Abstract
Silybum marianum is a medicinal plant that produces silybins A and B, which exhibit liver-protective activities. The He–Ne laser pre-sowing seed treatment has been used as a physical biostimulator in numerous plants to promote seed germination, plant growth, and secondary metabolite production. Thus, in this study, a He–Ne laser at 632.8 nm was used to irradiate S. marianum seeds with the aim of promoting plant growth as well as enhancing the production of silybin (A + B). The seeds were irradiated with a He–Ne laser for 2, 4, 6, 8, and 10 min. The growth attributes of the plants, as well as the silybin (A + B) content and bioactivities of fruit extracts from various treatment groups, were compared. The results indicated that all laser seed treatments resulted in a subsequent increase in plant growth, with the highest yield observed in the 10 min laser treatment. An increase in fruit silybin (A + B) content and in chalcone synthase gene expressions were observed in plants from the 6-, 8-, and 10-min laser-irradiated seeds. The highest silybin (A + B) contents were observed in plants from the 6-min-irradiated seeds. Furthermore, the corresponding fruit extract also exhibited elevated cell growth inhibition in the Dictyostelium discoideum cell model.
1. Introduction
The pre-sowing laser irradiation of seeds has been shown to improve seed germination, seedling and plant growth and development, as well as the quality and quantity of crop yields [1,2]. Mechanisms such as the activation of phytochrome, phytohormones, and fermentative systems have been proposed to be involved in the laser-induced enhancement effects [3]. A red laser with a wavelength of 632.8 nm, which is produced by the helium–neon (He–Ne) gas laser, has been commonly used for seed irradiation. Factors such as plant species, method of irradiation, dose of irradiation, and the thermal moisture conditions under which the plants were cultivated were reported to affect the impact of the laser stimulation effects, as proposed by Hernadez’s [3] review and individual reports on various plant species such as pea [4], scorzonera [5,6], white lupine, and faba bean [7].
Silybum marianum is a medicinal plant that produces silymarin, a compound mixture of flavonolignans and flavonoids predominantly found in the fruits of the plant [8]. Silymarin has demonstrated hepato-, renal-, and cardio-protective effects, as well as antiviral, neurodegenerative, and cancer preventive effects in in vitro and/or in vivo systems [9]. Silybin A and B are the major components that represent about 50 to 70% of the silymarin composition and have been implicated in the bioactivities of silymarin [10]. Due to the medicinal importance of silymarin, strategies to enhance its production have been actively pursued by various research groups. These include the use of gamma irradiation [11,12], sodium chloride [11], or biotic elicitors such as methyl jasmonate [13,14] or fungal extract [11]. Banisharif and Amooghaie [15] attempted the use of He–Ne lasers in promoting S. marianum seed germination and seedling growth; however, the corresponding study did not proceed further to analyze the effect of lasers on the subsequent plant growth and yield, as well as the silybin A and B production.
Chalcone synthase (EC 2.3.1.74) is a group of enzymes that catalyzes the biosynthesis of taxifolin, the flavonoid precursor that forms the flavonolignans mixture of silymarin. Sepideh and coworkers [16] identified and partially cloned three S. marianum chalcone synthase genes (CHS1, CHS2, and CHS3) and found that they were differentially expressed in various organs of the plants during the early and late flowering stages. El-Garhy et al. [12] reported that these CHS genes were differentially activated by salinity and gamma radiation, and a strong association was observed between the genes’ activation profiles and the silymarin contents in the fruit samples. CHS1 and CHS3 had the highest transcript levels in petals during the early and mid-flowering stages of S. marianum; therefore, they are likely to be involved in anthocyanin and silymarin biosynthesis [16,17].
Dictyostelium discoideum is a slime mold that shares similar cell structure, behavior, and intracellular signaling with mammalian cells. Therefore, it is one of the eight non-mammalian cell models recognized by the National Institute of Health (NIH), USA, to be widely used in various biomedical studies involving human cells-related diseases and processes, including cancer, wound healing, development, and immune cell response [18]. The D. discoideum cell model offers a great advantage by being an indicator of the potential therapeutic functions of various natural components, including flavonoid compounds such as naringenin and epigallocatechin gallate [19].
Due to the promising biostimulation effects of the He–Ne laser (wavelength 632.8 nm) on various seeds, including that of S. marianum, it was thus applied in the present study to further explore its potential in increasing S. marianum plant growth and yield, as well as silybin A and B production. The expressions of genes involved in the biosynthesis of silybin A + B were measured to elucidate the underlying mechanism that contributed to the change in silybin (A + B) contents. The biological activities of fruit extract obtained from these plants were examined using the D. discoideum cell model system, which has been used to study the therapeutic values of several flavonoid compounds. Our results indicated that laser pretreatment of S. marianum seeds promotes subsequent plant growth and fruit yield. In addition, it also increased the silybin (A + B) contents and the therapeutic value of the fruits, as measured with the Dictyostelium discoideum slime mold model system.
2. Materials and Methods
2.1. Helium–Neon (He–Ne) Laser Irradiation
S. marianum seeds obtained from the Institute for Horticultural Science, Agricultural Research Center, Department of Medicinal and Aromatic Plants (ARC), Giza, Egypt, were divided into six laser treatment groups, with 50 seeds per group. The seeds in the “control” group were not irradiated with laser rays, while those in the other treatment groups were irradiated with laser rays for durations of 2, 4, 6, 8, and 10 min. The set of durations was chosen based on our previous study, where 10 min of Curcuma longa rhizome irradiation resulted in optimal effects on C. longa plant growth and yield [20]. A linearly polarized He–Ne laser beam of 632.8 nm wavelength and 5.5 mW/mm2 power density (power = 1 mW, diameter of ray = 0.48 mm) was delivered by a LeyboldR 471,830 device (Department of Physics, College of Science, King Faisal University). The seeds were placed in a Petri dish positioned at a distance of 40 mm below the laser device [20].
2.2. S. marianum Cultivation in a Greenhouse
All experimental S. marianum seeds were cultivated under greenhouse conditions at the Agriculture and Veterinary Research and Training Center, King Faisal University [21], from 1 October 2020, to 31 July 2021. A completely randomized experimental design with 20 replicates was followed. The greenhouse conditions, soil and irrigation water properties, and cultivation practices had been documented [21].
2.3. Measurement of Photosynthetic Pigment Contents and Attributes of Growth and Yield
Ten randomly selected plants from each treatment group were harvested 270 days after sowing. The leaf photosynthetic pigment content and the growth and yield attributes of these plants were measured and noted according to [21].
2.4. Quantification of Silybin (A + B) in S. marianum Fruits Using High Performance Liquid Chromatography (HPLC)
The silybin (A + B) contents of the S. marianum fruits of the 10 harvested plants were quantified against an authentic silybin (A + B) standard (Santa Cruz Biotechnology, Dallas, TX, USA) using a Waters 2690 Alliance HPLC (Milford, MA, USA) system equipped with a C18 Kromasil column (4.6 × 150 mm, 5 µm) and a Waters 996 photodiode array detector. The preparation of the silybin (A + B) standard and S. marianum methanolic fruit extracts, as well as the chromatography conditions, were carried out according to previously described methods [21].
2.5. Gene Expression Analyses of Chalcone Synthase 1, 2, and 3 by Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
The expressions of chalcone synthase genes 1, 2, and 3 in the petals from six S. marianum plants of each treatment group were analyzed using gene-specific primers. The collection and storage of petal samples, storage, total RNA extraction, cDNA synthesis, and RT-PCR were conducted according to [21].
2.6. Assay of S. marianum Fruit Methanolic Extract Bioactivities Using D. discoideum Cell Culture System
2.6.1. Dictyostelium discoideum Cell Lines and Cell Culture
The wild-type D. discoideum axenically growing strain AX2 was obtained from Dictybase, an online database and stock center. The cells were grown at 22 °C in HL-5 medium, including glucose from FORMEDIUM (Norfolk, England). Dihydrostreptomysin from Sigma (Dorset, United Kingdom) was added to the medium at a concentration of 0.05 g/mL.
2.6.2. D. discoideum Growth Assay
The assay was applied, as previously described [22]. The cells were grown in a shaker to reach the early exponential phase (1.5–2.5 × 106 cells/mL); then, they were diluted in HL-5 medium to 2 × 104 cells/mL. Aliquots of 500 μL were transferred to 24-well plates and incubated with various concentrations of S. marianum fruit extracts that were derived from either the 6-min laser-treated or untreated (control) S. marianum seeds. The original concentration of the extracts was 50 mg/mL with 50% DMSO as the solvent. The extracts were diluted with HL-5 medium to reach the desired concentrations (100 μg, 300 μg, and 600 μg). Control treatments consisted of cells incubated with DMSO at the same concentrations as solvents present in various S. marianum fruit extracts. The cell densities per mL were calculated over 3 days using a hemocytometer.
2.6.3. D. discoideum Development Experiment
For development phenotype assessment, the AX2 cells were centrifuged at 700× g and washed with development buffer (DB) (5 mM NaH2PO4, 5 mM Na2HPO4, 2 mM MgSO4, and 0.2 mM CaCl2). The cells at 2 × 107 were plated onto a DB-agar (1.5%) plate (90 mm dia.) containing S. marianum fruit extract (600 μg or 1200 μg) from plants of either the 6-min-laser-treated or laser-untreated S. marianum seeds. For control groups, cells at 2 × 107 were plated onto DB-agar (1.5%) −/+ DMSO (0.6% or 1.2%). Images were taken after 24 h of starvation using a Nikon Eclipse 80i microscope and a Nikon DS-Ri1 camera.
2.7. Statistical Analysis
The tests of homogeneity of variances and mean difference between the treatment groups were evaluated at a probability level of p = 0.05, using Tukey’s test for post hoc analysis from the Statistica 6 program ANOVA/MANOVA [23].
3. Results
Seed germination began simultaneously three days after sowing, and the germination rate was 100% in control as well as in the laser-irradiated seeds of S. marianum.
3.1. The Effect of He–Ne Laser Pre-Sowing Seed Treatment on the Vegetative Growth and Yield of S. marianum Plants
The means of the vegetative growth parameters are presented in Table 1. The results indicated that S. marianum plants from different groups of laser-irradiated seeds, except those irradiated for 8 min, produced plants with increased plant height and numbers of leaves, as well as dried weights of root and aerial parts, as compared with those plants derived from the control (non-irradiated) seeds. The plants from 8-min irradiated seeds decreased in root and aerial parts dry weight but increased in plant height and in the number of leaves as compared with the control. Plants from all irradiated seeds, except those irradiated for 2 min, had a lower number of branches as compared with the control. Interestingly, we observed that different irradiation durations resulted in selectively enhanced distinct growth parameters of the plants derived from the seeds. For example, the seeds irradiated for 4 min produced plants with the maximum plant height and root dry weight, while those irradiated for 10 min produced plants with the highest number of leaves and dried weight of aerial parts. On the other hand, plants from seeds that had been laser-irradiated for 2 min had the highest number of branches.

Table 1.
Effects of laser treatment on Silybum marianum L. plant height (cm), numbers of leaves (n), numbers of branches (n), and respective dry weights of root and aerial parts (g).
All irradiated seeds were found to produce plants with more capitula and higher fruit dry weight as compared with the control plants (Table 2). Plants with the highest capitula number (12.0) and fruit dry weight (49.84 g) were obtained from 10-min irradiated seeds. The 6 min and 8 min treatment groups displayed the lowest fruit dry weights and capitula numbers, respectively.
Table 2.
Effects of laser treatment on Silybum marianum L. capitula number (n) and fruit dry weight (g).
3.2. Effect of He–Ne Laser Pre-Sowing Seed Treatment on the Leaf Photosynthetic Pigments of S. marianum Plants
The data in Table 3 show that all He–Ne laser treatments positively affected the chlorophyll a (Chl a), chlorophyll b (Chl b), and carotenoid contents, as all irradiated seeds produced plants with higher photosynthetic pigments as compared with the control. Consistent with the yield, the 10 min-irradiation treatment was found to produce plants with the highest Chl a, Chl b, and carotenoids contents. The enhancement trends for different photosynthetic pigments varied. The enhancement in Chl-b contents in 2, 4, 6, and 8 min were similar, while the 10 min treatment was distinctly higher as compared with the control. On the other hand, all laser treatments resulted in distinct enhancements in Chl-a and carotenoids contents. Among all of the treatments, the 8-min treatment resulted in the least enhancement as compared with other irradiation durations.
Table 3.
Effects of laser treatment on chlorophyll a, b, and carotenoids contents. The contents of the pigments are enumerated as milligrams per 100 g of fresh Silybum marianum L. leaves.
3.3. Effect of He–Ne Laser Pre-Sowing Seed Treatment on the Leaf Silybin A + B Contents of S. marianum Plants
The silybin (A + B) contents in the fruits of S. marianum were analyzed using high-performance liquid chromatography (HPLC). Our results revealed that the control plants from non-irradiated seeds had an average silybin (A + B) content of 48.98 mg/100 g of dried fruit weight (D.W.). Plants from 2- and 4-min-irradiated seeds had lower silybin (A + B) contents than control plants; on the other hand, plants from 6-, 8-, and 10-min-irradiated seeds had higher silybin (A + B) contents than control plants. The highest silybin (A + B) content (55.66 mg/100 g D.W.) was recorded from plants derived from 6-min-irradiated seeds (Figure 1).
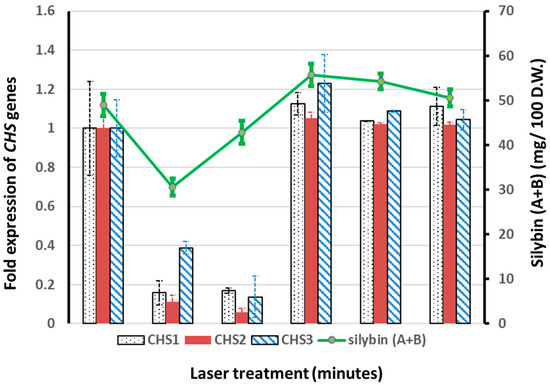
Figure 1.
Fold expressions of chalcone synthase (CHS) genes as related to the composition of silybin (A + B). The expressions of CHS1, CHS2, and CHS3 genes in plants from control (non-irradiated) treatments were normalized to one. The expressions of CHS genes for all other treatments (2, 4, 6, 8, and 10 min laser irradiation) were expressed as fold expressions with respect to the expressions of the corresponding genes from the control treatment. The content of silybin (A + B) was computed as milligrams (mg) per 100 g of fruit dry weight (D.W.), based on the HPLC quantification data.
3.4. Effect of He–Ne Laser Pre-Sowing Seed Treatment on the Expressions of Chalcone Synthase 1, 2, and 3 Genes in the Petals of S. marianum Plants
The effects of He–Ne laser seed pretreatment on the expression levels of the CHS1, 2, and 3 genes were investigated using quantitative reverse transcription-polymerase chain reaction (qRT-PCR). All chalcone synthase genes (CHS1, CHS2, and CHS3) were found to be down-regulated in plants derived from 2- and 4-min-irradiated seeds as compared with control plants. On the other hand, we observed a slight up-regulation of these genes (CHS1, CHS2, and CHS3) in plants derived from the 6-, 8-, and 10-min-irradiated seeds, with maxima of 1.1- (for the CHS1 and CHS2 genes) and 1.2- (for the CHS3 gene) fold expressions in the 6-min irradiated groups. In summary, consistent associations of the chalcone synthase gene expressions with the silybin (A + B) contents were observed in all the treatment groups (Figure 1).
3.5. Effect of He–Ne Laser Pre-Sowing Seed Treatment on the Bioactivities of S. marianum Fruit Extracts
3.5.1. The Anti-Dictyostelium discoideum Cell Growth Activity of S. marianum Fruit Extract
The high silybin (A + B) contents in fruit extracts of plants from 6-min-irradiated seeds prompted us to further investigate the bioactivities of these extracts as compared with those of plants from non-irradiated seeds. The growth inhibition effects of fruit extracts from the 6 min and control treatments were compared using the D. discoideum wild-type axenic strain Ax-2 cells. Cells were grown in HL-5 media in 24-well plates at a density of 2 × 104/mL per well. The cells were then treated with different concentrations (100 μg, 300 μg, and 600 μg) of laser-treated S. marianum extract or laser-untreated S. marianum. Then, the cells were counted with a hemocytometer over three days (Figure 2A–C). For control samples, cells were treated with DMSO at concentrations of 0.1%, 0.3%, and 0.6%, which were used as solvents for the S. marianum extracts.
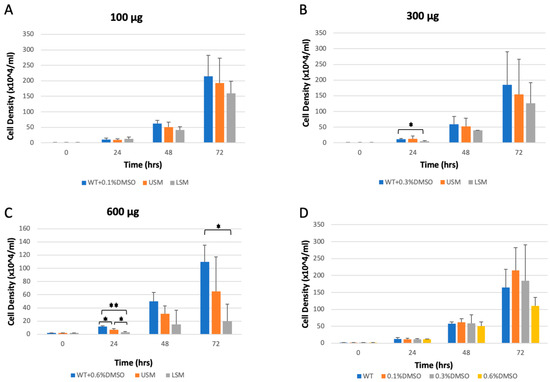
Figure 2.
Cell growth of Dictyostelium discoideum. Cells were grown in a 24-well plate, as described in Section 2.6.2. The AX2 cells (WT) were treated with a fruit extract of either 6-min laser-treated S. marianum (LSM) or untreated S. marianum (USM). Treatment with the solvent (DMSO) at a compatible percentage was used as a control. (A) AX2 cells treated with the extracts at the lowest concentration (100 μg). (B) AX2 cells treated with extracts at a concentration of 300 μg. (C) AX2 cells treated with the highest concentration (600 μg) of the extracts. (D) wild-type AX2 cell densities over three days compared to the DMSO-treated samples for the three concentrations used. The values represent three independent experiments +SD (* p < 0.05, ** p < 0.01).
The data showed no significant differences between the WT sample and the DMSO-treated samples as negative controls (Figure 2D). However, 6-min laser-treated S. marianum extract caused an inhibition of cell growth compared to the DMSO control and laser-untreated extract. A significant decrease was observed after 24 h of incubation with laser-treated S. marianum extract at concentrations of 300 μg and 600 μg compared to the DMSO control sample (Figure 2B,C). A significant decrease in cell growth was also observed after 72 h of incubation with laser-treated S. marianum extract at a concentration of 600 μg compared to the DMSO control sample (Figure 2C).
3.5.2. Effect of S. marianum Fruit Extracts on Dictyostelium discoideum Development Life Cycle
Dictyostelium cells are unicellular, feeding on bacteria and dividing by mitosis. When the food source is removed, cells start a development phase that begins with cAMP secretion and GPCR activation that links to heterotrimeric G proteins in order to initiate cell chemotaxis within the first 6 h. The D. discoideum development cycle completes with fruiting body formation after 24 h [18]. Chemotaxis is a very dynamic mechanism that is crucial for cancer metastasis. To determine the role of 6-min laser-treated S. marianum, the multicellular development cycle of D. discoideum was tested. The AX2 cell line was used for this study. The cells were washed with DB buffer and starved onto a DB agar plate along with 600 μg of either 6-min laser-treated S. marianum extract or laser-untreated S. marianum extract. The results showed that treatment with 600 μg of laser-treated S. marianum extract caused a noticeable delay in the development cycle and fruiting body formation (Figure 3G–I) compared to the laser-untreated S. marianum extract sample (Figure 3D–F) and the control sample (1.5% agar + 0.6% DMSO) (Figure 3B). To emphasize the effect of laser-treated S. marianum extract on the development cycle of D. discoideum, we doubled the concentrations of the treatments. Using 1200 μg of the extracts confirmed the conclusion reached. Adding 1200 μg of laser-treated S. marianum extract to the agar plate caused severe developmental delays (Figure 3M–O) compared to the sample treated with 1200 μg laser-untreated S. marianum extract (Figure 3J–L) and the control sample (1.5% agar + 1.2% DMSO) (Figure 3C). This effect was not due to the solvent (DMSO) concentration, as the results showed no differences between the development cycle of the AX2 cells on DB containing 1.5% agar only (Figure 3A) and the control samples (1.5% agar + 0.6% DMSO, Figure 3B, or 1.5% agar + 1.2% DMSO, Figure 3B). We also observed that treatment with 1200 μg of 6-min laser-treated S. marianum extract did not inhibit cells streaming; however, it caused a delay in this stage, as some of the population began to aggregate after 24 h of starvation (Figure 3M).
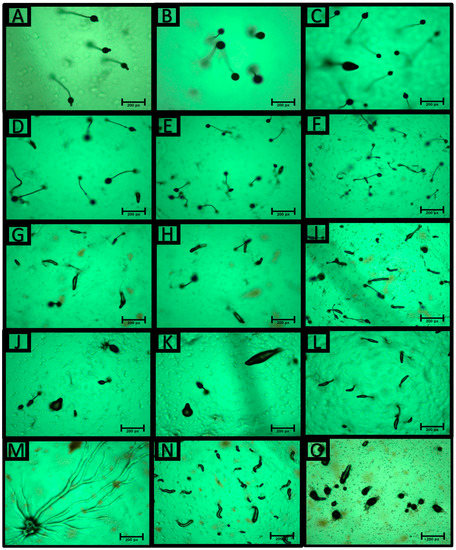
Figure 3.
Development phenotypes of Dictyostelium discoideum after 24 h of starvation. (A) Ax2 cells were washed with DB buffer and plated onto a DB (1.5% agar) plate at a density of 2 × 107 cells per plate (90 mm dia.). Ax2 cells were starved onto a DB agar plate (1.5%) containing 0.6% DMSO (B) or 1.2% DMSO (C) as controls. (D–F), Ax2 cells were starved onto a DB agar plate (1.5%) that contained 600 μg of laser-untreated S. marianum extract. (G–I), Ax2 cells were starved onto a DB agar plate (1.5%) containing 600 μg of laser-treated S. marianum extract. Images show the development phenotype of AX2 cells starved onto a DB agar plate (1.5%) containing 1200 μg of laser-untreated S. marianum extract (J–L) or 1200 μg of laser-treated S. marianum extract (M–O).
4. Discussion
In this study, S. marianum seeds were irradiated by a He–Ne laser with a wavelength of 632.8 nm for various durations (2, 4, 6, 8, and 10 min). The seeds were then grown, and the vegetative growth, fruit yield, photosynthetic pigments, silybin (A + B) content, and expressions of chalcone synthase genes in the mature plants were evaluated. The positive plant growth and crop yield impacts as a result of pre-sowing irradiated seeds have been documented in various plant species [1,2,7,20,24,25,26]. Consistent with reports from other plant species, we also observed similar enhancement effects in the growth, yield, and photosynthetic pigment contents of the S. marianum plants derived from seeds irradiated for different durations. Our observations further support the positive laser seed treatment effects observed by Banisharif and Amooaghaie [15] on S. marianum seeds and seedlings. The increases in photosynthetic pigment (Chl a and carotenoids) contents were highly correlated with improvements in growth and yield. The 10-min pre-sowing irradiation treatment showed the highest enhancement among the five irradiation durations examined in this study. It was reported that the effective range of laser treatments varied among plant species, and when the irradiation limit was reached, negative effects were observed [27,28]. In this study, we observed that the 8-min irradiation treatment showed the lowest enhancement in plant height, number of leaves, number of capitula, and contents of all photosynthetic pigments. In addition, the 8-min treatment also negatively impacted the dry weight of roots and aerial plant parts. However, the 10-min treatment increased values in all growth and yield parameters as well as photosynthetic pigment content compared with 8-min of irradiation. Therefore, we could not conclude that 8 min was the irradiation limit for S. marianum seeds.
Increased bioactive compounds have been reported in plants, such as fennel, coriander, anise, cumin, and curcuma, derived from laser-irradiated seeds or vegetative reproductive organs [20,29,30]. Similarly, in our study, in addition to laser-enhanced growth and yield, increased silybin (A + B) contents were also observed in the 6-, 8-, and 10-min laser treatment groups as compared with the control. For short irradiation durations (2 and 4 min), decreases in silybin (A + B) contents were observed despite the high growth rates and fruit yields. Despite having the highest growth and yield, plants from the 10-min-irradiated seeds were found to have lower silybin (A + B) contents as compared with those from the 6-min-irradiated seeds. This is consistent with the fact that secondary metabolites are produced in plants with lower growth and yield due to the redirection of plant resources from primary metabolism to secondary metabolism. Furthermore, in the present study, a strong correlation was also observed between the silybin (A + B) contents and expressions of the three chalcone synthase (CHS1, CHS2, and CHS3) genes. The stimulating effect of He–Ne laser irradiation on CHS gene transcription observed in the present study could likely be the contributing factor that led to the high silybin (A + B) contents, similar to the observed increase in phenolic compounds and flavonoids previously reported in Celosia argentea plants [31]. Using D. discoideum as a cellular model confirmed the fruit extract bioactivity differences between plants derived from the 6-min laser-irradiated S. marianum seeds and those derived from the non-laser-irradiated S. marianum seeds. Increasing the content of silybin (A + B) via a 6-min laser treatment seems to inhibit D. discoideum cell proliferation. A significant decrease in D. discoideum cell growth was observed after 24 h of incubation with laser-treated S. marianum extract at concentrations of 300 μg and 600 μg (Figure 2B,C). The significant inhibition of cell growth was also observed during the 72-h incubation with laser-treated S. marianum extract. Furthermore, adding 600 μg or 1200 μg of laser-treated S. marianum extract to the agar plate also caused noticeable delays in the D. discoideum development cycle. The high content of silybin (A + B) in the extracts may have been the major cause of the delay. The delay was severe with 1200 μg of laser-treated S. marianum extract, as we noticed some of the population proceeded through the streaming stage (Figure 3M) and mound formation (Figure 3N,O) after 24 h of starvation. These data suggested that silybin (A + B) could not prevent the streaming and the directed migration of the cells; however, silybin (A + B) caused a significant delay in the process, in addition to inhibiting cell growth. The data obtained here support and emphasize the anticancer activity of silybin (A + B), as previously reported [32,33].
5. Conclusions
The pre-sowing seed treatment with the He–Ne laser at 632.8 nm enhanced the growth, yield, and silybin (A + B) contents of S. marianum. Irradiating seeds for 10 min produced plants with the highest number of leaves, number of capitula, aerial part, and fruit dry weights, and photosynthetic pigment content, while 6 min of seed irradiation resulted in the highest silybin (A + B) content in the plants. The silybin (A + B) content was strongly correlated with the expression profiles of chalcone synthase genes. In addition, the increase in silybin (A + B) contents in the fruit extracts of the 6 min treatment was also reflected in the increase in cell inhibition and developmental delay activities in the D. discoideum cell model system. Our study suggests that 6 min of the He–Ne laser (632.8 nm) pre-sowing seed treatment could be a good strategy to produce S. marianum plants that yield fruits with increased silybin (A + B) contents and greater biological activities.
Author Contributions
Conceptualization, F.E.S. and S.K.; methodology, F.E.S. and Y.-K.Y.; formal analysis, F.E.S. and S.A.; investigation, F.E.S., Y.-K.Y. and S.A.; visualization, F.E.S., S.A. and Y.-K.Y.; writing—original draft preparation, F.E.S. and S.A.; writing—review and editing, F.E.S. and D.A.; project administration, S.K.; funding acquisition, F.E.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Deanship of Scientific Research, King Faisal University, grant number GRANT2,528.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank the greenhouse personnel from the King Faisal University’s Agriculture and Veterinary Research and Training Center for their valuable support and the Dictybase for providing the AX2 strain and for being a valuable source of information for protocols related to Dictyostelium.
Conflicts of Interest
The authors declare there are no conflict of interest.
References
- Dziwulska-Hunek, A.; Szymanek, M.; Stadnik, J. Impact of Pre-Sowing Red Light Treatment of Sweet Corn Seeds on the Quality and Quantity of Yield. Agriculture 2020, 10, 165. [Google Scholar] [CrossRef]
- Krawiec, M.; Dziwulska-Hunek, A.; Kornarzyński, K. The Use of Physical Factors for Seed Quality Improvement of Horticultural Plants. J. Hortic. Res. 2018, 26, 81–94. [Google Scholar] [CrossRef]
- Hernandez, A.C.; Dominguez, P.A.; Cruz, O.A.; Ivanov, R.; Carballo, C.A.; Zepeda, B.R. Laser in Agriculture-Review. Int. Agrophys. 2010, 2010, 407–422. [Google Scholar]
- Podlesna, A.; Gładyszewska, B.; Podlesny, J.; Zgrajka, W. Changes in the Germination Process and Growth of Pea in Effect of Laser Seed Irradiation. Int. Agrophys. 2015, 29, 485–492. [Google Scholar] [CrossRef]
- Krawiec, M.; Dziwulska-Hunek, A.; Sujak, A.; Palonka, S. Laser Irradiation Effects on Scorzonera (Scorzonera hispanica L.) Seed Germination and Seedling Emergence. Acta Sci. Pol. Hortorum Cultus 2015, 14, 145–158. [Google Scholar]
- Dziwulska-Hunek, A.; Krawiec, M.; Sujak, A. Laser Light Stimulation Effects on Scorzonera hispanica L. Seeds Germination, Field Emergence and Photosynthetic Pigments Content. J. Hortic. Res. 2016, 24, 57–62. [Google Scholar] [CrossRef]
- Podleśny, J.; Stochmal, A.; Podleśna, A.; Misiak, L.E. Effect of Laser Light Treatment on Some Biochemical and Physiological Processes in Seeds and Seedlings of White Lupine and Faba Bean. Plant Growth Regul. 2012, 67, 227–233. [Google Scholar] [CrossRef]
- Karkanis, A.; Bilalis, D.; Efthimiadou, A. Cultivation of Milk Thistle (Silybum marianum L. Gaertn.), a Medicinal Weed. Ind. Crops Prod. 2011, 34, 825–830. [Google Scholar] [CrossRef]
- Crocenzi, F.; Roma, M. Silymarin as a New Hepatoprotective Agent in Experimental Cholestasis: New Possibilities for an Ancient Medication. Curr. Med. Chem. 2006, 13, 1055–1074. [Google Scholar] [CrossRef]
- Ou, Q.; Weng, Y.; Wang, S.; Zhao, Y.; Zhang, F.; Zhou, J.; Wu, X. Silybin Alleviates Hepatic Steatosis and Fibrosis in NASH Mice by Inhibiting Oxidative Stress and Involvement with the Nf-ΚB Pathway. Dig. Dis. Sci. 2018, 63, 3398–3408. [Google Scholar] [CrossRef]
- El-Sherif, F.; Khattab, S.; Ibrahim, A.K.; Ahmed, S.A. Improved Silymarin Content in Elicited Multiple Shoot Cultures of Silybum marianum L. Physiol. Mol. Biol. Plants 2013, 19, 127–136. [Google Scholar] [CrossRef] [PubMed]
- El-Garhy, H.A.S.; Khattab, S.; Moustafa, M.M.A.; Abou Ali, R.; Abdel Azeiz, A.Z.; Elhalwagi, A.; El-Sherif, F. Silybin Content and Overexpression of Chalcone Synthase Genes in Silybum marianum L. Plants under Abiotic Elicitation. Plant Physiol. Biochem. 2016, 108, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Firouzi, A.; Mohammadi, S.A.; Khosrowchahli, M.; Movafeghi, A.; Hasanloo, T. Enhancement of Silymarin Production in Cell Culture of Silybum marianum (L) Gaertn by Elicitation and Precursor Feeding. J. Herbs. Spices Med. Plants 2013, 19, 262–274. [Google Scholar] [CrossRef]
- Madrid, E.; Corchete, P.N. Silymarin Secretion and Its Elicitation by Methyl Jasmonate in Cell Cultures of Silybum marianum Is Mediated by Phospholipase D-Phosphatidic Acid. J. Exp. Bot. 2010, 61, 747–754. [Google Scholar] [CrossRef]
- Banisharif, A.; Amooaghaie, R. The Effect of Laser Pretreatment on Seed Germination Indices and α-Amylase Activity and Growth of Silybum marianum L. Seedling in Normal Condition and under Pb Stress. J. Plant Res. (Iran. J. Biol.) 2020, 33, 285–298. [Google Scholar]
- Sepideh, S.; Shobbar, S.Z.; Ebrahimi, M.; Hasanloo, T.; Sadat-Noori, S.-A.; Tirnaz, S. Chalcone Synthase Genes from Milk Thistle (Silybum marianum): Isolation and Expression Analysis. J. Genet. 2015, 94, 611–617. [Google Scholar] [CrossRef]
- Drouet, S.; Tungmunnithum, D.; Lainé, É.; Hano, C. Gene Expression Analysis and Metabolite Profiling of Silymarin Biosynthesis during Milk Thistle (Silybum marianum (L.) Gaertn.) Fruit Ripening. Int. J. Mol. Sci. 2020, 21, 4730. [Google Scholar] [CrossRef]
- Martín-González, J.; Montero-Bullón, J.F.; Lacal, J. Dictyostelium Discoideum as a Non-Mammalian Biomedical Model. Microb. Biotechnol. 2021, 14, 111–125. [Google Scholar] [CrossRef]
- Schaf, J.; Damstra-Oddy, J.; Williams, R.S.B. Dictyostelium Discoideum as a Pharmacological Model System to Study the Mechanisms of Medicinal Drugs and Natural Products. Int. J. Dev. Biol. 2019, 63, 541–550. [Google Scholar] [CrossRef]
- El-Sherif, F.; Yap, Y.-K.; Ibrahim, H.I. Laser Irradiation Induces DNA Polymorphism and Alters Phytochemicals Compositions as Well as Growth and Yield of Curcuma longa. J. Dis. Med. Plants 2019, 5, 29–38. [Google Scholar] [CrossRef]
- Yap, Y.K.; El-sherif, F.; Habib, E.S.; Khattab, S. Moringa Oleifera Leaf Extract Enhanced Growth, Yield and Silybin Content While Mitigating Salt-induced Adverse Effects on the Growth of Silybum marianum. Agronomy 2021, 11, 2500. [Google Scholar] [CrossRef]
- Cocorocchio, M.; Baldwin, A.J.; Stewart, B.; Kim, L.; Harwood, A.J.; Thompson, C.R.L.; Andrews, P.L.R.; Williams, R.S.B. Curcumin and Derivatives Function through Protein Phosphatase 2A and Presenilin Orthologues in Dictyostelium Discoideum. DMM Dis. Model. Mech. 2018, 11, dmm032375. [Google Scholar] [CrossRef] [PubMed]
- StatSoft. STATISTICA Fur Windows; StatSoft: Tulsa, OK, USA, 2001. [Google Scholar]
- Urva; Shafique, H.; Jamil, Y.; ul Haq, Z.; Mujahid, T.; Khan, A.U.; Iqbal, M.; Abbas, M. Low Power Continuous Wave-Laser Seed Irradiation Effect on Moringa Oleifera Germination, Seedling Growth and Biochemical Attributes. J. Photochem. Photobiol. B 2017, 170, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Asghar, T.; Jamil, Y.; Iqbal, M.; Zia-ul-Haq; Abbas, M. Laser Light and Magnetic Field Stimulation Effect on Biochemical, Enzymes Activities and Chlorophyll Contents in Soybean Seeds and Seedlings during Early Growth Stages. J. Photochem. Photobiol. B 2016, 165, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Hanafiah, M.M.; Taha ZAeyad, A.I.; Said, M.N.M. Effect of Low-Intensity Laser Irradiation on Field Performance of Maize (Zea mays L.) Emergence, Phenological and Seed Quality Characteristics. Appl. Ecol. Environ. Res. 2020, 18, 6009–6923. [Google Scholar] [CrossRef]
- Podleoeny, J. Effect of Laser Irradiation on the Biochemical Changes in Seeds and the Accumulation of Dry Matter in the Faba Bean. Int. Agrophys. 2002, 16, 209–213. [Google Scholar]
- Musznski, S.; Gladyszewska, B. He-Ne Laser Irradiation Effect on Radish Seeds with Selected Germination Indices. Int. Agrophys. 2008, 22, 151–157. [Google Scholar]
- Osman, Y.A.H.; El Tobgy, K.M.K.; Sayed, E.; El Sherbini, A. Effect of Laser Radiation Treatments on Growth, Yield and Chemical Constituents of Fennel and Coriander Plants. J. Appl. Sci. Res. 2009, 5, 244–252. [Google Scholar]
- El Tobgy, K.M.K.; Osman, Y.A.H.; Sayed, E.; El Sherbini, A. Effect of Laser Radiation on Growth, Yield and Chemical Constituents of Anise and Cumin Plants. J. Appl. Sci. Res. 2009, 5, 522–528. [Google Scholar]
- Ali, S.I.; Gaafar, A.A.; Metwally, S.A.; Habba, I.E.; Khalek, M.R.A. The Reactive Influences of Pre-Sowing He-Ne Laser Seed Irradiation and Drought Stress on Growth, Fatty Acids, Phenolic Ingredients, and Antioxidant Properties of Celosia Argentea. Sci. Hortic. 2020, 261, 108989. [Google Scholar] [CrossRef]
- Agarwal, C.; Wadhwa, R.; Deep, G.; Biedermann, D.; Gažák, R.; Křen, V.; Agarwal, R. Anti-Cancer Efficacy of Silybin Derivatives—A Structure-Activity Relationship. PLoS ONE 2013, 8, e60074. [Google Scholar] [CrossRef] [PubMed]
- Deep, G.; Agarwal, R. Antimetastatic Efficacy of Silibinin: Molecular Mechanisms and Therapeutic Potential against Cancer. Cancer Metastasis Rev. 2010, 29, 447–463. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).