
Application of the Drosophila melanogaster Research Model to Evaluate the Toxicity Levels between Lead and Copper

 ,
,  ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical Products
2.2. Stocks
2.3. Preparing the Cultural Environment
2.4. Growth Medium and Experimental Variants of the Metals Cu2+ (CuSO4) and Pb2+ (C2H3O2)2
- -
- wheat flour-based control medium [55].
- -
- control medium based on wheat flour supplemented with a solution of copper sulphate CuSO4-5H2O (0.50 mM, 0.75 mM, 1.00 mM, and 2.00 mM);
- -
- control medium based on wheat flour supplemented with lead acetate solution (Pb(C2H3O2)2-3H2O (0.50 mM, 0.75 mM, 1.00 mM, and 2.00 mM).
2.5. Determination of the Inhibition Index (IC50) of Proliferation after Exposure to Cu2+ (CuSO4) and Pb2+ Pb(C2H3O2)2
2.6. Statistical Calculation Methods Used to Determine the Levels of Proliferation and Toxicity between Cu2+ and Pb2+
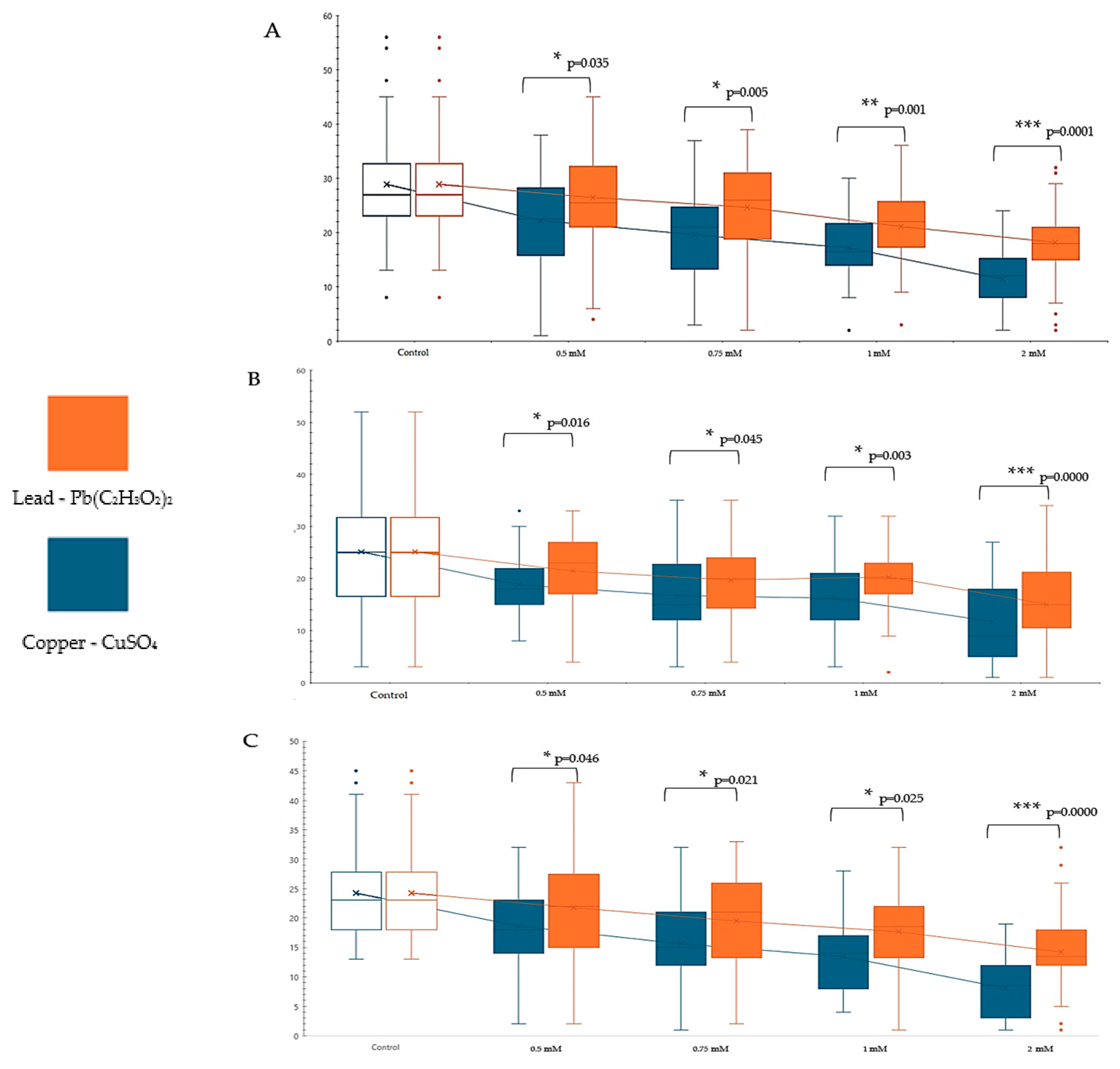
3. Results
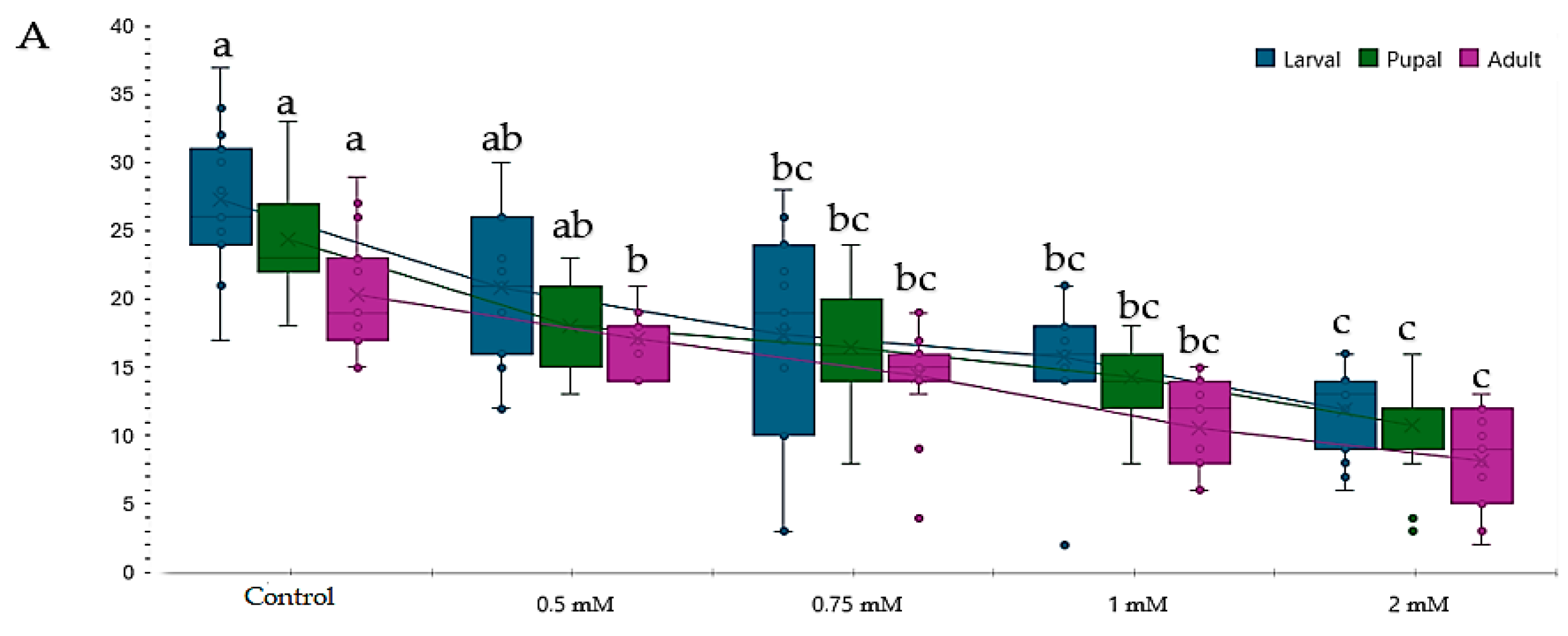
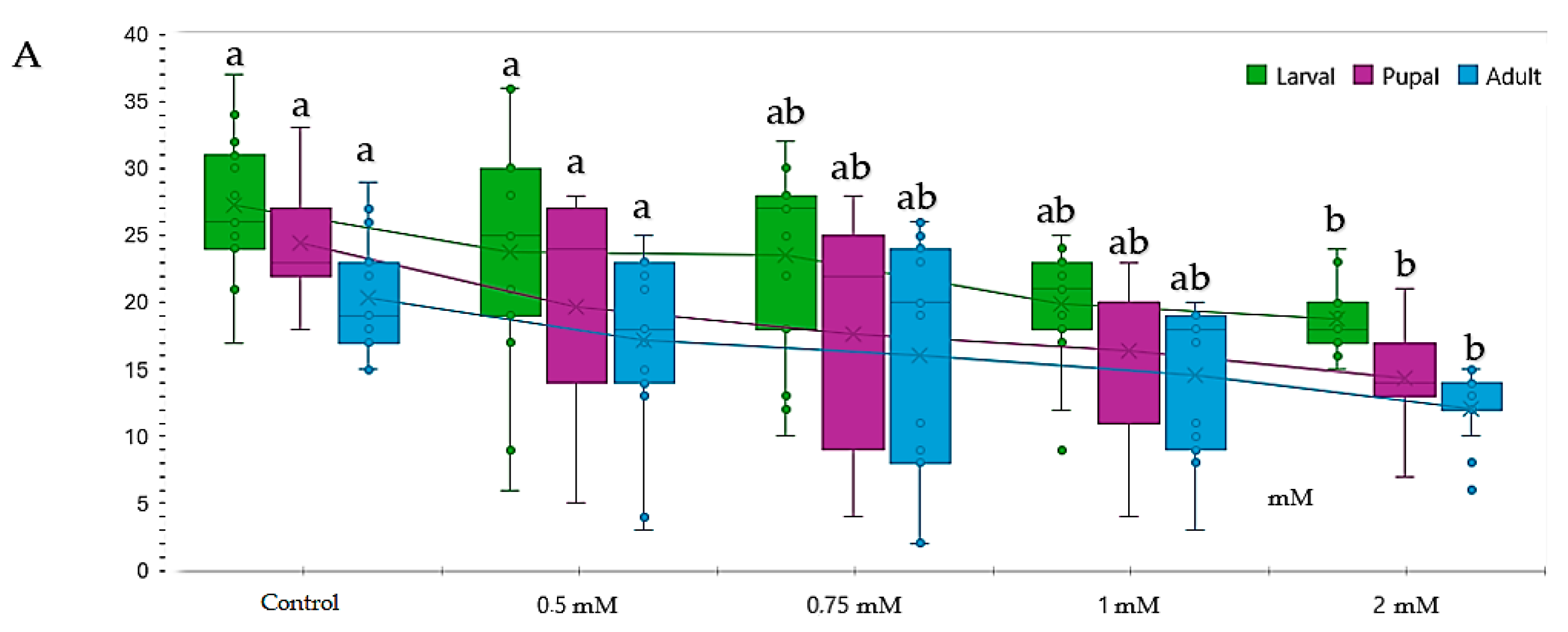
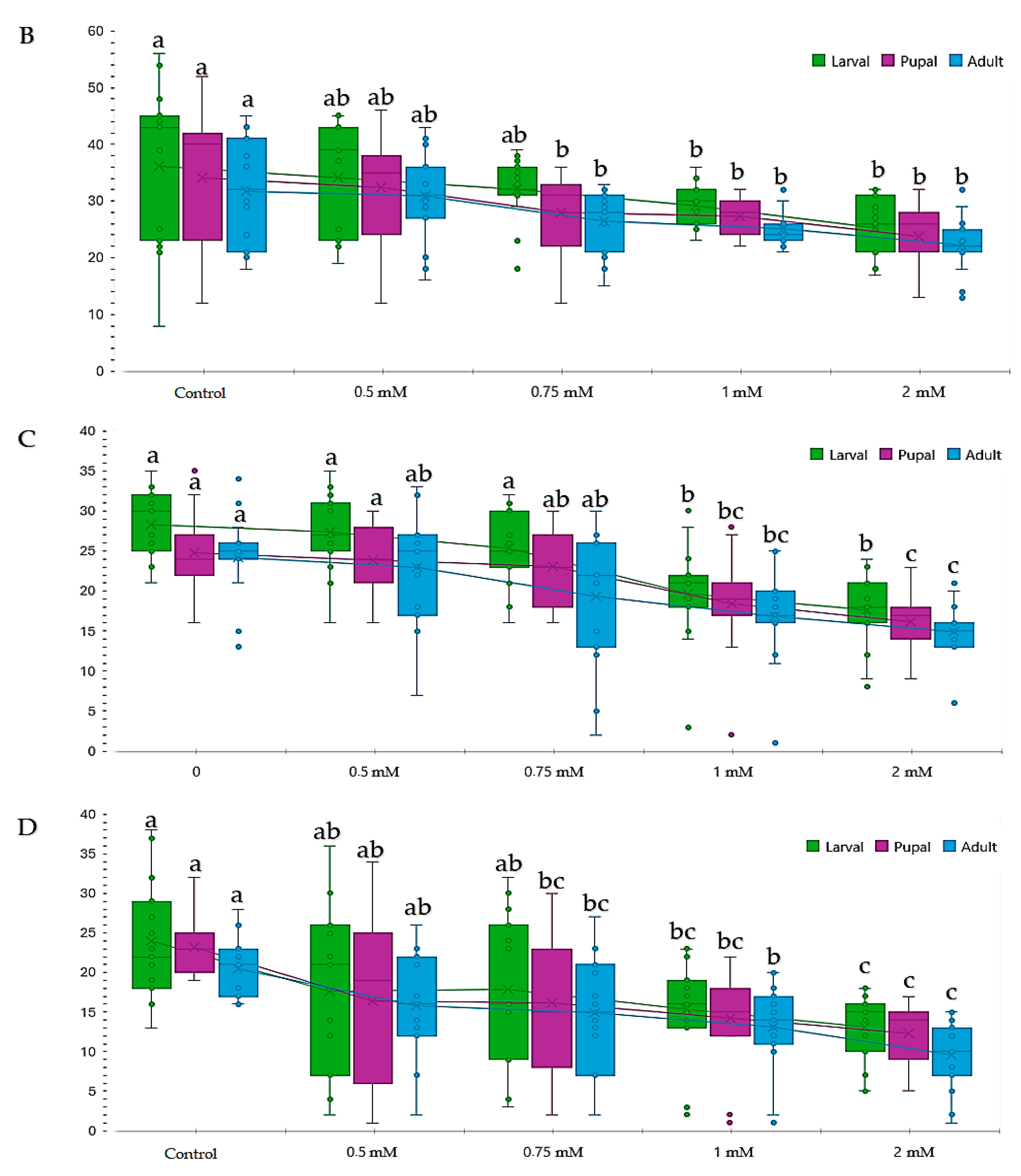
3.1. Influence of Genotype, Concentration, and Type of Metal Cu2+ and Pb2+ on Proliferation
3.2. Influence of Copper (CuSO4) on Proliferation during the Life Cycle (Larva III-Pupal-Adult)
3.3. Influence of Lead Pb(C2H3O2)2 on Proliferation during the Life Cycle (Larva III-Pupa-Adult)
3.4. Comparative Study of Different Concentrations of Lead and Copper Toxic Effects on Drosophila melanogaster
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Ali, M.M.; Hossain, D.; Al-Imran, A.; Khan, M.S.; Begum, M.; Osman, M.H. Environmental pollution with heavy metals: A public health concern. In Heavy Metals—Their Environmental Impacts and Mitigation; IntechOpen: London, UK, 2021; pp. 771–783. [Google Scholar]
- Jung, J.-M.; Kim, C.-J.; Chung, C.-S.; Kim, T.; Choi, K.-Y. Heavy metal characterization of land-based waste dumped at three ocean dumping sites in the Republic of Korea. Mar. Pollut. Bull. 2023, 193, 115205. [Google Scholar] [CrossRef]
- Alkhanjaf, A.A.M.; Sharma, S.; Sharma, M.; Kumar, R.; Arora, N.K.; Kumar, B.; Umar, A.; Baskoutas, S.; Mukherjee, T.K. Microbial strategies for copper pollution remediation: Mechanistic insights and recent advances. Environ. Pollut. 2024, 346, 123588. [Google Scholar] [CrossRef]
- McFarland, M.J.; Hauer, M.E.; Reuben, A. Half of US population exposed to adverse lead levels in early childhood. Proc. Natl. Acad. Sci. USA 2022, 119, e2118631119. [Google Scholar] [CrossRef]
- Karahan, F. Evaluation of Trace Element and Heavy Metal Levels of Some Ethnobotanically Important Medicinal Plants Used as Remedies in Southern Turkey in Terms of Human Health Risk. Biol. Trace Element Res. 2022, 201, 493–513. [Google Scholar] [CrossRef]
- Harmonisation, P.T. Derivation Methods of Soil Screening Values in Europe. Available online: https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=1315ea61e2c6e168e151175735cb6253ddef33d7 (accessed on 2 April 2024).
- Sexton, J.P.; Hangartner, S.B.; Hoffmann, A.A. Genetic Isolation by Environment or Distance: Which Pattern of Gene Flow is Most Common? Evolution 2013, 68, 1–15. [Google Scholar] [CrossRef]
- Huang, Y.; Zhu, M.; Ji, M.; Fan, J.; Xie, J.; Wei, X.; Jiang, X.; Xu, J.; Chen, L.; Yin, R.; et al. Air Pollution, Genetic Factors, and the Risk of Lung Cancer A Prospective Study in the UK Biobank. Am. J. Respir. Crit. Care Med. 2021, 204, 817–825. [Google Scholar] [CrossRef]
- Brumberg, H.L.; Karr, C.J.; Bole, A.; Ahdoot, S.; Balk, S.J.; Bernstein, A.S.; Byron, L.G.; Landrigan, P.J.; Marcus, S.M.; Nerlinger, A.L.; et al. Ambient Air Pollution: Health Hazards to Children. Pediatrics 2021, 147, e2021051484. [Google Scholar] [CrossRef]
- Rajendran, S.; Priya, T.; Khoo, K.S.; Hoang, T.K.; Ng, H.-S.; Munawaroh, H.S.H.; Karaman, C.; Orooji, Y.; Show, P.L. A critical review on various remediation approaches for heavy metal contaminants removal from contaminated soils. Chemosphere 2021, 287, 132369. [Google Scholar] [CrossRef] [PubMed]
- Bhat, V.S.; Kudva, A.K.; Naik, H.V.; Raghu, S.V.; De Padova, P.; Hegde, G. Toxicological Profiling of Onion-Peel-Derived Mesoporous Carbon Nanospheres Using In Vivo Drosophila melanogaster Model. Appl. Sci. 2022, 12, 1528. [Google Scholar] [CrossRef]
- Curcio, R.; Lunetti, P.; Zara, V.; Ferramosca, A.; Marra, F.; Fiermonte, G.; Cappello, A.R.; De Leonardis, F.; Capobianco, L.; Dolce, V. Drosophila melanogaster Mitochondrial Carriers: Similarities and Differences with the Human Carriers. Int. J. Mol. Sci. 2020, 21, 6052. [Google Scholar] [CrossRef]
- Everman, E.R.; Macdonald, S.J.; Kelly, J.K. The genetic basis of adaptation to copper pollution in Drosophila melanogaster. Front. Genet. 2023, 14, 1144221. [Google Scholar] [CrossRef] [PubMed]
- Alaraby, M.; Hernández, A.; Marcos, R. Copper oxide nanoparticles and copper sulphate act as antigenotoxic agents in drosophila melanogaster. Environ. Mol. Mutagen. 2017, 58, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Balinski, M.A.; Woodruff, R.C. Differential sexual survival of Drosophila melanogaster on copper sulfate. Genetica 2017, 145, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Gaetke, L.M.; Chow-Johnson, H.S.; Chow, C.K. Copper: Toxicological relevance and mechanisms. Arch. Toxicol. 2014, 88, 1929–1938. [Google Scholar] [CrossRef]
- Soylak, M.; Kirnap, M. Serum Copper and Zinc Concentrations of Patients with Rheumatoid Arthritis from Kayseri_Turkey. Fresenius Environ. Bull. 2001, 10, 409–410. [Google Scholar]
- Swain, J.; Gutteridge, J.M. Prooxidant iron and copper, with ferroxidase and xanthine oxidase activities in human atherosclerotic material. FEBS Lett. 1995, 368, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Lavrent`yeva, S.I.; Ivachenko, L.Y.; Golokhvast, K.S.; Nawaz, M.A. Ribonuclease activity of Glycine max and Glycine soja sprouts as a marker adaptation to copper sulphate and zinc sulphate toxicity. Biochem. Syst. Ecol. 2019, 83, 66–70. [Google Scholar] [CrossRef]
- Fiskin, K.; Kandemir, S.; Hamamci, D.; Yesilada, E.; Bozcuk, A. Age-related changes in catalase, glutathione reductase activities, the amount of glutathione in total body of oregon and vestigial Drosophila melanogaster. Arch. Gerontol. Geriatr. 1994, 19, 85–90. [Google Scholar] [CrossRef]
- Nwaka, S.; Hudson, A. Innovative lead discovery strategies for tropical diseases. Nat. Rev. Drug Discov. 2006, 5, 941–955. [Google Scholar] [CrossRef]
- Al-Saleh, I.; Nester, M.; Devol, E.; Shinwari, N.; Al-Shahria, S. Determinants of Blood Lead Levels in Saudi Arabian Schoolgirls. Int. J. Occup. Environ. Health 1999, 5, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Sekara, A.; Poniedzialeek, M.; Ciura, J.; Jedrszczyk, E. Cadmium and Lead Accumulation and Distribution in the Organs of Nine Crops: Implications for Phytoremediation. Pol. J. Environ. Stud. 2005, 14, 509–516. [Google Scholar]
- Geng, Y.; Faber, K.N.; de Meijer, V.E.; Blokzijl, H.; Moshage, H. How does hepatic lipid accumulation lead to lipotoxicity in non-alcoholic fatty liver disease? Hepatol. Int. 2021, 15, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Rehman, K.; Fatima, F.; Waheed, I.; Akash, M.S.H. Prevalence of exposure of heavy metals and their impact on health consequences. J. Cell. Biochem. 2018, 119, 157–184. [Google Scholar] [CrossRef] [PubMed]
- Flora, G.; Gupta, D.; Tiwari, A. Toxicity of lead: A review with recent updates. Interdiscip. Toxicol. 2012, 5, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Segin, T.B.; Hnatush, S.O.; Gorishniy, M.B. The processes of lipid peroxidation in the cells of Chlorobium limicola IMV K-8 under the influence of copper (II) sulphate. Biosyst. Divers. 2016, 24, 72–77. [Google Scholar] [CrossRef]
- Siddique, H.R.; Gupta, S.C.; Mitra, K.; Bajpai, V.K.; Mathur, N.; Murthy, R.C.; Saxena, D.K.; Chowdhuri, D.K. Adverse effect of tannery waste leachates in transgenic Drosophila melanogaster: Role of ROS in modulation of Hsp70, oxidative stress and apoptosis. J. Appl. Toxicol. 2008, 28, 734–748. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.P.; Himalian, R.; Shabir, S.; Obaid, A.A.; Alamri, A.S.; Galanakis, C.M.; Singh, S.K.; Vamanu, E. Protection of Phytoextracts against Rotenone-Induced Organismal Toxicities in Drosophila melanogaster via the Attenuation of ROS Generation. Appl. Sci. 2022, 12, 9822. [Google Scholar] [CrossRef]
- Mackay, T.F.C.; Huang, W. Charting the genotype-phenotype map: Lessons from the Drosophila melanogaster Genetic Reference Panel. Wiley Interdiscip Rev. Dev. Biol. 2018, 7, e289. [Google Scholar] [CrossRef]
- Kondrashov, A.S.; Houle, D. Genotype—Environment interactions and the estimation of the genomic mutation rate in Drosophila melanogaster. Proc. R. Soc. B Biol. Sci. 1994, 258, 221–227. [Google Scholar] [CrossRef]
- Anderson, S.; Sadinski, W.; Shugart, L.; Brussard, P.; Depledge, M.; Ford, T.; Hose, J.; Stegeman, J.; Suk, W.; Wirgin, I.; et al. Genetic and Molecular Ecotoxicology: A Research Framework. Environ. Health Perspect. 1994, 102, 3–8. [Google Scholar] [CrossRef]
- Wipf, I.J. Investigating the Role of ATP-Binding Cassette Transporters in Drosophila melanogaster Testis Stem Cells. Master’s Thesis, University of Northern Colorado, Greeley, CO, USA, 2021. [Google Scholar]
- Licht, A.; Schneider, E. ATP binding cassette systems: Structures, mechanisms, and functions. Cent. Eur. J. Biol. 2011, 6, 785–801. [Google Scholar] [CrossRef]
- Kang, J.; Park, J.; Choi, H.; Burla, B.; Kretzschmar, T.; Lee, Y.; Martinoia, E. Plant ABC Transporters. Arab. Book 2011, 9, e0153. [Google Scholar] [CrossRef]
- Xiao, C.; Qiu, S.; Robertson, R.M. The white gene controls copulation success in Drosophila melanogaster. Sci. Rep. 2017, 7, 17712. [Google Scholar] [CrossRef]
- Qian, S.; Pirrotta, V. Dosage compensation of the Drosophila white gene requires both the X chromosome environment and multiple intragenic elements. Genetics 1995, 139, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Slatis, H.M. Position Effects at the Brown Locus in Drosophila Melanogaster. Genetics 1955, 40, 5–23. [Google Scholar] [CrossRef]
- Grubbs, N.; Haas, S.; Beeman, R.W.; Lorenzen, M.D. The ABCs of Eye Color in Tribolium castaneum: Orthologs of the Drosophila white, scarlet, and brown Genes. Genetics 2015, 199, 749–759. [Google Scholar] [CrossRef]
- Babazadeh, R.; Schneider, K.L.; Fischbach, A.; Hao, X.; Liu, B.; Nystrom, T. The yeast guanine nucleotide exchange factor Sec7 is a bottleneck in spatial protein quality control and detoxifies neurological disease proteins. Sci. Rep. 2023, 13, 14068. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.R.; Wittinghofer, A. The Guanine Nucleotide-Binding Switch in Three Dimensions. Science 2001, 294, 1299–1304. [Google Scholar] [CrossRef]
- Mandel, M.; Marmur, J. [109] Use of ultraviolet absorbance-temperature profile for determining the guanine plus cytosine content of DNA. Methods Enzymol. 1968, 12, 195–206. [Google Scholar] [CrossRef]
- Maeda, T.; Chapman, D.L.; Stewart, A.F.R. Mammalian Vestigial-like 2, a Cofactor of TEF-1 and MEF2 Transcription Factors That Promotes Skeletal Muscle Differentiation. J. Biol. Chem. 2002, 277, 48889–48898. [Google Scholar] [CrossRef]
- Vaudin, P.; Delanoue, R.; Davidson, I.; Silber, J.; Zider, A. TONDU (TDU), a novel human protein related to the product of vestigial (vg) gene of Drosophila melanogaster interacts with vertebrate TEF factors and substitutes for Vg function in wing formation. Development 1999, 126, 4807–4816. [Google Scholar] [CrossRef]
- Halmenschelager, P.T.; da Rocha, J.B.T. Biochemical CuSO4 Toxicity in Drosophila melanogaster depends on sex and developmental stage of exposure. Biol. Trace Elem. Res. 2019, 189, 574–585. [Google Scholar] [CrossRef]
- Lints, F.; Lints, C.; Bullens, P.; Bourgois, M.; Delincé, J. Unexplained variations in life span of the Oregon-R strain of Drosophila melanogaster over a four-year period. Exp. Gerontol. 1989, 24, 265–271. [Google Scholar] [CrossRef]
- Siddique, Y.H.; Haidari, M.; Khan, W.; Fatima, A.; Jyoti, S.; Khanam, S.; Naz, F.; Rahul; Ali, F.; Singh, B.R.; et al. Toxic potential of copper-doped ZnO nanoparticles in Drosophila melanogaster (Oregon R). Toxicol. Mech. Methods 2015, 25, 425–432. [Google Scholar] [CrossRef]
- Lefevre, G.; Green, M.M. Genetic duplication in the white-split interval of the X chromosome in Drosophila melanogaster. Chromosoma 1972, 36, 391–412. [Google Scholar] [CrossRef]
- Green, M.M. 2010: A Century of Drosophila Genetics Through the Prism of the white Gene. Genetics 2010, 184, 3–7. [Google Scholar] [CrossRef]
- Rabinow, L.; Nguyen-Huynh, A.T.; Birchler, J.A. A trans-acting regulatory gene that inversely affects the expression of the white, brown and scarlet loci in Drosophila. Genetics 1991, 129, 463–480. [Google Scholar] [CrossRef]
- Campbell, J.L.; Nash, H.A. Volatile general anesthetics reveal a neurobiological role for the white and brown genes of Drosophila melanogaster. J. Neurobiol. 2001, 49, 339–349. [Google Scholar] [CrossRef]
- Aarts, J.M.M.J.G.; Alink, G.M.; Franssen, H.J.; Roebroeks, W. Evolution of Hominin Detoxification: Neanderthal and Modern Human Ah Receptor Respond Similarly to TCDD. Mol. Biol. Evol. 2020, 38, 1292–1305. [Google Scholar] [CrossRef]
- Gachon, F.; Olela, F.F.; Schaad, O.; Descombes, P.; Schibler, U. The circadian PAR-domain basic leucine zipper transcription factors DBP, TEF, and HLF modulate basal and inducible xenobiotic detoxification. Cell Metab. 2006, 4, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, A.K.; Pandey, P. Fecundity of inbred fruit fly Drosophila melanogaster on different solid culture media: An analysis. J. Appl. Nat. Sci. 2018, 10, 1109–1114. [Google Scholar] [CrossRef]
- Klimaczewski, C.V.; Ecker, A.; Piccoli, B.; Aschner, M.; Barbosa, N.V.; Rocha, J.B.T. Peumus boldus attenuates copper-induced toxicity in Drosophila melanogaster. Biomed. Pharmacother. 2018, 97, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Va, D.P.; Sa, A.A.; Paul, S.F. wonder animal model for genetic studies-Drosophila melanogaster-its life cycle and breeding methods-a review. Sri Ramachandra J. Med. 2009, 2, 33–38. [Google Scholar]
- Wang, Y.; Misto, M.; Yang, J.; Gehring, N.; Yu, X.; Moussian, B. Toxicity of Dithiothreitol (DTT) to Drosophila melanogaster. Toxicol. Rep. 2021, 8, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Burke, R. Molecular physiology of copper in Drosophila melanogaster. Curr. Opin. Insect Sci. 2022, 51, 100892. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.A.; Schneuwly, S. Copper and Zinc Homeostasis: Lessons from Drosophila melanogaster. Front. Genet. 2017, 8, 223. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Men, L.; Wang, J.; Zhang, Y.; Chickenyen, S.; Wang, Y.; Zhou, F. Redox Reactions of Copper Complexes Formed with Different β-Amyloid Peptides and Their Neuropathalogical Relevance. Biochemistry 2007, 46, 9270–9282. [Google Scholar] [CrossRef] [PubMed]
- Southon, A.; Burke, R.; Norgate, M.; Batterham, P.; Camakaris, J. Copper homoeostasis in Drosophila melanogaster S2 cells. Biochem. J. 2004, 383, 303–309. [Google Scholar] [CrossRef]
- Avram, O.R.; Caragea, G.; Varzaru, C.A. Copper and its role in the human body-the importance of establishing copper concentrations in the body. Rom. J. 2021, 124, 2. [Google Scholar]
- Demir, E.; Demir, F.T. Drosophila: A promising model for evaluating the toxicity of environmental pollutants. Karaelmas Mühendislik Derg. 2022, 12, 101–118. [Google Scholar]
- An, Y.-J. Assessment of comparative toxicities of lead and copper using plant assay. Chemosphere 2006, 62, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Komarnicka, U.K.; Lesiów, M.K.; Witwicki, M.; Bieńko, A. The Bright and Dark Sides of Reactive Oxygen Species Generated by Copper–Peptide Complexes. Separations 2022, 9, 73. [Google Scholar] [CrossRef]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Guan, X.; Zhou, L.; Asad, M.A.U.; Xu, Y.; Pan, G.; Cheng, F. ABA-triggered ROS burst in rice developing anthers is critical for tapetal programmed cell death induction and heat stress-induced pollen abortion. Plant Cell Environ. 2023, 46, 1453–1471. [Google Scholar] [CrossRef] [PubMed]
- Emilian, O.; Ioan, S.; Irina, P.; Raul, P.; Adriana, C.; Dorin, C.; Ciprian, S. Cytological Applications of the Vacuolization Phenomenon as a Means of Determining Saline Cytotoxicity. Appl. Sci. 2023, 13, 8461. [Google Scholar] [CrossRef]
- Ho, T.; Ahmadi, S.; Kerman, K. Do glutathione and copper interact to modify Alzheimer’s disease pathogenesis? Free. Radic. Biol. Med. 2022, 181, 180–196. [Google Scholar] [CrossRef] [PubMed]
- Babak, M.V.; Ahn, D. Modulation of intracellular copper levels as the mechanism of action of anticancer copper complexes: Clinical relevance. Biomedicines 2021, 9, 852. [Google Scholar] [CrossRef]
- Devóz, P.P.; Dos Reis, M.B.; Gomes, W.R.; Maraslis, F.T.; Ribeiro, D.L.; Antunes, L.M.G.; Batista, B.L.; Grotto, D.; Reis, R.M.; Barbosa, F., Jr. Adaptive epigenetic response of glutathione (GSH)-related genes against lead (Pb)-induced toxicity, in individuals chronically exposed to the metal. Chemosphere 2021, 269, 128758. [Google Scholar] [CrossRef]

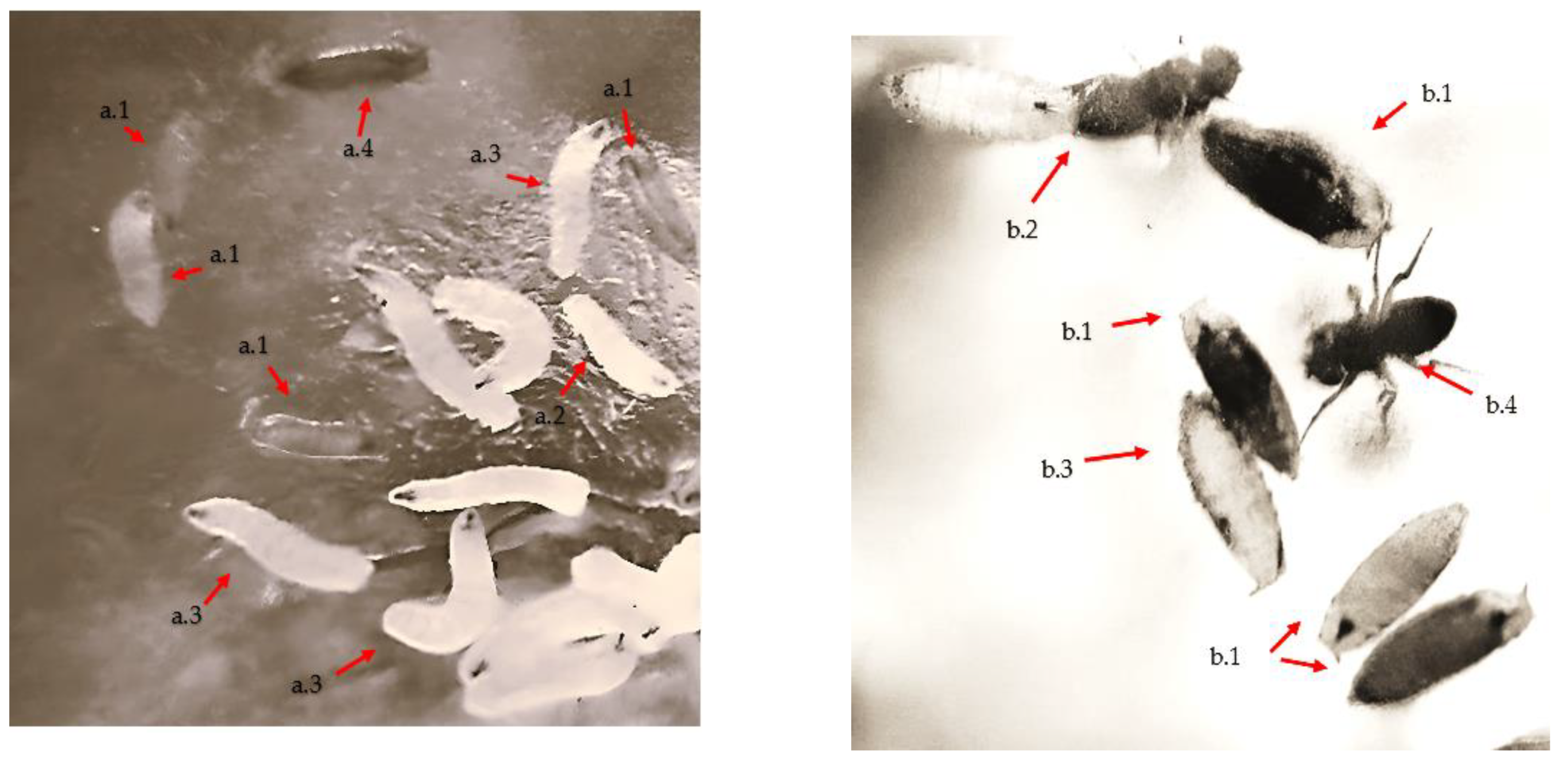


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | SS | df | S2 | F | Sig. |
|---|---|---|---|---|---|
| Genotype | 10,981,505 | 3 | 3,660,502 | 79,502 | *** |
| Concentrations | 13,289,452 | 4 | 3,322,363 | 72,158 | *** |
| Metal (Cu2+ vs. Pb2+) | 2408,925 | 1 | 2,408,925 | 52,319 | *** |
| Genotype * Concentrations | 190,491 | 12 | 15,874 | 0.345 | Ns |
| Genotype * Metal | 219,850 | 3 | 73,283 | 1.592 | Ns |
| Concentrations * Metal | 749,858 | 4 | 187,464 | 4.071 | ** |
| Genotype * Concentrations * Metal | 193,764 | 12 | 16,147 | 0.351 | Ns |
| Error | 25,784,145 | 560 | 46,043 | ||
| Total | 339,463,000 | 600 |
| Source | SS | Df | S2 | F | Sig. |
|---|---|---|---|---|---|
| Genotype | 9,901,500 | 3 | 3,300,500 | 82,113 | *** |
| Concentrations | 11,989,861 | 4 | 2,997,465 | 74,573 | *** |
| Metal (Cu2+ vs. Pb2+) | 1,989,970 | 1 | 1,989,970 | 49,508 | *** |
| Genotype * Concentrations | 316,201 | 12 | 26,350 | 0.656 | Ns |
| Genotype * Metal | 377,536 | 3 | 125,845 | 3.131 | * |
| Concentrations * Metal | 598,479 | 4 | 149,620 | 3.722 | ** |
| Genotype * Concentrations * Metal | 189,923 | 12 | 15,827 | 0.394 | Ns |
| Error | 22,509,100 | 560 | 40,195 | ||
| Total | 276,564,000 | 600 |
| Source | SS | Df | S2 | F | Sig. |
|---|---|---|---|---|---|
| Genotype | 10,508,245 | 3 | 3,502,748 | 106,420 | *** |
| Concentrations | 11,244,424 | 4 | 2,811,106 | 85,407 | *** |
| Metal (Cu2+ vs. Pb2+) | 1,825,496 | 1 | 1,825,496 | 55,462 | *** |
| Genotype * Concentrations | 319,035 | 12 | 26,586 | 0.808 | Ns |
| Genotype * Metal | 316,404 | 3 | 105,468 | 3.204 | * |
| Concentrations * Metal | 623,184 | 4 | 155,796 | 4.733 | *** |
| Genotype * Concentrations * Metal | 340,952 | 12 | 28,413 | 0.863 | Ns |
| Error | 18,432,016 | 560 | 32,914 | ||
| Total | 231,562,000 | 600 |
| Genotype | Concentration mM | Larvae | IC | Pupal | IC | Adult | IC |
|---|---|---|---|---|---|---|---|
| Mean | Mean | Mean | |||||
| Oregon R | Control | 36.13 a | 34.07 a | 31.73 a | |||
| 0.5 | 28.60 ab | 20.85 | 25.73 b | 24.46 | 22.53 bc | 28.99 | |
| 0.75 | 25.20 bcd | 30.26 | 21.67 bcde | 36.40 | 20.27 bcd | 36.13 | |
| 1 | 22.07 bcde | 38.93 | 19.07 bcde | 44.03 | 18.53 bcde | 41.60 | |
| 2 | 17.2 defg | 52.40 | 15.80 defg | 53.62 | 14.13 defgh | 55.46 | |
| brown | Control | 27.27 bc | 24.40 efg | 20.33 bcd | |||
| 0.5 | 20.80 bcdef | 23.72 | 18.07 fgh | 25.96 | 17.07 cdefg | 16.07 | |
| 0.75 | 17.47 defg | 35.94 | 16.47 defg | 32.51 | 14.40 defgh | 29.18 | |
| 1 | 15.73 efg | 42.30 | 14.33 efg | 41.26 | 10.60 fgh | 47.87 | |
| 2 | 11.93 gh | 56.23 | 10.80 fgh | 55.74 | 8.20 ghi | 59.67 | |
| white | Control | 28.33 ab | 24.80 bc | 24,27 b | |||
| 0.5 | 21.73 bcdef | 23.29 | 18.13 bcdef | 26.88 | 19.40 bcd | 20.05 | |
| 0.75 | 18.80 cdefg | 33.65 | 17.33 cdef | 30.11 | 16.40 cdef | 32.42 | |
| 1 | 17.40 defg | 38.59 | 15.67 defg | 36.83 | 15.27 def | 37.09 | |
| 2 | 10.20 gh | 64.00 | 8.87 gh | 64.25 | 7.67 hi | 68.41 | |
| white-vestigial | Control | 23.93 bcde | 23.20 bcd | 20,47 bcd | |||
| 0.5 | 16.92 defg | 29.29 | 15.54 defg | 33.02 | 14.92 defg | 27.09 | |
| 0.75 | 15.93 efg | 29.25 | 14.60 efg | 37.07 | 12.47 efgh | 39.09 | |
| 1 | 13.33 fgh | 44.29 | 10.53 fgh | 54.60 | 8.33 ghi | 59.28 | |
| 2 | 7.12 h | 70.26 | 5.76 h | 75.15 | 2.88 i | 85.92 |
| Genotype | Concentration | Larvae | IC | Pupal | IC | Adult | IC |
|---|---|---|---|---|---|---|---|
| Mean | Mean | Mean | |||||
| Oregon R | Control | 36.13 a | 34.07 a | 31.73 a | |||
| 0.5 | 34.13 ab | 5.54 | 32.47 ab | 4.70 | 30.87 ab | 2.73 | |
| 0.75 | 32.07 abc | 11.25 | 28.07 abc | 17.61 | 26.53 abc | 16.39 | |
| 1 | 29.20 abcd | 19.19 | 27.27 abcd | 19.96 | 25.13 abcd | 20.80 | |
| 2 | 25.20 cdefg | 30.26 | 23.80 cdefg | 30.14 | 22.27 bcdefg | 29.83 | |
| brown | Control | 27.27 bcdef | 24.40 cdefg | 20.33 bcdef | |||
| 0.5 | 23.73 bcdefg | 12.96 | 19.67 defgh | 19.40 | 17.13 cdefgh | 15.74 | |
| 0.75 | 23.53 efghij | 13.69 | 17.67 fghi | 27.60 | 16.07 bcdefg | 20.98 | |
| 1 | 19.93 efgh | 26.89 | 16.33 ghi | 33.06 | 14.60 efghi | 28.20 | |
| 2 | 18.80 ij | 31.05 | 14.40 hi | 40.98 | 12.07 fghi | 40.66 | |
| white | Control | 28.33 bcde | 24.80 abcde | 24.27 abcde | |||
| 0.5 | 27.40 bcdefg | 3.29 | 23.87 bcdef | 3.76 | 22.93 abcdef | 5.49 | |
| 0.75 | 25.33 cdefgh | 10.59 | 23.07 cdefgh | 6.99 | 19.33 bcdefg | 20.33 | |
| 1 | 19.67 efghij | 30.59 | 18.47 efgh | 25.54 | 16.80 ghi | 30.77 | |
| 2 | 17.67 fghij | 37.65 | 16.13 ghi | 34.95 | 14.87 cdefgh | 38.74 | |
| white-vestigial | Control | 23.93 cdefgh | 23.20 cdefg | 20.47 cdefgh | |||
| 0.5 | 19.85 defghij | 17.08 | 18.62 fghi | 19.76 | 15.38 defghi | 24.83 | |
| 0.75 | 17.47 ghij | 27.02 | 15.73 ghi | 32.18 | 16.00 ghi | 21.82 | |
| 1 | 15.80 hij | 33.98 | 14.73 ghi | 36.49 | 14.33 hi | 29.97 | |
| 2 | 11.88 j | 50.35 | 11.00 i | 52.59 | 8.59 i | 58.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ioan, S.; Irina, P.; Emilian, O.; Sorina, P.; Cerasela, P.; Adriana, C.; Dorin, C.; Alina-Maria, T.-C.; Dacian, L.; Ciprian, S.; et al. Application of the Drosophila melanogaster Research Model to Evaluate the Toxicity Levels between Lead and Copper. Appl. Sci. 2024, 14, 4190. https://doi.org/10.3390/app14104190
Ioan S, Irina P, Emilian O, Sorina P, Cerasela P, Adriana C, Dorin C, Alina-Maria T-C, Dacian L, Ciprian S, et al. Application of the Drosophila melanogaster Research Model to Evaluate the Toxicity Levels between Lead and Copper. Applied Sciences. 2024; 14(10):4190. https://doi.org/10.3390/app14104190
Chicago/Turabian StyleIoan, Sarac, Petrescu Irina, Onisan Emilian, Popescu Sorina, Petolescu Cerasela, Ciulca Adriana, Camen Dorin, Tenche-Constantinescu Alina-Maria, Lalescu Dacian, Stroia Ciprian, and et al. 2024. "Application of the Drosophila melanogaster Research Model to Evaluate the Toxicity Levels between Lead and Copper" Applied Sciences 14, no. 10: 4190. https://doi.org/10.3390/app14104190
APA StyleIoan, S., Irina, P., Emilian, O., Sorina, P., Cerasela, P., Adriana, C., Dorin, C., Alina-Maria, T.-C., Dacian, L., Ciprian, S., Anamaria Aurelia, M., Laura-Gratiela, V., & Mariana, G. (2024). Application of the Drosophila melanogaster Research Model to Evaluate the Toxicity Levels between Lead and Copper. Applied Sciences, 14(10), 4190. https://doi.org/10.3390/app14104190