Abstract
The objective of this study was to evaluate the antimicrobial effect of direct cold atmospheric plasma (CAPP) treatment for pre-harvest application using four different crop species: Hordeum vulgare L. (barley), Triticum aestivum L. (wheat), Brassica napus L. (rapeseed) and Lupinus angustifolius L. (lupine). The model bacterium Bacillus atrophaeus served as a proxy for spore-forming plant pathogens on the seed surface. After semi-dry inoculation of spores onto the seeds, treatment with two different plasma sources, a volume-dielectric barrier discharge and a corona discharge, and different exposure times was carried out. Subsequently, recovery of viable spores from the seeds’ surfaces was performed. Moreover, seed viability was determined based on maximum germination, as well as water contact angle as a measure for seed surface hydrophilicity. Direct CAPP treatment was efficient in reducing viable spores of B. atrophaeus with no significant differences between the plasma sources, reaching a mean inactivation of 1 log10 CFU/mL across all treatment times and crops species. Maximum germination of seeds was not negatively affected under any treatment condition. Seed hydrophilicity was increased for both plasma sources tested. Overall, this study provides valuable information on the efficiency of direct CAPP treatment of seeds with the purpose of seed hygienization with the premise of unaltered seed vitality and evaluates the potential application in comparison with previous investigated CAPP methods.
1. Introduction
Plant diseases caused by microbial pathogens still pose a considerable threat to crop growth and yield and ultimately food security. At the world scale, yield losses due to fungal pathogens of the most commonly planted staple food crops (wheat, rice, maize, potatoes and soybean) accounted for 44.5 Mt in 2018 [1,2]. In general, potential crop losses without crop protection for the above mentioned five crops are estimated to be in the range of 20 to 30% [2,3]. Moreover, documented worldwide yield loss in cereals caused by microbial pathogens despite using crop protection accounted for ~10% in wheat and ~9% in barley for the time period 1996–1998 [4].
One possible means of crop protection is the application of seed treatment using chemical, biological or physical methods before sowing. Among them, chemical seed dressing using, e.g., fungicides provides almost complete and long-lasting protection against losses caused by seed-borne microbial pathogens. On the other hand, commercial application of those chemicals is decreasing because of toxicological effects on non-target organisms, the emergence of antifungal-resistant strains of especially pathogenic fungi, and environmental aspects along with potential effects on human health and public acceptance of usage (e.g., endocrine-disrupting features of azoles) (e.g., [5,6,7]). Thus, roughly 70% of the registered active ingredients of fungicide products are estimated to be prohibited in the EU because they meet cutoff criteria or might be listed as candidates for substitution in the near future [8]. A recent example is the ban of application from 2018 onwards for the seed fungicide Thiram, a dithiocarbamate fungicide which was widely used in rapeseed and legume seed treatment to prevent seed- as well as soil-borne infections.
Alternatives for chemical seed treatment are needed to secure crop productivity and yield under high-risk situations (e.g., conducive soil properties and environmental conditions, as well as known load with soil-borne pathogens), especially in view of new emerging pathogens associated with climate change (e.g., [9,10,11]).
One possible alternative is represented by biological seed treatments using antagonists or biological extracts. A recent meta-analysis across 210 publications on the effect of biological seed treatment on disease control until 2020 highlighted that the efficiency largely depends on the scale of observation and was significantly lower under field conditions compared to controlled environments with an average of ~50% efficiency [12]. Moreover, the usage of small RNAs and short peptides in disease control is being explored, with major challenges in potential implementation including production costs, safety, stability and mode of delivery, as well as legal requirements [13].
Besides the above-mentioned methods, there is physical treatment, ranging from hot water or steam treatment of seeds (e.g., [14,15,16]) to methods like electron beam treatment (E-beam), which is already on the market [17], or cold atmospheric pressure plasma (CAPP), which is currently in the proof-of-concept stage. For post-harvest applications, CAPP has already been investigated in up-scaled environments under industrial-orientated circumstances (e.g., [18,19,20]).
Plasma is considered the fourth state of matter and can be classified into thermal and non-thermal plasmas. Under laboratory conditions, CAPP can be produced at room temperature and atmospheric pressure by the excitation and ionization of gases with an externally applied energy source, e.g., electric and/or magnetic fields. Hereby, gas molecules are ionized or dissociated after electron and ion collisions with the background gas to create a plasma. Thus, plasma comprises different quantities of chemical species, charged particles, radicals, heat and UV radiation depending on the process parameters and the gas used (e.g., [21,22,23]).
Exposure of the biological targets to CAPP can take place via two different methods. Firstly, direct exposure when the sample is in direct contact with the plasma discharge. In this case, all plasma-generated components can interact with the sample.
Secondly, indirect exposure, which involves treatment of an agent (gaseous or liquid) with CAPP and introducing this agent to the biological target, i.e., treatment of air with CAPP and transferring this plasma-processed air (PPA) to the biological target. Using the indirect configuration, not all CAPP components are able to interact with the target, e.g., UV light and short-lived reactive species are excluded (e.g., [24]).
The efficiency of the microbial inactivation of CAPP strongly depends on the mode of exposure (direct or indirect), the configuration of the plasma reactor, exposure time to CAPP, process parameter used for CAPP treatment (e.g., input voltage, frequency, mode of operation, i.e., continuous, burst or pulsed), post-treatment storage time and the working gas used. Moreover, the type of microorganism (virus, bacteria, fungi, vegetative or generative status), the type of carrier of the microorganism (liquid media, agar plate, crop seed) and the location of the microorganism on the carrier, especially when using seeds (on the seed coat, inside the seed), are important factors which determine the overall efficiency of inactivation and ultimately disease reduction during growth of the crop under natural surroundings (e.g., [25,26]). High efficiencies for inactivation of fungal pathogens using dielectric barrier discharges (DBD) were demonstrated in several publications, e.g., 80% reduction of viable spores of Fusarium on rice seeds using an air DBD [27], 3 to 4 log10 units reduction for Aspergillus, Alternaria and Fusarium on wheat seeds using a coplanar surface barrier discharge [28] and complete inactivation of Fusarium, Trichothecium and Aspergillus on maize seeds using an air DBD [29]. Even though complete inactivation was reached, process parameters were detrimental for seed vitality in some cases, reflected in significant decrease in seed germination for longer treatment times in both above-mentioned cases. Efficiency of corona discharge (CD) treatments displayed similar results, with significant reduced seed germination at longer treatment times. Reduction of natural loads of molds and yeasts after CD treatment of broccoli seeds reached ~1 log10 unit and 1.5 log10 units at 2 and 3 min treatment time, respectively, with a decrease in seed germination at 3 min treatment time of 32% [30]. Moreover, reduction of the natural fungal load (molds and yeasts) on seeds of rapeseed reached 2 log10 units at 3 min treatment along with a decrease in seed germination by ~30% at 3 min treatment time [31].
In our study, we investigate the inactivation efficiency of direct CAPP treatment using DBD [32] and CD [33] plasma sources and Bacillus atrophaeus spores in order to evaluate the potential of direct CAPP exposure as seed treatment against sporulated forms of microbial pathogens, e.g., fungal or bacterial spores, in potential future pre-harvest application. Because unimpaired seed vitality after CAPP treatment has to be ensured in pre-harvest application, maximum germination was monitored simultaneously, along with the status of seed coat hydrophilicity. Bacillus atrophaeus spores were chosen as test organisms for spore-forming pathogens on the seeds surface. This microorganism served as a proxy and is not a plant pathogen. The spores of this Gram-positive bacterium are commonly used as a bio-indicator for evaluating the efficiency of physical and/or chemical sterilization procedures in industry, biomedicine and sanitation [34]. The bacterium is ubiquitous distributed in the natural environment and no specific safety measures are needed due to its harmlessness. Four crop plant species with agricultural relevance and importance for the agricultural market in Germany were chosen for this study: Hordeum vulgare (barley, food and feed grain), Triticum aestivum L. (wheat, food and feed grain), Brassica napus (rapeseed, oil, protein-rich food and feed, biofuel) and Lupinus angustifolius (lupine, protein-rich food and feed).
Finally, inactivation efficiency after direct CAPP was compared to previously reported efficiency using indirect plasma treatment, e.g., plasma processed air.
2. Materials and Methods
2.1. Seed Material
Seeds of Hordeum vulgare L. (barley) accession Kosmos and Triticum aestivum L. (wheat) accession Informer were obtained from Ceravis AG (Rendsburg, Germany), while seeds of Brassica napus L. (rapeseed) accession Atora were provided by NPZ Innovation GmbH (Holtsee, Germany) and seeds of Lupinus angustifolius L. (lupine) accession Boregine from Saatzucht Steinach GmbH (Steinach, Germany).
Chemical pre-treatment was needed for barley seeds due to their high load of naturally occurring bacteria, which interfere with the inoculation and determination of viable spores after CAPP treatment. For sterilization before CAPP treatment, a 5% (v/v) sodium hypochlorite solution (NaClO, Chemsolut Th. Geyer GmbH & Co. KG, Renningen, Germany) was applied to 16 g of barley seeds and incubated for one min while constantly shaking, with one repetition of this process. Following these two incubations, barley seeds were washed three times with 32 mL deionized water and dried under a laminar flow safety cabinet (Herasafe KS 12 class II, Thermo Scientific, Langenselbold, Germany) on Petri dishes for approximately two hours.
2.2. Artificial Inoculation of Seeds with Bacillus atrophaeus Spores and Recovery of Viable Spores from Crop Seeds
The sporulated form of B. atrophaeus (DSM 675) originating from the German collection of microorganisms and cell cultures (DSMZ, Braunschweig, Germany) was used as a model in this study. A stock of spore suspension with concentration of 1 × 109 CFU/mL was stored in sterile NaCl (0.85% w/v) at 7 °C for further use.
Inoculation of seeds was carried out as described previously in Wannicke et al. [35]. In short: a diluted spore stock culture with a density of ~1 × 107 CFU/mL was applied with a defined quantity for each gram of seed (i.e., 20 µL for barley and wheat, 64 µL for rapeseed, 30 g for lupine), followed by vigorously mixing for approximately 30 s to ensure an even distribution of the spore solution. Effective loading on the seed surface with viable spores accounted for 7.41 ± 4.51 × 104 CFU/mL for barley, 5.61 ± 0.71 × 105 CFU/mL for wheat, 2.36 ± 0.39 × 105 CFU/mL for rapeseed and 1.40 ± 0.22 × 105 CFU/mL for lupine seeds. Afterwards, inoculated seeds were dried for 30 min at room temperature in a laminar flow safety cabinet (Herasafe KS 12 class II, Thermo Scientific, Langenselbold, Germany) to allow the attachment of microorganisms on the seed surface. In preparation for treatment, seeds were transferred to pressure-resistant one-liter glass bottles (Duran, Mainz, Germany) using three replicates each with four technical replicates from each individual replicate.
After plasma treatment (see below), recovery of spores from the seed surface was initiated by adding 40 mL of sterile soybean casein digest bouillon CASO broth (Carl Roth, Karlsruhe, Germany) to the bottles and incubating them for 30 min on a horizontal shaker (150 rpm, PSU-20i, Biosan, Riga, Latvia) at room temperature, followed by centrifugation and re-suspension of the pellet. Re-suspended pellets were serially diluted and aliquots (10 µL) of appropriate dilutions were plated on CASO agar plates (Carl Roth, Germany). Agar plates were incubated for 17 h at 30 °C (Binder BD 115, Tuttingen, Germany). Colony-forming units were counted manually. Finally, the logarithmic reduction in CFU was calculated using the difference in decimal logarithm of CFU/mL for CAPP-treated seeds and the CFU/mL determined for control samples without CAPP treatment. A second control was applied using gas flow through the reactors only (Cg), without igniting the plasma.
2.3. Plasma Sources and Treatment of Seeds
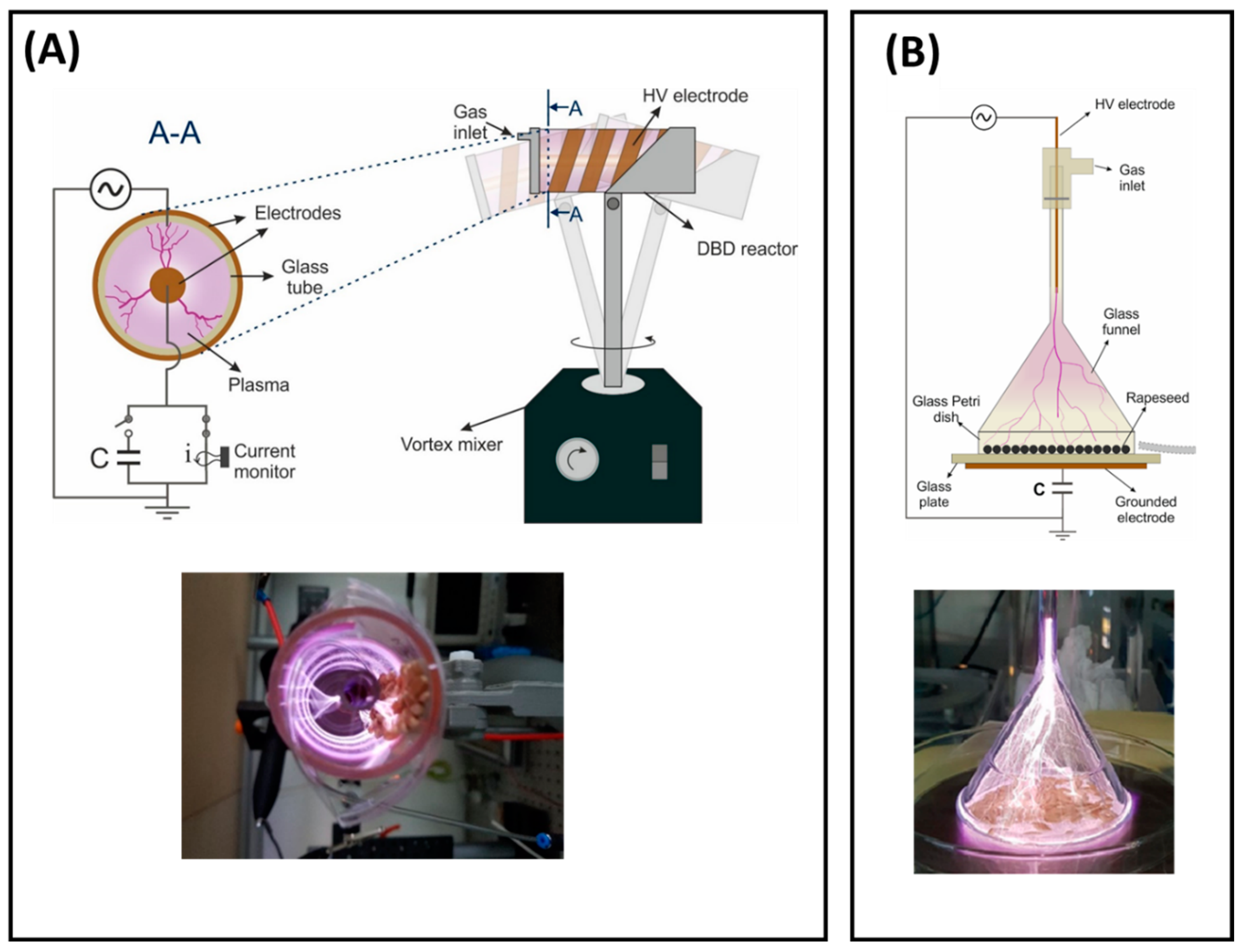
Two plasma sources were employed in this study, a volumetric dielectric barrier discharge (volume-DBD, [32]) and a corona discharge (CD, [33]) reactor. The volume-DBD reactor has a coaxial configuration and has been previously reported in [32]. The reactor consists of a borosilicate glass tube with one end closed and a copper tape glued around it in a spiral shape serving as the high-voltage electrode (Figure 1A). The second electrode comprises a grounded copper rod placed at the central axis and forming a gap of 13 mm. Because of the relatively large gap size for operation in ambient air, the reactor operates with argon using a gas flow of 2 slm. To ensure a homogeneous treatment of the seeds, the reactor was mounted on top of a vortex mixer, allowing the constant shaking of the seeds.
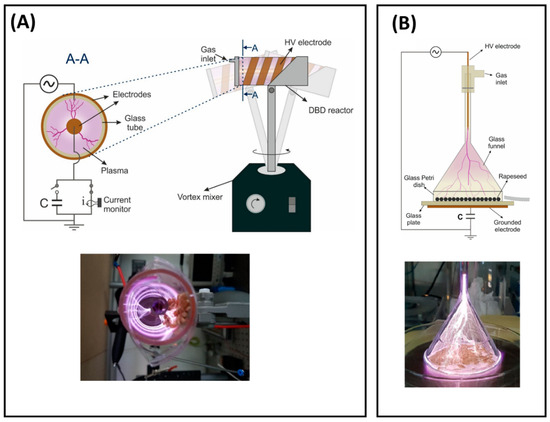
Figure 1.
Schemes and photographs of running plasma sources used in this study: (A) volume-DBD (see [32] for more details) front view (left) and complete set-up (right) with plasma on (below), (B) corona discharge (see [33] for more details) complete set-up (above) and plasma on (below).
The CD reactor consists of an upside-down borosilicate glass funnel mounted on top of a glass Petri dish (Figure 1B), inside of which the seeds are placed. This configuration was reported in [33] for the treatment of seeds from rapeseed. The reactor is set up on top of a 5.85 mm thick glass plate with a grounded copper tape attached on the lower bottom. The high-voltage electrode comprises a tungsten pin inserted in the straight part of the funnel. With a distance between the pin electrode and the Petri dish of more than 10 cm, argon was fed into the reactor via the HV-electrode holder (top part) to facilitate discharge formation. Applying a flow rate of 4 slm helps with the corona filaments’ propagation throughout the funnel, homogenizing their distribution.
Both reactors were operated with an AC power supply (AL-1400-HF-A, Amp-Line, West Nyack, NY, USA) at a frequency of 10 kHz using amplitude-modulated voltage with a repetition period of 100 ms. The voltage amplitude was defined for each reactor separately and chosen as the minimum operating voltage to ignite the discharge when seeds were placed inside of it. Therefore, a voltage of 12 kV p-p and duty cycle of 30% were chosen for the volume-DBD configuration and a voltage of 20 kV p-p and voltage duty cycle of 50% were selected for the corona discharge.
Incubation times of seeds with plasma on were 2, 5 and 10 min. After five minutes of operation with the respective parameters, the temperature in the reactor at the seed position was measured to be around 50 °C in both cases. To avoid the influence of high temperatures potentially originating from the plasma, the 10 min treatments for both reactors were split into 3 phases while the reactor was filled constantly with argon: 5 min induced plasma, 5 min without induced plasma and 5 min induced plasma without opening the reactor. More details on the temperature increase inside of the reactor during seed treatment are reported in [32,33].
In addition, two different control sets for each treatment and crop were implemented for the two plasma sources. The first control sample (control) encompassed artificially contaminated seeds without contact with the plasma sources inoculated with the spore suspension. This control enabled calculation of log reduction after CAPP treatment and served as a reference for total viable spores. The second control sample was a gas control with artificially contaminated seeds inside the plasma device which was filled with argon gas without plasma on for 2 min (Cg 2 min). This control elucidated whether gas flow of argon alone displayed decontaminating properties. A pre-test on the decontamination efficiency of argon gas only (gas flow 2 slm) applying treatment times of two minutes was carried out using the volume-DBD and lupine as a model.
For each parameter, the treatment was established with three biological parallels and four technical replicates each.
2.4. Seed Germination Tests
Maximum germination of CAPP-treated and untreated seeds was surveyed in Petri dishes mounted with layers of filter paper amended with 10 mL (barley, wheat), 12 mL (rapeseed) or 30 mL (lupine, 20 mL bottom + 10 mL top) of tap water. Four replicates with 50 seeds (barley, wheat, rapeseed) or 30 seeds (lupine) each were applied. Samples were incubated inside a climate chamber in the dark (WB, Mytron, Heilbad Heiligenstadt, Germany) at 75% humidity and 22 °C. Germination was monitored after 66 h for barley and wheat and 48 h for rapeseed and lupine. A seed was considered germinated once the radicle protruded ~1 mm in length. Calculation of germination percentage was conducted using max germination % = (n × 100%)/Nt, with n = number of seeds germinated and Nt = total number of seeds.
2.5. Water Contact Angle (WCA) Analysis
The surface hydrophobicity of all investigated plant species was examined by measuring the water contact angle (WCA). To do so, the sessile drop method was applied after CAPP treatment using a goniometer OCA 30 (DataPhysics Instruments, Filderstadt, Germany). The seeds were glued on top of a flat surface, where irregular seeds, such as wheat and barley, were glued with the crease facing down. Three separate drops of 2.0 µL (wheat, barley, lupine) or 0.5 µL (rapeseed) of deionized water were placed on top of each seed on the convex side, and for evaluation, the software SCA 20 (DataPhysics Instruments, Germany) was used. For each treatment parameter, a set of four seeds was analyzed with three measurements made on each seed.
2.6. Statistical Analysis
Comparison of mean values of the studied parameters between CAPP treatments and control/control gas were analyzed using One-Way Analysis of Variance (ANOVA) and subsequent multiple comparisons versus the respective control group (Bonferroni t-test):
In addition, to compare the efficiency of the two different plasma sources regarding WCA, log reduction, maximum germination and WCA, a one-way analysis of covariance (ANCOVA) with plasma source as factor and treatment time as covariates was performed. All statistical tests were two-sided and taken at the 5% level of significance (p ≤ 0.05). Statistical analyses were conducted using SigmaPlot 13 (Systat Software Inc., San Jose, CA, USA).
3. Results
3.1. Inactivation Efficiency of B. atrophaeus by Direct CAPP
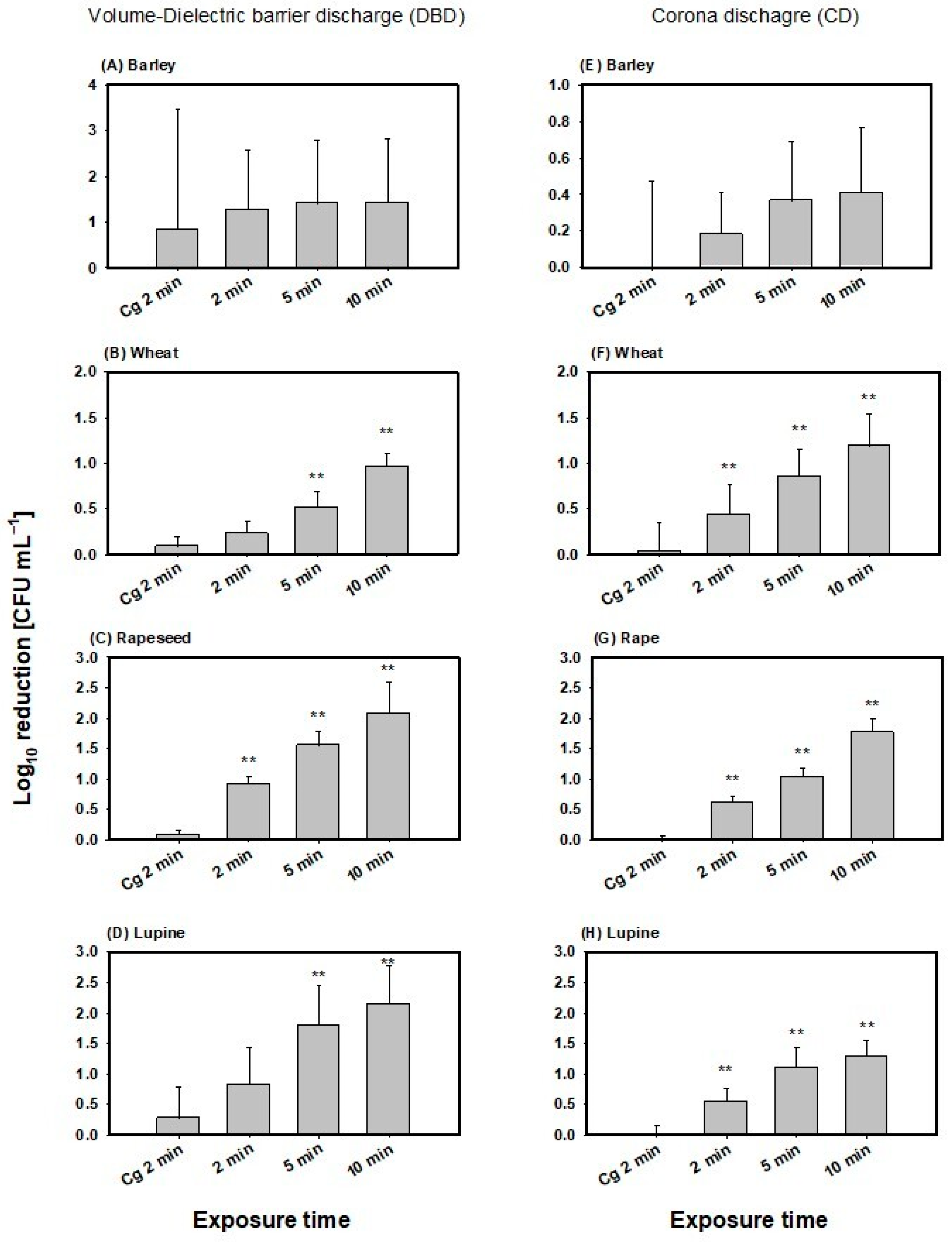
Recovery of viable spores on barley seeds displayed large standard deviations. Therefore, calculation of recovery from CAPP treatments, as well as gas control (Cg), also resulted in high standard deviations for both plasma sources (Figure 2). Inactivation efficiency in both plasma sources showed no significant differences compared to gas flow of argon alone (Cg 2 min 0.85 ± 2.6 log10 reduction CFU/mL), nor with increasing exposure time to CAPP (p = 0.718; Figure 2A,E). Moreover, differences between the two plasma sources were not significant (p = 0.633). The mean inactivation detected for the volume-DBD treatment was 1.37 ± 1.36 log10 reduction CFU/mL and 0.32 ± 0.32 lo10 g reduction CFU/mL for CD treatment across all exposure times to CAPP. Maximum mean decontamination was 1.43 ± 1.38 log10 reduction CFU/mL for 10 min volume-DBD treatment.
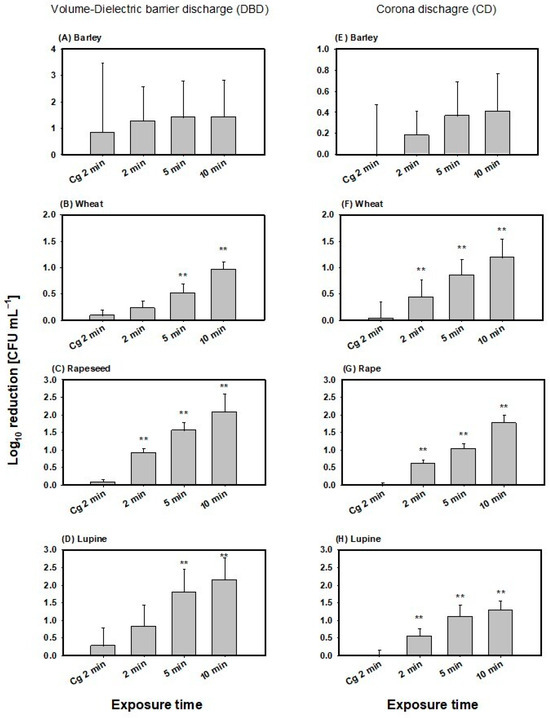
Figure 2.
Inactivation of Bacillus atrophaeus spores for the volume-DBD (left column) and CD (right column) treatment of barley (A,E), wheat (B,F), rapeseed (C,G) and lupine (D,H). The dotted line is equal to the zero reference line. ** denotes statistically significant differences in mean values relative to Cg 2 min with p ≤ 0.01 according to One Way Analysis of variance and multiple comparisons versus the control group (Bonferroni t-test).
For wheat, a significant reduction in B. atrophaeus spores was observed compared to the gas control and 5 and 10 min exposure times for the volume-DBD treatments and all exposure times in the CD treatment (Figure 2B,F). Maximum inactivation occurred after 10 min exposure in both plasma sources with 0.96 ± 0.15 log10 reduction CFU/mL for the volume-DBD and 1.19 ± 0.35 log10 reduction CFU/mL for the CD plasma source. Moreover, One-way-ANCOVA showed that the plasma source type did not significantly affect inactivation efficiency (p = 0.276), with a mean inactivation across all exposure times for volume-DBD treatment of 0.57 ± 0.32 log10 reduction CFU/mL and 0.83 ± 0.41 log10 reduction CFU/mL for CD treatment. On the other hand, exposure time significantly affected inactivation efficiency (One-way-ANCOVA, p ≤ 0.001), with no significant interactions between the factor plasma source and the covariate time of exposure (One-way-ANCOVA, p = 0.999).
Efficiency of inactivation in rapeseed increased with elevated exposure time in both plasma sources and was significantly higher compared to the gas control only (p ≤ 0.01; Figure 2C,G). Apart from this, there was no significant difference in inactivation between the plasma sources (p = 0.116), with mean values across all exposure times for volume-DBD of 1.52 ± 0.55 log10 reduction CFU/mL and 1.14 ± 0.49 log10 reduction CFU/mL for CD. Maximum mean inactivation occurred at 10 min volume-DBD treatment, reaching 2.08 ± 0.42 log10 CFU/mL reduction. Exposure time, on the other hand, displayed a significant impact on inactivation efficiency (One-way-ANCOVA, p ≤ 0.001). Plasma source, as well as the interaction of factor plasma source and co-variate exposure time, was not significant (One-way-ANCOVA, p = 0.118, p = 0.928, respectively).
Inactivation in lupine increased with elevated exposure time with significant differences between gas control and 5 and 10 min exposure time in the volume-DBD treatment, as well as in all exposure time in the CD treatment (p ≤ 0.01; Figure 2D,H). Mean inactivation of the volume-DBD treatment was 1.59 ± 0.77 log10 reduction CFU/mL and 0.98 ± 0.38 log10 reduction CFU/mL for the CD treatment across all exposure times. Maximum mean decontamination was detected at 10 min exposure to the volume-DBD with 2.15 ± 0.51 log10 CFU/mL reduction.
Differences in plasma sources were not significant (One-way ANCOVA, p = 0.664), nor was the interaction of factor plasma source and co-variate exposure time (One-way ANCOVA, p = 0.344). Exposure time displayed significant effects on the inactivation efficiency (One-way ANCOVA, p = 0.004).
3.2. Impact of Direct CAPP Treatment on Seed Viability
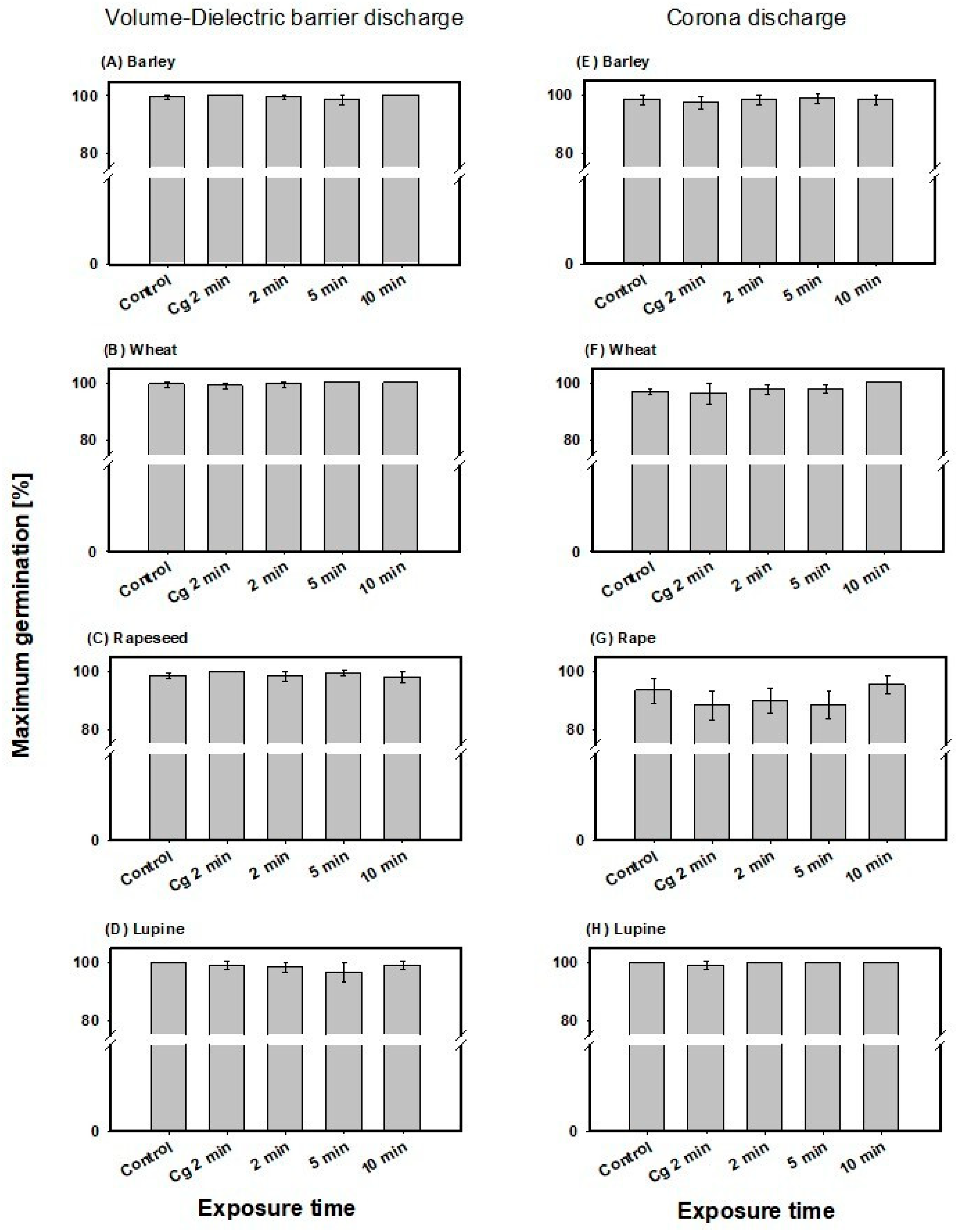
In barley, no significant effect on maximum germination was detected for the volume-DBD or CD treatments (Figure 3A,E). Mean values across all treatment times were 98.7 ± 1.7% for the volume-DBD and 99.3 ± 0.8% for the CD treatment.
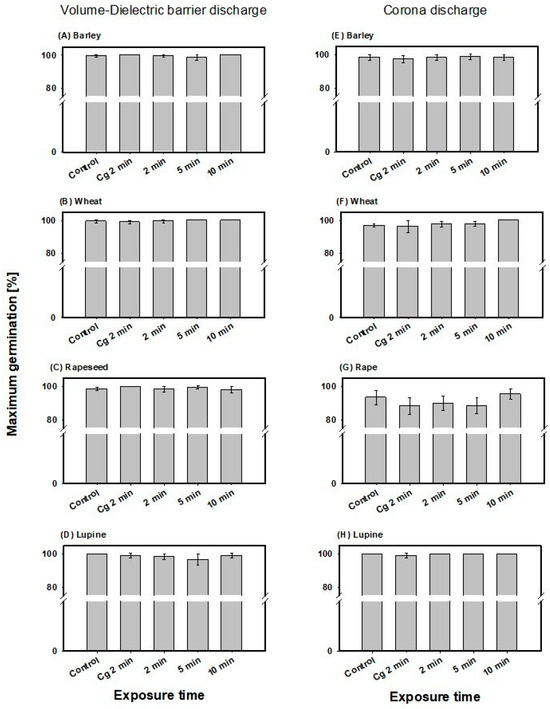
Figure 3.
Maximum germination determined for barley and wheat after 66 h and rapeseed and lupine after 48 h of incubation for the volume-DBD (left column) and CD treatment (right column), as well as the control and gas control (Cg 2 min). Note: no significant differences in mean values of CAPP-treated seeds compared to untreated control or gas control were detected using One-Way-Analysis-of-Variance.
In wheat too, no significant effect on maximum germination was detected for the volume-DBD or CD treatment (Figure 3B,F). Mean maximum germination values across all treatment times were 98.5 ± 1.0% for the volume-DBD and 99.8 ± 0.3% for the CD treatment.
Maximum germination in rapeseed showed no significant alteration after CAPP treatment compared to control and gas control, with mean values across the treatments of 96.9 ± 1.5% for volume-DBD and 91.3 ± 4.0% for the CD treatment (Figure 2C,G). Also, in lupine, no significant effect was detected (Figure 2D,H) after treatment with both CAPP plasma sources. Mean maximum germination of volume-DBD treatment was 100 ± 0% and 98.1 ± 2.1% for CD treatment.
3.3. Impact of Direct CAPP Treatment on Seed Surface Hydrophobicity
A significant decrease in WCA after CAPP treatment was observed for all treatment times compared to the untreated control as well as the gas control for all plant species (Table 1). Overall, CD treatment resulted in larger spread of data of individual replicates and higher standard deviations compared to volume-DBD treatment. In barley and lupine, differences in mean WCA were not significantly dependent on the kind of plasma source (One-way-ANCOVA, p = 0.320; p = 0.591, respectively), nor did increasing treatment time affect mean WCA significantly (One-way-ANCOVA, p = 0.711, p = 0.951, respectively). The mean WCA of wheat seeds significantly depended on plasma source (p = 0.010) and treatment time (One-way-ANCOVA, p = 0.055). Moreover, significant interaction between the factor, plasma source, and a covariate, treatment time, occurred (One-way-ANCOV, p = 0.028). In rapeseed, mean WCA showed significant dependency on treatment time (One-way-ANCOVA, p = 0.045) but not plasma source (One-way-ANCOVA, p = 0.990).

Table 1.
Effect of volume-DBD and CD treatment on the water contact angle (WCA) of the seed surface for the four plant species studied. Values represent mean ± SD of 12 individual measurements for each treatment. Bold numbers indicate a significant difference in mean value relative to the control and gas control at the level of p ≤ 0.05 (One-Way-Analysis-of-Variance, n = 12).
4. Discussion
The two plasma sources compared in this study displayed comparable results regarding inactivation of bacterial spores on four different types of crop seeds, maximum germination and changes in seed surface hydrophilicity. The only statistically significant plasma source effect was observed for wheat seeds and WCA, with the volume-DBD showing significant lower WCA values after treatment compared to the CD plasma source.
Specifically, inactivation of B. atrophaeus spores by direct CAPP treatment displayed different efficiency depending on the crop species tested. Inactivation in barley showed large variations and ranged from 0 to 3.3 log10 CFU/mL, with a mean inactivation of 0.85 ± 1.12 log10 CFU/mL across all treatments, leading to non-significant differences compared to the gas control set-up using argon flow only (Cg 2 min). From five minutes onwards, mean efficiency in inactivation of all other species tested was significantly higher than the gas control. Using a CAPP source with short plasma exposure did not result in significant inactivation of spores, while longer treatment times led to elevated reduction in viability. Overall mean inactivation in wheat, lupine and rapeseed reached 1.1 ± 0.7 log10 CFU/mL, 1.7 ± 0.5 log10 CFU/mL and 1.9 ± 0.4 log10 CFU/mL, respectively, for 10 min volume-DBD, which is equal to 90% to 99% inactivation. On the other hand, inactivation detected for two minutes of direct CAPP treatment did not show significant differences compared to the gas flow control of argon in all crop species except rapeseed. The overall low mean inactivation and large span of data recorded for barley might result from its seed coat structures, which differ from the other crop species tested. Wrinkles or crevices might allow better attachment of spores and/or shielding of spores and microorganisms (Supplementary Figure S1).
Altogether, alteration of plasma process parameters (input voltage, duty cycle, working gas, gas flow, exposure time, pots-storage time) might increase the detected efficiency in our study and might be pursued in a follow-up study.
In general, spore inactivation by direct CAP, from artificially inoculation onto seed surfaces to natural load and to application of pure spore solution, has been extensively demonstrated (e.g., [25,26]). Brasoveanu et al. [36] observed higher plasma inactivation levels of fungal load for smoother corn seeds compared to barley seeds, which was also detected in our study. Moreover, factors such as density of the pathogen loading and spatial distance, as well as the location of the microorganism towards the plasma source, play a role in determining the overall efficiency of inactivation on surfaces, as well as for processes such as pulsed electric fields [37]. Using a DBD with atmospheric air as a working gas, inactivation of a natural fungal of wheat seeds was determined, reaching maximum values of 2.5 log10 CFU/g seeds after 20 min of direct treatment and 24 h of post-treatment retention time, which was accompanied by a reduction in seed germination of 85% [38]. In the same study, a shorter exposure time of five minutes and 24 h post-retention time led to an inactivation of 0.9 log10 CFU/g without negative effects on seed germination. This is in line with results from our study, where CAPP exposure times < 20 min led to inactivation of 0.96 log10 CFU/mL in wheat seeds for the volume-DBD treatment, but with argon as the working gas, there was no impairment of seed germination. Moreover, a nearly complete reduction of Fusarium culmorum spores on wheat and barley seeds was accompanied by a decrease in seed germination of ~54% for treatment times > 120 s using an atmospheric pressure diffuse coplanar surface-DBD with ambient air as a working gas [39]. In the field of post-harvest treatment of spices, inactivation of B. atrophaeus spores on black pepper using a radio frequency (RF) plasma jet resulted in a reduction of viable spores of 1.3 log10 CFU/g peppercorns after 15 min of exposure [40]. Here, viability of seeds was not the target and was not studied. Using the same microorganism in its sporulated form as in our study, B. atrophaeus, Reineke et al. published a reduction of 3.1 log10 after five minutes of exposure using a pure spore solution which was semi-dry inoculated onto Petri-dishes and treated with a non-thermal RF-driven plasma jet with pure argon as a working gas [41]. The higher inactivation of spores compared to our study resulted from the type of plasma source used, as well as the media in which the spores were treated. Similar, high levels of inactivation of dried spore solution of B. atrophaeus on glass slides treated with an atmospheric-pressure Ar/O2 plasma jet were reached, with complete inactivation after 40 s of exposure [42]. Efficient inactivation was achieved using pure cultures of spores and argon as a working gas for CAP treatment due to the relatively higher amounts of reactive nitrogen and oxygen species (RONS), especially hydroxyl radicals (OH●), which might be quenched in the other process gas compositions using pure or mixtures of oxygen, nitrogen or helium (e.g., [41]). Argon as a working gas was also used in our study. Some admixture of air (including oxygen) might also have occurred because the two reactors were not airtight. Commonly detected reactive chemical species of the plasma sources used in this study have been previously published by Nishime et al. [32,33]. Due to its configuration, ambient air was able to diffuse into the reactor of the volume-DBD. Thus, reactive oxygen and nitrogen species (RONS) were produced in the discharge after admixture inside the reactor with weak emission of nitrogen oxide, hydroxyl radicals and molecular dinitrogen, as well as strong argon emission, with preferred energy transfer to oxygen. In the CD reactor, some excited species from air molecules (oxygen and dinitrogen) were observed in the emission spectrum, although the reactor was closed by a Petri dish. A strong argon emission, in addition to hydroxyl radicals, and weak emission of dinitrogen, nitrogen oxide and singlet oxygen was detected. Some of those reactive species possess known decontaminating properties and can oxidize lipids, proteins and nucleic acids and ultimately lead to pathogen destruction (e.g., [43]). More specific, inactivation mechanisms can encompass erosion the surface of microbial cells through etching (e.g., [44]), oxidative damage of intracellular macromolecules, such as membrane lipids, proteins, and DNA, and a reduction in intracellular pH from diffusion into the microbial cells disrupting pH homeostasis (e.g., [45]).
Comparing the inactivation efficiency detected in our study to previously investigated and published indirect CAPP treatment using plasma processed air (PPA), PPA proved to be more efficient in inactivation of B. atrophaeus spores with mean reduction of 3 log10 CFU/mL for barley, which is equal to ~99.99% decontamination, 2.56 ± 0.61 log10 CFU/mL in rapeseed and 2.44 ± 0.52 log10 CFU/mL in lupine (~99.9%) [35]. In contrast to the volume-DBD and CD, PPA is enriched in RNS, including nitride oxide and its possible reaction compounds nitrogen dioxide, nitrous acid and nitric acid. Those components form an acid environment known for favoring the inactivation of spores [46].
Although showing efficient spore reduction, PPA can lead to a reduction in seed vitality, as observed and published for rapeseed, depending on the applied process parameters [35]. In contrast to this, no significant reduction in maximum germination was detected in our study using direct CAPP treatment at any exposure time.
Direct CAPP treatment is also known for its stimulating effect on seed germination potential, germination speed and seedling development (e.g., [26,47,48,49,50]), which has been confirmed and previously published for the CAPP reactors presented in our study as well [32,33].
Moreover, direct CAPP treatment has been shown to change the physicochemical properties of seed surfaces and to increase seed surface hydrophilicity by means of attachment of polar groups such as hydroxyl-, carboxyl- and carbonyl- groups which lead to an increased interaction with water via dipole–dipole intermolecular forces (e.g., [50,51,52,53,54]). Ultimately, water contact angle, a proxy for hydrophilicity of a surface, is significantly decreased below 90° after direct CAPP treatment, indicating facilitated water uptake by the seed. In some cases, stimulated water uptake which is accompanied by reduced WCA has also been reported (e.g., [50,55,56]), while there was no effect on water uptake by seeds even though WCA was reduced after direct CAPP treatment (e.g., [57,58]). Indirect CAPP treatment does not alter the WCA (e.g., [35,57,59]).
This effect of elevated hydrophilicity was also detected in all crop species tested in our study. In wheat, significant differences occurred between the two tested plasma sources, as well as for the different exposure times, with a significantly lower and more homogeneous contact angle after treatment with the volume-DBD. Inhomogeneous treatment was observed, especially for the CD reactor, resulting in large spans of data ranging from, e.g., 30° to 108° in barley for 10 min exposure. In the CD source, plasma filaments are created, possibly leading to a treatment of mainly one side of the seed or an object with restricted treatment of the opposing side. As no shaking and turning of the seeds took place during treatment with the CD reactor, different WCA values for each side of the seed can occur and can lead to a higher span because it cannot be assured that the upper side of the treated seeds is also used in WCA determination. In contrast, seeds are constantly shaken during treatment in the volume-DBD, leading to more homogeneous treatment and effect on the WCA.
Future studies on the two CAPP reactors regarding detailed analysis of water uptake next to WCA, germination speed and uniformity should be pursued to further evaluate its application as a biostimulant. Elevated seed hydrophilicity is considered to be one possible explanation for an increase in germination speed due to facilitated water uptake and swelling, which ultimately initiates the germination process (e.g., [60]). Because WCA displayed significant differences for wheat seeds with lower WCA values for the volume-DBD plasma source, it might be speculated that water uptake, as well as germination speed, might differ between the plasma sources. This needs to be clarified in depth in future studies.
5. Conclusions
Direct treatment as applied in this study using volume-DBD and CD for the purpose of seed hygiene only displayed moderate efficiency for spore-forming microorganisms on the seed surface, with a mean inactivation of ~1 log unit, which is equivalent to a 90% reduction in viability. Efficiency was comparable between both plasma sources and increased with longer exposure times (≥5 min). On the other hand, maximum germination was not impaired at any plasma process parameter, and seed hydrophilicity significantly increased after volume-DBD or CD treatment. Comparing the applied plasma sources with the chosen process parameters to published reductions in spore vitality of B. atrophaeus using indirect treatment with plasma processes air (PPA) for the same crop seed, the inactivation efficiency investigated in this study is distinctly lower than that detected for PPA, which reached sporicidal inactivation of >3 log units (~99.99% inactivation). In the future, the stimulating effect of volume-DBD and CD applied in our study, as well as the observed changes in seed surface hydrophilicity, should be investigated in more depth.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app14219793/s1, Figure S1. Photos of seed used in this study. A and B photos taken with google Pixel 8. C Photo taken using Bresser Advance ICD stereo microscope, 10× magnification.
Author Contributions
Conceptualization, N.W. and H.B.; writing—review and editing, N.W., H.B., T.M.C.N. and J.M.D.; project administration, N.W. and H.B.; funding acquisition, N.W. and H.B.; laboratory work, data acquisition and processing, J.M.D. and T.M.C.N. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the by the German Federal Ministry of Education and Research (BMBF) FZK 03WIR2802B, FKZ 03WIR2805A and FZK 03WIR2812A. Funding for publication was awarded by the Federal Government, the state of Mecklenburg-Vorpommen (FKZ VIII-0639-INP00-2023/004-002).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Richard, B.; Qi, A.; Fitt, B.D. Control of crop diseases through Integrated Crop Management to deliver climate-smart farming systems for low-and high-input crop production. Plant Pathol. 2022, 71, 187–206. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Oerke, E.-C.; Dehne, H.-W. Safeguarding production—Losses in major crops and the role of crop protection. Crop Prot. 2004, 23, 275–285. [Google Scholar] [CrossRef]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal Agents in Agriculture: Friends and Foes of Public Health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef]
- Jørgensen, L.N.; Heick, T.M. Azole Use in Agriculture, Horticulture, and Wood Preservation–Is It Indispensable? Front. Cell. Infect. Microbiol. 2021, 11, 730297. [Google Scholar] [CrossRef]
- Lopez-Antia, A.; Ortiz-Santaliestra, M.E.; Mougeot, F.; Camarero, P.R.; Mateo, R. Birds Feeding on Tebuconazole Treated Seeds Have Reduced Breeding Output. Environ. Pollut. 2021, 271, 116292. [Google Scholar] [CrossRef]
- Bryson, R.; Brix, H. Challenges and prospects for fungicidal control of wheat diseases. In Integrated Disease Management of Wheat and Barley; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; pp. 239–254. [Google Scholar]
- Chakraborty, S. Migrate or evolve: Options for plant pathogens under climate change. Glob. Change Biol. 2013, 19, 1985–2000. [Google Scholar] [CrossRef]
- Garrett, K.A.; Thomas-Sharma, S.; Forbes, G.A.; Nopsa, J.H. Climate change and plant pathogen invasions. In Invasive Species and Global Climate Change; CABI: Wallingford, UK, 2014; pp. 22–44. [Google Scholar]
- Singh, B.K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J.E.; Liu, H.; Trivedi, P. Climate change impacts on plant pathogens, food security and paths forward. Nat. Rev. Microbiol. 2023, 21, 640–656. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Corrales, D.C.; Soltani, E. Biological seed treatments promote crop establishment and yield: A global meta-analysis. Agron. Sustain. Dev. 2022, 42, 45. [Google Scholar] [CrossRef]
- Rosa, S.; Pesaresi, P.; Mizzotti, C.; Bulone, V.; Mezzetti, B.; Baraldi, E.; Masiero, S. Game-changing alternatives to conventional fungicides: Small RNAs and short peptides. Trends Biotechnol. 2022, 40, 320–337. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Black, M.; Halmer, P. The Encyclopedia of Seeds: Science, Technology and Uses; CABI: Wallingford, UK, 2006. [Google Scholar]
- Jahn, M.; Koch, E.; Blum, H.; Nega, E.; Wilbois, K.-P. Leitfaden Saatgutgesundheit im Ökologischen Landbau-Gemüsekulturen; Forschungsinstitut für biologischen Landbau eV, FiBL Deutschland eV: Frankfurt am Main, Germany, 2007. [Google Scholar]
- Yamagishi, N.; Fujinaga, M.; Ishiyama, Y.; Ogiso, H.; Sato, T.; Tosa, Y. Life cycle and control of Colletotrichum nymphaeae, the causal agent of celery stunt anthracnose. J. Gen. Plant Pathol. 2015, 81, 279–286. [Google Scholar] [CrossRef]
- Schaal, R. Beizung auf Elektronisch. agrarzeitung. 11 August 2023. Available online: https://www.agrarzeitung.de/pflanzenbautipps/getreide/physikalisch-biologisches-verfahren-beizung-auf-elektronisch-108324 (accessed on 28 August 2024).
- Wagner, R.; Weihe, T.; Winter, H.; Weit, C.; Ehlbeck, J.; Schnabel, U. Reducing Storage Losses of Organic Apples by Plasma Processed Air (PPA). Appl. Sci. 2023, 13, 12654. [Google Scholar] [CrossRef]
- Ziuzina, D.; Misra, N.; Cullen, P.; Keener, K.M.; Mosnier, J.; Vilaró, I.; Gaston, E.; Bourke, P. Demonstrating the potential of industrial scale in-package atmospheric cold plasma for decontamination of cherry tomatoes. Plasma Med. 2016, 6, 397–412. [Google Scholar] [CrossRef]
- Cullen, P.J.; Lalor, J.; Scally, L.; Boehm, D.; Milosavljević, V.; Bourke, P.; Keener, K. Translation of plasma technology from the lab to the food industry. Plasma Process. Polym. 2018, 15, 1700085. [Google Scholar] [CrossRef]
- Deng, X.; Shi, J.; Kong, M.G. Physical mechanisms of inactivation of Bacillus subtilis spores using cold atmospheric plasmas. IEEE Trans. Plasma Sci. 2006, 34, 1310–1316. [Google Scholar] [CrossRef]
- Laroussi, M. Low temperature plasma-based sterilization: Overview and state-of-the-art. Plasma Process. Polym. 2005, 2, 391–400. [Google Scholar] [CrossRef]
- Moisan, M.; Barbeau, J.; Crevier, M.-C.; Pelletier, J.; Philip, N.; Saoudi, B. Plasma sterilization. Methods and mechanisms. Pure Appl. Chem. 2002, 74, 349–358. [Google Scholar] [CrossRef]
- Fridman, G.; Brooks, A.D.; Balasubramanian, M.; Fridman, A.; Gutsol, A.; Vasilets, V.N.; Ayan, H.; Friedman, G. Comparison of direct and indirect effects of non-thermal atmospheric-pressure plasma on bacteria. Plasma Process. Polym. 2007, 4, 370–375. [Google Scholar] [CrossRef]
- Adhikari, B.; Pangomm, K.; Veerana, M.; Mitra, S.; Park, G. Plant Disease Control by Non-Thermal Atmospheric-Pressure Plasma. Front. Plant Sci. 2020, 11, 77. [Google Scholar] [CrossRef]
- Brust, H.; Wannicke, N.; Park, G. Agriculture and Food Processing Applications. In Plasma Biosciences and Medicine; Springer: Singapore, 2023; pp. 111–227. [Google Scholar]
- Jo, Y.-K.; Cho, J.; Tsai, T.-C.; Staack, D.; Kang, M.-H.; Roh, J.-H.; Shin, D.-B.; Cromwell, W.; Gross, D. A Non-thermal Plasma Seed Treatment Method for Management of a Seedborne Fungal Pathogen on Rice Seed. Crop Sci. 2014, 54, 796–803. [Google Scholar] [CrossRef]
- Zahoranová, A.; Henselová, M.; Hudecová, D.; Kaliňáková, B.; Kováčik, D.; Medvecká, V.; Černák, M. Effect of Cold Atmospheric Pressure Plasma on the Wheat Seedlings Vigor and on the Inactivation of Microorganisms on the Seeds Surface. Plasma Chem. Plasma Process. 2016, 36, 397–414. [Google Scholar] [CrossRef]
- Zahoranová, A.; Hoppanová, L.; Šimončicová, J.; Tučeková, Z.; Medvecká, V.; Hudecová, D.; Kaliňáková, B.; Kováčik, D.; Černák, M. Effect of Cold Atmospheric Pressure Plasma on Maize Seeds: Enhancement of Seedlings Growth and Surface Microorganisms Inactivation. Plasma Chem. Plasma Process. 2018, 38, 969–988. [Google Scholar] [CrossRef]
- Kim, J.-W.; Puligundla, P.; Mok, C. Effect of corona discharge plasma jet on surface-borne microorganisms and sprouting of broccoli seeds. J. Sci. Food Agric. 2017, 97, 128–134. [Google Scholar] [CrossRef]
- Puligundla, P.; Kim, J.-W.; Mok, C. Effect of corona discharge plasma jet treatment on decontamination and sprouting of rapeseed (Brassica napus L.) seeds. Food Control 2017, 71, 376–382. [Google Scholar] [CrossRef]
- Nishime, T.M.C.; Wannicke, N.; Horn, S.; Weltmann, K.D.; Brust, H. A Coaxial Dielectric Barrier Discharge Reactor for Treatment of Winter Wheat Seeds. Appl. Sci. 2020, 10, 7133. [Google Scholar] [CrossRef]
- Nishime, T.M.; Werner, J.; Wannicke, N.; Mui, T.S.; Kostov, K.G.; Weltmann, K.-D.; Brust, H. Characterization and optimization of a conical corona reactor for seed treatment of rapeseed. Appl. Sci. 2022, 12, 3292. [Google Scholar] [CrossRef]
- ISO 11138-4; Sterilization of Health Care Products—Biological Indicators—Part 4: Biological Indicators for Dry Heat Sterilization Processes. International Organization for Standardization: Geneva, Switzerland, 2017.
- Wannicke, N.; Wagner, R.; Stachowiak, J.; Nishime, T.M.C.; Ehlbeck, J.; Weltmann, K.-D.; Brust, H. Efficiency of plasma-processed air for biological decontamination of crop seeds on the premise of unimpaired seed germination. Plasma Process. Polym. 2021, 18, 2000207. [Google Scholar] [CrossRef]
- Braşoveanu, M.; Nemţanu, M.; Surdu-Bob, C.; Karaca, G.; Erper, I. Effect of glow discharge plasma on germination and fungal load of some cereal seeds. Rom. Rep. Phys. 2015, 67, 617–624. [Google Scholar]
- Leadley, C.E.; Williams, A. Pulsed electric field processing, power ultrasound and other emerging technologies. Food Process. Handb. 2006, 201, 5–20. [Google Scholar]
- Los, A.; Ziuzina, D.; Akkermans, S.; Boehm, D.; Cullen, P.J.; Van Impe, J.; Bourke, P. Improving microbiological safety and quality characteristics of wheat and barley by high voltage atmospheric cold plasma closed processing. Food Res. Int. 2018, 106, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Hoppanová, L.; Medvecká, V.; Dylíková, J.; Hudecová, D.; Kaliňáková, B.; Kryštofová, S.; Zahoranová, A. Low-temperature plasma applications in chemical fungicide treatment reduction. Acta Chim. Slovaca 2020, 13, 26–33. [Google Scholar] [CrossRef]
- Hertwig, C.; Reineke, K.; Ehlbeck, J.; Knorr, D.; Schluter, O. Decontamination of whole black pepper using different cold atmospheric pressure plasma applications. Food Control 2015, 55, 221–229. [Google Scholar] [CrossRef]
- Reineke, K.; Langer, K.; Hertwig, C.; Ehlbeck, J.; Schlüter, O. The impact of different process gas compositions on the inactivation effect of an atmospheric pressure plasma jet on Bacillus spores. Innov. Food Sci. Emerg. Technol. 2015, 30, 112–118. [Google Scholar] [CrossRef]
- Lim, J.-P.; Uhm, H.S.; Li, S.-Z. Influence of oxygen in atmospheric-pressure argon plasma jet on sterilization of Bacillus atrophaeous spores. Phys. Plasmas 2007, 14, 093504. [Google Scholar] [CrossRef]
- Liao, X.; Liu, D.; Xiang, Q.; Ahn, J.; Chen, S.; Ye, X.; Ding, T. Inactivation mechanisms of non-thermal plasma on microbes: A review. Food Control 2017, 75, 83–91. [Google Scholar] [CrossRef]
- Moisan, M.; Barbeau, J.; Moreau, S.; Pelletier, J.; Tabrizian, M.; Yahia, L. Low-temperature sterilization using gas plasmas: A review of the experiments and an analysis of the inactivation mechanisms. Int. J. Pharm. 2001, 226, 1–21. [Google Scholar] [CrossRef]
- Laroussi, M.; Leipold, F. Evaluation of the roles of reactive species, heat, and UV radiation in the inactivation of bacterial cells by air plasmas at atmospheric pressure. Int. J. Mass Spectrom. 2004, 233, 81–86. [Google Scholar] [CrossRef]
- Ehlbeck, J.; Brandenburg, R.; von Woedtke, T.; Krohmann, U.; Stieber, M.; Weltmann, K.-D. PLASMOSE-antimicrobial effects of modular atmospheric plasma sources. GMS Krankenhaushygiene Interdiszip. 2008, 3, 1–12. [Google Scholar]
- Adhikari, B.; Adhikari, M.; Park, G. The Effects of Plasma on Plant Growth, Development, and Sustainability. Appl. Sci. 2020, 10, 6045. [Google Scholar] [CrossRef]
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M. Plasma Agriculture from Laboratory to Farm: A Review. Processes 2020, 8, 1002. [Google Scholar] [CrossRef]
- Starič, P.; Vogel-Mikuš, K.; Mozetič, M.; Junkar, I. Effects of Nonthermal Plasma on Morphology, Genetics and Physiology of Seeds: A Review. Plants 2020, 9, 1736. [Google Scholar] [CrossRef] [PubMed]
- Brust, H.; Nishime, T.M.C.; Wannicke, N.; Mui, T.S.M.; Horn, S.; Quade, A.; Weltmann, K.D. A medium-scale volume dielectric barrier discharge system for short-term treatment of cereal seeds indicates improved germination performance with long-term effects. J. Appl. Phys. 2021, 129, 044904. [Google Scholar] [CrossRef]
- Gómez-Ramírez, A.; López-Santos, C.; Cantos, M.; García, J.L.; Molina, R.; Cotrino, J.; Espinós, J.; González-Elipe, A.R. Surface chemistry and germination improvement of Quinoa seeds subjected to plasma activation. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Molina, R.; Lalueza, A.; López-Santos, C.; Ghobeira, R.; Cools, P.; Morent, R.; de Geyter, N.; González-Elipe, A.R. Physicochemical surface analysis and germination at different irrigation conditions of DBD plasma-treated wheat seeds. Plasma Process. Polym. 2020, 18, e2000086. [Google Scholar] [CrossRef]
- Štěpánová, V.; Slavíček, P.; Kelar, J.; Prášil, J.; Smékal, M.; Stupavská, M.; Jurmanová, J.; Černák, M. Atmospheric pressure plasma treatment of agricultural seeds of cucumber (Cucumis sativus L.) and pepper (Capsicum annuum L.) with effect on reduction of diseases and germination improvement. Plasma Process. Polym. 2018, 15, 1700076. [Google Scholar] [CrossRef]
- Recek, N.; Vesel, A.; Zaplotnik, R.; Paul, D.; Primc, G.; Gselman, P.; Mozetič, M. Hydrophilization of corn seeds by non-equilibrium gaseous plasma. Chem. Biol. Technol. Agric. 2021, 8, 32. [Google Scholar] [CrossRef]
- Švubová, R.; Kyzek, S.; Medvecká, V.; Slováková, Ľ.; Gálová, E.; Zahoranová, A. Novel insight at the Effect of Cold Atmospheric Pressure Plasma on the Activity of Enzymes Essential for the Germination of Pea (Pisum sativum L. cv. Prophet) Seeds. Plasma Chem. Plasma Process. 2020, 40, 1221–1240. [Google Scholar] [CrossRef]
- Švubová, R.; Slováková, Ľ.; Holubová, Ľ.; Rovňanová, D.; Gálová, E.; Tomeková, J. Evaluation of the Impact of Cold Atmospheric Pressure Plasma on Soybean Seed Germination. Plants 2021, 10, 177. [Google Scholar] [CrossRef]
- Los, A.; Ziuzina, D.; Boehm, D.; Cullen, P.J.; Bourke, P. Investigation of mechanisms involved in germination enhancement of wheat (Triticum aestivum) by cold plasma: Effects on seed surface chemistry and characteristics. Plasma Process. Polym. 2019, 16, 1800148. [Google Scholar] [CrossRef]
- Ji, S.H.; Ki, S.H.; Kang, M.H.; Choi, J.S.; Park, Y.; Oh, J.; Kim, S.B.; Yoo, S.J.; Choi, E.H.; Park, G. Characterization of physical and biochemical changes in plasma treated spinach seed during germination. J. Phys. D Appl. Phys. 2018, 51, 145205. [Google Scholar] [CrossRef]
- Pawlat, J.; Starek, A.; Sujak, A.; Kwiatkowski, M.; Terebun, P.; Budzeń, M. Effects of atmospheric pressure plasma generated in GlidArc reactor on Lavatera thuringiaca L. seeds’ germination. Plasma Process. Polym. 2018, 15, 1700064. [Google Scholar] [CrossRef]
- Bewley, J.; Bradford, K.J.; Hilhorst, H.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; Volume 10, p. 978-1. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).