
Protective Activity against Oxidative Stress in Dermal Papillae with Extracted Herbal Essential Oils

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Two-Dimensional (2D) and Three-Dimensional (3D) DPC Cultures
2.1.1. 2D-DPC Cultures
2.1.2. 3D-DPC Cultures
2.2. Preparation of Essential Oils Derived from Herbs
2.3. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium Bromide (MTT) Assay
2.4. Measurement of Ultraviolet B (UVB)-Induced Intracellular ROS Levels
2.5. Confocal Microscopy for Detecting NRF2 Expression in DPCs
2.6. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) for NRF2, Phase II Enzymes Genes, and the Hair Growth Factors Genes VEGF and IGF1 in DPCs
2.7. Real-Time Quantitative RT-PCR (qRT-PCR) for NRF2 and Phase II Enzyme Genes in 3D-DPC Cultures
2.8. ELISA Assay to Determine VEGF and IGF-1 Protein Levels in 3D-DPC Cultures
2.9. Statistical Analyses
3. Results
3.1. Extraction of Herb-Derived Essential Oils
3.2. The ROS-Scavenging Activity of Herb-Derived Essential Oils in UVB-Irradiated DPCs. DCF-DA Signals Are Reduced to Confirm Anti-Oxidant Activity
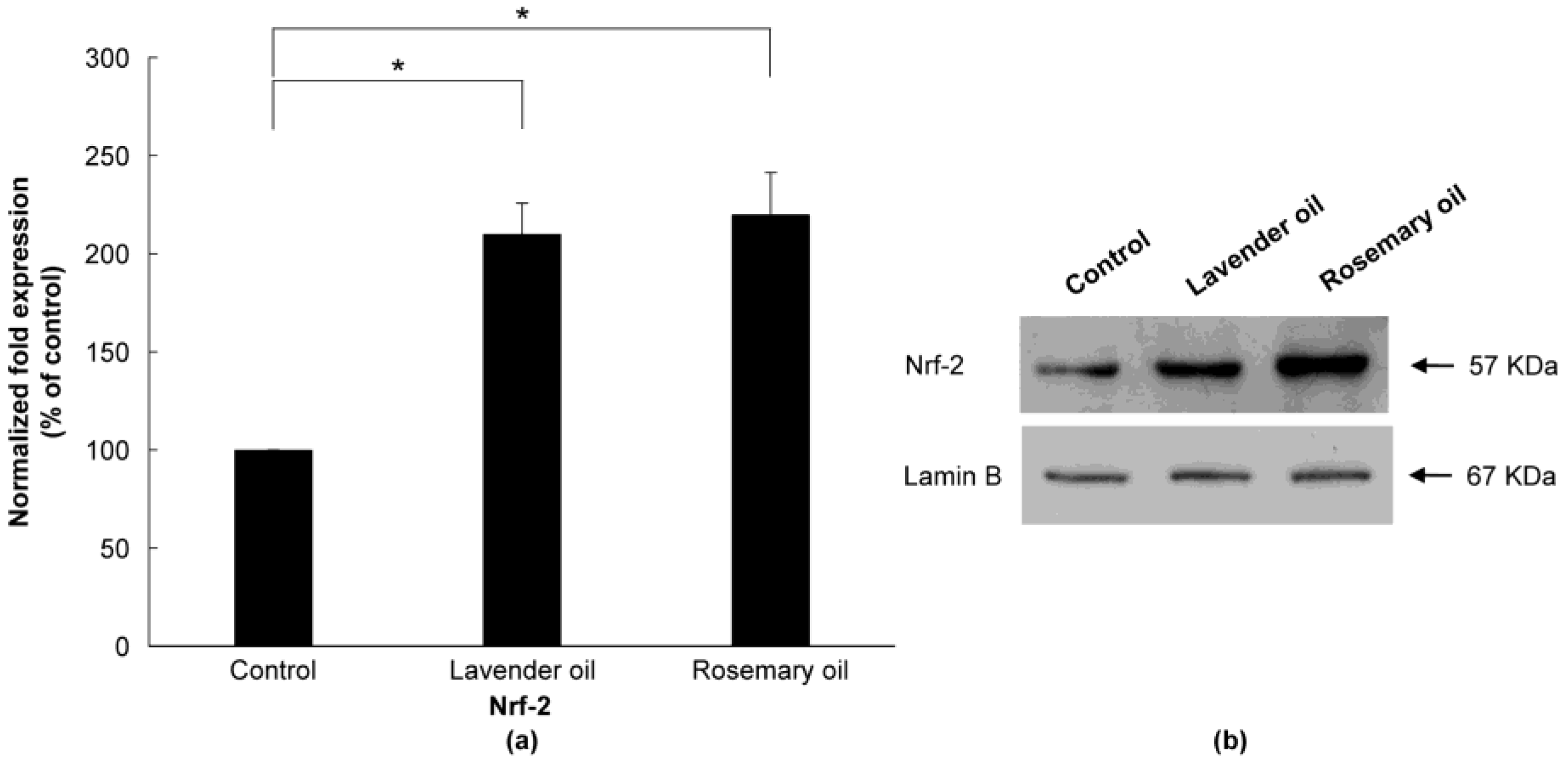
3.3. Upregulation of NRF2 by Herb-Derived Essential Oils in DPCs
3.4. Upregulation of Phase II Enzymes by Herb-Derived Essential Oils in DPCs
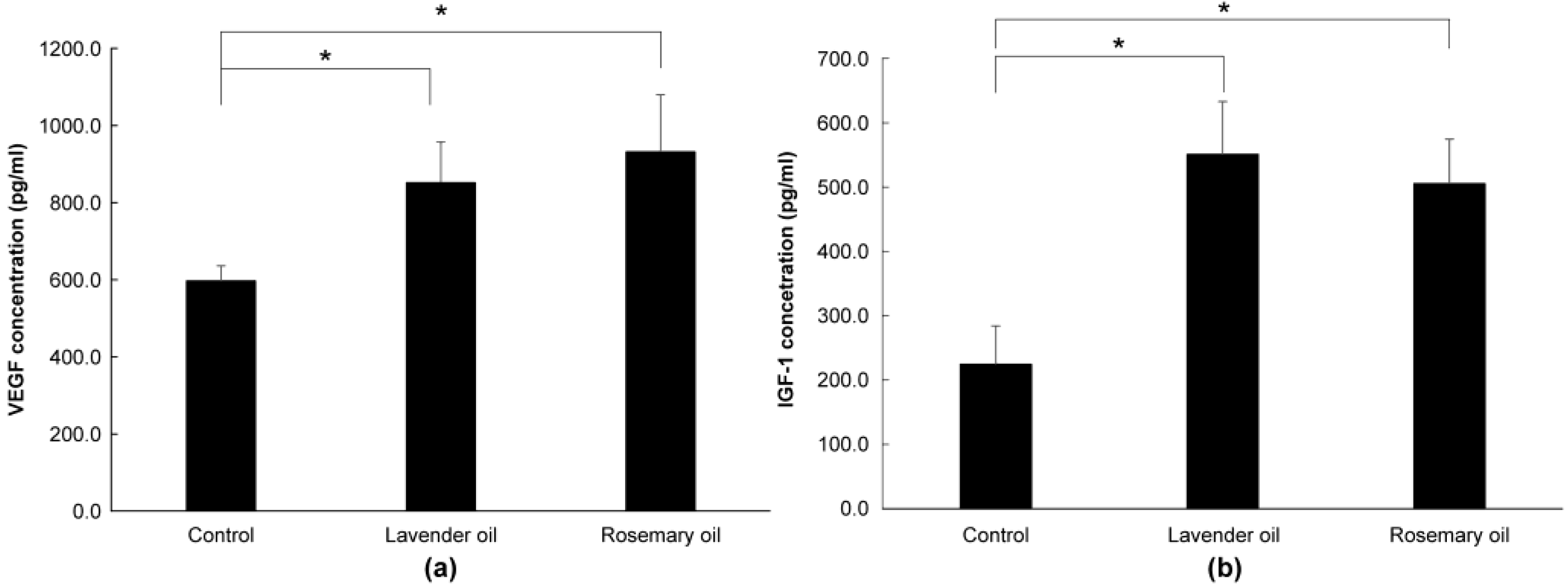
3.5. Upregulation of VEGF and IGF1, Hair Growth Factors by Herb-Derived Essential Oils in DPC Cultures
3.6. Upregulation of NRF2, Phase II Enzymes, and Hair Growth Factors by Herb-Derived Essential Oils in 3D-DPC Culture
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, C.C.; Cotsarelis, G. Review of hair follicle dermal cells. J. Dermatol. Sci. 2010, 57, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Madaan, A.; Verma, R.; Singh, A.T.; Jaggi, M. Review of hair follicle dermal papilla cells as in vitro screening model for hair growth. Int. J. Cosmet. Sci. 2018, 40, 429–450. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.R.; Schmidt-Ullrich, R.; Paus, R. The hair follicle as a dynamic miniorgan. Curr. Biol. 2009, 19, R132–R142. [Google Scholar] [CrossRef]
- Jahoda, C.A.B.; Reynolds, A.J. Dermal-epidermal interactions-follicle-derived cell populations in the study of hair-growth mechanisms. J. Investig. Dermatol. 1993, 101, S33–S38. [Google Scholar] [CrossRef]
- Harris, J.E. Cellular stress and innate inflammation in organ-specific autoimmunity: Lessons learned from vitiligo. Immunol. Rev. 2016, 269, 11–25. [Google Scholar] [CrossRef]
- Bakry, O.A.; Elshazly, R.M.A.; Shoeib, M.A.M.; Gooda, A. Oxidative stress in alopecia areata: A case-control study. Am. J. Clin. Dermatol. 2014, 15, 57–64. [Google Scholar] [CrossRef]
- Xing, L.; Dai, Z.; Jabbari, A.; Cerise, J.E.; Higgins, C.A.; Gong, W.; de Jong, A.; Harel, S.; DeStefano, G.M.; Rothman, L.; et al. Alopecia areata is driven by cytotoxic T lymphocytes and is reversed by JAK inhibition. Nat. Med. 2014, 20, 1043–1049. [Google Scholar] [CrossRef]
- Prie, B.E.; Voiculescu, V.M.; Ionescu-Bozdog, O.B.; Petrutescu, B.; Iosif, L.; Gaman, L.E.; Clatici, V.G.; Stoian, I.; Giurcaneanu, C. Oxidative stress and alopecia areata. J. Med. Life 2015, 8, 43–46. [Google Scholar]
- Yenin, J.Z.; Serarslan, G.; Yönden, Z.; Ulutaş, K.T. Investigation of oxidative stress in patients with alopecia areata and its relationship with disease severity, duration, recurrence and pattern. Clin. Exp. Dermatol. 2015, 40, 617–621. [Google Scholar] [CrossRef]
- Naziroglu, M.; Kokcam, I. Antioxidants and lipid peroxidation status in the blood of patients with alopecia. Cell Biochem. Funct. 2000, 18, 169–173. [Google Scholar] [CrossRef]
- Akar, A.; Arca, E.; Erbil, H.; Akay, C.; Sayal, A.; Gür, A.R. Antioxidant enzymes and lipid peroxidation in the scalp of patients with alopecia areata. J. Dermatol. Sci. 2002, 29, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Koca, R.; Armutcu, F.; Altinyazar, C.; Gürel, A. Evaluation of lipid peroxidation, oxidant/antioxidant status, and serum nitric oxide levels in alopecia areata. Med. Sci. Monit. 2005, 11, CR296–CR299. [Google Scholar] [PubMed]
- Abdel Fattah, N.S.A.; Ebrahim, A.A.; El Okda, E.S. Lipid peroxidation/antioxidant activity in patients with alopecia areata. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Picardo, M. Antioxidant activity, lipid peroxidation and skin diseases. What’s new. J. Eur. Acad. Dermatol. Venereol. 2003, 17, 663–669. [Google Scholar] [CrossRef]
- Ramadan, R.; Tawdy, A.; Abdel Hay, R.; Rashed, L.; Tawfik, D. The antioxidant role of paraoxonase 1 and vitamin E in three autoimmune diseases. Skin Pharmacol. Physiol. 2013, 26, 2–7. [Google Scholar] [CrossRef]
- Upton, J.H.; Hannen, R.F.; Bahta, A.W.; Farjo, N.; Farjo, B.; Philpott, M.P. Oxidative stress–associated senescence in dermal papilla cells of men with androgenetic alopecia. J. Investig. Dermatol. 2015, 135, 1244–1252. [Google Scholar] [CrossRef]
- Schäfer, M.; Werner, S. Nrf2–A regulator of keratinocyte redox signaling. Free Radic. Biol. Med. 2015, 88, 243–252. [Google Scholar] [CrossRef]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Fujie, T.; Katoh, S.; Oura, H.; Urano, Y.; Arase, S. The chemotactic effect of a dermal papilla cell–derived factor on outer root sheath cells. J. Dermatol. Sci. 2001, 25, 206–212. [Google Scholar] [CrossRef]
- Yano, K.; Brown, L.F.; Detmar, M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Investig. 2001, 107, 409–417. [Google Scholar] [CrossRef]
- Trüeb, R.M. Further clinical evidence for the effect of IGF-1 on hair growth and alopecia. Skin Appendage Disord. 2018, 4, 90–95. [Google Scholar] [CrossRef] [PubMed]
- McDonagh, A.J.G.; Elliott, K.R.; Messenger, A.G. Cytokines and dermal papilla function in alopecia areata. J. Investig. Dermatol. 1995, 104, S9–S10. [Google Scholar] [CrossRef] [PubMed]
- Jahoda, C.A.B.; Reynolds, A.J. Dermal-epidermal interactions: Adult follicle-derived cell populations and hair growth. Dermatol. Clin. 1996, 14, 573–583. [Google Scholar] [CrossRef]
- Millar, S.E. Molecular mechanisms regulating hair follicle development. J. Investig. Dermatol. 2002, 118, 216–225. [Google Scholar] [CrossRef]
- Fischer, T.W.; Herczeg-Lisztes, E.; Funk, W.; Zillikens, D.; Bíró, T.; Paus, R. Differential effects of caffeine on hair shaft elongation, matrix and outer root sheath keratinocyte proliferation, and transforming growth factor-β2/insulin-like growth factor-1-mediated regulation of the hair cycle in male and female human hair follicles in vitro. Br. J. Dermatol. 2014, 171, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Bosch, R.; Philips, N.; Suárez-Pérez, J.A.; Juarranz, A.; Devmurari, A.; Chalensouk-Khaosaat, J.; González, S. Mechanisms of photoaging and cutaneous photocarcinogenesis, and photoprotective strategies with phytochemicals. Antioxidants 2015, 4, 248–268. [Google Scholar] [CrossRef] [PubMed]
- Jadkauskaite, L.; Coulombe, P.A.; Schäfer, M.; Dinkova-Kostova, A.T.; Paus, R.; Haslam, I.S. Oxidative stress management in the hair follicle: Could targeting NRF2 counter age-related hair disorders and beyond. BioEssays 2017, 39, 1700029. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, J.; Fu, D.; Liu, Z.; Wang, H.; Wang, J.; Qu, Q.; Li, K.; Fan, Z.; Hu, Z.; et al. Transcriptome Analysis Reveals an Inhibitory Effect of Dihydrotestosterone-Treated 2D- and 3D-Cultured Dermal Papilla Cells on Hair Follicle Growth. Front. Cell Dev. Biol. 2021, 17, 724310. [Google Scholar] [CrossRef]
- Piao, M.S.; Choi, J.Y.; Lee, D.H.; Yun, S.J.; Lee, J.B.; Lee, S.C. Differentiation-dependent expression of NADP(H): Quinone oxidoreductase-1 via NF-E2 related factor-2 activation in human epidermal keratinocytes. J. Dermatol. Sci. 2011, 62, 147–153. [Google Scholar] [CrossRef]
- Mu, Y.; He, J.; Yan, R.; Hu, X.; Liu, H.; Hao, Z. IGF-1 and VEGF can be used as prognostic indicators for patients with uterine fibroids treated with uterine artery embolization. Exp. Ther. Med. 2016, 11, 645–649. [Google Scholar] [CrossRef]
- El-Sayed, S.M.; Hassan, K.M.; Abdelhamid, A.N.; Yousef, E.E.; Abdellatif, Y.M.R.; Abu-Hussien, S.H.; Naseerm, M.A.; Elshalakany, W.A.; Eldin Darwish, D.B.; Abdulmajeed, A.M.; et al. Exogenous Paclobutrazol Reinforces the Antioxidant and Antimicrobial Properties of Lavender (Lavandula officinalis L.) Oil through Modulating Its Composition of Oxygenated Terpenes. Plants 2022, 19, 1607. [Google Scholar] [CrossRef]
- Aziz, Z.A.A.; Ahmad, A.; Setapar, S.H.M.; Karakucuk, A.; Azim, M.M.; Lokhat, D.; Rafatullah, M.; Ganash, M.; Kamal, M.A.; Ashraf, G.M. Essential oils: Extraction techniques, pharmaceutical and therapeutic potential—A review. Curr. Drug Metab. 2018, 19, 1100–1110. [Google Scholar] [CrossRef] [PubMed]
- Itami, S.; Kurata, S.; Takayasu, S. Androgen induction of follicular epithelial cell growth is mediated via insulin-like growth factor-I from dermal papilla cells. Biochem. Biophys. Res. Commun. 1995, 212, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Trüeb, R.M. Oxidative stress in ageing of hair. Int. J. Trichology 2009, 1, 6–14. [Google Scholar] [CrossRef]
- Bae, S.; Lim, K.M.; Cha, H.J.; An, I.S.; Lee, J.P.; Lee, K.S.; Lee, G.T.; Lee, K.K.; Jung, H.J.; Ahn, K.J.; et al. Arctiin blocks hydrogen peroxide-induced senescence and cell death though microRNA expression changes in human dermal papilla cells. Biol. Res. 2014, 47, 50. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.J.; Jahoda, C.A. Hair matrix germinative epidermal cells confer follicle-inducing capabilities on dermal sheath and high passage papilla cells. Development 1996, 122, 3085–3094. [Google Scholar] [CrossRef] [PubMed]
- Uliasz, A.; Spencer, J.M. Chemoprevention of skin cancer and photoaging. Clin. Dermatol. 2004, 22, 178–182. [Google Scholar] [CrossRef]
- Choi, J.; Yun, S.; Lee, J.; Piao, M.; Park, E.; Lee, S. A set of plant essential oils have antioxidant activity in ultraviolet B-irradiated epidermal keratinocytes by upregulating antioxidant and detoxifying enzymes. J. Investig. Dermatol. 2017, 5, 233–242. [Google Scholar] [CrossRef]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation—A review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef]
- Furue, M.; Fuyuno, Y.; Mitoma, C.; Uchi, H.; Tsuji, G. Therapeutic agents with AHR inhibiting and NRF2 activating activity for managing chloracne. Antioxidants 2018, 7, 90. [Google Scholar] [CrossRef]
- Scapagnini, G.; Davinelli, S.; Renzo, L.D.; Lorenzo, A.D.; Olarte, H.H.; Micali, G.; Cicero, A.F.; Gonzalez, S. Cocoa bioactive compounds: Significance and potential for the maintenance of skin health. Nutrients 2014, 6, 3202–3213. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Wang, X.; Zhao, S.; Ma, C.; Cui, J.; Zheng, Y. Sulforaphane Protects against Cardiovascular Disease via Nrf2 Activation. Oxid. Med. Cell. Longev. 2015, 2015, 407580. [Google Scholar] [CrossRef] [PubMed]
- Limat, A.; Hunziker, T.; Waelti, E.R.; Inaebnit, S.P.; Wiesmann, U.; Braathen, L.R. Soluble factors from human hair papilla cells and dermal fibroblasts dramatically increase the clonal growth of outer root sheath cells. Arch. Dermatol. Res. 1993, 285, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, H.F.; Brown, L.F.; Detmar, M.; Dvorak, A.M. Vascular permeability factor/vascular endothelial growth factor, microvascular hyperpermeability, and angiogenesis. Am. J. Pathol. 1995, 146, 1029–1039. [Google Scholar] [PubMed]
- Brown, L.F.; Yeo, K.T.; Berse, B.; Yeo, T.K.; Senger, D.R.; Dvorak, H.F.; van de Water, L. Expression of vascular permeability factor (vascular endothelial growth factor) by epidermal keratinocytes during wound healing. J. Exp. Med. 1992, 176, 1375–1379. [Google Scholar] [CrossRef]
- Detmar, M.; Brown, L.F.; Claffey, K.P.; Yeo, K.T.; Kocher, O.; Jackman, R.W.; Berse, B.; Dvorak, H.F. Overexpression of vascular permeability factor/vascular endothelial growth factor and its receptors in psoriasis. J. Exp. Med. 1994, 180, 1141–1146. [Google Scholar] [CrossRef]
- Lachgar, S.; Moukadiri, H.; Jonca, F.; Charveron, M.; Bouhaddioui, N.; Gall, Y.; Bonafe, J.L.; Plouët, J. Vascular endothelial growth factor is an autocrine growth factor for hair dermal papilla cells. J. Investig. Dermatol. 1996, 106, 17–23. [Google Scholar] [CrossRef]
- Meephansan, J.; Thummakriengkrai, J.; Ponnikorn, S.; Yingmema, W.; Deenonpoe, R.; Suchonwanit, P. Efficacy of topical tofacitinib in promoting hair growth in non-scarring alopecia: Possible mechanism via VEGF induction. Arch. Dermatol. Res. 2017, 309, 729–738. [Google Scholar] [CrossRef]
- Lachgar, S.; Charveron, M.; Gall, Y.; Bonafe, J.L. Minoxidil upregulates the expression of vascular endothelial growth factor in human hair dermal papilla cells. Br. J. Dermatol. 1998, 138, 407–411. [Google Scholar] [CrossRef]
- Rho, S.S.; Park, S.J.; Hwang, S.L.; Lee, M.H.; Kim, C.D.; Lee, I.H.; Chang, S.Y.; Rang, M.J. The hair growth promoting effect of Asiasari radix extract and its molecular regulation. J. Dermatol. Sci. 2005, 38, 89–97. [Google Scholar] [CrossRef]
- Junlatat, J.; Sripanidkulchai, B. Hair growth-promoting effect of Carthamus tinctorius floret extract. Phytother. Res. 2014, 28, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.S.; Hong, E.J.; Gwak, K.S.; Park, M.J.; Choi, K.C.; Choi, I.G.; Jang, J.W.; Jeung, E.B. The essential oils of Chamaecyparis obtusa promote hair growth through the induction of vascular endothelial growth factor gene. Fitoterapia 2010, 81, 17–24. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (Forward/Reverse) | PCR Product Size (bp) |
|---|---|---|
| NRF2 | 5′-aaaccagtggatctgccaac-3′/ 5′-gaccgggaatatcaggaaca-3 | 164 |
| HO-1 | 5′-caggcagagaatgctgagttc-3′/ 5′-gatgttgagcaggaacgcagt-3 | 554 |
| NQO-1 | 5′-cagcgccccggactgcaccagagcc-3′/ 5′-gggaagcctggaaagatacccaga-3′ | 142 |
| GSTpi | 5′-cagcgccccggactgcaccagagcc-3′/ 5′-gggaagcctggaaagatacccaga-3 | 428 |
| VEGF | 5′-tcttgggtgcattggagcctc-3′/ 5′-aagctcatctctcctatgtgc-3’ | 350 |
| IGF1 | 5′-tcaacaagcccacagggtat-3′/ 5′-actcgtgcagagcaaaggat-3′ | 307 |
| GAPDH | 5′-gtcttcaccaccatggagaaggc-3/ 5′-cggaaggccatgccagtgagctt-3′ | 400 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, D.-I.; Choi, J.-Y.; Lee, J.-B.; Yun, S.-J.; Moon, B.-K.; Ahn, Y.-G.; Lee, S.-Y.; Lee, S.-C. Protective Activity against Oxidative Stress in Dermal Papillae with Extracted Herbal Essential Oils. Appl. Sci. 2023, 13, 3985. https://doi.org/10.3390/app13063985
Choi D-I, Choi J-Y, Lee J-B, Yun S-J, Moon B-K, Ahn Y-G, Lee S-Y, Lee S-C. Protective Activity against Oxidative Stress in Dermal Papillae with Extracted Herbal Essential Oils. Applied Sciences. 2023; 13(6):3985. https://doi.org/10.3390/app13063985
Chicago/Turabian StyleChoi, Da-In, Jee-Young Choi, Jee-Bum Lee, Sook-Jung Yun, Byung-Kwon Moon, Yong-Gook Ahn, Sook-Young Lee, and Seung-Chul Lee. 2023. "Protective Activity against Oxidative Stress in Dermal Papillae with Extracted Herbal Essential Oils" Applied Sciences 13, no. 6: 3985. https://doi.org/10.3390/app13063985
APA StyleChoi, D.-I., Choi, J.-Y., Lee, J.-B., Yun, S.-J., Moon, B.-K., Ahn, Y.-G., Lee, S.-Y., & Lee, S.-C. (2023). Protective Activity against Oxidative Stress in Dermal Papillae with Extracted Herbal Essential Oils. Applied Sciences, 13(6), 3985. https://doi.org/10.3390/app13063985