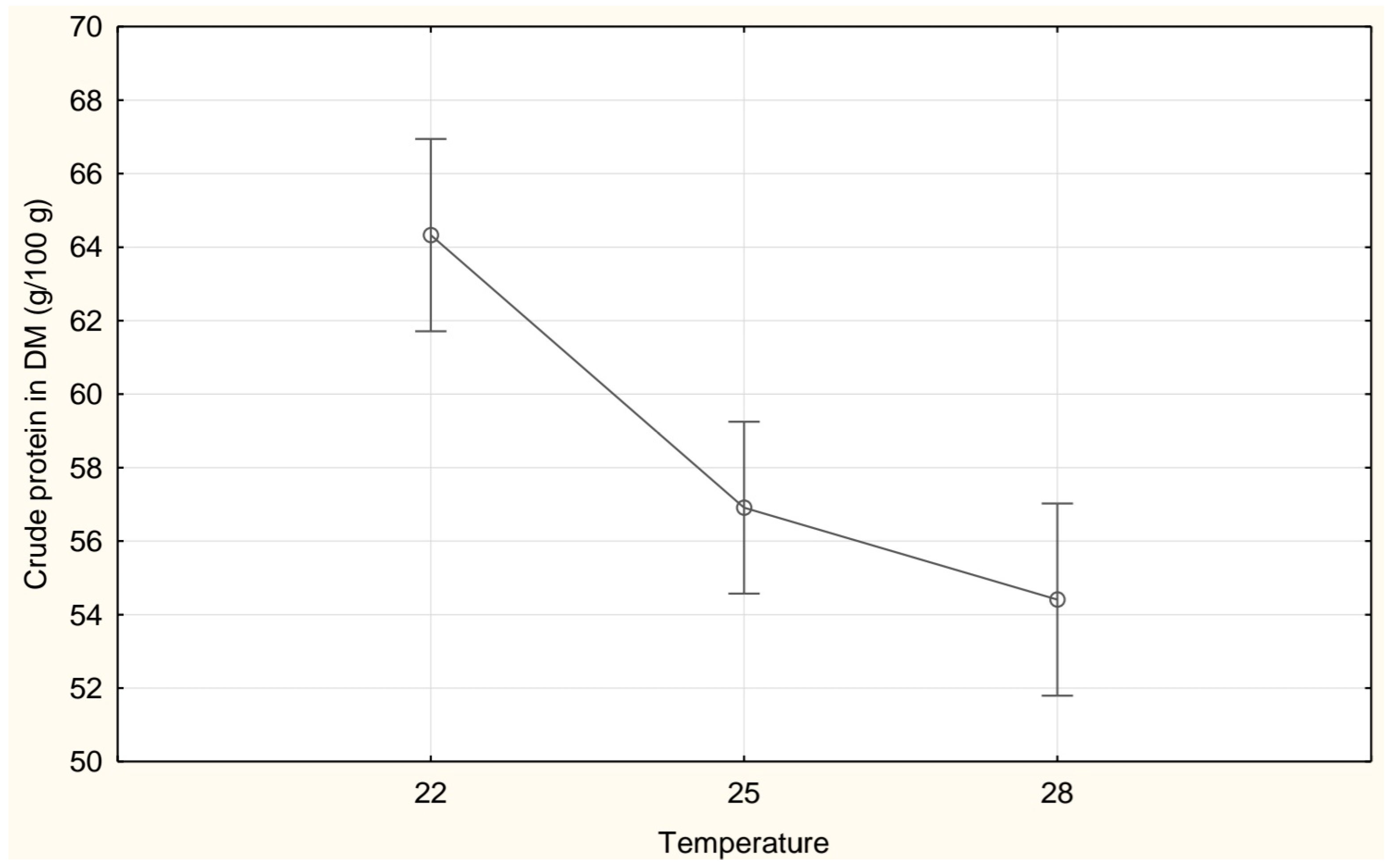
| Crude protein (g/100 g DM) | 64.34 ± 2.98 c | 56.91 ± 6.29 b | 54.41 ± 2.47 a |
| First emergence of larvae (days) | 10.00 ± 2.87 a | 7.50 ± 5.54 ab | 5.78 ± 1.69 b |
| Terminal larval stage (days) | 107.66 ± 12.10 a | 84.41 ± 13.18 b | 47.43 ± 6.61 c |
| Amino Acid | 22 °C | 25 °C | 28 °C |
|---|---|---|---|
| Aspartic acid | 3.395 ± 0.142 a | 3.375 ± 0.111 a | 3.030 ± 0.180 a |
| Threonine | 1.962 ± 0.073 a | 2.128 ± 0.229 a | 1.907 ± 0.025 a |
| Serine | 2.208 ± 0.106 ab | 2.425 ± 0.250 a | 1.940 ± 0.041 b |
| Glutamic acid | 3.332 ± 0.370 ab | 3.538 ± 0.311 a | 2.647 ± 0.090 b |
| Proline | 3.343 ± 0.031 ab | 3.747 ± 0.308 a | 3.040 ± 0.064 b |
| Glycine | 2.990 ± 0.062 a | 2.693 ± 0.263 a | 2.617 ± 0.050 a |
| Alanine | 3.568 ± 0.056 a | 3.638 ± 0.213 a | 2.523 ± 0.058 b |
| Valine | 2.388 ± 0.035 a | 2.415 ± 0.212 a | 2.417 ± 0.176 a |
| Isoleucine | 1.538 ± 0.040 ab | 1.790 ± 0.193 a | 1.453 ± 0.063 b |
| Leucine | 2.000 ± 0.050 a | 2.090 ± 0.229 a | 1.820 ± 0.093 a |
| Tyrosine | 1.768 ± 0.097 ab | 1.845 ± 0.100 a | 1.583 ± 0.061 b |
| Phenylalanine | 1.143 ± 0.078 a | 1.075 ± 0.120 a | 1.107 ± 0.041 a |
| Histidine | 1.055 ± 0.034 a | 1.153 ± 0.164 a | 0.873 ± 0.062 a |
| Lysine | 1.635 ± 0.030 a | 1.810 ± 0.068 b | 1.177 ± 0.033 c |
| Arginine | 1.668 ± 0.049 ab | 1.757 ± 0.197 a | 1.330 ± 0.022 b |
| Cysteine | 0.123 ± 0.004 a | 0.248 ± 0.068 b | 0.197 ± 0.037 ab |
| Methionine | 0.445 ± 0.018 a | 0.558 ± 0.080 a | 0.470 ± 0.051 a |
| Essential Amino Acid | 22 °C | 25 °C | 28 °C |
|---|---|---|---|
| Threonine | 59.81 ± 2.22 a | 73.30 ± 7.89 b | 68.71 ± 0.90 c |
| Valine | 50.83 ± 0.74 ab | 58.13 ± 5.11 a | 60.84 ± 4.43 b |
| Isoleucine | 36.21 ± 0.95 a | 47.66 ± 5.15 b | 40.47 ± 1.77 c |
| Leucine | 35.32 ± 0.88 ab | 41.73 ± 4.58 a | 38.01 ± 1.94 b |
| Lysine | 39.71 ± 0.74 a | 49.69 ± 1.87 b | 33.79 ± 0.95 c |
| Methionine | 21.61 ± 0.88 a | 30.61 ± 4.38 b | 26.99 ± 2.93 ab |
| Phenylalanine | 30.62 ± 2.09 a | 32.57 ± 3.64 a | 35.07 ± 1.30 b |
| EAAI (%) | 37 a | 46 b | 41 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopecká, A.; Kouřimská, L.; Škvorová, P.; Kurečka, M.; Kulma, M. Effect of Temperature on the Nutritional Quality and Growth Parameters of Yellow Mealworm (Tenebrio molitor L.): A Preliminary Study. Appl. Sci. 2024, 14, 2610. https://doi.org/10.3390/app14062610
Kopecká A, Kouřimská L, Škvorová P, Kurečka M, Kulma M. Effect of Temperature on the Nutritional Quality and Growth Parameters of Yellow Mealworm (Tenebrio molitor L.): A Preliminary Study. Applied Sciences. 2024; 14(6):2610. https://doi.org/10.3390/app14062610
Chicago/Turabian StyleKopecká, Anežka, Lenka Kouřimská, Petra Škvorová, Michal Kurečka, and Martin Kulma. 2024. "Effect of Temperature on the Nutritional Quality and Growth Parameters of Yellow Mealworm (Tenebrio molitor L.): A Preliminary Study" Applied Sciences 14, no. 6: 2610. https://doi.org/10.3390/app14062610
APA StyleKopecká, A., Kouřimská, L., Škvorová, P., Kurečka, M., & Kulma, M. (2024). Effect of Temperature on the Nutritional Quality and Growth Parameters of Yellow Mealworm (Tenebrio molitor L.): A Preliminary Study. Applied Sciences, 14(6), 2610. https://doi.org/10.3390/app14062610