Abstract
β-N-acetylhexosaminidases have great potential in applied biocatalysis owing to their ability to act on a wide range of natural and modified substrates. In this work, β-N-acetylhexosaminidases from four Penicillium crustosum strains (PcHex) were studied. The production strains showed the highest enzymatic activity in the culture medium after 11–14 days of cultivation. The specific activity of the isolated and purified PcHex of hydrolysis of 4-nitrophenyl-N-acetyl-β-D-galactopyranoside was 15–20 U/mg protein. All isolated β-N-acetylhexosaminidases showed similar pH–activity profiles, with the optimum pH being 4.0–5.0 and the optimum temperature being 40 °C–50 °C. Apart from standard substrates, two synthetic substrates (5-bromo-4-chloro-3-indolyl-N-acetyl-β-D-galactoside and 2-chloro-4-nitrophenyl-β-D-galactopyranoside) were tested and successfully hydrolyzed using β-N-acetylhexosaminidase preparations. Maximum activity toward the fluorogenic substrate 4-methylumbelliferyl-N-acetyl-β-D-galactopyranoside (4MU-β-GalNAc) was observed for enzyme preparations from PcHex1 (15.38 U/mg protein) isolated from swab samples of books in the Slovak National Museum in Martin (Slovak Republic) and stored in our laboratory. The same enzyme preparation was used for the selective hydrolysis of β-anomer of 4MU-GalNAc from an anomeric mixture of 4MU-α/β-GalNAc. Thus, pure α-anomer (with the total yield being 90%) was separated from the mixture, which suggests the application potential of these enzymes.
1. Introduction
β-N-acetylhexosaminidases (Hex, EC 3.2.1.52) are a class of exo-glycosidases catalyzing the hydrolysis of the terminal β-linked N-acetylglucosamine (GlcNAc) and N-acetylgalactosamine (GalNAc) residues from glycoconjugates. They are ubiquitous in most living organisms (both prokaryotic and eukaryotic) and participate in several biological processes depending on the type of organism and their cellular localization. According to their origin, Hex can hydrolyze not only β-N-acetylhexosaminides but also β-glycosides [1]. These enzymes have gained attention owing to their structure, function, and specificity toward various substrates. They have great potential in applied biocatalysis based on transglycosylation or reverse hydrolysis reactions. In particular, fungal and recombinant Hex are capable to act over a broad range of natural and modified substrates and have been extensively used for synthetic purposes. Kinetically controlled transglycosylation reactions are predominantly used in synthetic applications. Different approaches and experimental conditions, such as high acceptor concentrations, the addition of co-solvents, choice of enzyme source, and removal of the transglycosylation product, have been chosen to promote the transglycosylation and prevent hydrolysis. Because of their synthetic potential, Hex have received great attention in the preparation of new bioactive glycosides (glycostructures, glycoconjugates) showing important effects on various biological processes such as immunoactivity, cancer metastasis, or hormone regulation [2,3].
To expand the current group of natural β-N-acetylhexosaminidases, we investigated the production and properties of fungal β-N-acetylhexosaminidases from Penicillium crustosum that have not been studied so far. The fungus P. crustosum was first isolated by Thom in 1930 [4]. Since then, it has been extensively used as an active producer of pectolytic and cellulolytic enzymes [5], a novel lipase [6], and schizophyllan degrading β-glucanase [7]. Moreover, P. crustosum has been used for the production of secondary metabolites such as roquefortine, penitrem, viridicatols, terrestric acid, and andrastin [8]. However, to the best of our knowledge, none of the previous studies have reported the production of β-N-acetylhexosaminidases by P. crustosum. Burtseva et al. [9] assessed active glucanases and glucosidases from 92 strains of fungi from various marine habitats, one of which was P. crustosum, which, however, showed no β-N-acetylhexosaminidase activity [9]. Thus far, the potential application of Hex in the production of α-anomers from synthetic substrates by enzymatic hydrolysis of the anomeric mixture has been described in the literature only twice, that is, once using bacterial Hex in the preparation of α-anomer from GlcNAc [10], and the other, using fungal Hex in the preparation of 4-nitrophenyl-N-acetyl-α-D-galactopyranoside (4-NP-α-GalNAc), which is our previous study [11].
In this research, we aimed to explore the cultivation conditions, biochemical properties, and substrate specificity of β-N-acetylhexosaminidases isolated from four P. crustosum (henceforth referred to as PcHex) strains. The ability of PcHex to hydrolyze modified glycosides used as synthetic substrates was also investigated, which has not yet been reported. Furthermore, the purified enzyme was used in the specific hydrolysis of b-anomer from chemically synthetized anomeric mixture of the fluorogenic substrate 4-metylumbelliferyl-N-acetyl-α/β-D-galactopyranoside (4MU-α/β-GalNAc).
2. Materials and Methods
2.1. Chemicals and Culture Media
4-Nitrophenyl-N-acetyl-β-D-gluco/galactopyranoside (4NP-β-Glc/GalNAc), 2-nitrophenyl-N-acetyl-β-D-gluco/galactopyranoside (2NP-β-Glc/GalNAc), 4-metylumbelliferyl-N-acetyl-β-D-galactopyranoside (4MU-β-GalNAc), 2-chloro-4-nitrophenyl-β-D-galactopyranoside (2Cl-4NP-β-Gal), and 5-bromo-4-chloro-3-indolyl-N-acetyl-β-D-galactoside (X-β-GalNAc) were purchased from Carbosynth, Ltd. (Compton, UK). The anomeric mixture of 4-methylumbelliferyl-N-acetyl-α/β-D-galactopyranoside (α:β anomer ratio = 4:3) was kindly provided by SynthCluster Co., Ltd. (Bratislava, Slovakia). All other chemicals were of analytical grade.
The cultivation medium contained the following constituents (g/L; pH 6.0 ± 0.2): KH2PO4, 3.0; NH4H2PO4, 5.0; (NH4)2SO4, 2.0; yeast extract, 0.5; NaCl, 15.0; MgSO4, 0.5; N-acetylglucosamine, 5.0. MgSO4 and N-acetylglucosamine were added after sterilization. Malt extract agar (MEA) contained the following constituents (g/L; pH 5.3 ± 0.2): malt extract, 20.0; peptone, 1.0; glucose, 20.0; agar, 20.0; stock solution of trace elements, 1 mL/L (0.5 g of CuSO4·5H2O, 0.1 g of ZnSO4·7H2O in 100 mL of dH2O). Czapek yeast autolysate agar (CYA) contained the following constituents (g/L; pH 7.3 ± 0.2): sucrose, 30.0; NaNO3, 3.0; KH2PO4, 1.0; KCl, 0.5; MgSO4·7H2O, 0.5; FeSO4·7H2O, 0.01; agar, 15.0. These media were sterilized in an autoclave at 121 °C for 20 min.
2.2. Microorganisms
The strain P. crustosum PcHex1 was isolated from samples taken by swabbing books in the Slovak National Museum in Martin (Slovak Republic) and stored in our laboratory. The strains P. crustosum CCF 3210 and CCF 3437 were obtained from the Culture Collection of Fungi (CCF), Department of Botany, Charles University in Prague (Czech Republic). The strain P. crustosum NRRL 62558 was obtained from the Agricultural Research Service Culture Collection (NRRL) in Peoria, Illinois. The strains were stored on MEA plates at 4 °C.
2.3. Cell Cultivation and Purification of ß-N-Acetylhexosaminidase
The medium (100 mL) in 500-mL flasks was inoculated with a suspension of spores in 0.1% (v/v) Tween 80. Cultivations were carried out in an orbital shaker at 28 °C and 200 rpm. After cultivation, the mycelium was removed by filtration, and the clarified medium was used for enzyme purification with cation exchange chromatography (Fractogel® EMD SO3−; Merck, Darmstadt, Germany) using the ÄKTA Purifier protein chromatography system (GE Healthcare, Uppsala, Sweden) according to the method described by Slámová et al. [12]. The purified enzyme was concentrated with an ultrafiltration membrane (Amicon® Ultra-4 10K, Merck, Kenilworth, NJ, USA) and stored in 1 M (NH4)2SO4 at 4 °C. Protein quantification was performed using the Bradford assay calibrated for bovine serum albumin.
2.4. Enzyme Assay
β-N-acetylhexosaminidase activity was measured spectrophotometrically using 4NP-β-GalNAc as the substrate at a starting concentration of 4 mM. One unit of enzyme activity was defined as the amount of enzyme that released 1 µmol of nitrophenol per minute in 50 mM citrate–phosphate buffer at pH 4.5, 35 °C, and under shaking (550 rpm) using a ThermoMixer® Comfort Shaker (Eppendorf, Cambridge, UK). Aliquots of 20 µL were taken at various time intervals and added to 2.0 mL of 0.1 M Na2CO3, and the amount of nitrophenol released was determined spectrophotometrically at 420 nm. All experiments were performed in triplicate. The results of spectrophotometric measurements were confirmed with high-performance liquid chromatography (column—Poroshell 120 EC-C18, 3 × 100 mm, 37 °C, mobile phase—methanol–water (15:85, 0.5 mL·min−1), detector diode array detector VL+). The enzyme activity toward the substrates 4NP-GlcNAc and 2NP-Glc/GalNAc was determined in the same manner.
Alternatively, other chromogenic and fluorogenic substrates were used to investigate substrate specificity, such as 4NP-β Glc/GalNAc, 2NP-β-Glc/GalNAc (the same procedure as that for 4NP-β-GalNAc), 4MU-β-GalNAc, 2Cl-4NP-β-Gal, and X-β-GalNAc (experimental conditions described in Table 1).

Table 1.
Experimental conditions for the tested substrates.
The enzyme activity of β-N-acetylhexosaminidases was calculated from the linear part of a plot where the hydrolysis course is represented as a function of the concentration of aglycon released and time. One unit of enzyme activity was defined as the amount of enzyme that releases 1 µmol of aglycone per minute. The specific activity was calculated as the enzyme activity divided by the amount of protein used for hydrolysis. All experiments were performed in triplicate.
2.5. Resolution of the Anomeric Mixture of 4MU-β-GalNAc by Enzyme Hydrolysis
The reaction mixture contained the substrate 4MU-α/β-GalNAc (4.5 mM solution in citrate–phosphate buffer, 50 mM, pH 4.5, 10 mL) and the purified enzyme PcHex1 (0.22 U). Hydrolysis was performed in 50 mL glass reactors at 35 °C for 4 h under stirring at 450 rpm. The reaction was stopped by heating the mixture solution to 100 °C for 5 min. The α-anomer of 4MU-GalNAc was isolated by extraction with organic solvents according to the protocol established by Hronská et al. [11].
3. Results and Discussion
3.1. Production, Purification, and Characterization of ß-N-Acetylhexosaminidases from P. crustosum Strains
β-N-acetylhexosaminidases are an important member of the binary chitinolytic system of filamentous fungi, which is responsible for the formation of septa and hyphenation of fungal cells. The activity and excretion of β-N-acetylhexosaminidases into the extracellular space are constantly controlled by the cell and depend on the stage of the fungal growth [13]. To our knowledge, no studies have investigated the enzymatic activity of Hex in P. crustosum strains. In this work, Hex activity was tested in four strains of P. crustosum. The strain P. crustosum PcHex1 was obtained from a set of microscopic fungal isolates from damaged archived materials of the Slovak National Museum in Martin (Slovak Republic). Colonies of the strain grown on CYA medium appeared a grayish green color with white margins (Figure 1A). Additionally, they had asymmetrically branched conidiophores with a typical warty stem, and the conidia were smooth and oval, arranged in 3.0–4.0 µm chains when observed under the microscope (Figure 1B). Based on the micromorphological and colony characteristics, the strain PcHex1 was identified as P. crustosum according to the Culture Collection of Fungi (CCF, Prague, Czech Republic). The other tested P. crustosum strains were obtained as isolates from international collections of microorganisms.
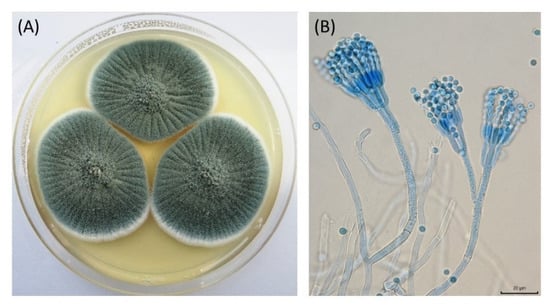
Figure 1.
Colony morphology (A) and microscopic characteristics (B) of Penicillium crustosum PcHex1. The scale bar represents 20 μm.
The strains of P. crustosum were grown in a culture medium containing N-acetylglucosamine as the β-N-acetylhexosaminidase inducer for 15 days. The highest level of enzyme activity in the medium was achieved after 11–12 days of cultivation for the P. crustosum strains CCF 3437 and PcHex1 and after 13–14 days for the strains CCF 3210 and NRRL 62558 (typically 0.15–0.25 U/mL and 9–11 U/mg protein, respectively; Figure 2). These results are comparable with those of previous studies wherein the specific activity of fungal Hex from P. oxalicum CCF 1959 and Aspergillus versicolor CCF 2491 was 10.8 U/mg protein [14] and 11 U/mg protein (0.4 U/mL), respectively [15]. The enzyme was purified using cation exchange chromatography and stored in 1 M ammonium sulfate at 4° C (SDS-PAGE electrophoresis is shown in Figure S1). The specific activity of the stock enzyme was typically 15–20 U/mg protein, and after 6 months, the decrease was not more than 5% of its original activity.
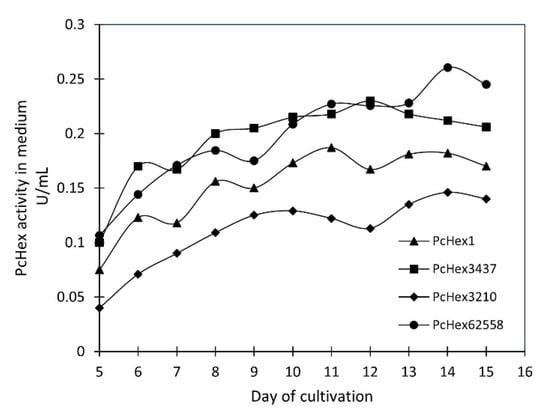
Figure 2.
Time course of the extracellular Hex activity of PcHex3437, PcHex3210, PcHex1, and PcHex62558 in the culture medium containing 0.5% N-acetylglucosamine as the inducer.
3.2. Effect of pH and Temperature on the Activity of ß-N-Acetylhexosaminidases
In general, the pH optimum of fungal hexosaminidases lies in the acidic region [16], and it usually has quite a broad range. The maximum hydrolytic activity of all four PcHex strains tested was in the same pH range of 3.5–5.0 (Figure 3), which is similar to that of hexosaminidases isolated from P. oxalicum (pH optimum 2–4; [14]), Talaromyces flavus (pH 2.5–5.0; [12]), and Aspergillus versicolor (pH 3.0–6.0; [15]). PcHex3210 and PcHex62558 were more sensitive to minor pH changes, whereas PcHex3437 and PcHex1 showed good stability over the entire tested pH range.
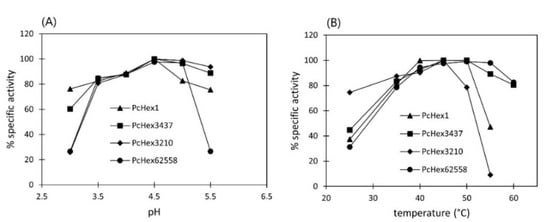
Figure 3.
Effect of pH (A) and temperature (B) on the specific activity of Hex. Reaction conditions: reaction volume 2 mL, 4-NP-β-GalNAc concentration 4 mM, enzyme activity 0.2–0.3 U. The pH profile was investigated at 37 °C, and the temperature profile was investigated at pH 4.5.
The optimal temperature of fungal hexosaminidases also varies in a wider range, usually 35–60 °C [14,16]. Hex isolated from P. crustosum showed temperature optimum at around 40–50 °C. Both PcHex1 and PcHex3210 were sensitive to a temperature increase beyond the optimum, and they lost up to 90% of their activity at 55 °C. The temperature profile of PcHex3437 and PcHex62558 showed a broader temperature optimum; however, after 15 min of incubation at 55 °C, both enzymes started to lose their activity, and after 60 min of incubation, they became completely inactive.
3.3. Substrate Specificity toward 4NP-ß-Glc/GalNAc and 2NP-ß-Glc/GalNAc
β-N-acetylhexosaminidases are characterized by their inherent dual catalytic activity, which allows them to act over two substrates differing in the configuration of the hydroxyl group on C-4 of the glycon, which may be N-acetylglucosamine (GlcNAc) or N-acetylgalactosamine (GalNAc). This specificity characteristic is commonly tested on substrates such as 4NP-β-GalNAc and 4NP-β-GlcNAc, which are largely accepted by a wide range of β-N-acetylhexosaminidases. Moreover, in this study, we used nitrophenyl glycosides with a nitro group binding to the phenyl group in position 2, namely, 2NP-β-GalNAc and 2NP-β-GlcNAc, because the GlcNAcase/GalNAcase ratio may also be affected by the aglycon structure [17]. In general, β-N-acetylhexosaminidases prefer D-glucose structures, with the GlcNAcase-to-GalNAcase ratio ranging from 1.5 to 4.0 [3].
As shown in Table 2, Hex produced by P. crustosum strains showed higher specific activity toward 2-nitrophenyl substrates, which indicated their preference for 2-nitrophenyl aglycon over 4-nitrophenyl, apart from the strain PcHex3437, for which the difference was not very significant. Concerning the preferences for glucose or galactose, Hex isolated from all four strains of P. crustosum exhibited a GalNAcase/GlcNAcase ratio similar to that of various species of Penicillium, namely, 0.9 for Penicillium chrysogenum [18], 1.2 for Penicillium funiculosum, or 1.15 for Penicillium purpurogenum [19], but still lower than that of P. oxalicum strains, for which the GalNAcase/GlcNAcase ratio usually ranged from 1.4 to 2.3 [19,20]. The determination of the preference for GalNAcase/GlcNAcase is a delicate matter, as the preference not only depends on the producing strain but also varies during microorganism cultivation and can be influenced by the presence of some inorganic salts or organic solvents [19].
Table 2.
Substrate specificity of the isolated β-N-acetylhexosaminidases.
Kinetic constants describing the hydrolysis of the substrates 4NP-β-GalNAc, 4NP-β-GalNAc, 2NP-β-GalNAc, and 2NP-β-GlcNAc were determined for all four PcHex. The kinetics of the hydrolysis of 4NP-β-GalNAc, 4NP-β-GalNAc, 2NP-β-GalNAc, and 2NP-β-GlcNAc was described using the Michaelis–Menten equation. The kinetic parameters kcat (catalytic constant), and Km (Michaelis constant) were determined using nonlinear regression from the initial reaction rate versus substrate concentration data using MATLAB R2022a (lsqnonlin procedure). When comparing the kcat/Km values shown in Figure 4, the most efficient catalyst overall was PcHex1, with the highest kcat/Km values for almost all tested substrates, except for 2NP-β-GlcNAc. The catalytic efficiency of PcHex1 was more than threefold higher than that of PcHex3437. Three of the four PcHex catalyzed the hydrolysis of 4NPGlcNAc the most efficiently. Only the strain PcHex3210 operated most efficiently with the substrate 2NP-β-GlcNAc.
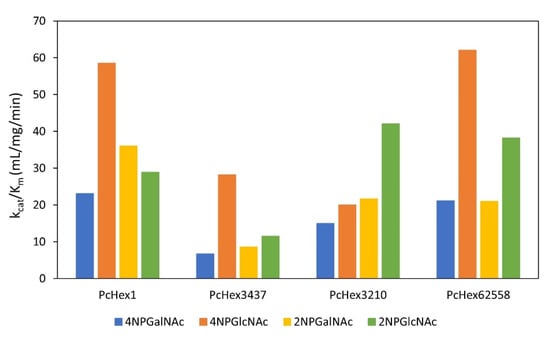
Figure 4.
Kinetic parameter analysis describing hydrolysis of 4NP-β-Gal/GlcNAc and 2NP-β-Gal/GlcNAc by PcHex.
3.4. Substrate Specificity toward Nonconventional Substrates
The specific activities of the purified PcHex were measured with three different nonconventional substrates: 4-methylumbeliferyl-2-acetamido-2-deoxy-β-D-galactopyranoside (4MU-β-GalNAc), 5-bromo-4-chloro-3-indolyl-N-acetyl-β-D-galactopyranoside (X-β-GalNAc), and 2-chloro-4-nitrophenyl-β-D-galactopyranoside (2Cl-4NP-β-Gal; Figure 5). They belong to the group of synthetic fluorogenic/chromogenic enzyme substrates, and the properties of Hex acting on these substrates have not yet been studied.
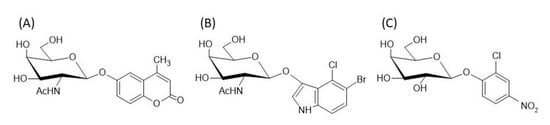
Figure 5.
Structures of the synthetic substrates (A) 4MU- β -GalNAc, (B) X-β-GalNAc, and (C) 2Cl-4NP-β-Gal.
Most of the fluorogenic substrates contain a fluorescent coumarin heterocycle, such as 4-methylumbelliferone (4-MU), which is often used, for example, for the detection of glycosidases and phosphatases [21]. The fluorescence of 4-MU is strongly dependent on pH and reaches the maximum at pH 9–10 [22]. Derivatives of 4-methylumbelliferyl provide a more sensitive detection method than nitrophenyl glycosides, which are frequently used for carbohydrate-active enzymes [23]. 4-Methylumbelliferyl-N-acetyl-β-D-glucosamide has been successfully used for the early detection of fungal growth on various cultural heritage materials before it was visible to the human eye [24]. PcHex1 (15.38 U/mg protein) showed the highest specific activity against 4-MUGalNAc with 98% hydrolysis after 120 min. The other three enzymes showed threefold lower activity, approximately 5 U/mg protein (Table 3).
Table 3.
Enzyme hydrolysis of 4MU-β-GalNAc, X-β-GalNAc, and 2Cl-4NP-β-Gal.
X-β-GalNAc is a synthetic substrate consisting of N-acetylgalactopyranoside and aglycone 5-bromo-4-chloroindoxyl, which is commonly referred to as X. Indoxyl substrates are very popular among chromogenic substrates because of their intensive color, which appears when two indoxyl molecules dimerize under oxidative conditions. This compound is insoluble, which provides highly localized labeling of the target. Probably the best-known indoxyl substrate is 5-bromo-4-chloro-3-indolyl β-D-galactopyranoside (X-Gal), which is used in molecular biology applications to distinguish nonrecombinant transformant colonies (blue) from recombinant colonies carrying the Lac Z gene (white) [25]. The hydrolysis of X-β-GalNAc was catalyzed by all four purified PcHex, although their specific activity toward this substrate was significantly lower than that toward the other two substrates. The highest specific activity was observed for the enzyme PcHex 62558 (0.28 U/mg protein), with an almost 94% degree of hydrolysis after 22 h.
Testing hydrolysis reactions using hexosaminidase with 2Cl-4NP-β-Gal as the substrate has not yet been described in the literature, although this substrate was used to measure the level of salivary α-amylase in athletes [26] and to analyze the correlation between pain and salivary α-amylase levels during dental treatment in children with Down syndrome [27]. During the hydrolysis of 2Cl-4NP-β-Gal, 2-chloro-4-nitrophenol is released, which causes a more intense yellow color than that generated by 4-nitrophenol. Hydrolysis of this substrate was surprising for us because, unlike other substrates assessed in this study, 2Cl-4NP-β-Gal lacks the N-acetyl group on galactopyranoside. This group is usually crucial for β-N-acetylhexosaminidases, as they are sensitive to its modification. However, fungal β-N-acetylhexosaminidases tolerate certain N-acyl-modified substrates that have sterical changes at C-2 (shorter or longer acyls, a hydroxyl group instead of a hydrogen atom) [17]. The hexosaminidase PcHex3437 could hydrolyze this substrate easily (enzyme activity: 3.97 U/mg), with the degree of hydrolysis reaching 96% in 30 min of reaction, similar to PcHex1.
The observation of the ability to catalyze the hydrolysis of chlorinated derivatives of other than N-acetylated saccharides is so interesting that it would be useful to focus future research on verifying this ability using β-N-acetylhexosaminidases in a highly purified form and, this way, exclude the presence of contaminating activities of other enzymes those presence in pre-purified preparations cannot be excluded with certainty.
3.5. Separation of an Anomeric Mixture of 4MU-α/β-GalNAc by Selective Hydrolysis
Fungal β-N-acetylhexosaminidases can selectively hydrolyze β-anomer in a mixture of α- and β-substituted GalNAc derivatives. It is a convenient method for preparing α-anomers of synthetic substrates suitable for the rapid identification and quantification of specific enzymes. This method is favorable because the enzymes are inexpensive and well-characterized, and the reaction is carried out under mild reaction conditions. After the hydrolysis reaction, α-anomer can be easily separated from the reaction mixture by extraction [11].
This is the first study to report the ability of hexosaminidases to efficiently hydrolyze β-anomer of 4MU-GalNAc in a chemically prepared anomeric mixture of 4MU-α/β-GalNAc (Scheme 1). The main product of this hydrolysis reaction, namely, 4MU-α-GalNAc, is a fluorogenic synthetic substrate, which allows the sensitive determination of α-N-acetylgalactosaminidase (α-NAGA), an important marker monitored in clinical practice. It is naturally produced by the human body and is required by cells to degrade glycolipids and glycoproteins. Genetic mutations in the NAGA gene can cause a loss of enzyme activity, leading to a serious disease called Schindler’s disease [28].
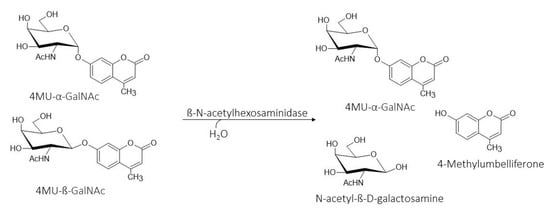
Scheme 1.
Preparation of anomerically pure 4MU-α-GalNAc from a chemically prepared anomeric mixture of 4MU-α/β-GalNAc.
For the separation of the anomeric mixture, we chose the hexosaminidase from the P. crustosum strain PcHex1, which showed the highest specific activity toward the substrate 4MU-β-GalNAc (Table 2). The hydrolysis reaction was carried out under standard conditions in citrate–phosphate buffer (pH 4.5). After 4 h, 99.1% of the β-anomer was hydrolyzed (Figure 6). The reaction mixture consisted of α-anomer of 4-MU-GalNAc, 4-MU aglycone, and N-acetylgalactosamine. After double-solvent extraction with ethyl acetate, 98% of 4-MU was removed from the reaction mixture, leaving 91% of the α-anomer in the aqueous phase along with N-acetylgalactosamine. The aqueous phase was subsequently extracted thrice with n-butanol. The total yield of 4MU-α-GalNAc extracted into n-butanol was 90%.
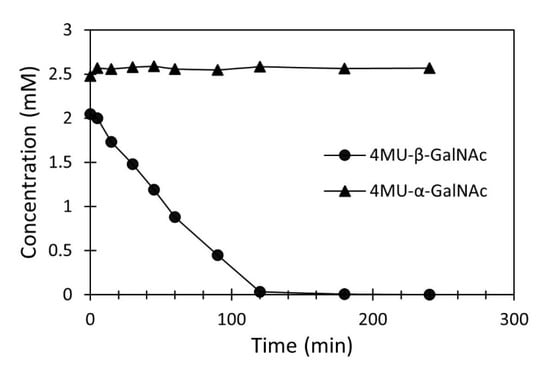
Figure 6.
Enzymatic hydrolysis of 4MU-β-GalNAc in the chemically synthesized mixture of 4MU-α/β-GalNAc.
As for further simplification of the process, the buffer in the reaction medium was replaced with distilled water. The 4MU-α/β-GalNAc anomer mixture was thus dissolved in distilled water, and the pH of the mixture was adjusted to pH 4.5 at the beginning of the reaction and was then controlled during hydrolysis. The enzymatic activity of Hex and the product yield remained the same. The improved procedure for the preparation of α-anomeric derivatives of glycosides enables the effective production of 4MU-α-GalNAc in a buffer-free environment, thus, minimizing the impurity content in the final product.
β-N-acetylhexosaminidases from Penicillium crustosum presented in this work have proved a great potential in the hydrolytic resolution of glycoside anomeric mixtures. Moreover, the acceptance of novel synthetic substrates by these enzymes additionally opens up opportunities for research in the biosynthesis of non-natural oligosaccharides.
4. Conclusions
In conclusion, β-N-acetylhexosaminidases from P. crustosum were isolated and characterized for the first time in terms of their biochemical properties, kinetic parameters, and substrate specificity toward several synthetic substrates. The relatively broad substrate promiscuity and easy handling of β-N-acetylhexosaminidases were promising properties for their use in the hydrolysis and synthesis of various glycosides. In this research, PcHex1 was successfully applied to prepare anomerically pure 4MU-α-GalNAc from a chemically synthesized anomeric mixture of 4MU-α/β-GalNAc through total enzymatic hydrolysis of the corresponding β-anomer. The findings of this research will aid in the preparation of α-anomers of substituted glycosides as the synthetic substrates for rapid enzyme-based diagnosis.
5. Patents
The above process is patented under Slovak Patent Application no. 50042–2018 filed on 9 September 2018.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app13063399/s1, Figure S1: SDS-PAGE electrophoresis of PcHex1 enzyme preparation after purification with cation exchange chromatography.
Author Contributions
Conceptualization, E.O. and H.H.; methodology, H.H., E.O. and V.Š.; formal analysis, V.Š.; investigation, E.O., M.B. and H.H.; data curation, E.O., M.B., H.H. and V.Š.; writing—original draft preparation, E.O. and H.H.; writing—review and editing, H.H., E.O., M.B. and V.Š.; supervision, H.H. and M.R.; funding acquisition, M.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Agency for supporting research and development, according to the agreement No. APVV-20–0208.
Acknowledgments
The authors would like to thank Alena Kubátová, curator of the Collection of Cultures of Fungi (CCF) in Prague, for her precise work in identifying the culture of the filamentous fungus Penicillium crustosum PcHex1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mayer, C.; Vocadlo, D.J.; Mah, M.; Rupitz, K.; Stoll, D.; Warren, R.A.J.; Withers, S.G. Characterization of a beta-N-acetylhexosaminidase and a beta-N-acetylglucosaminidase/beta-glucosidase from Cellulomonas fimi. FEBS J. 2006, 273, 2929–2941. [Google Scholar] [CrossRef]
- Ogata, M.; Zeng, X.; Usui, T.; Uzawa, H. Substrate specificity of N-acetylhexosaminidase from Aspergillus oryzae to artificial glycosyl acceptors having various substituents at the reducing ends. Carbohydr. Res. 2007, 342, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Slámová, K.; Bojarová, P.; Petrásková, L.; Křen, V. β-N-Acetylhexosaminidase: What’s in a name…? Biotechnol. Adv. 2010, 28, 682–693. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef]
- Hamlyn, P.F.; Wales, D.; Sagar, B.F. Extracellular Enzymes of Penicillium. In Penicillium and Acremonium; Peberdy, J.F., Ed.; Springer: Boston, MA, USA, 1987; pp. 245–284. [Google Scholar] [CrossRef]
- Rigo, E.; Ninow, J.L.; Tsai, S.M.; Durrer, A.; Foltran, L.L.; Remonatto, D.; Sychoski, M.; Vardanega, R.; de Oliveira, D.; Treichel, H.; et al. Preliminary Characterization of Novel Extra-cellular Lipase from Penicillium crustosum Under Solid-State Fermentation and its Potential Application for Triglycerides Hydrolysis. Food Bioprocess Technol. 2012, 5, 1592–1600. [Google Scholar] [CrossRef]
- Sutivisedsak, N.; Leathers, T.; Bischoff, K.; Nunnally, M.; Peterson, S.W. Novel sources of β-glucanase for the enzymatic degradation of schizophyllan. Enzym. Microb. Technol. 2013, 52, 203–210. [Google Scholar] [CrossRef]
- Sonjak, S.; Frisvad, J.; Gunde-Cimerman, N. Comparison of secondary metabolite production by Penicillium crustosum strains, isolated from Arctic and other various ecological niches. FEMS Microbiol. Ecol. 2005, 53, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Burtseva, I.V.; Sova, V.; Pivkin, M.; Anastiuk, S.; Gorbach, V.; Zviagintseva, T.N. Distribution of O-glycosylases in marine fungi of the Sea of Japan and the Sea of Okhotsk: Characterization of exocellular N-acetyl-beta-D-glucosaminidase of the marine fungus Penicillium canescens. Prikl. Biokhim. Mikrobiol. 2010, 46, 700–708. [Google Scholar] [PubMed]
- Wang, S.Y.; Laborda, P.; Lu, A.M.; Wang, M.; Duan, X.C.; Liu, L.; Voglmeir, J. Chemo-enzymatic approach to access diastereopure α-substituted GlcNAc derivatives. J. Carbohydr. Chem. 2016, 35, 423–434. [Google Scholar] [CrossRef]
- Hronská, H.; Štefuca, V.; Ondrejková, E.; Bláhová, M.; Višňovský, J.; Rosenberg, M. Chemo-Enzymatic Production of 4-Nitrophenyl-2-acetamido-2-deoxy-α-D-galactopyranoside Using Immobilized β-N-Acetylhexosaminidase. Catalysts 2022, 12, 474. [Google Scholar] [CrossRef]
- Slámová, K.; Bojarová, P.; Gerstorferová, D.; Fliedrová, B.; Hofmeisterová, J.; Fiala, M.; Pompach, P.; Křen, V. Sequencing, cloning and high-yield expression of a fungal β-N-acetylhexosaminidase in Pichia pastoris. Protein Expr. Purif. 2012, 82, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Plíhal, O.; Sklenář, J.; Hofbauerová, K.; Novák, P.; Man, P.; Pompach, P.; Kavan, D.; Ryšlavá, H.; Weignerová, L.; Charvátová-Pišvejcová, A.; et al. Large propeptides of fungal beta-N-acetylhexosaminidases are novel enzyme regulators that must be intracellularly processed to control activity, dimerization, and secretion into the extracellular environment. Biochemistry 2007, 46, 2719–2734. [Google Scholar] [CrossRef]
- Ryšlavá, H.; Kalendová, A.; Doubnerová, V.; Skočdopol, P.; Kumar, V.; Kukačka, Z.; Pompach, P.; Vaněk, O.; Slámová, K.; Bojarová, P.; et al. Enzymatic characterization and molecular modeling of an evolutionarily interesting fungal β-N-acetylhexosaminidase: β-N-Acetylhexosaminidase from Penicillium oxalicum. FEBS J. 2011, 278, 2469–2484. [Google Scholar] [CrossRef] [PubMed]
- Bojarová, P.; Kulik, N.; Slámová, K.; Hubálek, M.; Kotik, M.; Cvačka, J.; Pelantová, H.; Křen, V. Selective β-N-acetylhexosaminidase from Aspergillus versicolor—A tool for producing bioactive carbohydrates. Appl. Microbiol. Biotechnol. 2019, 103, 1737–1753. [Google Scholar] [CrossRef]
- Scigelova, M.; Crout, D.H.G. Microbial β-N-acetylhexosaminidases and their biotechnological applications. Enzym. Microb. Technol. 1999, 25, 3–14. [Google Scholar] [CrossRef]
- Fialová, P.; Weignerová, L.; Rauvolfová, J.; Přikrylová, V.; Pišvejcová, A.; Ettrich, R.; Kuzma, M.; Sedmera, P.; Křen, V. Hydrolytic and transglycosylation reactions of N-acyl modified substrates catalysed by β-N-acetylhexosaminidases. Tetrahedron 2004, 60, 693–701. [Google Scholar] [CrossRef]
- Pócsi, I.; Pusztahelyi, T.; Bogáti, M.; Szentirmai, A. The formation of N-acetyl-β-D-hexosaminidase is repressed by glucose inPenicillium chrysogenum. J Basic Microbiol. 1993, 33, 259–267. [Google Scholar] [CrossRef]
- Weignerová, L.; Vavrušková, P.; Pišvejcová, A.; Thiem, J.; Křen, V. Fungal β-N-acetylhexosaminidases with high β-N-acetylgalactosaminidase activity and their use for synthesis of β-GalNAc-containing oligosaccharides. Carbohydr. Res. 2003, 338, 1003–1008. [Google Scholar] [CrossRef]
- Yamamoto, K.; Lee, K.; Kumagai, H.; Tochdcura, T. Purification and Characterization of β-N -Acetylhexosaminidase from Penicillium oxalicum. Agric. Biol. Chem. 1985, 49, 611–619. [Google Scholar] [CrossRef]
- Orenga, S.; James, A.; Manafi, M.; Perry, J.; Pincus, D.H. Enzymatic substrates in microbiology. J. Microbiol. Methods 2009, 79, 139–155. [Google Scholar] [CrossRef]
- Profeta, G.S.; Pereira, J.A.; Costa, S.G.; Azambuja, P.; Garcia, E.S.; Moraes, C.D.S.; Genta, F.A. Standardization of a Continuous Assay for Glycosidases and Its Use for Screening Insect Gut Samples at Individual and Populational Levels. Front. Physiol. 2017, 8, 308. [Google Scholar] [CrossRef] [PubMed]
- Van Tilbeurgh, H.; Loontiens, F.; De Bruyne, C.; Claeyssens, M. Fluorogenic and chromogenic glycosides as substrates and ligands of carbohydrases. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1988; Volume 160, pp. 45–59. [Google Scholar] [CrossRef]
- Konkol, N.; McNamara, C.; Mitchell, R. Fluorometric detection and estimation of fungal biomass on cultural heritage materials. J. Microbiol. Methods 2010, 80, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Juers, D.H.; Matthews, B.; Huber, R.E. LacZ β-galactosidase: Structure and function of an enzyme of historical and molecular biological importance: LacZ β-Galactosidase. Protein Sci. 2012, 21, 1792–1807. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.M.D.; Jaramillo, O.B.; Teixeira, R.; Espindola, F.S. Salivary Surrogates of Plasma Nitrite and Catecholamines during a 21-Week Training Season in Swimmers. PLoS ONE 2013, 8, e64043. [Google Scholar] [CrossRef]
- Yumasdhika, F.; Suharsini, M.; Indiarti, I.S.; Anggraeni, H.D. Correlation between FLACC Pain Score and Salivary Alpha-Amylase Level (A Review on Children with Down Syndrome). J. Int. Dent. Med. Res. 2017, 10, 529–532. [Google Scholar]
- Ferreira, C.R.; Gahl, W.A. Lysosomal storage diseases. TRD 2017, 2, 1–71. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).