Abstract
This study aimed to investigate the changes in Paraoxonase (PON), Carbonic Anhydrase (CA), and β-glucosidase levels of different aged rats and scopolamine-induced memory impairment rats. This study used young, adult, and middle-aged male Wistar Albino rats. Scopolamine was administered as a single dose/multiple doses and a Morris water maze (MWM) was used for spatial learning testing in rats. Enzyme-linked immunosorbent assays (ELISAs) were used to determine serum and liver PON and β-glucosidase levels. The CA enzyme activity was assayed following the hydration of CO2. As a result of the comparison of age-related and scopolamine-related changes in PON and β-glucosidase levels in liver and serum samples, no significant age-related and scopolamine-effective changes were observed in serum, while liver PON and liver β-glucosidase levels were found to change significantly. CA activity studies, on the other hand, showed that adults have the lowest CA activity compared to young and middle-aged groups and scopolamine inhibited CA activity in vivo. We found that adult rats modeled with memory impairment had statistically lower levels of liver PON and liver β-glucosidase. CA activity was also found to be significantly reduced. β-glucosidase and CA should be further investigated in terms of neurodegenerative disease risk factors, just like PON, whose importance has been determined by numerous studies in the literature.
1. Introduction
The scopolamine-induced memory impairment model is the most widely used pharmacological model for anti-dementia drug screening. The anti-muscarinic receptor drug scopolamine efficiently causes memory disruption by blocking the cholinergic nerve system [1,2,3]. Models of amnesia have been established using scopolamine; mice or rats can be utilized with dosages ranging from 0.5 mg/kg to 5 mg/kg for single/or multiple doses [4]. Repeated doses of scopolamine have been shown to cause more impairment in memory function in the passive avoidance test than a single dose of scopolamine [5].
The role of oxidative damage in human diseases, such as neurodegenerative disorders (NDs), and changes in the activity of the antioxidant and/or other enzymes in the organisms, has become an increasingly important research field for NDs. Scopolamine is known to increase the production of endogenous reactive oxygen species and pro-inflammatory cytokines leading to severe nervous system damage, especially in the hippocampus and/or prefrontal cortex regions [2,6,7]. In oxidative stress conditions, brain neurotransmitters such acetylcholine (Ach) and monoamine neurotransmitters are likely inactivated. Lipid peroxidation yields malondialdehyde (MDA) and antioxidant enzymes superoxide dismutase (SOD) and catalase (CAT) are thought to be crucial in combating oxidative stress [8,9,10,11,12]. Although many enzymes related to oxidative stress in NDs have been studied, Paraoxonase (PON), Carbonic Anhydrase (CA), and β-glucosidase enzymes are within the scope of our study. The PON enzyme family has a well-established role in the prevention of cardiovascular disease and demonstrates anti-inflammatory and antioxidant capabilities. Additionally, they have been connected to the beginning and development of numerous neurological conditions and neurodegenerative diseases [13,14,15]. β-glucosidases play important roles in a variety of crucial physiological processes, particularly cytosolic β-glucosidase, which demonstrates important activity towards many common dietary xenobiotics including drugs, although the question regarding the impact of scopolamine on liver and serum cytosolic β-glucosidase in mammals has not been answered yet [16,17]. CA dysfunction affects cognition and is linked to mental retardation, Alzheimer’s disease, and aging according to an increasing body of research [18]. Patients with Carbonic anhydrase II (CAII) deficiencies exhibit cognitive impairments that range in severity from mild deficits to severe mental retardation [19,20]. In the hippocampi of Alzheimer’s disease (AD) patients, an increase in oxidized proteins has been also reported, including CAII [21,22].
The fact that approximately 152 million people—three times the present estimate of 50 million—are predicted to suffer dementia by 2050 serves as a stark reminder of the significance of neurodegenerative diseases [23]. Therefore, this study aimed to evaluate CA activity changes from erythrocytes with PON and β-glucosidase level changes in serum and liver tissues of young, adult, and middle-aged rats and scopolamine-induced memory impairment rats.
2. Materials and Methods
2.1. Experimental Animals
In total, 1-month-old (young), 4–5-month-old (adult), and 14–16-month-old (middle-aged) male Wistar Albino rats were used in this study. Rats were housed in groups of five per cage under temperature- (21 ± 3 °C), humidity- (55–60%), and light- (12 h light, 12 h dark cycle, lights on at 08.00) controlled conditions. The animals were handled daily for one week before the experiments. They were taken to the experimental room at least 24 h before the behavioral experiments and adapted to the environment. Standard rodent pellet feed was given to all groups of rats and ad libitum access to food and water was provided. The animal experiment was conducted in accordance with the recommendations made by Balikesir University’s Local Ethics Committee for Animal Research (27 April 2023, number 2023/3–6 and number 2023/3–7). The animal care procedures were conducted according to the guidelines of the European Community Council Directive (86/609/EEC). The Experimental Animal Production, Care, Application and Research Center at Balikesir University provided all the animal supplies and animal care.
2.2. Treatment
Scopolamine hydrobromide was prepared by dissolving it in 0.9% saline and administered intraperitoneally.
2.3. Experimental Groups
1 month-old (young) group (n:10): No application
4–5-month-old (adult) group (n:10): No application
14–16-month-old (middle-aged) group (n:10): No application
Adult (4–5 months old) animals were randomly divided into the following 3 groups:
- Control (Morris water maze (MWM) only) group (n:10): The MWM test was applied for five days, and 1 mL/kg saline was applied 30 min before the probe test on the last day of the test.
- MWM + single-dose scopolamine group (n:10): The MWM test was applied for five days, and 1 mg/kg scopolamine was applied 30 min before the probe test on the last day of the test.
- MWM + multiple-dose scopolamine group (n:10): The MWM test was applied for five days, and 1 mg/kg scopolamine was administered 30 min before the test every day of the test.
2.4. Morris Water Maze Test
A cylindrical tank with a diameter of 150 cm and a height of 50 cm was filled with water to a height of approximately 30 cm. The temperature of the water was adjusted to be 25 ± 1 °C. The tank was hypothetically divided into 4 equal parts and defined as north, south, east, and west. A platform with a diameter of 12 cm was placed 1 cm below the water, near the center of one of the quarter quadrants (the southeast quadrant was used in this study). The acquisition period of the test was applied for 4 consecutive days, with each rat undertaking 3 swims. In the first 4 days, it was determined from which direction and in which order the rat would be released into the tank. In each swimming session, the rats were given 60 s to find the platform. The rats that could not find the platform during this period were placed on the platform by the experimenter and allowed to stay on the platform for 20 s. In each swimming session, the time they stepped onto the platform was recorded.
On the 5th day, the last day of the experiment, the platform was removed for the probe test and the animals were removed as a single session and floated for 60 s. During this period, the time spent in the quarter where the platform was previously was recorded. All the experiments were recorded with a camera and analyzed via the EthoVision XT (Version10, Nol-dus Information Technology, Wageningen, The Netherlands,) video tracking program.
2.5. Statistical Analysis for the MWM Test
For statistical analysis, GraphPad Prism 6.0 software (GraphPad Software, Inc., La Jolla, CA, USA) was utilized. The mean and SEM of the results are given. A two-way analysis of variance (ANOVA) was used to analyze the MWM acquisition test, followed by Tukey post hoc tests. One-way ANOVA and Tukey post hoc tests were used to analyze the results of the probe test. p-values less than 0.05 were considered significant.
2.6. The Enzyme-Linked Immunosorbent Assay (ELISA)
Serum and liver β-Glucosidase (rat β-Glucosidase ELISA kit, E1599Ra, Bioassay Technology Laboratory, Shanghai, China), and serum and liver PON (Rat paraoxonase (PON) ELISA Kit, E1563Ra Bioassay Technology Laboratory, Shanghai, China) levels were measured using commercially available kits and were carried out in accordance with the company’s instructions. ELISA analyses were carried out in Atlas Biotechnologies/Ankara laboratory. Briefly, liver samples were rinsed in ice-cold PBS (pH 7.4) and weighed before homogenization. The homogenates were then centrifuged for 5 min at 5000× g to obtain the supernatant. Serum samples were allowed to clot for 10–20 min at room temperature and centrifuged at 3000 rpm for 20 min. The supernatant without sediment was collected.
2.7. CA Activity Assay
CA activity was performed as described previously [17]. Briefly, dry tubes containing blood samples were centrifuged at 1500 rpm for 15 min. With cold water, the packed red blood cells were hemolyzed and hemolysate was utilized for CA enzyme activity tests. The CO2-hydratase activity of the enzyme was tested at room temperature in a veronal buffer (pH 8.6) using phenol-red as an indicator and saturated carbon dioxide solution as a substrate in a final volume of 4.2 mL. The amount of time it took for the solution to transition from red to yellow was measured in seconds. Using the equation (t0 − tc/tc), where t0 and tc are the timings for the pH changes of the non-enzymatic and enzymatic processes, respectively, the enzyme unit (EU) was calculated.
2.8. Statistical Analysis for ELISA Assay and CA Activity Assay
The SPSS version 22.0 software (IBM SPSS Statistics for Windows, Armonk, NY, USA) was used to perform all statistical analyses. Levene’s test was used to determine the homogeneity of variance before the statistical comparison. The Shapiro–Wilk test, a numerical approach for determining data normality, was used to determine whether the findings of experimental analyses had a normal distribution. To assess for significant differences between the groups, the one-way analysis of variance with Tukey’s post hoc test was used to evaluate all of the normally distributed data. The nonparametric Kruskal Wallis test and Tamhane’s T2 were used to evaluate data that were not normally distributed in order to compare the means and statistical differences across all groups. p < 0.05 results were considered significant. Graph Pad Prism 9 was used for the graphs.
3. Results
3.1. The Effect of Scopolamine Administration on the MWM Test
The escape latency time to find the platform during the acquisition period in all groups decreased from day 1 to day 4 (day effect, two-way ANOVA, F (3.36) = 48.21, p < 0.05), except on day 2 of multiple-dose scopolamine administration (Figure 1A). In the probe trial, a significant difference was found between the control and the scopolamine groups in terms of the time spent in the escape platform quadrant (one-way ANOVA, F(2.27) = 4.287, p < 0.05, Figure 1B) (p < 0.05). There was no significant difference in the groups administered single and multiple doses (Figure 1B). There was no significant difference between the control and scopolamine groups in terms of mean speed and total distance moved during the probe test (one-way ANOVA, F (2.27) = 1.009, p = 0.3779, Figure 1C and F (2.27) = 0.9324, p = 0.4059, Figure 1D).
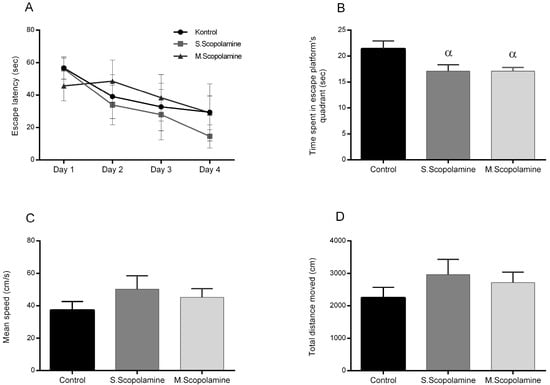
Figure 1.
The effect of scopolamine administration on the MWM test. (A) The results of the acquisition test of MWM. Each value represents the mean ± SEM (n = 10). (B) The results of the probe test of MWM test. Each value represents the mean ± SEM (n = 10). α p < 0.05 compared with the control group. (C) The total distance moved during the probe test of the MWM test. Each value represents the mean ± SEM (n = 10). (D) The mean speed during the probe test of the MWM test. Each value represents the mean ± SEM (n = 10).
3.2. Determination of PON and β-Glucosidase Levels from Serum and Liver
The result of the ELISA assay from rat liver tissues and serum samples and the changes in the PON and β-glucosidase levels for young, adult, and middle-aged rats are presented in Figure 2A,B, respectively. However, serum PON levels did not differ significantly for young, adult, and middle-aged rats, while the liver PON levels of the middle-aged group were found to be statistically significantly higher compared to young (F = 49.913; df = 2.12; p = 0.004) and adult (F = 49.913; df = 2.12; p = 0.005) groups (Figure 2A). Liver β-glucosidase levels of adult rats were found to be lower than those for the young (F = 16.145; df = 2.12; p = 0.0005) and middle-aged groups (F = 16.145; df = 2.12; p = 0.010) but no statistically significant changes were observed for the serum β-glucosidase levels between young, adult and middle-aged rats (Figure 2B).
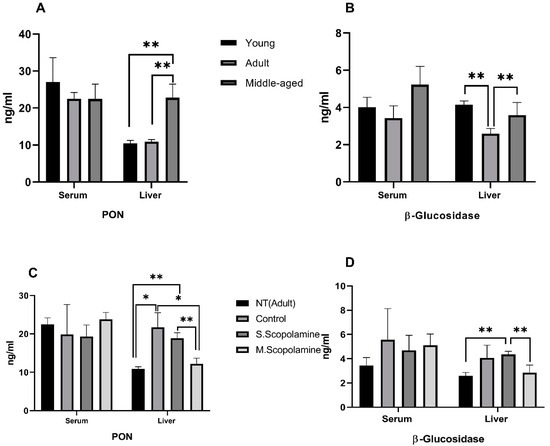
Figure 2.
Changes in enzyme levels. (A) Serum and liver PON levels for young, adult, and middle-aged rats. (B) Serum and liver β-glucosidase levels for young, adult, and middle-aged rats. (C) In vivo effects of single/multiple dose scopolamine administration on the levels of serum and liver PON. (D) In vivo effects of single/multiple dose scopolamine administration on the levels of serum and liver β-glucosidase levels. ELISA assay was performed to determine the PON and β-glucosidase levels. The data represent the means ± SD (n = 5 mice/group) and results are analyzed using one-way ANOVA. * p ≤ 0.05; ** p ≤ 0.01.
The in vivo effects of single dose and multiple doses of scopolamine treatment on serum and liver PON levels are given in Figure 2C. The Morris water maze test increased the liver PON level of the control group compared to the non-treated group (F = 27.934 df = 3.16; p = 0.018). While single-dose scopolamine treatment decreased the level of liver PON compared to the control group, the decrease was not found to be significant. Administration of multiple doses of scopolamine caused a statistically significant decrease in the liver PON level compared to the control (F = 27.934 df = 3.16; p = 0.019). Differences in liver PON levels between single dose and multiple doses of scopolamine-administrated groups were also significant (F = 27.934 df = 3.16; p = 0.001). In general, serum and liver beta glucosidase levels are increased for MWM, single and multiple dose scopolomamine groups compared to non-treated group.But only increase of single scopolamine for liver was found statistically significant (F = 9.378; df = 3.16; p = 0.0005). Multiple doses of scopolamine decreased the liver β-glucosidase significantly compared to the single-dose scopolamine-treated group (F = 9.378; df = 3.16; p = 0.022) (Figure 2D).
3.3. Determination of CA Enzyme Activity from Blood Samples
The results of the CA activity assay from blood samples for young, adult, and middle-aged rats are presented in Figure 3A. The CA activity of adult rats was found to be significantly lower than young (F = 167.081; df = 2.12; p = 0.003) and middle-aged (F = 167.081; df = 2.12; p = 0.0005) groups, but the differences between young and middle-aged rats were not found to be significant. The MWM test increased the CA enzyme activity of the control group rats compared to the non-treated group (F = 13.672; df = 3.26; p = 0.034) The administration of a single dose and multiple doses of scopolamine decreased the CA activity in a dose-dependent manner compared to the control group, as shown in Figure 3B (F = 13.672; df = 3.26; p = 0.027 for single-dose scopolamine control groups, F = 13.672; df = 3.26; p = 0.006 for multiple-dose scopolamine control groups).
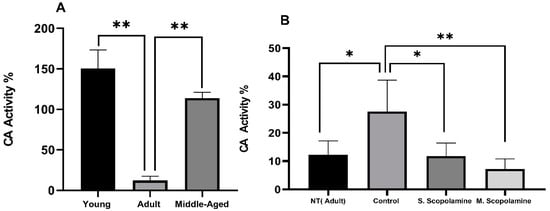
Figure 3.
Changes in the carbonic anhydrase activity. (A) Young, adult, and middle-aged rats. (B) In vivo effects of scopolamine administration and MWM test on the carbonic anhydrase activity for NT, control, S. scopolamine, and M. scopolamine groups. Dry tubes containing blood samples were centrifuged at 1500 rpm for 15 min. With cold water, the packed red blood cells were hemolyzed and hemolysate was utilized for CA activity assay. The data represent means ± SD (n = 5 mice/group) and the results are analyzed using one-way ANOVA. * p ≤ 0.05; ** p ≤ 0.01.
4. Discussion
The complex origin of various neurodegenerative disorders remains unclear. Since distinct cell populations with highly specialized and distinct roles are distributed throughout the central nervous system (CNS), there are several neurodegenerative disorders. Nonetheless, certain risk factors, like exposure to specific chemicals, the existence of specific polymorphisms, modifications in cholesterol metabolism, reduced antioxidant activity and elevated oxidative stress, are shared by various illnesses [24,25]. In this work, we compared for the first time the changes in the PON, β-glucosidase, and CA of young, adult, and middle-aged rats and scopolamine-induced memory impairment rats. Our results revealed that PON and CA have led to more impressive results compared to β-glucosidase as scopolamine administration decreased the liver PON level and CA activity in a dose-dependent manner. Scopolamine-induced memory impairment was assessed by the MWM test with an acquisition test and probe test. Scopolamine, a well-known anticholinergic drug, is frequently used as a standard drug with the experimental purpose of causing cognitive impairment in animals. Scopolamine induces cholinergic neuron loss and ACh reduction, oxidative stress damage, neuroinflammation, and amnesia [2]. Several studies have specifically evaluated the effects of scopolamine on tasks that were developed to study attentional processes. Intraperitoneal injection of scopolamine could cause learning and memory disorders in mice [5,26]. In our study, scopolamine administration was performed as a single dose on the 5th day of the MWM test and multiple doses every day for five days. The MWM is a spatial learning test for rodents. The need to model the assumed mechanism of NDs and, thus detect the therapeutic response of putative cognition enhancers, is necessary for understanding NDs and for treating it pharmacologically [1]. In this test, rodents try to find a hidden escape platform underwater, starting from different starting points in an open swimming tank. The test relies on using distant cues in the environment for this purpose. No significant difference in the learning abilities of rats was found in the single and multiple doses of scopolamine-administered groups.
Over the last few decades, there has been a growing interest in the role of antioxidant enzymes related to NDs. The potential of PONs to provide antioxidant and anti-inflammatory counter-regulatory mechanisms that increase neuroprotection in the brain has made them one of the most prominent enzymes in this field [15]. In our studies, in which PON and β-glucosidase levels were determined in young, adult, and middle-aged rats, no significant change was observed in serum PON and serum β-glucosidase protein levels, while PON protein levels in the liver were significantly increased in middle-aged individuals compared to adult and young individuals. The liver β-glucosidase level was found to show a significant decrease in adults compared to young and middle-aged rats. Studies investigating PON and β-glucosidase protein levels in rats that were administered single dose and multiple doses of scopolamine and subjected to the MWM test revealed that there was no significant change in the serum PON and serum β-glucosidase levels between the groups. However, it was determined that the liver PON level and liver β-glucosidase level increased in the MWM-tested control groups compared to the NT groups, but single-dose and multiple-dose scopolamine administration decreased the liver PON and liver β-glucosidase levels of scopolamine-induced memory impairment rats. An increasing number of studies have shown the association of PON activity with neurodegenerative diseases as multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), AD and Parkinson’s disease. In the study investigating the effect of ozone therapy on rats in which a demyelination model (multiple sclerosis model) was created using ethidium bromide, it was determined that PON1 activity in the brain was significantly reduced compared to the control group [27]. Rajashri et al. (2020) tried to explain the relationship of oleoresin species with memory and cognitive disorders in rats with scopolamine-induced Alzheimer’s disease. Their studies showed that acetylcholinesterase activity was found to be longer in rats given scopolamine compared to the control animals [28]. The enzymatic changes caused by scopolamine in rats will not fully correspond to human NDs, but the results are interesting when the data are compared with studies on this subject. Oxidative stress is linked to increased motor neuron demyelination and has a role in the pathophysiology of MS [29,30,31]. It has been noted that blood PON1 activity and total antioxidant status were low in recurrent-remitting multiple sclerosis (RRMS) patients, although total antioxidant status was high [24,32]. Another multifactorial neuro-degenerative illness with altered mitochondria and dysfunctional brain cells is ALS [33]. Saeed et al. (2006) have shown a significant link between the polymorphisms present in the PON gene and ALS [34]. Dementia is another neurodegenerative disorder characterized by a progressive decline in cognitive function. About 80% of dementia cases identified in elderly populations are related to AD [24]. Cervellati et al. (2015) report that arylesterase and PON-1 levels were lower in individuals with moderate cognitive impairment (MCI), vascular dementia (VAD), and late-onset AD than in healthy controls [35]. Furthermore, Saeidi et al. (2017) evaluated the genotype, allele frequency, and activity of PON-1 arylesterase in late-onset AD patients and healthy controls, concluding that PON-1 activity is considerably lower in AD patients than in controls [36]. By comparing the PON-high-density lipoprotein (HDL) enzyme activity in the AD group to that of healthy people, Paragh et al. (2002) demonstrated a connection between cholesterol levels and the progression of AD and VAD. In comparison to controls, patients with AD and VAD had considerably higher serum cholesterol levels. When compared to healthy controls, PON-1 HDL-associated activity in AD and VAD patients was significantly lower [37]. Our PON results are partially consistent with the literature, as it was determined that scopolamine administration decreases the liver PON level. Since there is limited information about scopolamine application and examination of β-glucosidase levels, the data obtained in this study made a significant contribution to the literature. Determining the serum and liver β-glucosidase levels for adult rat models of neurodegenerative diseases will be useful as an early marker for potential risk. Dimmitt et al. (2003) performed a similar study to test whether serum levels of cytosolic β-glucosidase are elevated in a young rat model of necrotizing enterocolitis and showed the serum concentration of cytosolic β-glucosidase was found to significantly increase in animal models with ischemia and bowel obstruction [38]. Recently, several studies have shed light on CAs as possible new targets for memory impairment and cognitive problems [39]. Our carbonic anhydrase activity studies showed that the CA activities of adult rats are significantly less than young and middle-aged individuals. MWM-tested animals have higher CA activity compared to the non-treated group and scopolamine administration decreased the activity of CA in single-dose and multiple-dose scopolamine-administrated groups compared to the control group rats. In addition to being involved in long-term synaptic conversion and pH regulation, carbonic anhydrases are linked to a number of diseases, including Down syndrome, ALS, mental retardation, and AD. In the study conducted by Liu et al. (2016), CAI expression was shown in human spinal cord cells and it was hypothesized that CAI expression may have an important function in motor neuron degeneration in ALS disease [40]. Golabek et al. (2007) investigated the level of CAII in developing brains with Down syndrome and determined that the level of CAII increased in the brains of Down syndrome model mice and also in humans, especially in infants and young children with Down syndrome [41]. Jang et al. (2010) focused on the determination of CAII protein levels in blood plasma samples using immunoblot and ELISA methods to determine whether the level of carbonic anhydrase increases in Alzheimer’s disease. Accordingly, plasmas from Alzheimer’s disease, amnestic mild cognitive impairment, and cognitively normal control groups were compared. The plasma level of CAII was significantly increased in Alzheimer’s patients compared with the control group [21]. These results provided further evidence that changes in carbonic anhydrase levels may play a role in the pathogenesis of neurodegenerative diseases [21].
5. Conclusions
Our study showed the learning abilities of rats and the changes in PON, β-glucosidase, and CA levels from the points of single- and multiple-dose scopolamine administration. Comparison of the age-dependent and scopolamine-related changes in the PON and β-glucosidase levels indicated that only liver PON and liver β-glucosidase levels were affected but serum PON and serum β-glucosidase levels did not change significantly by age or by scopolamine administration. However, CA activity was affected significantly by both age and scopolamine administration. More attention should be paid to the impact of scopolamine on changes in enzyme levels in relation to neurodegenerative diseases.
Author Contributions
Conceptualization H.Y. and E.A.; methodology B.E.D., F.Ş. and M.Ç.; formal analysis B.E.D., F.Ş. and M.Ç.; software H.Y. and E.A.; writing—original draft preparation H.Y. and E.A.; writing—review and editing H.Y.; supervision H.Y.; project administration H.Y.; funding acquisition H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported partially by the Scientific Research Council of Balikesir University (Project No.2022/070).
Institutional Review Board Statement
The animal experiment was conducted in accordance with the recommendations made by Balikesir University’s Local Ethics Committee for Animal Research (27 April 2023, Number 2023/3-6 and Number 2023/3-7). The Experimental Animal Production, Care, Application and Research Center at Balikesir University provided all the animal supplies and animal care.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Klinkenberg, I.; Blokland, A. The validity of scopolamine as a pharmacological model for cognitive impairment: A review of animal behavioral studies. Neurosci. Biobehav. Rev. 2010, 34, 1307–1350. [Google Scholar] [CrossRef]
- Wang, Z.-X.; Lian, W.-W.; He, J.; He, X.-L.; Wang, Y.-M.; Pan, C.-H.; Li, M.; Zhang, W.-K.; Liu, L.-Q.; Xu, J.-K. Cornuside ameliorates cognitive impairments in scopolamine induced AD mice: Involvement of neurotransmitter and oxidative stress. J. Ethnopharmacol. 2022, 293, 115252. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Patel, C.; Pande, S.; Acharya, S. Potentiation of anti-Alzheimer activity of curcumin by probiotic Lactobacillus rhamnosus UBLR-58 against scopolamineinduced memory impairment in mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1955–1962. [Google Scholar] [CrossRef]
- Rahimzadegan, M.; Soodi, M. Comparison of memory impairment and oxidative stress following single or repeated doses administration of scopolamine in rat hippocampus. Basic Clin. Neurosci. 2018, 9, 5–14. [Google Scholar] [CrossRef]
- Juszczyk, G.; Mikulska, J.; Kasperek, K.; Pietrzak, D.; Mrozek, W.; Herbet, M. Chronic Stress and Oxidative Stress as Common Factors of the Pathogenesis of Depression and Alzheimer’s Disease: The Role of Antioxidants in Prevention and Treatment. Antioxidants 2021, 10, 1439. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Feng, X.; Meng, Q.; Li, Y.; Chen, S.; Wang, G.; Nie, K. Gingerol ameliorates cisplatin-induced pica by regulating the TPH/MAO-A/SERT/5-HT/5-HT3 receptor system in rats. Drug Des. Dev. Ther. 2020, 14, 4085–4099. [Google Scholar] [CrossRef]
- Zarrouk, A.; Hammouda, S.; Ghzaiel, I.; Hammami, S.; Khamlaoui, W.; Ahmed, S.H.; Lizard, G.; Hammami, M. Association between oxidative stress and altered cholesterol metabolism in Alzheimer’s disease patients. Curr. Alzheimer Res. 2020, 17, 823–834. [Google Scholar] [CrossRef]
- Kong, D.; Yan, Y.; He, X.Y.; Yang, H.; Liang, B.; Wang, J.; He, Y.; Ding, Y.; Yu, H. Effects of resveratrol on the mechanisms of antioxidants and estrogen in Alzheimer’s Disease. BioMed Res. Int. 2019, 20, 8983752. [Google Scholar] [CrossRef] [PubMed]
- Ming, Z.; Wotton, C.A.; Appleton, R.T.; Ching, J.C.; Loewen, M.E.; Sawicki, G.; Bekar, L.K. Systemic lipopolysaccharide-mediated alteration of cortical neuromodulation involves increases in monoamine oxidase-A and acetylcholinesterase activity. J. Neuroinflamm. 2015, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.J.; Xu, Q.P.; Shoyama, Y. Extract of Yi Zhi Fang improves learning and memory behaviours of mice and its possible mechanisms. Phytother. Res. 2002, 16, 449–454. [Google Scholar] [CrossRef]
- Youdim, M.B.; Bakhle, Y.S. Monoamine oxidase: Isoforms and inhibitors in Parkinson’s disease and depressive illness. Br. J. Pharmacol. 2006, 147, S287–S296. [Google Scholar] [CrossRef] [PubMed]
- Erlich, P.M.; Lunetta, K.L.; Cupples, L.A.; Abraham, C.R.; Green, R.C.; Baldwin, C.T.; Farrer, L.A. Serum paraoxonase activity is associated with variants in the PON gene cluster and risk of Alzheimer disease. Neurobiol. Aging 2012, 33, 1015.e7–1015.e23. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, H. Proteomic exploration of paraoxonase 1 function in health and disease. Int. J. Mol. Sci. 2023, 24, 7764. [Google Scholar] [CrossRef]
- Khalaf, F.K.; Connolly, J.; Khatib-Shahidi, B.; Albehadili, A.; Tassavvor, I.; Ranabothu, M.; Eid, N.; Dube, P.; Khouri, S.J.; Malhotra, D.; et al. Paraoxonases at the Heart of Neurological Disorders. Int. J. Mol. Sci. 2023, 24, 6881. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, G.; Tehrani, S.S.; Fana, S.E.; Moradi-Sardareh, H.; Panahi, G.; Maniati, M.; Meshkani, R. Crosstalk between Alzheimer’s disease and diabetes: A focus on anti-diabetic drugs. Metab. Brain Dis. 2023, 38, 1769–1800. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, H.; Sunay, F.B.; Sinan, S.; Köçkar, F. In vivo effects of curcumin on the paraoxonase, carbonic anhydrase, glucose-6-phosphate dehydrogenase and β-glucosidase enzyme activities in dextran sulphate sodium-induced ulcerative colitis mice. J. Enzym. Inhib. Med. Chem. 2016, 31, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.K.; Alkon, D.L. Carbonic anhydrase gating of attention: Memory therapy and enhancement. Trends Pharmacol. Sci. 2002, 23, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Sly, W.S.; Hewett-Emmett, D.; Whyte, M.P.; Yu, Y.S.; Tashian, R.E. Carbonic anhydrase II deficiency identified as the primary defect in the autosomal recessive syndrome of osteopetrosis with renal tubular acidosis and cerebral calcification. Proc. Natl. Acad. Sci. USA 1983, 80, 2752–2756. [Google Scholar] [CrossRef]
- Sly, W.S.; Whyte, M.P.; Sundaram, V.; Tashian, R.E.; Hewett-Emmett, D.; Guibaud, P.; Vainsel, M.; Jorge Baluarte, H.; Gruskin, A.; Al-Mosawi, M.; et al. Carbonic anhydrase II deficiency in 12 families with the autosomal recessive syndrome of osteopetrosis with renal tubular acidosis and cerebral calcification. N. Engl. J. Med. 1985, 313, 139–145. [Google Scholar] [CrossRef]
- Jang, B.G.; Yun, S.M.; Ahn, K.; Song, J.H.; Jo, S.A.; Kim, Y.Y.; Kim, D.K.; Park, M.H.; Han, C.; Koh, Y.H. Plasma carbonic anhydrase II protein is elevated in Alzheimer’s disease. J. Alzheimers Dis. 2010, 21, 939–945. [Google Scholar] [CrossRef]
- Sultana, R.; Boyd-Kimball, D.; Poon, H.F.; Cai, J.; Pierce, W.M.; Klein, J.B.; Merchant, M.; Markesbery, W.R.; Butterfield, D.A. Redox proteomics identification of oxidized proteins in Alzheimer’s disease hippocampus and cerebellum: An approach to understand pathological and biochemical alterations in AD. Neurobiol. Aging 2006, 27, 1564–1576. [Google Scholar] [CrossRef]
- Weidner, W.S.; Barbarino, P. P4-443: The State of The Art of Dementia Research: New Frontiers. Alzheimers Dement. 2019, 15, P1473. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Reichert, C.O.; Levy, D.; Bydlowski, S.P. Paraoxonase Role in Human Neurodegenerative Diseases. Antioxidants 2020, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Drummond, E.; Wisniewski, T. Alzheimer’s disease: Experimental models and reality. Acta Neuropathol. 2017, 133, 155–175. [Google Scholar] [CrossRef]
- Salem, N.A.; Assaf, N.; Ismail, M.F.; Khadrawy, Y.A.; Samy, M. Ozone Therapy in Ethidium Bromide-Induced Demyelination in Rats: Possible Protective Effect. Cell. Mol. Neurobiol. 2015, 36, 943–954. [Google Scholar] [CrossRef]
- Rajashri, K.; Mudhol, S.; Serva Peddha, M.; Borse, B.B. Neuroprotective effect of spice oleoresins on memory and cognitive impairment associated with scopolamine-induced Alzheimer’s disease in rats. ACS Omega 2020, 5, 30898–30905. [Google Scholar] [CrossRef]
- Lassmann, H.; Van Horssen, J.; Mahad, D. Progressive multiple sclerosis: Pathology and pathogenesis. Nat. Rev. Neurol. 2012, 8, 647–656. [Google Scholar] [CrossRef]
- Mahad, D.H.; Trapp, B.D.; Lassmann, H. Pathological mechanisms in progressive multiple sclerosis. Lancet Neurol. 2015, 14, 183–193. [Google Scholar] [CrossRef]
- Sayre, L.M.; Perry, G.; Smith, M.A. Oxidative stress and neurotoxicity. Chem. Res. Toxicol. 2008, 21, 172–188. [Google Scholar] [CrossRef]
- Kirbas, A.; Kirbas, S.; Anlar, O.; Efe, H.; Yilmaz, A. Serum paraoxonase and arylesterase activity and oxidative status in patients with multiple sclerosis. J. Clin. Neurosci. 2013, 20, 1106–1109. [Google Scholar] [CrossRef] [PubMed]
- Obrador, E.; Salvador, R.; López-Blanch, R.; Jihad-Jebbar, A.; Vallés, S.L.; Estrela, J.M. Oxidative Stress, Neuroinflammation and Mitochondria in the Pathophysiology of Amyotrophic Lateral Sclerosis. Antioxidants 2020, 9, 901. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.; Siddique, N.; Hung, W.Y.; Usacheva, E.; Liu, E.; Sufit, R.L.; Heller, S.L.; Haines, J.L.; Pericak-Vance, M.; Siddique, T. Paraoxonase cluster polymorphisms are associated with sporadic ALS. Neurology 2006, 67, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Trentini, A.; Romani, A.; Bellini, T.; Bosi, C.; Ortolani, B.; Zurlo, A.; Passaro, A.; Seripa, D.; Zuliani, G. Serum paraoxonase and arylesterase activities of paraoxonase-1 (PON-1), mild cognitive impairment, and 2-year conversion to dementia: A pilot study. J. Neurochem. 2015, 135, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Saeidi, M.; Shakeri, R.; Marjani, A.; Khajeniazi, S. Alzheimer’s disease and paraoxonase 1 (pon1) gene polymorphisms. Open Biochem. J. 2017, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Paragh, G.; Balla, P.; Katona, E.; Seres, I.; Égerházi, A.; Degrell, I. Serum paraoxonase activity changes in patients with Alzheimer’s disease and vascular dementia. Eur. Arch. Psychiatry Clin. Neurosci. 2002, 252, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Dimmitt, R.A.; Glew, R.; Colby, C.; Brindle, M.; Skarsgard, E.; Moss, R.L. Serum cytosolic β-glucosidase activity in a rat model of necrotizing enterocolitis. Pediatr. Res. 2003, 54, 462–465. [Google Scholar] [CrossRef]
- Provensi, G.; Carta, F.; Nocentini, A.; Supuran, C.T.; Casamenti, F.; Passani, M.B.; Fossati, S. A new kid on the block? Carbonic anhydrases as possible new targets in Alzheimer’s disease. Int. J. Mol. Sci. 2019, 20, 4724. [Google Scholar] [CrossRef]
- Liu, X.; Lu, D.; Bowser, R.; Liu, J. Expression of Carbonic Anhydrase I in Motor Neurons and Alterations in ALS. Int. J. Mol. Sci. 2016, 17, 1820. [Google Scholar] [CrossRef]
- Palminiello, S.; Kida, E.; Kaur, K.; Walus, M.; Wisniewski, K.E.; Wierzba-Bobrowicz, T.; Rabe, A.; Albertini, G.; Golabek, A.A. Increased levels of carbonic anhydrase II in the developing Down syndrome brain. Brain Res. 2008, 1190, 193–205. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).