
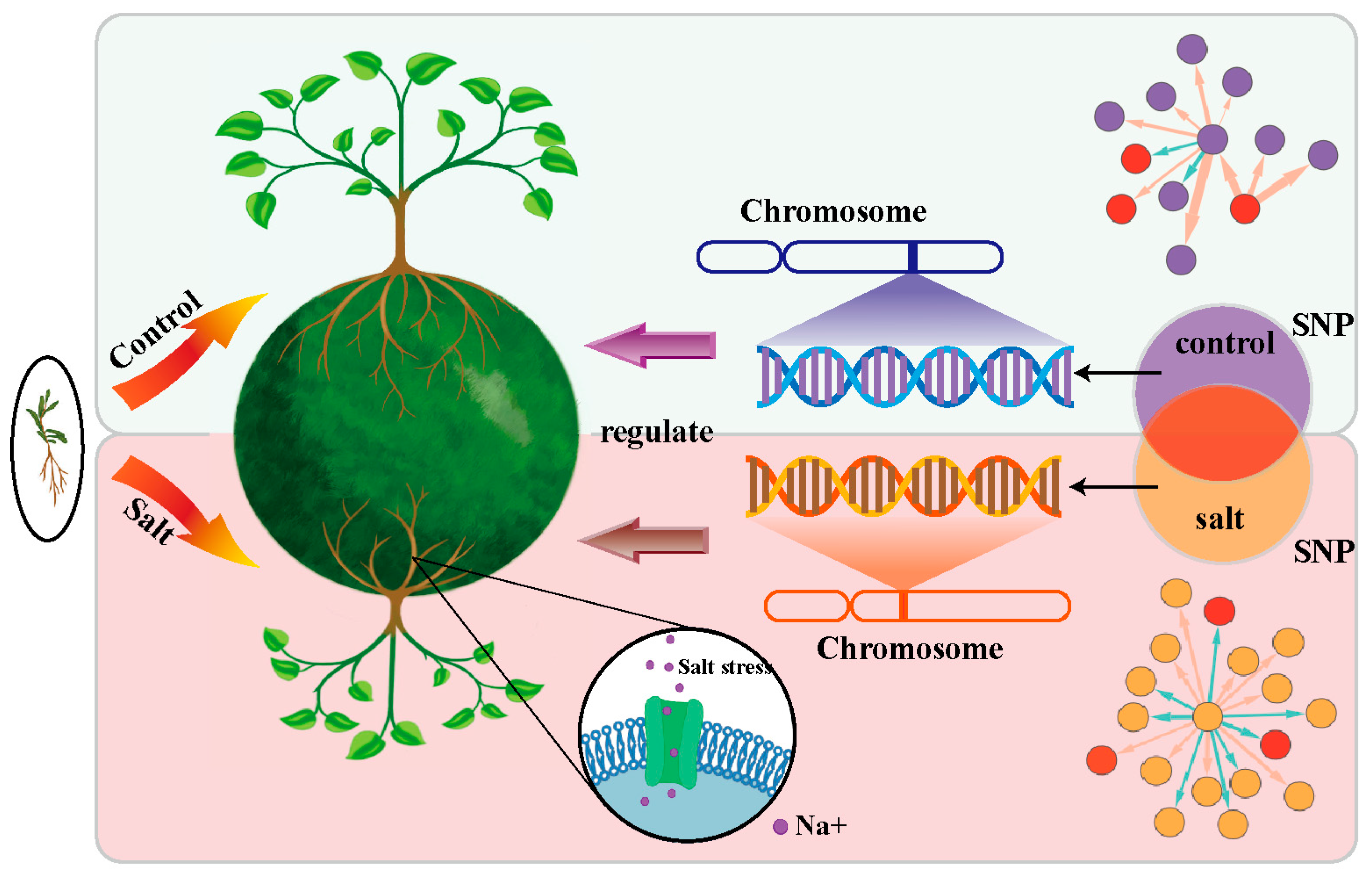
The Genetic Architecture of the Root System during Seedling Emergence in Populus euphratica under Salt Stress and Control Environments

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Environment-Induced Differential Interaction Equation (EDIE)
2.3. Modeling Framework of Systems Mapping
2.4. Inferring QTL Networks
3. Results
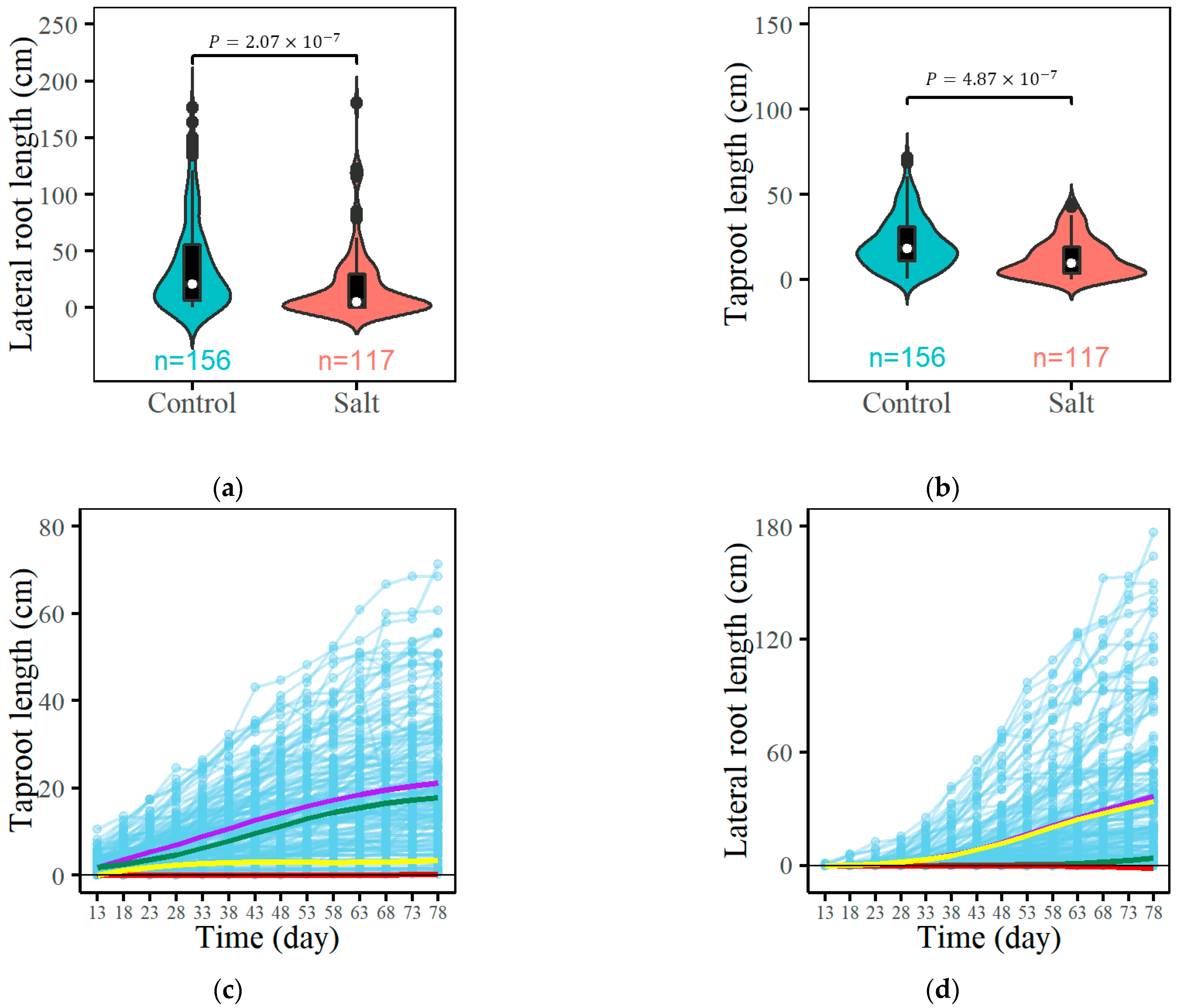
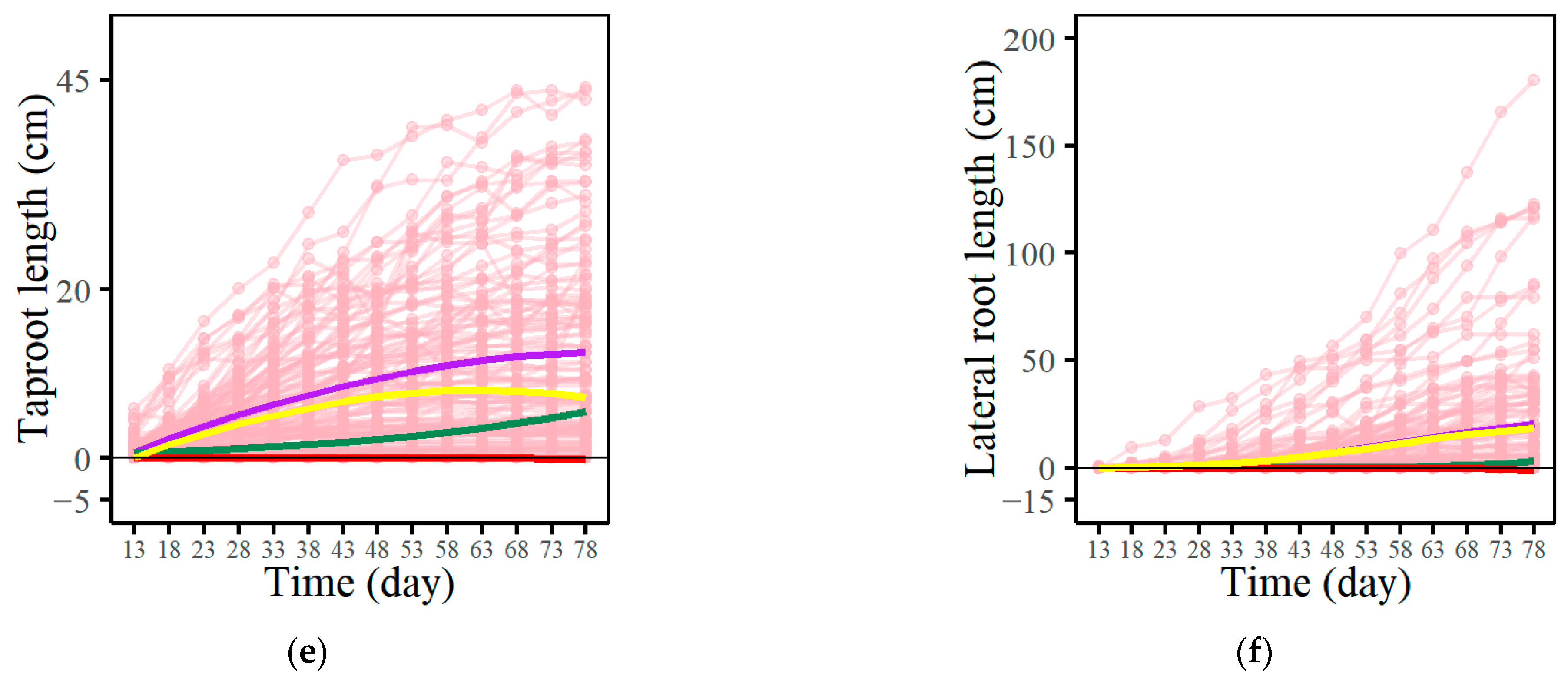
3.1. Growth Trajectory Fitting Based on EDIE
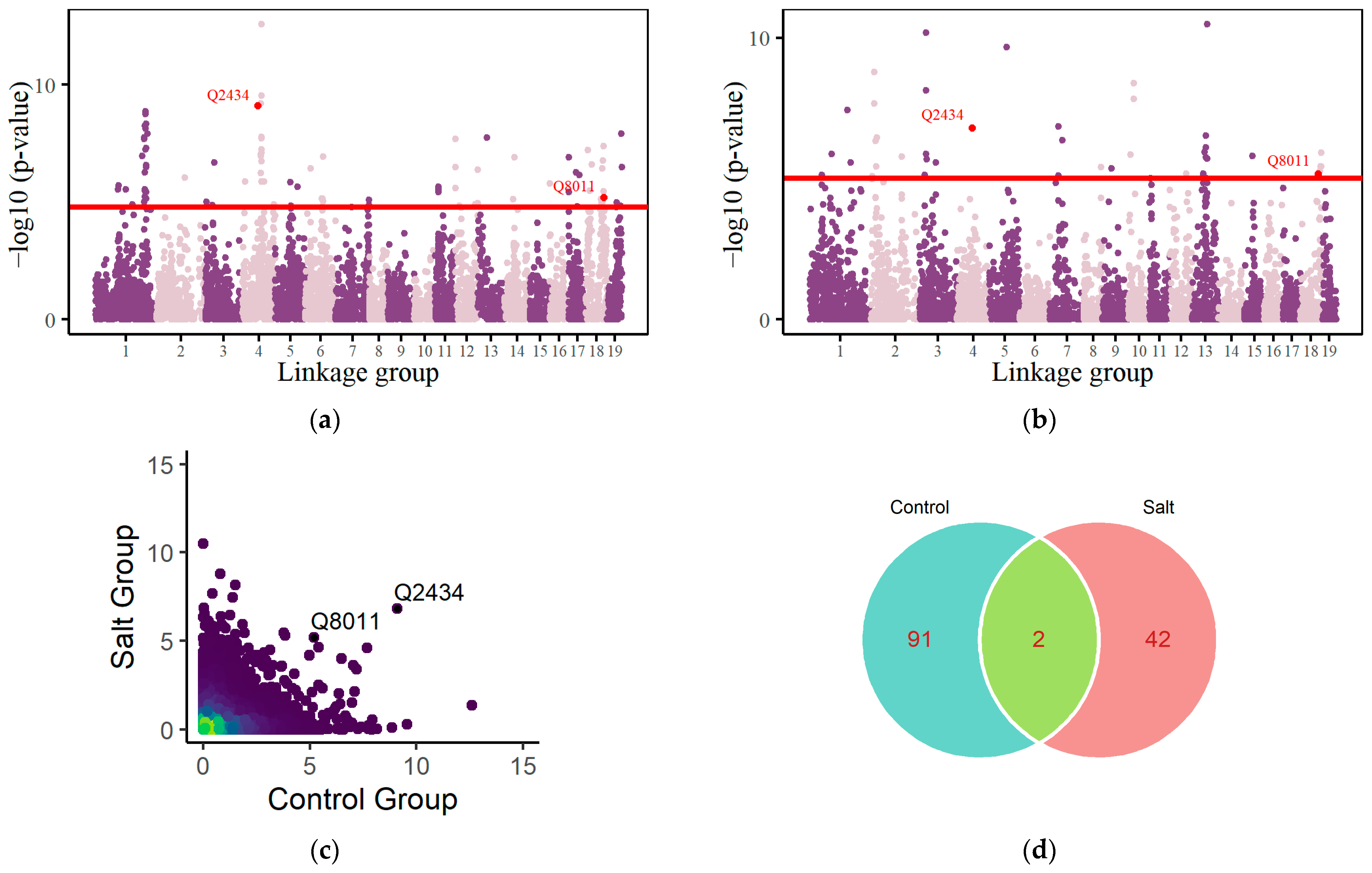
3.2. QTL Mapping of the Root System
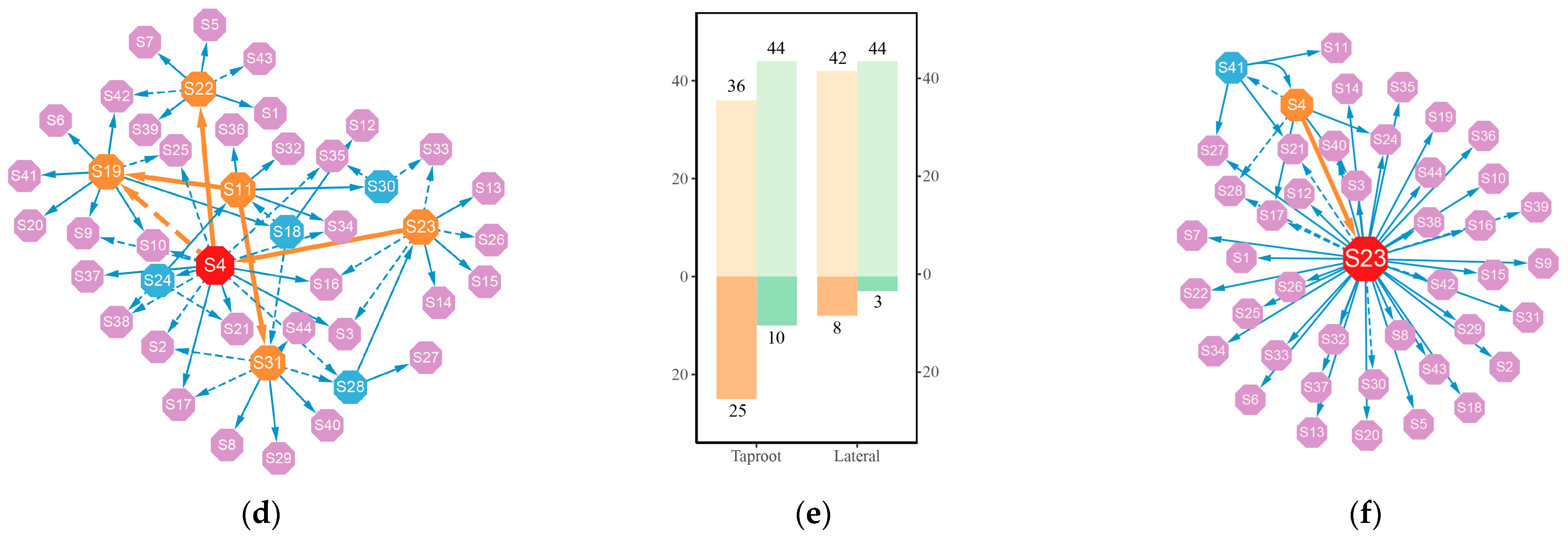
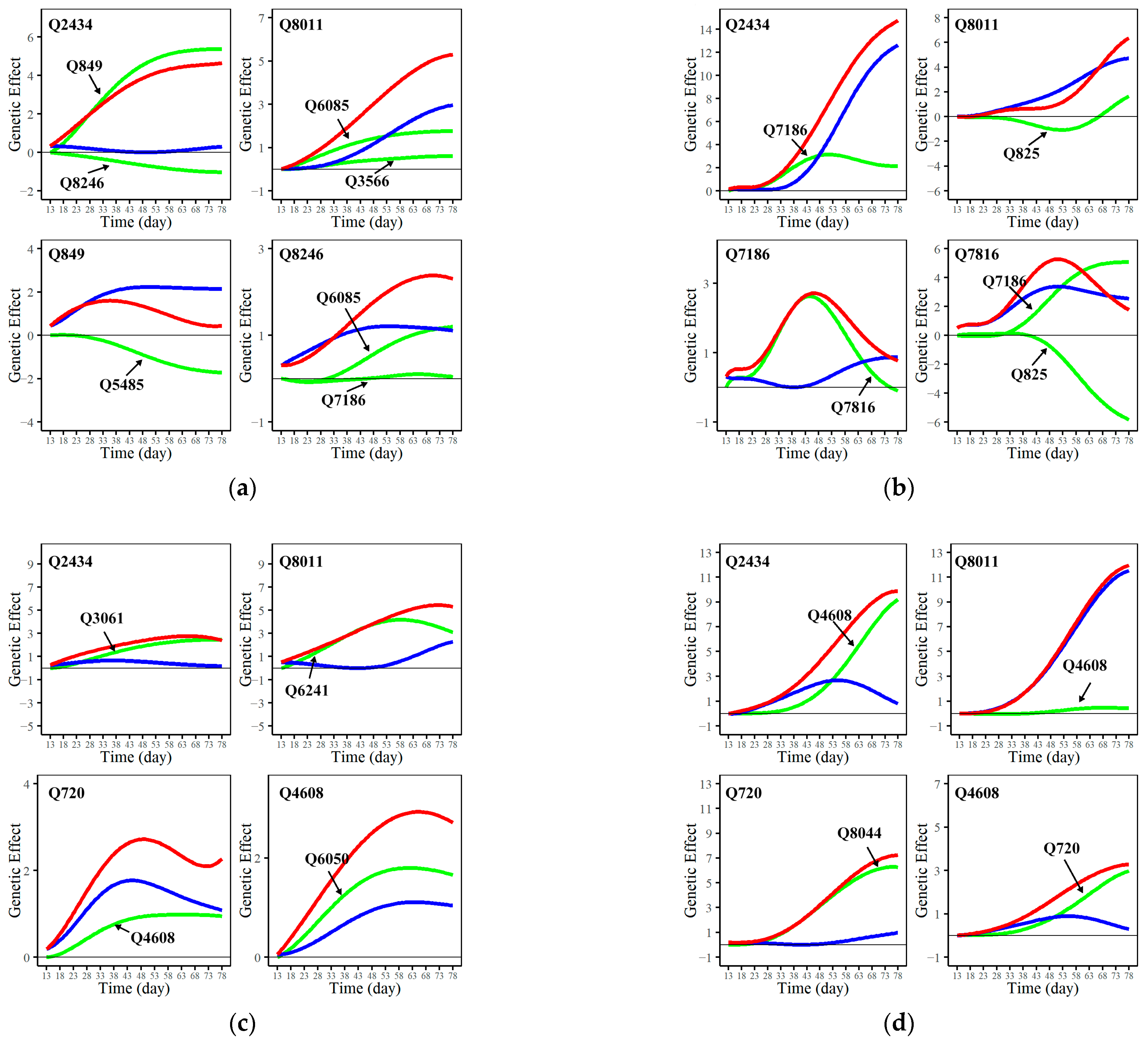
3.3. Genetic Network of Significant QTLs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wyatt, J. Grain and Plant Morphology of Cereals and How Characters Can Be Used to Identify Varieties. In Encyclopedia of Food Grains; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Bloom, A.J.; Chapin III, F.S.; Mooney, H.A. Resource Limitation in Plants—An Economic Analogy. Annu. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Eziz, A.; Yan, Z.; Tian, D.; Han, W.; Tang, Z.; Fang, J. Drought Effect on Plant Biomass Allocation: A Meta-analysis. Ecol. Evol. 2017, 7, 11002–11010. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, R.L.; Reynolds, J.F.; Strain, B.R. Allometric Relations and Growth in Pinus Taeda: The Effect of Elevated CO2, and Changing N Availability. New Phytol. 1996, 134, 85–93. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass Allocation to Leaves, Stems and Roots: Meta-analyses of Interspecific Variation and Environmental Control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Puglielli, G.; Laanisto, L.; Poorter, H.; Niinemets, Ü. Global Patterns of Biomass Allocation in Woody Species with Different Tolerances of Shade and Drought: Evidence for Multiple Strategies. New Phytol. 2021, 229, 308–322. [Google Scholar] [CrossRef]
- Freschet, G.T.; Violle, C.; Bourget, M.Y.; Scherer-Lorenzen, M.; Fort, F. Allocation, Morphology, Physiology, Architecture: The Multiple Facets of Plant Above- and Below-ground Responses to Resource Stress. New Phytol. 2018, 219, 1338–1352. [Google Scholar] [CrossRef]
- Ma, T.; Zeng, W.; Li, Q.; Yang, X.; Wu, J.; Huang, J. Shoot and Root Biomass Allocation of Sunflower Varying with Soil Salinity and Nitrogen Applications. Agron. J. 2017, 109, 2545–2555. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops–What Is the Cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef]
- Shahzad, B.; Rehman, A.; Tanveer, M.; Wang, L.; Park, S.K.; Ali, A. Salt Stress in Brassica: Effects, Tolerance Mechanisms, and Management. J. Plant Growth Regul. 2022, 41, 781–795. [Google Scholar] [CrossRef]
- Ismail, A.M.; Horie, T. Genomics, Physiology, and Molecular Breeding Approaches for Improving Salt Tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef]
- Morton, M.J.; Awlia, M.; Al-Tamimi, N.; Saade, S.; Pailles, Y.; Negrão, S.; Tester, M. Salt Stress under the Scalpel–Dissecting the Genetics of Salt Tolerance. Plant J. 2019, 97, 148–163. [Google Scholar] [CrossRef]
- Galvan-Ampudia, C.S.; Testerink, C. Salt Stress Signals Shape the Plant Root. Curr. Opin. Plant Biol. 2011, 14, 296–302. [Google Scholar] [CrossRef]
- Luo, D.; Shi, Y.-J.; Song, F.-H.; Li, J.-C. Effects of Salt Stress on Growth, Photosynthetic and Fluorescence Characteristics, and Root Architecture of Corylus Heterophylla× C. avellan Seedlings. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2019, 30, 3376–3384. [Google Scholar] [CrossRef]
- Lovelli, S.; Scopa, A.; Perniola, M.; Di Tommaso, T.; Sofo, A. Abscisic Acid Root and Leaf Concentration in Relation to Biomass Partitioning in Salinized Tomato Plants. J. Plant Physiol. 2012, 169, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Julkowska, M.M.; Hoefsloot, H.C.; Mol, S.; Feron, R.; de Boer, G.-J.; Haring, M.A.; Testerink, C. Capturing Arabidopsis Root Architecture Dynamics with ROOT-FIT Reveals Diversity in Responses to Salinity. Plant Physiol. 2014, 166, 1387–1402. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, Y.; Hoch, G.; Wang, Z.; Gu, J. Linkage of Root Morphology to Anatomy with Increasing Nitrogen Availability in Six Temperate Tree Species. Plant Soil 2018, 425, 189–200. [Google Scholar] [CrossRef]
- Avolio, M.L.; Hoffman, A.M.; Smith, M.D. Linking Gene Regulation, Physiology, and Plant Biomass Allocation in Andropogon Gerardii in Response to Drought. Plant Ecol. 2018, 219, 1–15. [Google Scholar] [CrossRef]
- Liu, R.; Yang, X.; Gao, R.; Hou, X.; Huo, L.; Huang, Z.; Cornelissen, J.H. Allometry Rather than Abiotic Drivers Explains Biomass Allocation among Leaves, Stems and Roots of Artemisia across a Large Environmental Gradient in China. J. Ecol. 2021, 109, 1026–1040. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, Q.; Gao, X.; Ma, Y.; Liang, K.; Yue, H.; Huang, X.; Wu, K.; Wang, X. Land Degradation Changes the Role of Above-and Belowground Competition in Regulating Plant Biomass Allocation in an Alpine Meadow. Front. Plant Sci. 2022, 13, 822594. [Google Scholar] [CrossRef]
- Ribeiro, P.R.; Zanotti, R.F.; Deflers, C.; Fernandez, L.G.; de Castro, R.D.; Ligterink, W.; Hilhorst, H.W. Effect of Temperature on Biomass Allocation in Seedlings of Two Contrasting Genotypes of the Oilseed Crop Ricinus Communis. J. Plant Physiol. 2015, 185, 31–39. [Google Scholar] [CrossRef] [PubMed]
- OlaOlorun, B.M.; Shimelis, H.A.; Mathew, I. Variability and Selection among Mutant Families of Wheat for Biomass Allocation, Yield and Yield-related Traits under Drought-stressed and Non-stressed Conditions. J. Agron. Crop Sci. 2021, 207, 404–421. [Google Scholar] [CrossRef]
- Shamuyarira, K.W.; Shimelis, H.; Mathew, I.; Zengeni, R.; Chaplot, V. A Meta-analysis of Combining Ability Effects in Wheat for Agronomic Traits and Drought Adaptation: Implications for Optimizing Biomass Allocation. Crop Sci. 2022, 62, 139–156. [Google Scholar] [CrossRef]
- López-Pérez, M.; Aguirre-Garrido, F.; Herrera-Zúñiga, L.; Fernández, F.J. Gene as a Dynamical Notion: An Extensive and Integrative Vision. Redefining the Gene Concept, from Traditional to Genic-Interaction, as a New Dynamical Version. Biosystems 2023, 234, 105060. [Google Scholar] [CrossRef]
- Fu, L.; Sun, L.; Hao, H.; Jiang, L.; Zhu, S.; Ye, M.; Tang, S.; Huang, M.; Wu, R. How Trees Allocate Carbon for Optimal Growth: Insight from a Game-Theoretic Model. Brief. Bioinform. 2018, 19, 593–602. [Google Scholar] [CrossRef]
- Sun, L.; Wu, R. Mapping Complex Traits as a Dynamic System. Phys. Life Rev. 2015, 13, 155–185. [Google Scholar] [CrossRef]
- Wu, R.; Lin, M. Functional Mapping—How to Map and Study the Genetic Architecture of Dynamic Complex Traits. Nat. Rev. Genet. 2006, 7, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Brauer, F.; Castillo-Chavez, C.; Castillo-Chavez, C. Mathematical Models in Population Biology and Epidemiology; Springer: Berlin/Heidelberg, Germany, 2012; Volume 2. [Google Scholar] [CrossRef]
- Freedman, H.I. Deterministic Mathematical Models in Population Ecology; M. Dekker: New York, NY, USA, 1980; Volume 38, pp. 876–877. [Google Scholar] [CrossRef]
- Hoppensteadt, F. Predator-Prey Model. Scholarpedia 2006, 1, 1563. [Google Scholar] [CrossRef]
- Si, J.; Zhou, T.; Bo, W.; Xu, F.; Wu, R. Genome-Wide Analysis of Salt-Responsive and Novel microRNAs in Populus euphratica by Deep Sequencing. In BMC Genetics; BioMed Central: London, UK, 2014; Volume 15, pp. 1–11. [Google Scholar] [CrossRef]
- Yao, J.; Shen, Z.; Zhang, Y.; Wu, X.; Wang, J.; Sa, G.; Zhang, Y.; Zhang, H.; Deng, C.; Liu, J. Populus euphratica WRKY1 Binds the Promoter of H+-ATPase Gene to Enhance Gene Expression and Salt Tolerance. J. Exp. Bot. 2020, 71, 1527–1539. [Google Scholar] [CrossRef]
- Ye, Z.; Wang, J.; Wang, W.; Zhang, T.; Li, J. Effects of Root Phenotypic Changes on the Deep Rooting of Populus euphratica Seedlings under Drought Stresses. PeerJ 2019, 7, e6513. [Google Scholar] [CrossRef]
- Zhang, M.M. QTL Mapping and Epistatic Analysis of the Response of Populus euphratica Root Growth Dynamics to Salt Stress. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2019. [Google Scholar] [CrossRef]
- Ma, T.; Wang, J.; Zhou, G.; Yue, Z.; Hu, Q.; Chen, Y.; Liu, B.; Qiu, Q.; Wang, Z.; Zhang, J.; et al. Genomic Insights into Salt Adaptation in a Desert Poplar. Nat. Commun. 2013, 4, 2797. [Google Scholar] [CrossRef]
- Wu, R.; Cao, J.; Huang, Z.; Wang, Z.; Gai, J.; Vallejos, E. Systems Mapping: How to Improve the Genetic Mapping of Complex Traits through Design Principles of Biological Systems. BMC Syst. Biol. 2011, 5, 84. [Google Scholar] [CrossRef]
- Jaffrézic, F.; Thompson, R.; Hill, W.G. Structured Antedependence Models for Genetic Analysis of Repeated Measures on Multiple Quantitative Traits. Genet. Res. 2003, 82, 55–65. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, Y.Q.; Casella, G.; Cheverud, J.M.; Wu, R. A Non-Stationary Model for Functional Mapping of Complex Traits. Bioinformatics 2005, 21, 2469–2477. [Google Scholar] [CrossRef]
- Blum, E.K. A Modification of the Runge-Kutta Fourth-Order Method. Math. Comput. 1962, 16, 176–187. [Google Scholar] [CrossRef]
- Do, C.B.; Batzoglou, S. What Is the Expectation Maximization Algorithm? Nat. Biotechnol. 2008, 26, 897–899. [Google Scholar] [CrossRef]
- Moon, T.K. The Expectation-Maximization Algorithm. IEEE Signal Process. Mag. 1996, 13, 47–60. [Google Scholar] [CrossRef]
- Wu, R.; Jiang, L. Recovering Dynamic Networks in Big Static Datasets. Phys. Rep. 2021, 912, 1–57. [Google Scholar] [CrossRef]
- Busiello, D.M.; Suweis, S.; Hidalgo, J.; Maritan, A. Explorability and the Origin of Network Sparsity in Living Systems. Sci. Rep. 2017, 7, 12323. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Lin, Y. Model Selection and Estimation in Regression with Grouped Variables. J. R. Stat. Soc. Ser. B Stat. Methodol. 2006, 68, 49–67. [Google Scholar] [CrossRef]
- Dinneny, J.R. Developmental Responses to Water and Salinity in Root Systems. Annu. Rev. Cell Dev. Biol. 2019, 35, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Korver, R.A.; van den Berg, T.; Meyer, A.J.; Galvan-Ampudia, C.S.; Ten Tusscher, K.H.; Testerink, C. Halotropism Requires Phospholipase Dζ1-mediated Modulation of Cellular Polarity of Auxin Transport Carriers. Plant Cell Environ. 2020, 43, 143–158. [Google Scholar] [CrossRef]
- Warton, D.I.; Duursma, R.A.; Falster, D.S.; Taskinen, S. Smatr 3–an R Package for Estimation and Inference about Allometric Lines. Methods Ecol. Evol. 2012, 3, 257–259. [Google Scholar] [CrossRef]
- Zhang, X.; Gonzalez-Carranza, Z.H.; Zhang, S.; Miao, Y.; Liu, C.-J.; Roberts, J.A. F-Box Proteins in Plants. Annu Plant Rev 2019, 2, 307–328. [Google Scholar] [CrossRef]
- ul Hassan, M.N.; Zainal, Z.; Ismail, I. Plant Kelch Containing F-Box Proteins: Structure, Evolution and Functions. RSC Adv. 2015, 5, 42808–42814. [Google Scholar] [CrossRef]
- Mathieu, N.A.; Levin, R.H.; Spratt, D.E. Exploring the Roles of HERC2 and the NEDD4L HECT E3 Ubiquitin Ligase Subfamily in P53 Signaling and the DNA Damage Response. Front. Oncol. 2021, 11, 659049. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, M.; Liu, S.; Teng, Q.; Li, S.; Jiang, Y. Functions of PPR Proteins in Plant Growth and Development. Int. J. Mol. Sci. 2021, 22, 11274. [Google Scholar] [CrossRef] [PubMed]
- Comas, L.H.; Eissenstat, D.M. Patterns in Root Trait Variation among 25 Co-existing North American Forest Species. New Phytol. 2009, 182, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A. Root Decisions. Plant Cell Environ. 2009, 32, 628–640. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J. Root Architecture and Plant Productivity. Plant Physiol. 1995, 109, 7. [Google Scholar] [CrossRef] [PubMed]
- Bengough, A.G.; McKenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root Elongation, Water Stress, and Mechanical Impedance: A Review of Limiting Stresses and Beneficial Root Tip Traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef]
- Atkinson, J.A.; Wingen, L.U.; Griffiths, M.; Pound, M.P.; Gaju, O.; Foulkes, M.J.; Le Gouis, J.; Griffiths, S.; Bennett, M.J.; King, J. Phenotyping Pipeline Reveals Major Seedling Root Growth QTL in Hexaploid Wheat. J. Exp. Bot. 2015, 66, 2283–2292. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Mu, P.; Zhang, H.; Chen, C.Y.; Gao, Y.; Tian, Y.; Wen, F.; Li, Z. Mapping QTLs of Root Morphological Traits at Different Growth Stages in Rice. Genetica 2008, 133, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Pierret, A.; Moran, C.J.; Doussan, C. Conventional Detection Methodology Is Limiting Our Ability to Understand the Roles and Functions of Fine Roots. New Phytol. 2005, 166, 967–980. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taproot–Lateral-Root | |||
|---|---|---|---|
| Control | Saline Stress | ||
| Lateral Root | Taproot | Lateral Root | Taproot |
| 31.3696 | 19.0193 | 33.4926 | 22.4966 |
| () | () | () | () |
| 0.0815 | 0.0743 | 0.0923 | 0.0368 |
| () | () | () | () |
| −0.0007 | 0.0004 | −0.0028 | −0.0007 |
| () | () | () | () |
| 0.0215 | 2.3801 | 0.0221 | 0.8944 |
| () | () | () | () |
| 0.0569 | −0.1686 | 0.0333 | −0.0742 |
| () | () | () | () |
| 0.8597 | 0.5530 | ||
| 0.9997 | 0.9994 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Z.; Gong, H.; Lu, K.; Zhang, X. The Genetic Architecture of the Root System during Seedling Emergence in Populus euphratica under Salt Stress and Control Environments. Appl. Sci. 2024, 14, 2225. https://doi.org/10.3390/app14062225
Liang Z, Gong H, Lu K, Zhang X. The Genetic Architecture of the Root System during Seedling Emergence in Populus euphratica under Salt Stress and Control Environments. Applied Sciences. 2024; 14(6):2225. https://doi.org/10.3390/app14062225
Chicago/Turabian StyleLiang, Zhou, Huiying Gong, Kaiyan Lu, and Xiaoyu Zhang. 2024. "The Genetic Architecture of the Root System during Seedling Emergence in Populus euphratica under Salt Stress and Control Environments" Applied Sciences 14, no. 6: 2225. https://doi.org/10.3390/app14062225
APA StyleLiang, Z., Gong, H., Lu, K., & Zhang, X. (2024). The Genetic Architecture of the Root System during Seedling Emergence in Populus euphratica under Salt Stress and Control Environments. Applied Sciences, 14(6), 2225. https://doi.org/10.3390/app14062225