Featured Application
Authors are encouraged to provide a concise description of the specific application or a potential application of the work. This section is not mandatory.
Abstract
The production of pikeperch in a recirculation aquaculture system (RAS) is of growing importance. However, the use of combined technologies may also be justified, especially in countries with larger pond areas. However, this requires the most effective adaptation of pellet-consuming individuals to pond conditions. Foraging training, a form of environmental enrichment to compensate for the poor environment in hatchery rearing, can play a major role in this. The aim of our study was to investigate the prey size preference already observed in wild pikeperch, during prey capture by intensively reared naive individuals. We investigated predation success, latency, number of attempts during the process, and size distribution, in a total of 152 foraging trials of 38 pikeperch fingerlings (mean TL = 90.65 mm) at three different prey sizes (predator–prey length ratio (PPR): 0.11, 0.22, 0.33). Although predation on medium–large prey was more successful overall, it was achieved in more attempts and with greater latency. Small prey was captured with the fewest attempts. These results suggest that when calculating prey size in the foraging training of pikeperch, it is recommended to use a 0.11–0.22 PPR less than the optimal PPR described for wild individuals.
1. Introduction
In central and eastern European countries where the traditional pond culture is frequent, the combined way of low-cost juvenile pond production followed by RAS production is profitable [1]. Although RAS is safer for percid fish health due to the reduced risk of pathogen entry [2], the combined culture method has several benefits compared to the whole cycle RAS production at the present state of the culture technology. High production efficiency and low level of morphological deformities characterize this method, resulting in high quality and quantity of fingerlings [3]. The reduced feed intake and growth rate of the individuals in the size range of 400–1000 g [4] also confirms the increase in the economy of the grow-out phase by raising them on live fish food in pond culture.
Different species can be introduced into the natural environment with varying success, following hatchery rearing, and those that can be established well are generally not sensitive to the artificial environment [5]. The intensively reared pikeperch can be introduced into lake conditions [6]; however, its survival is significantly lower in the presence of other predatory species [7]. RAS, as a stimulus-poor rearing environment, changes the behavioral variability, plasticity, and cognitive capacity of intensively reared fish [8]. In the case of pikeperch, habituation to pelleted food could affect the behavior spectrum of the population. Individuals that start consuming food late or refuse it are more exploratory and less sensitive to stress; thus, less bold, and competitive individuals may be selected as a result of the intensive culture [9]. Ahlbeck and Holliland [10] pointed out that individuals raised in pond culture or RAS show different exploratory and foraging behaviors. Pond-raised juveniles (TL: 63–76 mm) begin to start foraging faster on live invertebrate prey and are more active in their antipredator response.
Under natural conditions, exogenous feeding starts 4–5 days after hatching at a length of 6–9 mm in Perca and Sander species. Thereafter, they feed on small planktonic organisms up to 20–35 mm and then switch to larger food with higher energy content at 30–35 mm. At this point, there are differences between species. For walleye (Sander vitreum) and pikeperch, a growth-dependent shift to piscivory is observed from 35 to 100 mm in size, depending on prey abundance and size, with a preferred prey size of one-fifth of their length [11]. In the case of perch (Perca fluviatilis) and Volga pikeperch (Sander volgensis), this can be observed later at larger sizes. In the case of pikeperch, the lack of large zooplankton or invertebrates can lead to poor growth and condition resulting in significant cannibalism due to the large size variance of the stock [12].
The culture environment presents many challenges to the fish, on the one hand providing a stable predictable environment with the absence of predation, a balanced food supply, and a physical environment, and on the other hand a significant source of stress through increased stocking densities and handling. The adult phenotype of an individual is significantly influenced by the environment in which its early development occurs. The environment during the development of the behavior significantly influences the later phenotype and cognitive abilities through brain development (reviewed by [8,13]). Environment (predation pressure) also influences boldness and, through energy expenditure linked to personality types, the growth of the individual [14]. In addition, boldness can be significantly influenced by the social environment of the individual [15]. Sensory use also varies significantly across environments, and although behavioral lateralization is involved in an antipredator mechanism, it also significantly impacts foraging behavior. Lateralized individuals can capture more prey than their peers [16], but lateralization is less observed in predation-free environments [17].
Environmental enrichment serves to reduce the selection effects of intensive rearing by providing a more complex environment during development. Its role is complex, as it not only improves fish welfare under intensive conditions but also allows individuals to perform better under natural conditions. Of the five categories of environmental enrichment, structural enrichment has been the main focus in recent years [18]. However, the other four strategies (sensory, occupational, social, and dietary enrichment) are also worthy of attention from a fish farming perspective. In the case of pikeperch, there are publications addressing animal welfare (structural enrichment [19], occupational [20], social [19,21], and dietary enrichment [22]), however, the consequence of dietary enrichment is not yet widely studied. The most important skills for captive-reared individuals during release into the wild are predator avoidance, foraging, and reproductive behavior. In the natural environment, for carnivorous fish, capturing live prey and learning tactics are of primary importance, which is facilitated by foraging training, mostly using direct exposure techniques [5]. Although there are publications available on the predation behavior of the pikeperch [23,24,25,26,27] information on the foraging of live prey by intensively farmed individuals is still limited.
Foraging training studies in other species report variable successes, yet mostly result in improvements in the foraging behavior of naïve fish. Studies generally aim to establish learning curves in order to determine the number of attempts sufficient to achieve an already acceptable post-release performance [5]. The training induces learning either through direct exposure to live prey or through social learning [13]. Increased foraging success has been found for Atlantic salmon (Salmo salar) [28,29], largemouth bass (Micropterus salmoides) [30], jade perch, (Scortum barcoo) [31], turbot (Scophthalmus maximus) [32], and European grayling (Thymallus thymallus) [33]. In esocids (muskellunge Esox masquinongy and tiger muskellunge (muskellunge × northern pike E. lucius)), pre-release feeding experience did not significantly alter post-release prey consumption or growth, although the survival of naïve (pellet-consuming) individuals was poorer [34]. In the case of the walleye, under laboratory conditions, the experienced group consumed more prey compared to the naïve group, but this difference could not be confirmed in pond experiments [35]. Changes in food preference have been observed in juvenile Atlantic salmon as a result of foraging training compared to the initial naïve state [36].
The pikeperch, similar to its North American relative the walleye, could show an improvement in the success rate of the behavior based on practice [27,35]. Exercise has probably a significant role in the kinematics of the behavior [37,38]. The size of the prey can affect the learning process, larger prey can quickly lose (energetic) value due to higher handling costs and possibly lower predation success [39]. In wild-caught pikeperch, the preferred PPR value was 0.23 on average, while a maximum PPR of 0.63 was measured for 98 mm-TL individuals [40]. Based on a laboratory prey selection study [24], active prey choice can be observed in the species with a preference of 0.25 PPR.
The aim of our study was to investigate whether and how intensively reared pikeperch show a prey size preference when foraging on live fish. In addition, we investigated how different prey sizes affect the prey capture process (number of attempts, predation latency) and whether the size of the prey ultimately captured is related to the prey size observed during attempts and predation success.
2. Materials and Methods
2.1. Ethical Approval
The protocol was approved by the Committee on the Ethics of Animal Experiments of the Hungarian University of Agriculture and Life Sciences Kaposvár Campus (permit number: 3/2016-MÁB).
2.2. Experimental Fish and Environment
The experiment was carried out at the Fish Laboratory of the Hungarian University of Agriculture and Life Sciences Kaposvár Campus (Hungary). Juvenile pikeperch (Sander lucioperca L.) (mean standard length 38.4 ± 0.6 mm, mean body weight 0.8 ± 0.1 g) were purchased from BO-FA Fish Farm (Attala, Hungary). After acclimation to pellets (in a 12-day weaning period), the fingerlings were reared in a recirculation system, under low stocking density (1.8 g/L) and consumed the pelleted food, Skretting Alterna 2P, until satiation. Cannibal individuals were sorted daily. The system with a total volume of 2600 L was an experimental aquarium system consisting of 30 tanks (60 L) and an external filter. A daily water change rate of about 10% was applied during the study. The water flow rate was set at 1.5 L/min and the temperature was maintained at 20.4 ± 0.3 °C. The lighting provided a luminance of 50 lx for a 12/12 h cycle. Water quality parameters were determined twice weekly using a Compact Photometer PF-12 Plus (Macherey-Nagel). The parameters measured were dissolved oxygen = 7.6 ± 0.4 mg/L, pH = 7.4 ± 0.1, ammonia (NH4+) = 0.09 ± 0.05 mg/ L and nitrite (NO2−) = 0.04 ± 0.03 mg/L.
The standard length and weight of the fish were measured, after which they were anesthetized with clove oil (10 drops in 10 L water). At the beginning of the experiment, the 38 fingerlings had an average standard length of 90.6 ± 4.8 mm and an average body weight of 11.1 ± 1.8 g. They were selected to have the average size of the stock and the smallest possible variance so as not to influence the PPR value during the experiment.
2.3. Behavior Testing
Live fish foraging behavior was studied in a 110 L (90 cm × 35 cm × 35 cm) aquarium. The walls of the aquarium were painted black except for the front, and it contained one artificial plant, and its bottom was covered with small black gravel. The three-prey fish of different sizes (immature Pethia conchonius, one prey of each size) was placed in a partitioned section before testing and the test started with the removal of the partition wall. The pikeperch was placed in the tank 12 h prior to the test to acclimate to the environment. Each pikeperch was subjected to four prey foraging tests with 12 h intervals between tests to ensure motivation (hunger). The test was terminated with a successful predation event (the pikeperch was allowed to eat only one prey), or after 60 min.
We used three prey size categories: 10 mm, 20 mm, and 30 mm standard body length in the small, medium, and large categories, respectively. The categories were formed according to the determination of the average mouth gap of the pikeperch based on the equation: G = 0.127 Lstandard + 0.891 [26]. The average mouth size of the pikeperch was calculated to be 12.4 mm from an average standard body length of 90.65 mm. After measuring 50 specimens of prey fish, the body length–height ratio was 0.42 ± 0.03, based on which the maximum prey length was determined to be 29.52 mm with a pikeperch mouth size of 12.4 mm. With a length of 10 and 20 mm, the height of the prey was 4.2 and 8.4 mm, respectively. The medium category corresponded to the preferred PPR given in the literature (0.22), while the small and large categories had PPR values of 0.11 and 0.33, respectively.
During the test, video recordings of the pikeperch behavior were made (Sony HDR-XR camera). From the recordings, the latency of successful predation, the size of the prey captured, the number of attempts made up to that point, and the size of the prey at each attempt were evaluated. A total of 152 recordings were analyzed (four trials for 38 pikeperch). The video recordings were analyzed manually by the same person for each fish, using PMB version 5.2 software.
2.4. Statistical Analysis
Factor analysis (extraction method: Principal component analysis, varimax rotation) was used to determine the types of foraging behavior. The PCA was performed on the number of attempts for each of the three size categories observed in the tests. The relationship between factor scores and the number of attempts was determined using regression analysis. Based on the factor scores of the first factor, the foraging tests were clustered using a Two-Step cluster analysis (number of iterations 15). The effect of repeated tests was investigated using a general linear model (GLM), where foraging clusters were included as a fixed variable and the individual identifier as a random variable. The model also included the interaction of the two variables. The effect of the foraging clusters (types) was further analyzed using a Cox proportional-hazard regression model performed on the latency of predation measured in the total of 152 and 72 of the successful life foraging tests. The success rate and the distribution of the number of attempts between different prey sizes and foraging clusters were compared by the Chi-square test. Bonferroni correction was applied to compare the number of attempts across the three prey sizes at a p = 0.016 significance level. Statistical analyses were performed using SPSS 17.0 software [41].
3. Results
3.1. Foraging Clusters and Their Influence on Patterns of Predatory Behavior
For a total of 152 tests with varying numbers of prey capture attempts within the small, medium, and large categories, factor analysis identified two factors covering 75.9% of the total variance. The first factor (which covered 38.9% of the variance) was predominantly determined by large prey (component coefficients of −0.182, 0.424, and 0.830 for small, medium, and large prey, respectively), while the second factor was determined by small prey (component coefficients of 0.884, 0.339, and −0.231 for small, medium, and large prey, respectively). The first factor and the number of attempts showed a correlation (r = 0.791, p < 0.001, Figure S1), and the 152 tests were classified into two clusters based on factor scores (Two-Step cluster centroids: 1. cluster: −0.543 ± 0.407; cluster 2: 1.252 ± 0.817). Out of the 152 foraging tests, 106 (69.7%) belonged to cluster 1, while 46 (30.3%) belonged to cluster 2, the latter predominantly foraging on medium–large prey animals. The number of attempts was higher in cluster 2 (Figure 1).
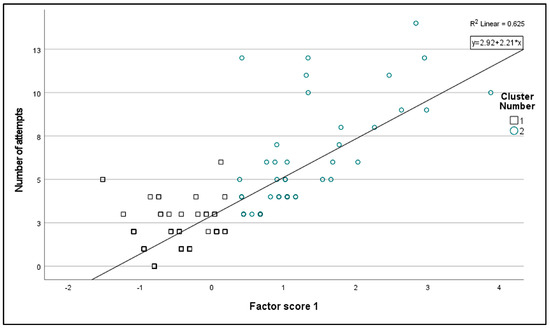
Figure 1.
Separation of tests by foraging behavior type, based on the number of trials in each prey size category. The factor score 1 was obtained using PCA based on the number of attempts observed in each of the three prey-size categories. The two clusters were separated using a Two-Step cluster analysis.
The GLM results show that the repeated tests had no significant effect (p = 0.803) on the latency of the predation, although the effect of foraging clusters (p < 0.001) and the interaction of repeated tests and clusters (p = 0.001) were significant (Table 1).

Table 1.
The effect of repeated tests and foraging clusters on foraging latency (general linear model).
The proportion of large prey fish in predation attempts affects not only the number of attempts, yet also the latency of predation. The effect of large prey is not significant for all 152 trials, which include ultimately unsuccessful trials, (Cox regression: Overall: Chi2 = 3.182, df = 1, p = 0.074; cluster count: Wald Chi2 = 3.135, df = 1, p = 0.077, ExpB = 0.653). However, for the trials that resulted in successful predation (n = 72), the predation attempt on the medium–large prey significantly increased the latency to predation (cluster 1: n = 43, mean = 354.8 sec, SD = 472.8; cluster 2: n = 29, mean = 911.2 sec, SD = 140.1), resulting in different temporal patterns of foraging behavior (Cox regression: Overall: Chi2 = 12.749, df = 1, p < 0.001; cluster number: Wald Chi2 = 12.047, df = 1, p < 0.001, ExpB = 2.463). The predation event occurs about 500 s earlier in the small–medium prey predation test compared to the medium–large prey predation test (Figure 2).
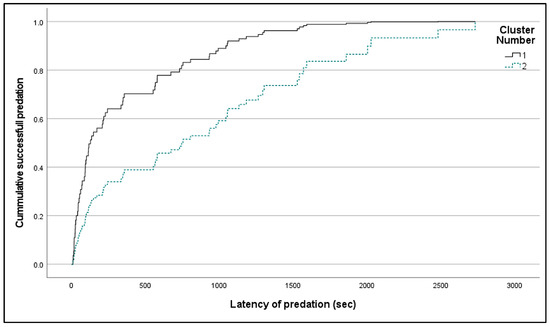
Figure 2.
Cumulative event curves showing the effect of foraging clusters on the probability that fish had captured the prey before a given time during an experimental trial.
3.2. The Effect of Foraging Clusters and Predation Success on the Number of Attempts for Prey of Different Sizes within the Tests
Of a total of 38 individuals, eight did not result in a successful capture in any of the tests. Test success rates differed significantly between the two foraging clusters (Chi2 = 6. 458, df = 1, p = 0.011; Table 2). In the tests belonging to cluster 2 (preferring medium–large fish), 63.0% of the tests (29/46) resulted in successful predation, while in tests classified in cluster 1, this rate was only 40.5% (43/106). Within the 72 successful trials, the distribution of the three prey size categories differed significantly within the two clusters (Chi2 = 11.849, df = 1, p < 0.001). The distribution indicated the preference of the clusters, although captures on medium prey were similar in both clusters.
Table 2.
The distribution of the number of successful predations on the prey of different sizes in the two clusters during the tests.
Table 3 shows the number of attempts in the tests, ranked by the number of times the pikeperch attempted the given prey size. If we consider the attempts as a function of the size of the prey finally caught, significant differences were found in the distribution of frequencies (between small and medium sizes: Chi2 = 44.168, df = 1, p < 0.001; small and large sizes: Chi2 = 34.323, df = 1, p < 0.001; medium and large sizes: Chi2 = 30.987, df = 1, p < 0.001). Moreover, the total number of attempts was higher when the larger prey was finally captured (n = 113), the difference was significant both with the small prey (n = 79) (Chi2 = 9.361, df = 1, p = 0.002) and the medium prey (n = 77) (Chi2 = 10.545, df = 1, p = 0.001). The numbers of attempts on the small and the medium prey were not significantly different (Chi2 = 0.036, df = 1, p = 0.849). When considered independently of the final captured prey size, the total number of attempts also differed for the three prey categories: 67 (43 + 7 + 17), 110 (19 + 56 + 35), and 92 (17 + 14 + 61) for small, medium, and large prey, respectively. However, the difference was only significant between the small and medium prey (Chi2 = 15.568, df = 1, p = 0.001), and marginal between the small and large prey (Chi2 = 5.579, df = 1, p = 0.018), after the Bonferroni correction. The total attempt number was similar between the medium and large prey (Chi2 = 2.568, df = 1, p = 0.109).
Table 3.
Distribution of the number of attempts to capture prey of different sizes during successful tests according to the size of the prey captured.
4. Discussion
Our study investigated the prey-foraging behavior of pikeperch juveniles when they received prey of different sizes. One of the most important results was that the intensively reared pikeperch were unsuccessful in 52% of the trials, despite attempts, during the one-hour test period. This explains that survival will be lower when these individuals are reared in ponds under non-ideal conditions (the presence of other predators as competitors [7]). As one-fifth of the individuals failed the test in all four cases, this highlights the importance of foraging training and confirms that adaptation to an intensive environment shows large individual differences among pikeperch and leads to selection in behavior [9]. The high failure rate can be explained by several factors. When rearing pikeperch in RAS, they must respond to a number of changes compared to their natural environment. The most significant of these is the consumption of pellets, however, there are also changes in the physical environment and the social environment. Individual differences in the success of foraging may be due to their personality. Although group modification effects are not present in our study (behavior was examined individually), our previous study has already shown that exploration is associated with the timing of the transition to pellet consumption [9]. It is likely that exploration levels associated with different personality types also influence the capture of live prey [27]. On the other hand, the complexity of the environment also influences the ability to learn. In a highly heterogeneous environment, fish are often forced to change behavior through learning, while a stable, stationary environment typical to RAS may lock in individual behaviors over time. This influences not only the learning to forage live prey, but also the need to ‘forget’ to search for pellets [42].
Predatory behavior could be classified into two major groups based on the prey size attempted during the tests: small–medium prey and medium–large prey preference behavior. Although the medium prey attempt was present in the process, it was not dominant in any of the tests. In the intensively reared pikeperch, small–medium prey attempts were more prevalent (two-thirds of the cases), indicating a preference for PPR values of 0.11 and 0.22, respectively. This is consistent with the preference of wild-caught pikeperch, where PPR was similar for different prey species: perch 0.11, pikeperch 0.23, ruffe 0.14, roach 0.23 [40]. Turesson et al. [24] examining the prey size preference of the species, found it to be 0.25 when examined in both the 0.25–0.45 and the 0.20–0.30 PPR ranges. Under natural conditions (in stomach contents), an average PPR of 0.28 was also obtained. Their study showed that pikeperch prefer smaller prey, and although approach rates did not differ, attack rates were lower for larger prey. Our study also confirms this for intensively reared naïve fish, although in our case the preference for smaller fish was observed at lower PPR values.
Although the three size categories were almost equally represented in the successful tests, the number of trials that preferred the medium–large category was only one-third of the total, so these trials were about 20% more successful. There are several possible explanations for this. The first is differences in prey behavior. It is possible that differences in size may also cause differences in predator avoidance behavior [43] or that swimming performance varies as a function of size [44], which could result in altered predation risk. The other possibility could be due to differences between pikeperch individuals. It is possible that the two different size preference categories were used by pikeperch individuals with different personalities, with larger prey sizes being preyed upon by more active, exploratory individuals [45]. In the case of the transition to pellet consumption, more exploratory individuals tended to consume live food during the transition, accepting pellets later or not at all [9]. Even more, a combination of the two factors could have shaped the outcome.
Foraging on larger prey resulted in more attempts and longer predation latency. Four mechanisms may underlie the consumption of costly prey [46]: the presence of direct benefits (e.g., limiting nutrients), individual speciation, optimal foraging, and compensatory growth. Direct benefits are unlikely in the present case, as individuals of different sizes of the same prey species were used. Optimal foraging theory suggests a PPR size of 0.26 is optimal for pikeperch [24], with larger sizes rewarding costly prey consumption only in the presence of different proportions of resources, which was not the case. It is more reasonable to hypothesize the remaining two mechanisms, either individual specialization (based on behavioral, and sensory differences) or, more likely, greater motivation (hunger) behind the attempt to prey on larger prey. These individuals were temporarily compensating for their possibly lower previous food intake by consuming larger prey. Higher motivation also seems to be confirmed by the fact that, despite the higher cost, predation success was also higher in these trials. Finally, an additional factor may have caused the preference for larger fish. Since cannibalism was not controlled during larval rearing in our study, it is possible that non-naive fish were included in the study, whose preference was already different from the others. Although in practice it is unlikely since cannibalism, at any stage of rearing, causes a significant size difference that persists later (resulting in significantly larger individuals), and we used in our study individuals that have the average size of the stock.
For different prey sizes, the number of attempts was found to be significantly lower for the small size. Out of a total of 67 attempts, 43 attempts were in a prey capture process that ultimately resulted in the capture of the small prey. Nevertheless, the proportion of small prey within successful prey captures did not differ. These results suggest that although the small prey had a suboptimal PPR, it was beneficial for the learning process of intensively reared naïve individuals. Learning not only results in faster predation but also provides energetic benefits by altering attack latency and prey acquisition efficiency [47]. Our study suggests that a slightly suboptimal PPR (compared to wild fish) during foraging training is recommended.
Finally, it should be mentioned that foraging training may be subject to bias in the practical implementation. The main problem may be the production of the right forage fish. When foraging training is carried out in a closed system, the production of the necessary forage fish entails additional costs, the return on which requires further investigation. If large quantities of pond-produced prey fish are available (e.g., invasive species: Carassius gibelio, Pseudorasbora parva), this could be a solution to the problem, but the introduction of prey fish from ponds into a closed system poses a high health risk. However, it may be appropriate to carry out foraging training in small ponds immediately prior to stocking, in which case other forms of environmental management could be implemented. However, this requires further investigation to develop the correct method.
5. Conclusions
Our results suggest the use of foraging training prior to pond rearing of intensively reared pikeperch. Prey acquisition tests showed a small–medium prey preference similar to wild fish. Although predation on medium–large prey was more successful overall, it was achieved in more attempts and with greater latency. Small prey was captured with the fewest attempts. These results suggest that when calculating prey size in the foraging training of pikeperch, it is recommended to use a PPR less than the 0.25 optimal PPR described for wild individuals.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app13042259/s1, Figure S1: Changes in the number of attempts as a function of PCA factor scores; Table S1: The raw data of the experiment.
Author Contributions
Conceptualization, T.M. and I.B.; methodology, T.M.; software, T.M.; validation, T.M. and I.B.; formal analysis, T.M.; investigation, T.M. and I.B.; resources, T.M.; writing—original draft preparation, T.M. and I.B.; writing—review and editing, T.M. and I.B.; visualization, T.M.; funding acquisition, T.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Hungarian Scientific Research Fund, OTKA K112070, and OTKA K143458.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board (or Ethics Committee) of the Hungarian University of Agriculture and Life Sciences Kaposvár Campus (permit number: 3/2016-MÁB).
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data of the experiment is available as Supplementary Materials (Table S1).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Policar, T.; Stejskal, V.; Kristan, J.; Podhorec, P.; Svinger, V.; Blaha, M. The effect of fish size and stocking density on the weaning success of pond-cultured pikeperch Sander lucioperca L. juveniles. Aquacult. Int. 2013, 21, 869–882. [Google Scholar] [CrossRef]
- Rodger, H.D.; Phelps, N.B.D. Percid fish health and disease. In Biology and Culture of Percid Fishes; Springer: Dordrecht, The Netherlands, 2015; pp. 799–813. [Google Scholar]
- Policar, T.; Blecha, M.; Křišťan, J.; Mráz, J.; Velíšek, J.; Stará, A.; Stejskal, V.; Malinovskyi, O.; Svačina, P.; Samarin, A.M. Comparison of production efficiency and quality of differently cultured pikeperch (Sander lucioperca L.) juveniles as a valuable product for ongrowing culture. Aquacult. Int. 2016, 24, 1607–1626. [Google Scholar] [CrossRef]
- Ende, S.S.; Larceva, E.; Bögner, M.; Lugert, V.; Slater, M.J.; Henjes, J. Low turbidity in recirculating aquaculture systems (RAS) reduces feeding behavior and increases stress-related physiological parameters in pikeperch (Sander lucioperca) during grow-out. Transl. Anim. Sci. 2021, 5, txab223. [Google Scholar] [CrossRef] [PubMed]
- Näslund, J. Reared to become wild-like: Addressing behavioral and cognitive deficits in cultured aquatic animals destined for stocking into natural environments—A critical review. Bull. Mar. Sci. 2021, 97, 489–538. [Google Scholar] [CrossRef]
- Blecha, M.; Kristan, J.; Policar, T. Adaptation of intensively reared pikeperch (Sander lucioperca) juveniles to pond culture and subsequent re-adaptation to a recirculation aquaculture system. Turk. J. Fish. Aquat. Sci. 2016, 16, 15–18. [Google Scholar] [CrossRef]
- Zakęś, Z.; Szczepkowski, M.; Szczepkowska, B.; Kowalska, A.; Kapusta, A.; Jarmołowicz, S.; Piotrowska, I.; Kozłowski, M.; Partyka, K.; Wunderlich, K.; et al. Effects of stocking earthen ponds with pikeperch (Sander lucioperca (L.)) fingerlings reared in recirculating aquaculture systems-effects of Fish size and the presence of predators. Bulg. J. Agric. Sci. 2015, 21 (Suppl. S1), 5–11. [Google Scholar]
- Salena, M.G.; Turko, A.J.; Singh, A.; Pathak, A.; Hughes, E.; Brown, C.; Balshine, S. Understanding fish cognition: A review and appraisal of current practices. Anim. Cogn. 2021, 24, 395–406. [Google Scholar] [CrossRef]
- Molnár, T.; Csuvár, A.; Benedek, I.; Molnár, M.; Kabai, P. Domestication affects exploratory behaviour of pikeperch (Sander lucioperca L.) during the transition to pelleted food. PLoS One 2018, 13, e0196118. [Google Scholar] [CrossRef]
- Ahlbeck, I.; Holliland, P.B. Rearing environment affects important life skills in pikeperch (Sander lucioperca). Boreal. Environ. Res. 2012, 17, 291–304. [Google Scholar]
- Feiner, Z.S.; Höök, T.O. Environmental biology of Percid fishes. In Biology and Culture of Percid Fishes; Springer: Dordrecht, The Netherlands, 2015; pp. 799–813. [Google Scholar]
- Specziár, A. Size-dependent prey selection in piscivorous pikeperch Sander lucioperca and Volga pikeperch Sander volgensis shaped by bimodal prey size distribution. J. Fish Biol. 2011, 79, 1895–1917. [Google Scholar] [CrossRef]
- Braithwaite, V.A.; Ahlbeck Bergendahl, I. The Effects of Early Life Experience on Behavioural Development in Captive Fish Species. In The Welfare of Fish; Kristiansen, T., Fernö, A., Pavlidis, M., van de Vis, H., Eds.; Animal Welfare Book Series; Springer: Cham, Switzerland, 2020; pp. 111–128. [Google Scholar] [CrossRef]
- Conrad, J.L.; Weinersmith, K.L.; Brodin, T.; Saltz, J.B.; Sih, A. Behavioural syndromes in fishes: A review with implications for ecology and fisheries management. J. Fish Biol. 2011, 78, 395–435. [Google Scholar] [CrossRef] [PubMed]
- Magnhagen, C. Behaviour of Percid fishes in the wild and its relevance for culture. In Biology and Culture of Percid Fishes; Springer: Dordrecht, The Netherlands, 2015; pp. 799–813. [Google Scholar]
- Berlinghieri, F.; Panizzon, P.; Penry-Williams, I.L.; Brown, C. Laterality and fish welfare—A review. Appl. Anim. Behav. Sci. 2021, 236, 105239. [Google Scholar] [CrossRef]
- Brown, C.; Gardner, C.; Braithwaite, V.A. Population variation in lateralized eye use in the poeciliid Brachyraphis episcopi. Proc. R. Soc. B Biol. Sci. 2004, 271 (Suppl. S6), S455–S457. [Google Scholar] [CrossRef] [PubMed]
- Arechavala-Lopez, P.; Cabrera-Álvarez, M.J.; Maia, C.M.; Saraiva, J.L. Environmental enrichment in fish aquaculture: A review of fundamental and practical aspects. Rev. Aquac. 2022, 14, 704–728. [Google Scholar] [CrossRef]
- Thomas, M.; Reynaud, J.G.; Ledoré, Y.; Pasquet, A.; Lecocq, T. Enrichment in a Fish Polyculture: Does it Affect Fish Behaviour and Development of Only One Species or Both? Appl. Sci. 2022, 12, 3674. [Google Scholar] [CrossRef]
- Thomas, M.; Lecocq, T.; Abregal, C.; Nahon, S.; Aubin, J.; Jaeger, C.; Wilfart, A.; Schaeffer, L.; Ledoré, Y.; Puillet, L.; et al. The effects of polyculture on behaviour and production of pikeperch in recirculation systems. Aquac. Rep. 2020, 17, 100333. [Google Scholar] [CrossRef]
- Lepič, P.; Buřič, M.; Hajíček, J.; Kozák, P. Adaptation to pelleted feed in pikeperch fingerlings: Learning from the trainer fish over gradual adaptation from natural food. Aquat. Living Resour. 2017, 30, 8. [Google Scholar] [CrossRef]
- Pěnka, T.; Malinovskyi, O.; Imentai, A.; Kolářová, J.; Kučera, V.; Policar, T. Evaluation of different feeding frequencies in RAS-based juvenile pikeperch (Sander lucioperca) aquaculture. Aquaculture 2023, 562, 738815. [Google Scholar] [CrossRef]
- Turesson, H.; Brönmark, C. Foraging behaviour and capture success in perch, pikeperch and pike and the effects of prey density. J. Fish Biol. 2004, 65, 363–375. [Google Scholar] [CrossRef]
- Turesson, H.; Persson, A.; Brönmark, C. Prey size selection in piscivorous pikeperch (Stizostedion lucioperca) includes active prey choice. Ecol. Freshw. Fish 2002, 11, 223–233. [Google Scholar] [CrossRef]
- Colchen, T.; Dias, A.; Gisbert, E.; Fontaine, P.; Pasquet, A. The onset of piscivory in a freshwater fish species: Analysis of behavioural and physiological traits. J. Fish Biol. 2020, 96, 1463–1474. [Google Scholar] [CrossRef] [PubMed]
- Malinovskyi, O.; Veselý, L.; Yanes-Roca, C.; Policar, T. The effect of water temperature, prey availability and presence of conspecifics on prey consumption of pikeperch (Sander lucioperca). Czech J. Anim. Sci. 2022, 67, 465–473. [Google Scholar] [CrossRef]
- Molnár, T.; Urbányi, B.; Benedek, I. Impact of exploration behavior, aptitude for pellet consumption, and the predation practice on the performance in consecutive live prey foraging tests in a piscivorous species. Anim. Cogn. 2023. accepted. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; Davidson, T.; Laland, K. Environmental enrichment and prior experience of live prey improve foraging behaviour in hatchery-reared Atlantic salmon. J. Fish Biol. 2003, 63, 187–196. [Google Scholar] [CrossRef]
- Brown, C.; Markula, A.; Laland, K. Social learning of prey location in hatchery-reared Atlantic salmon. J. Fish Biol. 2003, 63, 738–745. [Google Scholar] [CrossRef]
- Diana, M.J.; Diffin, B.J.; Einfalt, L.M.; Wahl, D.H. Effect of rearing experience on the survival, growth, and behavior of hatchery-reared largemouth bass. N. Am. J. Fish. Manag. 2018, 38, 794–802. [Google Scholar] [CrossRef]
- Reid, A.L.; Seebacher, F.; Ward, A.J.W. Learning to hunt: The role of experience in predator success. Behaviour 2010, 147, 223–233. [Google Scholar] [CrossRef]
- Ellis, T.E.; Hughes, R.N.; Howell, B.R. Artificial dietary regime may impair subsequent foraging behaviour of hatchery-reared turbot released into the natural environment. J. Fish Biol. 2002, 61, 252–264. [Google Scholar] [CrossRef]
- Czerniawski, R.; Domagała, J.; Krepski, T.; Pilecka-Rapacz, M. The effect of the live diet given to hatchery-reared fry of the European grayling (Thymallus thymallus) on their survival and growth in the wild. Turk. J. Fish. Aquat. Sci. 2015, 15, 633–638. [Google Scholar] [CrossRef]
- Szendrey, T.A.; Wahl, D.H. Effect of feeding experience on growth, vulnerability to predation, and survival of esocids. N. Am. J. Fish. Manag. 1995, 15, 610–620. [Google Scholar] [CrossRef]
- Wahl, D.H.; Einfalt, L.M.; Hooe, M.L. Effect of experience with piscivory on foraging behavior and growth of walleyes. Trans. Am. Fish. Soc. 1995, 124, 756–763. [Google Scholar] [CrossRef]
- Reiriz, L.; Nicieza, A.G.; Braña, F. Prey selection by experienced and naive juvenile Atlantic salmon. J. Fish Biol. 1998, 53, 100–114. [Google Scholar] [CrossRef]
- Wintzer, A.P.; Motta, P.J. A comparison of prey capture kinematics in hatchery and wild Micropterus salmoides floridanus: Effects of ontogeny and experience. J. Fish Biol. 2005, 67, 409–427. [Google Scholar] [CrossRef]
- Caldentey, P.; Brennan, N.P.; Heimann, T.; Gardiner, J.M. Prey capture kinematics of wild and hatchery juvenile common snook Centropomus undecimalis. Bull. Mar. Sci. 2021, 97, 539–558. [Google Scholar] [CrossRef]
- Gill, A.B. The dynamics of prey choice in fish: The importance of prey size and satiation. J. Fish Biol. 2003, 63, 105–116. [Google Scholar] [CrossRef]
- Dörner, H.; Hülsmann, S.; Hölker, F.; Skov, C.; Wagner, A. Size-dependent predator–prey relationships between pikeperch and their prey fish. Ecol. Freshw. Fish 2007, 16, 307–314. [Google Scholar] [CrossRef]
- SPSS Inc. Released SPSS Statistics for Windows; Version 17.0; SPSS Inc.: Chicago, IL, USA, 2008. [Google Scholar]
- Braithwaite, V.A. Cognitive ability in fish. In Fish Physiology: Behaviour and Physiology of Fish; Sloman, K.A., Balshine, S., Wilson, R.W., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2006; pp. 1–29. [Google Scholar]
- Krause, J.; Loader, S.P.; McDermott, J.; Ruxton, G.D. Refuge use by fish as a function of body length–related metabolic expenditure and predation risks. Proc. R. Soc. Lond B Biol. Sci. 1998, 265, 2373–2379. [Google Scholar] [CrossRef]
- Rubio-Gracia, F.; García-Berthou, E.; Guasch, H.; Zamora, L.; Vila-Gispert, A. Size-related effects and the influence of metabolic traits and morphology on swimming performance in fish. Curr. Zool. 2020, 66, 493–503. [Google Scholar] [CrossRef]
- Toscano, B.J.; Gownaris, N.J.; Heerhartz, S.M.; Monaco, C.J. Personality, foraging behavior and specialization: Integrating behavioral and food web ecology at the individual level. Oecologia 2016, 182, 55–69. [Google Scholar] [CrossRef]
- Langerhans, R.B.; Goins, T.R.; Stemp, K.M.; Riesch, R.; Araujo, M.S.; Layman, C.A. Consuming costly prey: Optimal foraging and the role of compensatory growth. Front. Ecol. Evol. 2021, 8, 603387. [Google Scholar] [CrossRef]
- Dill, L.M. Adaptive flexibility in the foraging behavior of fishes. Can. J. Fish. Aquat. Sci. 1983, 40, 398–408. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).